行政院國家科學委員會專題研究計畫 成果報告
家禽白血病 J 亞群病毒基因、抗原性及病原性之演變(3/3)
計畫類別: 個別型計畫 計畫編號: NSC92-2313-B-002-128- 執行期間: 92 年 08 月 01 日至 93 年 07 月 31 日 執行單位: 國立臺灣大學獸醫學系暨研究所 計畫主持人: 王金和 報告類型: 完整報告 處理方式: 本計畫可公開查詢中 華 民 國 93 年 8 月 23 日
行政院國家科學委員會補助專題研究計畫
█ 成 果 報 告 □期中進度報告家禽白血病 J 亞群病毒基因、抗原性及病原性之演變(3/3)
計畫類別:█ 個別型計畫 □ 整合型計畫
計畫編號:NSC 92-2313-B-002-128-
執行期間: 92 年 08 月 01 日至 93 年 07 月 31 日
計畫主持人:王金和
共同主持人:無
計畫參與人員: 陳素貞 陳慧真
成果報告類型(依經費核定清單規定繳交):□精簡報告 █完整報告
本成果報告包括以下應繳交之附件:無
□赴國外出差或研習心得報告一份
□赴大陸地區出差或研習心得報告一份
□出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份
處理方式:除產學合作研究計畫、提升產業技術及人才培育研究計畫、
列管計畫及下列情形者外,得立即公開查詢
□涉及專利或其他智慧財產權,█一年□二年後可公開查詢
執行單位:台灣大學獸醫學系
中 華 民 國 93 年 08 月 20
目錄
目錄---Ⅰ 表次---Ⅲ 中文摘要---Ⅳ 英文摘要---Ⅴ 1. 前言---1 2. 研究目的---1 3. 文獻探討---2 3.1 歷史背景---2 3.2 家禽白血病病毒的基因體結構與病毒蛋白---3 3.3 外源性家禽白血病 J 亞群病毒---3 3.4 病毒的傳播---5 3.5 家禽白血病病毒之實驗室診斷---5 4. 研究方法---7 4.1 採樣方式---7 4.2 樣本來源---7 4.3 抗體檢測---7 4.4 抗原檢測---7 4.5 細胞培養---8 4.6 病毒增殖與病毒核酸偵測---8 4.6.1 病毒來源---8 4.6.2 病毒增殖---8 4.6.3 病毒核酸偵測---8 4.6.3.1 病毒 RNA 之萃取---8 4.6.3.2 細胞之 DNA 萃取---9 4.6.3.3 反轉錄反應 (RT)---9 4.6.3.4 聚合酵素鏈反應 (PCR)---9 4.7 聚合酵素鏈反應產物之選殖 (cloning)---10 4.7.1 聚合酵素鏈反應產物之純化---10 4.7.2 聚合酵素鏈反應產物之選殖---10 4.8 建構表現載體---11 4.8.1 目標基因之增幅---11 4.8.2 構築表現載體---11 4.9 重組蛋白之表現與純化---11 4.9.1 勝任細胞的製備與表現載體的轉殖---11 4.9.2 重組蛋白之表現---11 4.9.3 重組蛋白之純化---12 4.10 重組蛋白之應用---124.10.1 gp85N 塗鍍 ELISA 平盤之製備與應用-Indirect ELISA---12 4.10.2 不同 gp85N 塗鍍量對抗體偵測結果之影響---13 4.10.3 不同血清種類與血清稀釋倍數對抗體偵測結果之影響---13 4.11 單株抗體之生產與應用---13 4.11.1 單株抗體之生產---13 4.11.2 單株抗體之確認---13
4.11.2.1 Immunodot blot assay---13
4.11.2.2 免疫螢光分析 (immunofluorescence assay, IFA)---14
4.11.2.3 單株抗體之應用-Sandwich ELISA---14 5. 結果---15 5.1 台灣有色雞群之血清學調查---15 5.2 2002 年至 2004 年肉雞、蛋種雞與有色雞種雞之家禽白血病 J 亞群病毒分離情形 ---15 5.3 建構表現載體---18 5.3.1 表現載體之確認---18 5.4 重組蛋白之表現與純化---18 5.4.1 重組蛋白之表現---18 5.4.2 重組蛋白之純化---18 5.5 重組蛋白之應用---18 5.5.1 應用 gp85N 塗鍍 ELISA 平盤偵測 ALV-J 抗體---18 5.5.1.1 不同 gp85N 塗鍍量對抗體偵測結果之影響---18 5.5.1.2 不同血清種類與血清稀釋倍數對抗體偵測結果之影響---18 5.5.1.3 使用 gp85N 塗鍍 ELISA 平盤測試 SPF 雞隻血清樣本---19 5.6 單株抗體抗原性之確認---19
5.6.1 Immunodot blot assay---19
5.6.2 西方墨點法---19
5.6.3 免疫螢光分析 (immunofluorescence assay, IFA)---19
5.7 單株抗體之力價測定---19
5.8 Sandwich ELISA 之開發---19
6. 討論---21
表次
Table 1: History of breeder flocks for serological investigation. ---32
Table 2: Serological investigation of meat-type hybrid native chicken in 2004. ---32
Table 3: PCR primers. ---33
Table 4: History of clinical cases during 2002-2004. ---33
Table 5: Investigation of GSA (p27) antigen and clinical signs in case 3067, 3068, 3069 and 3070. ---34
Table 6: Investigation of anti-ALV-J antibody in clinical case 3067, 3068, 3069 and 3070. ---35
Table 7: Investigation of GSA (p27) antigen in clinical case 3114. ---35
Table 8: Detection of expressed ALV proteins by sandwich ELISA. ---35
家禽白血病
J 亞群病毒基因、抗原性及病原性之演變(3/3)
計畫主持人:王金和
家禽白血病J 亞群 (subgroup J avian leukosis virus) 在控制上主要以撲滅計畫為主,因 此最重要的是區分出感染和未感染的雞隻,在檢測方面主要是偵測病毒抗原、核酸,或是 做抗體的檢測,不過目前所用的方式都有一些限制存在。因此本研究欲試圖發展更好的診 斷方法。本研究利用基因選殖技術將本實驗室ALV-J 分離株 2921 的部分 gp85 基因 (含 vr2、 hr1 及 hr2 三個高變異區) 選殖出來,進行重組蛋白 ”gp85N”的表現並針對選殖部分的胺基 酸片段發展單株抗體,再同時利用重組蛋白gp85N 發展間接 ELISA 來偵測 ALV-J 特異性 抗體及使用單株抗體發展一三明治 ELISA 來偵測 ALV-J 抗原。其中偵測 ALV-J 特異性抗 體間接 ELISA 使用上有非特異性反應。發展單株抗體方面一共獲得 2 株單株抗體 mAb14 及mAb22,進一步利用單株來偵測 ALV-J 抗原,結果 mAb14 及 mAb22 都可成功偵測到重 組蛋白gp85N,但是只有 mAb22 可以偵測到 ALV-J 病毒。另外,國內的有色雞族群為了增 加產能,多以台灣本地雞與進口肉種雞做雜交以改良後代,因而將ALV-J 引入台灣有色雞 群中,但詳細的感染狀況卻仍不明,因此針對有色雞群做血清學調查,共調查10 個種用雞 群與40 個肉用雞群,其中公母各半,種用雞群不論公母,全部為 ALV-J 抗體陽性;肉用雞 群方面,公雞群有15% (3/20) 為 ALV-J 抗體陽性,母雞群則有 10% (2/ 20) 為 ALV-J 抗體 陽性,結果顯示ALV-J 普遍存於台灣有色雞族群中。此外,在 2002 至 2004 年間我們仍陸 續自白肉種雞與蛋種雞病例見有腫瘤發生且分離出 ALV-J,顯示除了有色雞族群外,在白 肉種雞與蛋種雞族群間ALV-J 感染問題仍然存在。
關鍵詞:
家禽白血病J 亞群 重組蛋白 gp85 免疫酵素連結反應English summary
Control of subgroup J avian leukosis virus (ALV-J) is mainly through elimination, and the first challenge is to differentiate between the infected and noninfected chickens. Methods for direct detection of ALV-J include the detection of viral RNA and viral antigen, or indirect detection for antibodies. However all of the currently available diagnostic methods have some limitation. The specific aim of this study is to develop more useful diagnostic methods for detecting ALV-J. In this study, partial gp85 gene, which included three variable regions: vr3, hr1 and hr2, was cloned into pRSET B and expressed in BL21(DE3). The expressed protein gp85N was used to develop indirect ELISA for detecting ALV-J specific antibody but this indirect ELISA had nonspecific reaction. The same gene fragment was used to prepare monoclonal antibodies. Two monoclonal antibodies, mAb14 and mAb22, were obtained and used to detect ALV-J antigen. The results indicate that both mAb14 and mAb22 react with gp85N, but only the later could detect ALV-J virus. In order to increase economic benefits, the poultry industries hybrid broiler breeder and native chicken in Taiwan and ALV-J was introduced into Taiwan by this way. But the situation of ALV-J infection in hybrid native chickens was unclear. The serological investigation was done for hybrid native chickens. In this study, 10 flocks of breeder and 40 flocks of meat-type chicken were investigated. Breeder flocks was 100% (10/10) ALV-J antibody positive no matter the male or female, meat-type-chicken flocks was 15% (3/20) ALV-J antibody positive in male and 10% (2/ 20) ALV-J antibody positive in female. The results of serological investigation indicate ALV-J is widespread among hybrid native chickens. Furthermore, tumors induced by ALV-J were seen in broiler breeder and layer breeder, and viruses were isolated from these clinical cases during 2002-2004 reveal that ALV-J still exists in broiler breeder and layer breeder flocks.
1. 前言
家禽白血病是由家禽白血病病毒 (avian leukosis virus, ALV) 所引起的禽鳥腫瘤性疾 病。對家禽產業而言,家禽白血病所造成的產能下降、免疫抑制等問題往往導致很嚴重的
經濟損失,不過在種雞業者施行家禽白血病病毒撲滅計畫後,家禽白血病A 亞群及 B 亞群
病毒已獲得良好的控制 (Payne et al., 1997)。不過至 1988 年,一個新的家禽白血病病毒亞 群的出現再次衝擊家禽產業 (Payne et al., 1991)。
家禽白血病J 亞群 (subgroup J avian leukosis virus, ALV-J) 在分類上屬於外源性家禽白 血病病毒,可以藉由垂直或是水平方式來傳播,經由母雞垂直傳播的小雞往往會導致免疫 耐性 (immuno-tolerance),而持續排毒,感染其他雞隻,除了導致骨髓球性白血病外 (myeloid leukosis, ML) (Payne et al., 1998),還會造成肉雞生長遲緩 ,以及影響雞隻的疫苗免疫 (Stedman et al., 1999; Spackman et al., 2003)。在所有雞隻品系中,以肉雞對 ALV-J 的感受性 最高,且雞隻間水平傳播速度最快 (Fadly et al., 1999),而使得 ALV-J 的控制較其他家禽白 血病病毒困難。另外 Payne 研究發現來亨雞品系 (Leghorn strains) 對 ALV-J 較有抗性 (Payne, 1992)。家禽白血病的控制主要是從撲滅計畫、飼養管理,以及以人為育種方式培養 出對家禽白血病病毒有抗性的雞隻著手 (Payne, 1998)。
在台灣,1995 年時 Wang 等人證實台灣肉雞場發生致死率很高的 myelocytomatosis (Wang et al., 1993),至 1997 年才進一步證實台灣有 ALV-J 的存在 (Wang et al., 2002);2002 年,徐針對台灣土雞進行調查,證實台灣土雞有ALV-J 的感染 (徐, 2002),但是詳細的感 染狀況仍然不明,因此本研究希望能針對台灣有色雞族群 (hybrid native chicken) 進行血清 學調查,以了解 ALV-J 在台灣有色雞族群內的分佈情形。
由於此病的控制仍是以撲滅計畫為主,因此找出感染雞隻並加以撲滅是相當重要的, 在病毒檢測方面主要是偵測病毒抗原、核酸,或是做抗體的檢測。在核酸檢測方面主要是 利用PCR 及 RT-PCR 的方式,但由於 ALV-J 毒株變異相當大 (Silva et al., 2000),因此發生 某些毒株無法被偵測出的問題 (Smith et al., 1998),也有人利用 nested PCR 來區分內外源性 病毒,不過仍有高達30%的偽陽性 (García et al., 2003)。抗原、抗體的檢測主要是利用商用 的ELISA 診斷套組進行,檢測抗體的 ELISA 套組在使用上可能發生偽陽性的情形 (Hunt et al., 2000);而檢測抗原的 ELISA 套組主要是針對 ALV 之 group specific antigen (p27),但是 內源性ALV 也會產生 group specific antigen (p27),因此檢測抗原的 ELISA 套組無法區分內 外源性的病毒 (Payne et al., 1993),雖然可配合病毒分離來使用檢測抗原的 ELISA 套組,但 是病毒分離工作費時又費力。
2. 研究目的
本研究希望利用 E. coli 表現之重組蛋白 gp85N 為抗原發展特異性更高的間接酵素連結 免疫吸附分析 (indirect ELISA) 來偵測 ALV-J 特異性抗體。同時生產針對 ALV-J SU (gp85) protein 包含 vr2、hr1、hr2 三個高變異區之部分片段生產單株抗體,並利用此單株抗體發 展間接三明治酵素連結免疫吸附分析 (indirect sandwich ELISA) 來偵測 ALV-J SU (gp85) protein,以利疾病診斷之用。
3. 文獻探討 3.1 歷史背景
家禽白血病 (avian leukosis) 追溯到最早的病例是於 1868 年 Roloff 以淋巴肉瘤 (lymphosarcomata )所發表出來 (Roloff, 1868),至 1908 年 Ellermann 和 Bang 於哥本哈根發 現紅血球性白血病 (erythroid leukosis) (Ellermann et al., 1921),至 1910 年 Rous 於紐約證實 家禽發生的白血病與肉瘤是由病毒所引起 (Rous, 1910),而後的數十年間家禽白血病造成 全球的家禽產業嚴重經濟損失。
家禽白血病病毒 (avian leukosis virus, ALV) 可感染許多不同品系的雞隻,甚至是其他 禽鳥 (Payne, 1987)。除了侵襲造血細胞,包括紅血球、淋巴球、骨髓球等,而導致白血病 的發生外,也會在腎臟、卵巢、睪丸、肝臟、胰臟、神經系統等等實質臟器造成腫瘤 (Beard, 1980; Purchase, 1987; Payne et al., 1997)。除了導致腫瘤的發生外,ALV 還會造成雞隻產蛋 率下降 (Gavora et al., 1980)。
ALV 可根據其宿主範圍的不同、同一亞群病毒間互相干擾的現象以及病毒本身抗原性 的不同而分為A-J 十個不同的亞群 (Payne et al., 1997),而其中以 A 亞群和 B 亞群為野外最 常見的外源性 (exogenous) 家禽白血病病毒 (Calnek, 1968);J 亞群最早是在 1988 年從肉雞 品系中分離出來,(Payne et al., 1991),其中一株 HPRS-103 被進ㄧ步研究且證實其病毒抗 原性與宿主範圍皆與已知的家禽白血病病毒亞群有所差異,因此將之歸類為新的亞群—J 亞群 (Payne et al., 1991)。 在台灣,最早是在1995 年 Wang 等人證實雞隻發生骨髓細胞瘤(myelocytomatosis) 且 造成很高的死亡率,不過當時並未分離出病毒 (Wang et al., 1995)。一直到 1997 年台灣一家 原種雞場因發生家禽白血病J 亞群 (subgroup J avian leukosis virus, ALV-J) 的感染而有嚴重 的經濟損失 (Wang et al., 2002),至此家禽白血病逐漸受到重視。至 2002 年徐亦證實台灣土 雞有ALV-J 的感染 (徐, 2002),不過台灣土雞感染的詳細情況仍然不明朗。 其實台灣現有的肉用雞大約可分為白肉雞、仿土雞與土雞 3 種。白肉雞主要是由歐 美國際性大育種公司所育成的,經由原源種雞場、種雞場而肉雞場等生產系列所生產出來 最有效率的雞種。仿土雞與土雞則是國內種雞場自行交配出來的雞種,因都具有非白色的 羽毛所以統稱『有色雞』以別於『白肉雞』。目前一般所稱謂的土雞並不是一個品種,而 只是養雞界、雞販與消費者對本省一類雞隻的稱呼。牠通常是具有大而直立的單冠、金黃 至紅色或其他花色(一般以金黃為多)的羽毛、鉛色的腳脛,而最重要的是要有性成熟的 外表才可上市。而仿土雞者通常體型較土雞為大,羽色原來多數為黑色,目前則多具紅羽 只是色澤比土雞為深;腳脛顏色則無一定標準,黑色居多,但也夾雜有黃色、白色或綠色 腳脛。仿土雞在 12∼13 週齡以前未達性成熟就出售,雞冠也較土雞為小 (李)。 如果按照遺傳形質與飼養方式,台灣目前的土雞應可分為 3 大類。第一類土雞為遺 傳與飼養環境方面都符合本地雞要件,數量極少;第二類是飼養環境符合者,而第三類則 是外表類似本地雞之一般商業生產的土雞 (李)。 第一類土雞:在全省各地原來都有,可是現在已接近絕跡,除非有心人士保存血統, 否則均已混雜外來雞血統。此類雞隻在各地均有所不同,因小群飼養,近親程度嚴重,因 此生長慢,較晚成熟,著肉不多,且產蛋少,母雞藾抱性強,生產效率極差 (李)。 第二類土雞:多飼於山坡地果林間,平時雞隻有廣大的運動空間,所以肉質較為堅實 富彈性。在鄉村農家副業飼養的雞隻多屬此類,其雞隻或購自飼料店、雛雞販,或自留母 雞孵蛋繁殖或種蛋託孵化場代孵,因此雞種在遺傳方面頗為混雜 (李)。 第三類土雞:則是目前飼養數量最多的商用土雞。這類土雞的飼養環境類似白肉雞, 均為大群同齡雞隻飼養於平飼雞舍,並給予完全配合飼料,育雛時給予保溫,也有防疫計 劃,幾乎與白肉雞相同,只是在雞舍旁給予運動場,在 5-6 週齡以後讓牠們有較大的活動
空間。由於空間較大,且有較乾燥的砂浴場所,雞隻外表看來都較活潑且羽毛光潔。一般 在 8 週齡時公母分開飼養,而在 13-15 週齡時出售。此類雞隻在遺傳上也不是本地雞, 也沒有固定的配種方式。各個雞場有不同的配種方式,但外表都是非白羽,主要以紅黃色 羽居多,具有大紅直立的單冠與鉛色的腳脛。因為這些性狀都是簡單幾個主基因控制,只 要引進基因雜交就很容易可將之純化。一般雞種來源有幾類,有自國外引進有色專用種肉 雞(單冠但為黃色腳脛),而以具有鉛色腳脛的公雞(基因來源可能是本地雞)與之交配, 再加以純化。此類土雞由於具有專用種肉雞的血統,因此生長較快,著肉較多,但種雞產 蛋性能較差,且體型較粗大。這些土雞的生產方式並不像白色肉雞或蛋雞以系統雜交生產, 而沒有利用雜交優勢以保護自己的雞種。較有規模的種雞場是由自己的肉雞場挑選較好的 雞隻作為種雞,而很多種雞場甚至由其他肉雞場挑選雞隻來作為種雞。由於產肉性能好的 雞隻通常產蛋性能較差,而且以肉雞方式育成的種雞,產蛋性能也較差,因此種土雞的產 蛋性能都很差。而種公雞則四處搜尋,由於較佳者不易獲得,因此都以人工授精進行繁殖 (李)。 3.2 家禽白血病病毒的基因體結構與病毒蛋白
家禽白血病病毒 (avian leukosis virus, ALV) 內部核蛋白中心含有正向雙套的 RNA 基 因體和三種病毒酵素: 反轉錄酶 (reverse transcriptase, RT)、嵌入酶 (integrase, IN) 及蛋白酶 (protease),它們的功能分別是將病毒 RNA 反轉錄成 DNA、將病毒基因以原病毒 (provirus) 的形式嵌入宿主基因體中,以及切割蛋白質以形成病毒蛋白。ALV RNA 基因體結構是 5’ R-U5-gag-pol-env-U3-R 3’,核酸序列上由 5 端到 3 端所 encode 的基因為 gag、pro、pol、 及 env,env 基因後面有一段 non-coding 的區域,此外核酸的兩端都有 LTR (long terminal repeate),為病毒基因表現所必須。gag 所轉譯的蛋白質為與病毒亞群性專一性有關的抗原 (group-specific antigen, GSA)包括 p27、p19、p12,其中 p27 為形成病毒蛋白鞘 (capsid) 的 蛋白質 (CA);pro 所轉譯的蛋白質為蛋白酵素(protease, PR);pol 所轉譯的蛋白質為反轉 錄酶(reverse transcriptase, RT),以及嵌入酶(intergrase, IN),env 所轉譯的蛋白質為 SU (gp85) 與TM (gp37)。SU 位在 env 所轉譯蛋白質的胺基酸端(amino-terminal),位於病毒封套外, 與病毒進入宿主細胞有關。env 轉譯的蛋白質具有一 signal peptide,可被宿主細胞辨識及切 割而形成完整的 env 基因產物,即 SU 及 TM。TM (Transmembrane Envelope)為 env 基因所 轉譯的蛋白質經蛋白酶(protease) 切割後的產物之一,在病毒進入宿主細胞時與細胞膜融合 有關 (Payne et al., 1997; Venugopal, 1999)。
病毒的基因體大小為7.2kb。除了以上的基本基因外,在非缺陷型的雞肉瘤病毒(Rous sarcoma virus, RSV)還另外存有一段致癌基因 (v-onc)-- src 基因,使其基因體組成為
gag/pro-pol-env-src(Enrietto et al., 1987)。RSV 因本身具有致癌基因,可使宿主快速的癌化,
屬於快速致癌型病毒(acute transforming virus)(Wang et al., 1988)。除了 src 外,快速致癌 型的病毒還有帶有 erbA、 erbB(AEV)、myb(AMV)等。相對於快速致癌型病毒,有的 病毒本身不帶有致癌基因,其致癌需要靠宿主本身帶有的致癌基因,因此往往需要較長的 時間才能導致腫瘤的發生,如LL 等,因此稱為慢速致癌型病毒(slow transformation virus) (Weiss et al., 1985)。
3.3 外源性家禽白血病 J 亞群病毒
家禽白血病J 亞群病毒 (subgroup J avian leukosis virus, ALV-J) 最早是在 1988 年 Payne 等人自肉種雞分離出來的,當時一共分離出5 株病毒,分別命名為 HPRS-100、HPRS-101、 HPRS-102、HPRS-103 及 HPRS-104 (Payne et al., 1991)。當時先利用病毒間的干擾作用
(interference assay) 來鑑別這些新病毒株是屬於哪個家禽白血病病毒亞群,結果發現這些新 分離株與先前已知的亞群都不同,此外也同時或單獨使用已知的家禽白血病病毒亞群的抗 血清同樣無法中和掉這些新病毒。另外針對其中一個毒株HPRS-103 進行宿主範圍的研究, 發現HPRS-103 除了可以感染雞、jungle fowl 和火雞胚胎纖維母細胞外,還可以感染多種鳥 類來源的細胞株 (Payne et al., 1992a)。將 HPRS-103 與其他家禽白血病亞群病毒 (A-I)的宿 主範圍作比較後,決定以HPRS-103 為原型病毒,定義為家禽白血病 J 亞群 (Payne et al., 1991)。
HPRS-103 具 有 慢 速 致 癌 型 病 毒 (slow transforming virus) 的 基 因 體 結 構 , 即 LTR-leader-gag/pol-env-LTR (Bai et al., 1995)。HPRS-103 的 LTR (long terminal repeat) 由 U3-R-U5 組成,與其他亞群的相似度高達 90%以上。HPRS-103 的 U3 相對於其他亞群而言, 其缺少enhancer protein (EFⅡ) 的結合位 (binding sites)。此外它的 leader 序列相較於其他 亞群 gag 基因 ATG 密碼子 (codon) 的上游還多了二個獨特的 ATG 密碼子 (codon),不過它 們的功能還不清楚。HPRS-103 的 gag 基因和 pol 基因與其他亞群有 96-97%的相似度,不 過 pol 基因有一個 stop codon,因此 pol 基因產物較其他亞群短少 22 個胺基酸,不過這並不 影響病毒的複製 (Katz et al., 1988)。
HPRS-103 的 env 基因序列與其他 ALV 亞群的差異較大 (Bai et al., 1995b)。SU (gp85) surface domain 與 A-E 亞群相比只有 40%的相似度,而 A-E 亞群間的 SU (gp85) surface domain 卻有 77-87%的相似度;而 TM (gp37) transmemberane domain 與 A-E 亞群相比有 65% 的相似度,而A-E 亞群間的 TM (gp37) transmemberane domain 的相似度高達 92-95%,此外 HPRS-103 的 TM domain 還有一個 219 bp 的 insertion (Bai et al., 1995a) 形成一個額外的區 域稱為redundant TM (rTM),此部份序列與其他亞群有 97%的相似度。ALV-J 的 env 基因與 其他亞群的 env 基因有極高度的差異,在基因庫搜尋的結果顯示 HPRS-103 的 env 基因序列 最接近內源性病毒EAV 中的 E51 的 env 基因序列 (Boyce-Jacino et al., 1992),它們的相似 度有75%以上,因此推測這些內源性病毒可能是 ALV-J env 基因的來源。不過另一方面 E51 缺失多且與HPRS-103 env 基因序列相似度不算高,因此認為 HPRS-103 env 基因可能是由 其他內源性病毒而來。為了證明這個理論,後來有人在雞隻的染色體上發現一個新的內源 性病毒EAV-HP,其與 HPRS-103 env 基因序列有 97%的相似度 (Bai et al., 1995b),這個新 的內源性病毒也稱為ev/J (Benson et al., 1998),在所有雞隻品系及古老的 jungle fowl 的染色 體上都有數個重複,但不存於其他鳥類染色體內 (Smith et al., 1999)。綜合目前已知的證據 顯示HPRS-103 是由外源性病毒與 EAV-HP 的 env 基因序列所重組出來的一個新的家禽白血 病亞群。HPRS-103 基因體上還有一個 150 bp 的片段,稱為 E element (也稱作 F2 或 XSR), E element 位於 3’non-coding 的區域,DR1 的下游 (Bai et al., 1995b),這個 element 先前只有 在Rous sarcoma virus 的 src 基因上游或下游看見 (Bizub et al., 1984),雖然這個 element 的 功能不明,不過它有轉錄因子 C/ EBP 的結合位 (Ryden et al., 1993),可能具有 enhancer 的 功能 (Laimins et al., 1984)。
反轉錄病毒有一個很顯著的特性,就是它們基因體不穩定且變異大,這是因為病毒聚 合酶的序列錯誤率和重組率高 (Nichol, 1996)。ALV-J 也不例外,不同分離株的抗血清不一 定能中和掉彼此 (Venugopal et al., 1998),經分析 12 個 ALV-J 分離株 env 基因序列後發現, ALV-J 的 env 基因序列上有數個高變異區,尤其是 hr1、hr2 和 vr3,而其中又以 hr2 最為重 要 (Venugopal et al., 1998)。推測這些序列的變化可能是由於病毒受到宿主免疫反應等的篩 選的壓力影響而改變其序列,進而使得抗原性有所變化,病毒不至於被宿主辨識而淘汰
(Venugopal et al., 1998)。
3.4 病毒的傳播
所 有 的 外 源 性 家 禽 白 血 病 病 毒 都 可 以 經 由 垂 直 (vertical transmission) 和 水 平 (horizontal transmission) 方式來傳播 (Rubin et al., 1962; Payne et al., 1997),而內源性病毒則 隨宿主染色體傳給子代 (genetic transmission)。ALV 主要是以垂直感染的方式傳播,也就是 受病毒感染的母雞將病毒經輸卵管至蛋白,在經蛋白感染將疾病傳染給胚胎 (Payne et al., 1982),因此雞隻在孵化前便受到病毒的感染 (Dougherty, 1967),這樣的小雞會產生免疫耐 性 (immunologically tolerant),而無法將病毒自體內清除而持續排毒 (viremia, V+; antibody, A-; shedding, S+),這種情況下雞隻很可能會產生腫瘤,這些小雞的胎糞含有大量的病毒, 是孵化室內病毒水平傳播的主要來源。
水平傳播主要是雞隻傷口、口、鼻及結膜直接接觸含病毒的糞便和唾液而被感染 (Spencer et al., 1977; Burmester, 1956),這也是 ALV-J 撲滅計畫中的一大挑戰,因為病毒水 平傳播的速度相當快 (Fadly et al., 1999; Witter et al., 2000)。孵化後被水平傳染的小雞會因 感染的ALV 不同而有不同的結果,感染 ALV-J 以外的亞群病毒後,雞隻通常會產生免疫力 而不會排毒 (V- A+ S-)。蛋雞品系與肉雞品系的雞隻對 ALV-J 水平感染後的反應不同,蛋 雞品系的brown leghorn 於孵化後注射病毒或是接觸感染雞隻會產生免疫反應而不排毒 (V- A+ S-);以相同方式處理肉雞,則不是像垂直感染的雞隻ㄧ樣產生免疫耐性 (V+ A- S+), 就是發生暫時性的病毒血症後產生免疫力,有部份產生免疫力的雞隻仍會排毒 (V- A+ S+) (Russell et al., 1997)。由於 ALV-J 傳播情形較其他亞群複雜,因此在疾病的控制上也較為困 難。
雞隻與病毒接觸的年齡與是否發病有關 (Coffin, 1996),接觸病毒的時間越早雞隻越容 易發病,如在孵化後數週才接觸病毒則雞隻較不易發病 (Fung et al., 1983)。由於病毒對熱
不 穩 定 , 因 此 ALV 的水平傳播主要是靠直接接觸傳染,間接接觸的傳染較少發生
(Dougherty, 1967; Pham et al., 1999)。
3.5 家禽白血病病毒之實驗室診斷
家禽白血病最主要的特徵是會導致腫瘤,在疾病的診斷上可以根據臨床症狀、腫瘤病 變來做診斷外,尚可用血清學方法檢測病毒的抗原或是抗體來作初步的診斷 (Fadly et al., 1998; Fadly, 2000),而後可利用雞胚胎或雞纖維母細胞(chicken embryo fibroblast)進ㄧ步 做病毒分離。
在病毒分離方面,ALV 可由血漿、血清、腫瘤等病材來進行分離(Fadly et al., 1998), 其中血漿最常被用來進行分離病毒(Fadly et al., 1998)。將家禽肉瘤病毒(Rous sarcoma virus)接種於 11 日齡的雞胚胎或雞纖維母細胞(chicken embryo fibroblast)時,會在雞的 尿囊絨毛膜(chorioallantoic membrane, CAM)造成 pork(Dougherty et al.,1960),或在雞纖 維母細胞造成細胞病變 (CPE)(Temin et al., 1958),不過大部分的 ALV 並不會造成 CPE, 因此在診斷時還需要配合 (1) Resistance inducing factor(RIF)(Rubin, 1960);(2) Phenotypic mixing(PM)(Okazaki et al., 1975);(3) Complement fixation test for avian leucosis(COFAL)、 Complement fixation(CF)(Sarma et al., 1964)。(4) Nonproducer cell activation(NP)(Rispens et al., 1970)等方法作病毒的檢測,不過這些用來檢測外源性病毒的方法需要靠先做病毒分 離,較為費時費力,故已很少使用。在進行病毒分離時需要使用具有感受性的細胞,目前
已發展出對各個亞群有抗性的細胞株。(1) C/0:對所有亞群均有感受性(Crittenden et al 1987);(2) C/E:對 E 亞群有抗性(Crittenden et al 1987); (3) C/AE:對 A、E 亞群有抗性 (Crittenden et al 1992);(4) C/J:對 J 亞群有抗性(Hunt et al., 1999)。
到了1980 年代,利用酵素反應結合抗體 (Wisdom, 1976) 而使得應用免疫分析來診斷 特殊蛋白或病原的方式得以迅速的發展。酵素連結免疫分析 (enzyme-linked immunosorbent assay, ELISA) 主要有三種系統 (1) 直接酵素連結免疫分析 (direct ELISA):為最簡單的 ELISA 設計,抗原被吸附在 ELISA 平盤底部後,洗去未吸附的抗原,接著直接加入酵素標 定的抗體來偵測抗原,經一段時間的作用後再次洗去多餘的抗體,最後加入呈色系統進行 呈色;(2) 間接酵素連結免疫分析 (indirect ELISA):抗原被吸附在 ELISA 平盤底部後,先 加入一特定可與抗原作用的抗體後,再加入抗此特定抗體且經酵素標定之抗血清,最後再 以呈色系統進行呈色;(3) 三明治酵素連結免疫分析 (sandwich ELISA):可在細分成兩類 a. 直接三明治酵素連結免疫分析 (direct sandwich ELISA),將抗體吸附在 ELISA 平盤底部來 捕捉抗原,再以另一個酵素標定抗體來偵測抗原,其中捕捉抗體 (capture antibody) 與偵測 抗體 (detecting antibody) 可以是相同的血清或是來自同種類不同隻動物的血清,也可以是 不同種類動物的血清,而被偵測的抗原至少需要有兩個以上的抗原決定位;b. 間接三明治 酵素連結免疫分析 (indirect sandwich ELISA),抗原與塗鍍在 ELISA 平盤底部的抗體 (coating antibody) 結合後,再以另一動物品種之抗體 (detecting antibody) 來偵測抗原,接 著再加入抗偵測抗體之酵素標定抗體,因此塗鍍抗體與偵測抗體須為來自不同品種的動物 血清,而使酵素標定抗體不會與塗鍍抗體作用 (Crowther, 2001)。
此後陸續有人使用 ELISA 來偵測 ALV 的抗原或抗體 (Clark et al., 1981; Smith et al., 1979; Smith et al., 1985; Tsukamoto et al., 1991; Venugopal et al., 1997; Youshio, 1986)。後來發 展出針對病毒GSA (p27) 的 ELISA(Fadly et al., 1998),可以用來檢測血液樣本、蛋白及陰 道拭子等,目前已有商業化的產物(Fadly, 2000)。不過這類偵測病毒 GSA (p27) ELISA 最 大缺點為無法區分內源及外源性的病毒,因此是否能用於檢測外源性病毒仍被懷疑(Fadly et al., 1981; Payne et al., 1993)。除此之外,現在也有發展出偵測抗 ALV-J gp85 特異性抗體 的 ELISA 套組(Venugopal et al., 1997),不過實際使用上仍有偽陽性發生 (Hunt et al., 2000)。因此若能開發偵測 ALV-J 抗體特異性更高的 ELISA 套組將更有助於疾病的診斷之 用。
在偵測病毒核酸方面,有針對各個亞群所發展出來的PCR 及 RT-PCR(Smith et al., 1998; Pham et al., 1999a)。而這些PCR 或 RT-PCR 其引子的設計大部分都是位於病毒的 env 或 LTR 片段,目的在為與內源性的病毒或其它亞群有所區別(Pham et al., 1999b)。此外,目前也 有人利用 nested PCR 來區分內外源性病毒,不過仍有高達 30%的偽陽性 (García et al., 2003)。
家禽白血病是很常見的病毒,因此對於腫瘤引起的疾病作病毒的分離與鑑定的意義有 限,不過就整個撲滅計劃而言這些檢測仍是必需(Fadly, 2000)。
4. 研究方法 4.1 採樣方式 本實驗依據統計學方式先做前置試驗,預估台灣土雞 ALV-J 特異性抗體陽性率約為 20%,再依據此預估值計算得知,每個雞群至少須採 14 個以上的樣本才具代表性,若其中 一個以上樣本測試為抗體陽性,則視此雞群為ALV-J 抗體陽性雞群。 4.2 樣本來源 實驗樣本來源有兩種,其中種雞部分的樣本為全台各地種雞場採得,並將血液以EDTA 抗凝後送至實驗室,再於實驗室中收集個樣本之血漿測試其中ALV-J 特異性抗體並保存於 -20℃冰箱中。而肉雞部分的樣本則是至中部與南部兩處的土雞屠宰場收集,血液樣本同樣 以EDTA 做抗凝後,再回實驗室收集血漿做 ALV-J 特異性抗體的測試,測試後同樣將血漿 保存於-20℃冰箱中。各樣本雞群之基本資料詳見於 Table 1. 及 Table 2.。 4.3 抗體檢測
本研究以商用酵素連結免疫吸附分析套組 (Enzyme-linked immunosorbent assay kit, ELISA kit, IDEXX Laboratories, Inc., Westbook, Maine)來檢測血漿樣本中的抗 ALV-J gp85 的 抗體 (anti-ALV-J gp85 antibody)。首先取出 ELISA 試劑套組於室溫下回溫,再將血漿樣本
以稀釋液做 500 倍稀釋,而後紀錄各樣本位置。以二重複方式將陽性與陰性對照組依序加
入A1 到 A4 孔中,再依紀錄好的位置加入 100µL 稀釋好的樣本,置於室溫下作用 30 分鐘。 30 分鐘後倒去盤中內容物後,每孔以 300µL 的清洗液清洗 5 次。完成 5 次清洗後,每孔加 入100µL 的二次抗體 (goat anti-chicken : HRPO conjugate),並置於室溫下作用 30 分鐘。30 分鐘後重複清洗步驟5 次,每孔再加 100µL TMB 溶液於室溫下作用 15 分鐘以呈色,隨後 每 孔 加 入 100µL 0.12% hydrofluoric acid 以 終 止 反 應 。 終 止 反 應 後 以 分 光 光 度 計 (Bio-Kinetics reader, EL 312e) 650 nm 波長讀取吸光值。之後計算各樣本的 S/P 值,若樣本 S/P 值大於或等於 0.6 則定為抗體陽性樣本。
4.4 抗原檢測
在 家 禽 白 血 病 病 毒 抗 原 檢 測 方 面 , 除 了 使 用 商 用 酵 素 連 結 免 疫 吸 附 分 析 套 組 (Enzyme-linked immunosorbent assay kit, ELISA kit, IDEXX, Westbook, Maine)來檢測各樣本 中 的 家 禽 白 血 病 GSA (p27)抗原外,也應用自行塗鍍兔子抗 GSA (p27)抗原抗血清 (rabbit-anti-p27 antiserum, Charles River, North Franklin, CT) 之 ELISA 平盤檢測樣本中之抗
原。在商用酵素連結免疫吸附分析套組使用方面,首先取出ELISA 試劑套組置於室溫,並
紀錄各樣本位置。以二重複加陽性、陰性對照樣品100µL 於檢測盤 A1 至 A4 孔中,然後再 依紀錄加入100µL 待測的樣品,在室溫下靜置 60 分鐘。倒除盤中所有液體,每孔加入去 離子水或蒸餾水 200µL 清洗,此步驟重複 4 次。結束清洗步驟後,每孔加入二次抗體 (Rabbit anti-p27: HRPO conjugate) 100 µL,放置室溫下 60 分鐘,倒掉二次抗體並重複上述的清洗步 驟5 次後,加入 100 µL 的 TMB 溶液,靜置於暗處 15 分鐘,加入 100 µL 終止溶液 (Stop solution, 0.12% hydrofluoric acid) 以終止呈色反應。用分光光度計 (Bio-Kinetics reader, EL 312e) 以波長 650 nm 來讀取結果。之後計算樣品的 S/P 值 (sample – NC / PC – NC),若樣 品的S/P 值大於或等於 0.2,則判定樣品為 GSA 陽性。
4.5 細胞培養
本實驗用於增殖病毒的細胞株為購自 American Type Culture Collection (ATCC, Manassas, VA) 之 DF1 細胞株 (ATCC CRL-12203),其為對內源性家禽白血病病毒具有 抗性的雞胚胎纖維母細胞。
當細胞長滿25 cm2細胞培養瓶時即需進行細胞繼代工作。首先將培養瓶中變黃變 酸的培養液倒掉,以5 mL PBS 溶液 (GIBCO BRL, CA) 清洗三次,在加入 1 mL 0.15% Trypsin 溶液覆滿細胞表面,於 37℃培養箱中作用 2 到 3 分鐘,待細胞圓形化可被輕拍 下 來 後 即 加 入 含 10% 胎 牛 血 清 (Hyclone, Logan, Utah) 及 廣 效 性 抗 生 素 (Life Technologies, Grand Island, NY) 之 Dulbecco’s Minimum Essential Medium (DMEM, pH 7.0-7.2) (Life Technologies, Grand Island, NY) (Himly et al., 1998) (Schaefer-Klein et al., 1998),以 pipetmen 吸放數次使細胞懸浮液均勻後留下 1/4 到 1/6 的細胞後再加入新鮮的 培養液,並置於39℃,含 5%二氧化碳的培養箱培養。 4.6 病毒增殖與病毒核酸偵測 4.6.1 病毒來源 本實驗選用兩個實驗室台灣家禽白血病 J 亞群分離株,分別為分離株 2921/ 00 及 N2/ 01。其中分離株 2921/ 00 為肉種雞腫瘤分離出之病毒; 分離株 N2/ 01 則為有色雞 來源的分離株,且兩者之 env 基因序列分別位於台灣家禽白血病 J 亞群分離株親緣樹狀 關係圖之不同群內 (徐, 2002)。 4.6.2 病毒增殖 將長滿25 cm2培養瓶的DF1 細胞以 0.15%Trypsin 消化下來後分裝於 6 孔細胞培養 盤。隨後每孔加入200µL 的病毒液 (Schaefer-Klein et al., 1998),並加入已回溫且含 10% 胎牛血清及2µg/ mL DEAE-dextran (Sigma, St. Louis, MO) (Fadly et al., 1998) 之DMEM 細胞培養液2 mL,隔天更換不含 DEAE-dextran 並含有 0.1% 胎牛血清 (Hyclone, Logan, Utah) 之 DMEM 細胞培養液 (Life Technologies, Grand Island, NY),並放入 39℃,含 5% 二氧化碳的培養箱培養。培養7 到 9 天後,收集細胞上清液,再以 RT-PCR 的方式檢測 是否有病毒釋出。
4.6.3 病毒核酸偵測 4.6.3.1 病毒 RNA 之萃取
將250µL 之細胞上清液或是血漿加入 750µL 之 TRIzol® Reagent (Life Technologies, Frederick, MD) 後混勻,並在室溫下靜置 5 分鐘,再加入 200µL chloroform (Merck, Darmstadt, Germany) 劇烈搖晃混勻,而後再於室溫下靜置 5 分鐘,接著於 4℃下,以 12000 rpm 離心 15 分鐘,離心後可見到液體分為三層,最下面為有機層,中間為蛋白質 層,最上面則為水層。取出500µL 含 RNA 的水層放入新的 1.5mL 離心管中,再加入 500µL 的isopropanol (Merck, Darmstadt, Germany),混合均勻後,於室溫下靜置 10 分鐘,再於 4℃下以 12000 rpm 離心 15 分鐘。完成離心後,將上清液去除,再加入 1mL 75%酒精, 然後在4℃下以 12000 rpm 離心 5 分鐘後,除去酒精,並以真空乾燥 5-10 鐘。完全乾燥 後,取20µL 經 0.1% DEPC (Diethylpyrocarbonate) (Sigma, St Louis, MO) 處理過之二次 蒸餾水將RNA 溶解,在置入 70℃水浴 3 分鐘以解開 RNA 之二級結構,隨後馬上做冰
浴。
4.6.3.2 細胞之 DNA 萃取
將已形成單一層細胞(mono-layer)的細胞以 Trypsin 將細胞消化下來,再以 1000 rpm 離心10 分鐘。去除上清液後,再加入適量的消化溶液(digestion buffer)(100mM NaCl、 10 mM Tris-Cl,PH8.0、25 mM EDTA,pH8.0、0.5﹪sodium dodecyl sulfate、0.1 mg/ mL proteinase K(Promega, Madison, WI)),置 56℃隔夜進行消化反應。隔日再加入 50 µL RNase A , 並 於 37℃ 作 用 1 小 時 。 完 成 後 即 加 入 同 體 積 的 25 : 24 : 1 phenol/chloroform/isoamyl alcohol(Amresco®, Solon, OH)進行萃取,上下混勻後以 13000 rpm 離心 10 分鐘。此時可見消化液分成上下 2 層,上成為層清透明的水層,下層為有 機層。吸取上層含 DNA 的水層置新離心管,再加入 2 倍體積冰的 100﹪酒精(Merck, Darmstadt, Germany)進行沉澱,此時應可見成雲絮狀之 DNA 產物。以 2000 rpm 離心 20 分鐘後,去除上清液,並加入 75%的冰酒精,上下混勻。再以 2000 rpm 離心 10 分鐘 後,移除酒精,將離心管倒置並以紙沾除多餘酒精,而後以真空抽氣裝置抽乾剩餘酒精。 抽乾後加入適量的二次蒸餾水溶解DNA。
4.6.3.3 反轉錄反應 (RT)
取200µL 之微量離心管(AB gene House, Surrey, UK),依序加入 1µL 40 pmol/ µL 之反向引子 (湃瑪士, 台北市, 台灣)、1µL 10 mM 之 dNTP (Promega, Madison, WI)、6.5µL 之0.1% DEPC (Diethylpyrocarbonate) (Sigma, St Louis, MO) 處理過之二次蒸餾水,最後 再加入 5µL RNA,共 13.5µL。接著將微量離心管置於 60℃ 2min,而後使其降至室溫 27℃,以增進引子與 RNA 模板的粘合。然後再依序加入:4µL 的 5 x buffer(250 mM Tris-HCl [pH 8.3], 375 mMKCl, 1.5 mM MgCl2)、2µL DTT(Life Technologies, Frederick,
MD)、0.4µL,濃度為 40 U/µL 的 Recombinant ribonuclease inhibitor (RNASEOUTTM) (Life Technologies, Frederick, MD)、0.1µL,濃度為 200 U/µL Moloney Murine Leukemia Virus (M-MuLV) reverse transcriptase (RevertAidTM M-MuLV reverse transcriptase) ( Life Technologies, Frederick, MD),使總體積為 20µL。置於 Gene Amp®PCR System 2700® (Applied Biosystem, CA) 中進行反應。反應條件設定為 42℃,90 分鐘,然後再以 70℃, 15 分鐘終止反轉錄酵素的作用。每管取 2µL 以進行聚合酵素鏈反應(PCR)的反應。
4.6.3.4 聚合酵素鏈反應(PCR)
取0.2 mL 之 PCR 微量離心管(AB gene House., Surrey, UK),依次加入:39.5µL 之 2 次 蒸餾水、5µL Taq DNA polymerase 10x buffer containing 15 mM MgCl2 (50 mM KCl, 10
mM Tris-HCl [pH 9.0 at 25℃], 1.5 mM MgCl2, 0.1﹪X-100®) (Promega, Madison, WI)、1µ
之10 mM 之 dNTP (Promega, Madison, WI)、各 1µL 之上下游引子(湃瑪士有限公司, 台北市, 台灣),其濃度為 40 pmole/µL、0.5µL Taq DNA polymerase (Promega, Madison, WI),其濃度為 5 U/µL。使其體積為 48µL,最後再加入 2µL 之 DNA 樣本使最終總體積 為 50µL。接著將 0.2 ml 微量離心管置入 GeneAmp® PCR System 2700 (Applied Biosystems,Foster City, CA)中進行 touch dowm PCR 反應(Don et al., 1991)。PCR 反應 條件方面,除了煉合反應(annealing temperature)的溫度因引子不同而有異外 (Table 3.),其餘條件均相似。以 H5/ H7 為例,其反應條件如下:
94℃、1 分鐘;60℃、1 分鐘;72℃、1 分 30 秒,而煉合反應的溫度每循環一次下降一 度,由60℃降至 48℃,共 13 個循環。最後再以 94℃、1 分鐘;48℃、1 分鐘;72℃、 1 分 30 秒進行 30 個循環,最終以 72℃ 10 分鐘使反應完全。反應完之 PCR 產物如馬上 使用,則置於-20℃冰箱中保存。 4.7 聚合酵素鏈反應產物之選殖 (cloning) 4.7.1 聚合酵素鏈反應產物之純化 聚合酵素鏈反應 (PCR) 產物在進行 TA cloning 前須先進行產物的純化,以除去多 餘的引子以及其他非特異性的PCR 產物。PCR 產物之純化是以 Gel-MTM gel extraction system 進行。首先將 PCR 產物以 50 伏特進行電泳,已使非特異性之雜 band 與目標產 物區分明顯,電泳完成後以EtBr 染色,染色後將膠體置於 UV box 上,並以乾淨的刀片 切下目標大小處之膠體,而後將膠體放入乾淨的小管中,加入 3 倍膠體體積的 GEX buffer。接著將小管置於 60℃加熱約 5-10 分鐘,且每 1-2 分鐘翻轉混合小管內物質,至 膠體完全溶解後使之恢復室溫。接著取一個Gel-MTM column 置於收集小管上,再取以 恢復室溫的膠體混合物放入Gel-MTM column 內,以 12000 rpm 離心 1 分鐘後除去離出 的液體。而後加入0.5 mL WF buffer,以 12000 rpm 離心 1 分鐘後同樣移除離出的液體。 接著再加入0.7 mLWS buffer,以 12000 rpm 離心分鐘並除去離出液體後,再以 12000 rpm 離心3 分鐘以除去殘餘的酒精。離心除去殘餘酒精後,將 Gel-MTM column 取出放於新 的小管上,並於Gel-MTM column 中心加入 30-50 µL 二次蒸餾水,靜置 2 分鐘後,以 12000 rpm 離心 2 分鐘將 PCR 產物自 Gel-MTM column 洗出。洗出之 PCR 產物可置於-20℃保 存。 4.7.2 聚合酵素鏈反應產物之選殖
TA-cloning 的方法依照廠商(Invitrogen., Carlsbad, CA)所附之說明書,其原理主 要是利用 Taq 酵素在反應進行完時會於序列 3’末端再多加一個 A,利用這一多出來的 A 與可與一 3’端多出一個 T 的載體(vector)有效的進行接合反應(ligation)。在進行接 合反應前,須估算加入之PCR 產物的量(ng),使其最終與載體(50ng)的比例為 1-3: 1。將 pCR® 2.1 置於冰上解凍,再依序加入:適量的 PCR 產物、1µL 10X Ligation Buffer、 1µL pCR® 2.1 vector、再加入 DDW 使總體積至 9µL,最後加入 1µL 的 T4 DNA Ligase, 使總體積為10µL,而後於 14℃放置隔夜。隔日進行轉形反應(transformation),如不馬 上進行,則將產物置於-20℃冰箱保存。轉形反應皆於冰上操作,在反應進行前先取出 SOC 培養液(Life Technologies, Grand Island, NY)及含 100µΜ /mL ampicillin(Sigma, St Louis, MO)之 LB 培養基(Life Technologies, Frederick, MD)回溫,並將水浴槽溫度定 為42℃。再於冰上解凍一 50µL 之勝任細胞(competent cell),勝任細胞解凍後加入2µL 之接合反應產物,接著靜置冰上30 分鐘。30 分鐘後取出含勝任細胞之離心管於 42℃水 浴槽,進行heat shock 60 秒,期間注意不可搖晃或震盪離心管。反應完後隨即置於冰上 2 分鐘,再加入 250µL 回溫之 SOC 培養液,接著於 37℃,225rpm 培養 1 小時,使細胞 恢復活性。同時取出 LB 培養基塗抹 40µL,濃度為 40 mg/mL 之 X-Gal (Promega, Madison, WI) 及 40µL,濃度為 100 mM IPTG(Promega, Madison, WI)。1 小時後取出 50µL 菌液塗於上述之 LB 培養基,並將剩餘之菌液以 6000 xg 離心 1 分鐘,倒掉多餘之 SOC 培養液使體積剩約 100µL,再將這 100µL 經 vortex 後,均勻的塗於另一 LB 培養
基上。將 2 個培養基置於 37℃隔夜培養,隔日挑出白色的菌落進行質體的檢定,並將 挑剩下之培養基置於4℃冰箱中保存。
4.8 建構表現載體 4.8.1 目標基因之增幅
針對分離株2921/ 00 之 env 基因 N 端序列設計一組引子 ALVa/ ALVb 並分別於引子的 5´端接上 EcoRⅠ與 BamHⅠ的切位,以 ALVa/ ALVb 可夾出 662 bp 片段,此片段主要 encode 的蛋白質為ALV-J 之 SU (gp85) 的 N 端胺基酸序列,此片段包括了 vr2 (variable region 2)、 hr1 (host range region 1)、hr2 (host range region 1) 三個高變異區。
4.8.2 構築表現載體
將ALVa/ ALVb 所增幅出之 PCR 產物經限制酶 EcoRⅠ與 BamHⅠ處理後,將 PCR 產 物與相同處理之載體 pRSET B (Invitrogen, CH Groningen, the Netherlands)進行接合,此表現 載體命名為pRSET 2921,而其經 IPTG 誘導表現出之重组蛋白命名為 ”gp85N ”,其分子量 為28.15 kDa。
除此之外,我們還有王金和老師自美國Avian Disease and Oncology Laboratory (ADOL) 所帶回的表現載體pGEX RavSu。pGEX RavSu 為含有家禽白血病 A 亞群病毒部份 env 基因 (gp85 gene),而其經 IPTG 誘導表現出之重组蛋白命名為 ”RAV1 ”。
4.9 重組蛋白之表現與纯化
4.9.1 勝任細胞的製備與表現載體的轉殖
以無菌操作方式取ㄧ E. coli BL21(DE3) 或是 E. coli BL21 (Invitrogen, Carlsbad, CA) 單一菌落於50 mL LB broth 中,於 37℃,250 rpm 下隔夜培養。隔日取 4 mL 菌液加入 400 mL LB broth 中,再於 37℃,250 rpm 下培養至菌液吸光值 OD590為0.375 後,將菌液分裝 於8 管 50 mL 離心管中,並冰浴 5 到 10 分鐘後,再於 4℃下以 1600g 離心 7 分鐘。離心後 去除上清液,每管再以10 mL 預冷之氯化鈣溶液 (60 mM CaCl2, 15% (v/v) glycerol, 10mM PIPES pH7.0) 輕緩的將菌體懸浮起來,於 4℃下以 1100g 離心 5 分鐘。去除上清以後每管 再加入10 mL 預冷氯化鈣輕緩的將菌體懸浮起來,接著置於冰上冰浴 30 分鐘後,在 4℃下 以1100g 離心 5 分鐘。而後去除上清液再以 2 mL 預冷氯化鈣輕緩的將菌體懸浮起來,最後 每250µL 分裝成一管,馬上放於-70℃冷凍保存。 取10 ng 建構好的表現載體 DNA (約 10-25 µL) 於 1.5 mL 小管中,再加入 100 µL 勝 任細胞後進行冰浴10 分鐘,此時 DNA 會因勝任細胞上鈣離子所帶的正電而被吸引至勝任 細胞外圍,故此時不可隨意移動反應小管。待10 分鐘後取出冰浴中的反應小管並將之置於 42℃水浴,進行 Heat shock,此時勝任細胞因熱刺激而於其細胞外壁產生小孔而使原本附 於勝任細胞外之DNA 被吸入細胞內,完成轉殖。完成轉殖後的細胞仍然相當脆弱,因此馬 上加入1 mL 不含任何抗生素之 LB broth,於 37℃,250 rpm 下培養 1 小時,使其恢復。1 小時後將菌液做適當的稀釋後塗於含100 µg/ mL ampicillin 之 LB agar 上,以篩選出具有表 現載體之菌落。篩選出具ampicillin 抗性之菌落更進一步做限制酶切割及定序來確定載體含 有目標片段DNA。 4.9.2 重組蛋白之表現
RNA polymerase 的基因,而 T7 RNA polymerase 基因可由 lac UV5 promoter 控制,因此可 利用IPTG (isopropyl β-D-thiogalatopyranoside)來誘導基因表現,進而合成出大量的重組蛋 白。將確定轉殖成功的細胞以無菌操作的方式取ㄧ單一菌落於SOB 培養液中,在 37℃,225 rpm 下隔夜培養。隔日取 3 mL 隔夜培養菌液至 200 mL SOB 培養液中,於 37℃,225 rpm 下培養至菌液吸光值 OD600為 0.3。而後加入 IPTG 並使其最終濃度為 1 mM。6 小時後以
5000 g 離心 15 分鐘後去除上清液。再以 20 mL Lysis buffer 1 (50 mM Tris-HCl pH7.8, 5% glycerol, 0.1 mM EDTA, 200 mM NaCl) 懸浮菌體後,在加入 10 mL Lysis buffer 2 (50 mM EDTA, 10% Triton X-100),於 4℃下作用一整晚。隔日以超音波震盪機破菌。完成破菌後於 4℃下以 12000 g 離心 15 分鐘,收集上清液與沉澱物,進行蛋白質電泳以確定重組蛋白為 可溶或不可溶蛋白。若重組蛋白位於沉澱物中,則將沉澱物先以4.5 M urea 溶液 (10 mM Tris-HCl pH 7.0, 4.5 M urea, 0.1 M 2-mercaptoethanol, 5% glycerol) 隔夜溶解後,於 4℃下以 12000 g 離心 15 分鐘,收取上清液,剩下之沉澱物再加入 6 M urea 溶液 (10 mM Tris-HCl pH 7.0, 6 M urea, 0.1 M 2-mercaptoethanol, 5% glycerol) 處理 6 小時後,再於 4℃下以 12000 g 離心15 分鐘,收取上清液,殘餘之沉澱物再加入 8 M urea 溶液 (10 mM Tris-HCl pH 7.0, 8 M urea, 0.1 M 2-mercaptoethanol, 5% glycerol) 隔夜處理後,於 4℃下以 12000 g 離心 15 分鐘,
收取上清液。將不同濃度urea 溶液所收得之蛋白質溶液進行電泳以確認重組蛋白較易溶解
於何種濃度中。
4.9.3 重組蛋白之純化
重組蛋白純化主要是使用商業純化套組 Ni-TEDTM (Protein Purification Silica-based purification of 6x His-tagged protein) (Active Motif, Carlsbad, USA) 來進行,首先將粗蛋白加 入純化管柱中,再以2000g 離心 2 分鐘,去除流出液後加入 600µL Buffer A,以 2000g 離心 2 分鐘,去除流出液後再重複加入 600µL Buffer A,以 2000g 離心 2 分鐘,最後加入 100µL Elution Buffer 以 2000g 離心 3 分鐘洗出重組蛋白,此步驟可重覆 4 次,一共可收得 400µL 純化好之重組蛋白。
4.10 重組蛋白之應用
4.10.1 gp85N 塗鍍 ELISA 平盤之製備與應用-Indirect ELISA
將純化好的gp85N 以 coating buffer 稀釋成 50ng/ dL 與 100ng/ dL,再分別以 50ng/ well 及100ng/ well 之 gp85N 加入 ELISA 平盤中 (Immuno-plate, NUCK),並置於室溫下隔夜塗 鍍後,用340µL/ well 之 PBST (Phosphate buffer saline + 0.1% Tween-80)清洗 1 次,以除去未 結合在ELISA 平盤上的 gp85N。接著加入 100µL/ well 的 blocking buffer (Phosphate buffer saline + 0.1% Tween-80 + 5% nonfat dried milk) 於 ELISA 平盤中,置於室溫下作用 30 分鐘, 以將各well 內的間隙補滿。而後再以 340µL/ well 之 PBST 清洗 1 次以除去多餘的 blocking buffer。將血清以 PBS 做一定倍數稀釋後,分別以 100µL/ well 加入 ELISA 平盤中,再將之 置於37℃恆溫箱中作用 30 分鐘,而後以 340µL/ well 之 PBST 清洗 3 次,除去多餘的血清 後再加入100µL/ well 以 PBS 做 2500 倍稀釋 HRP labeled goat-anti-chicken IgG conjugate (Kirkegaard & Perry Laboratories INC., Gaithersburg, MD),於 37℃恆溫箱中作用 30 分鐘後, 以340µL/ well 之 PBST 清洗 5 次,最後加入 100µL/ well 的呈色劑 (1 tablet OPD + 12mL substrate buffer + 8µL 30% H2O2) 避光作用 10 分鐘後,加入 100µL/ well 之 stop solution (1M
nm 波長之吸光值。
4.10.2 不同 gp85N 塗鍍量對抗體偵測結果之影響
為了找出最好的抗原塗鍍量,因此使用不同稀釋倍數 (100 倍、200 倍及 400 倍稀釋) 的 家禽白血病病毒抗血清 (Charles River, North Franklin, CT ) 分別與 50 ng/ dL 及 100 ng/ dL 純化好的gp85N 作用,藉由 OD490的結果來評估抗原的最佳塗鍍量,且不同處理組合均有
3 個血清樣本。
4.10.3 不同血清種類與血清稀釋倍數對抗體偵測結果之影響
為尋求最佳的血清稀 釋條件,先利用不同 亞群之家禽白血病病 毒抗血清 (anti-J antiserum、anti-A antiserum、anti-B antiserum 及 14 週齡 SPF 雞隻血清) 與不同的血清稀釋 倍數 (100 倍、200 倍及 400 倍稀釋) 來觀察 gp85N 的抗原性及不同稀釋倍數下,各血清樣 本與gp85N 間的作用情形,以進一步找出可區分不同亞群之家禽白血病病毒抗血清之最佳 稀釋條件,應用於ELISA 上。將各血清以 PBS 做 100 倍、200 倍及 400 倍稀釋後,分別以 100µL/ well 加入 ELISA 平盤中。不同處理組合都有 3 個血清樣本。 4.11 單株抗體之生產與應用 4.11.1 單株抗體之生產 單株抗體之生產主要是委託濁水溪公司 (台北, 台灣)進行,其中 BALB/c 小鼠之免 疫計畫如下:將含有目標DNA 片段之質體以 25µg/ mL,每次 2 mL 的劑量,用靜脈注射方 式免疫4 隻 BALB/c 小鼠,每二週免疫一次,共免疫四次,而後以 2921/ 00 分離株感染之 DF 1 細胞株進行免疫化學染色,確認小鼠血清免疫效價。接著選擇具最高效價者進行細胞 融合,而後對細胞培養液進行二次陽性孔的篩選,最後選定 2 株陽性株進行單株化選殖, 分別命名為,mAb14 及 mAb22。 4.11.2 單株抗體之確認
4.11.2.1 Immunodot blot assay
取已純化 2921/ 00 分離株、未純化之 gp85N 粗蛋白、pRSET B 空載體經 IPTG 誘導表 現之粗蛋白,以及未純化之RAV1 粗蛋白 (美國 ADOL) 與 mAb14 及 mAb22 進行 Immunodot blot assay 以初步確認單株抗體 mAb14 及 mAb22 是否只會辨識 ALV-J 的抗原。首先將所有 樣本稀釋到 1µg/ mL 後,取 1 µL 樣本點至硝基纖維濾紙 (nitrocellulose membrane) (Protran® nitrocellulose membrane, 0.45µm, Schleicher & Schuell),再於室溫下乾燥 30 到 60 分鐘。接 著將點上樣本的硝基纖維濾紙 (nitrocellulose membrane) (Protran® nitrocellulose membrane, 0.45µm, Schleicher & Schuell)置入 0.25% Gelatin-NET 溶液 (0.25% gelatin, 0.15 M NaCl, 5mM EDTA-Na, 0.5% Tween 20, 50mM Tris) 於室溫下緩慢震盪 60 到 90 分鐘以將無蛋白質 部份的硝基纖維濾紙做阻隔。完成阻隔後加入以Gelatin-NET 溶液做 5000 倍及 10000 倍單 株抗體mAb14 及 mAb22,於室溫下緩慢震盪作用 1 小時。接著以 washing buffer (PBS, 0.05% Tween 20, pH 7.4) 洗硝基纖維濾紙 4 次,每次 5 分鐘。然後加入以 Gelatin-NET 溶液做 2500 倍稀釋的Goat anti-mouse-HRP conjugate (Kirkegaard & Perry Laboratories INC., Gaithersburg, MD),於室溫下感作 1 小時。重複清洗步驟 4 次後,取出硝基纖維濾紙,加入 TMB (Kirkegaard & Perry Laboratories INC., Gaithersburg, MD) 進行呈色。
4.11.2.2 免疫螢光分析 (immunofluorescence assay, IFA) 取培養於24 孔細胞培養盤上且確定感染分離株 2921/ 00 之 DF1 細胞,以 acetone: 95% 酒精 (6: 4) 於-20℃下進行細胞固定 10 分鐘。10 分鐘後倒除固定液並將細胞培養盤置於室 溫下乾燥至少30 分鐘。完成乾燥後將 primary antibody 以 PBS 稀釋 500 倍後再於感染細胞 於室溫下作用1 小時,期間要時常觀察避免 PBS 乾掉。1 小時後取出玻片以 PBS 洗去多餘 的primary antibody,清洗時需小心勿直接沖洗到細胞,接著加入以 PBS 稀釋 150-200 的螢 光標示的secondary antibody,同樣於室溫下避光作用 1 小時。1 小時後重複清洗步驟以移 除多餘的抗體。接著於細胞滴上10% mounting solution (10% glycerol, 90% PBS) 並進行封 片,再以螢光顯微鏡觀察結果。
4.11.2.3 單株抗體 之應用-Sandwich ELISA
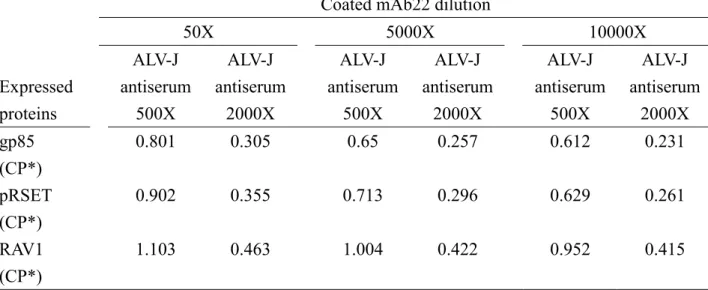
確認單株抗體辨識 ALV-J 抗原能力後,將單株抗體或 Subgroup J avian leukosis antiserum (Charles River, North Franklin, CT) 以 coating buffer 做 500 倍、5000 倍及 10000 倍 稀釋,再分別以100 µL/ well 之稀釋抗體加入 ELISA 平盤中 (Immuno-plate, NUCK),並置 於室溫下隔夜塗鍍後,用340µL/ well 之 PBST (Phosphate buffer saline + 0.1% Tween-80)清洗 1 次,以除去未結合在 ELISA 平盤上的 gp85N。接著加入 100µL/ well 的 blocking buffer (Phosphate buffer saline + 0.1% Tween-80 + 5% nonfat dried milk) 於 ELISA 平盤中,置於室 溫下作用30 分鐘,以將各 well 內的間隙補滿。而後再以 340µL/ well 之 PBST 清洗 1 次以 除去多餘的 blocking buffer。將病毒或重组蛋白以 PBS 稀釋至相同的濃度後,再分別以 100µL/ well 加入 ELISA 平盤中,將之置於 37℃恆溫箱中作用 30 分鐘,而後以 340µL/ well 之PBST 清洗 3 次,除去未結合的抗原後再加入 100µL/ well 以 blocking buffer 做 500 倍和 2000 倍稀釋的 Subgroup J avian leukosis antiserum (Charles River, North Franklin, CT) 或單株 抗體,於37℃恆溫箱中作用 30 分鐘後,再以 340µL/ well 之 PBST 清洗 5 次,再加入 100µL/ well 以 blocking buffer 做 2500 倍稀釋 HRP labeled goat-anti-mouse IgG conjugate,於 37℃ 恆溫箱中作用30 分鐘後,以 340µL/ well 之 PBST 清洗 5 次,最後加入 100µL/ well 的呈色 劑 (1 tablet OPD + 12mL substrate buffer + 8µL 30% H2O2) 避光作用 15 分鐘後,加入 100µL/
well 之 stop solution (1M H2SO4) 以終止呈色反應。再以分光光度計 (Bio-Kinetics reader, EL
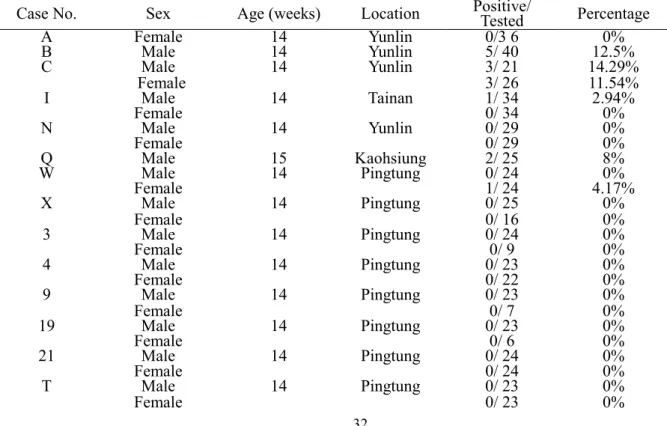
5. 結果 5.1 台灣有色雞群之血清學調查 自2002 年 11 月起至 2003 年 4 月止,一共調查了 16 群有色雞群,其中公母各半。血 液樣本大致分為兩個來源,其一是種雞場來源的雞隻抗凝血樣本,另外則為屠宰場來源之 抗凝血樣本。各群雞隻之基本資料詳見於Table 1.,其中病例 3068 於病雞剖檢時,在其肝 臟見有腫塊;病例3070 之母雞有產蛋率下降的情形;病例 3093 的雞隻則有蒼白、跛腳的 情形發生;病例3094 的公雞正常,剖檢也未見有異常,但是母雞則外觀蒼白,剖檢時發現 肝臟、脾臟腫大;病例3095 有跛腳情形,剖檢後見有肝脾腫大且有壞死點。本實驗所檢測 的有色雞品種包括安樂、珍珠雞、紅羽土雞等;採樣年齡主要針對12 週齡以上 ALV-J 抗體 陽轉的雞群 (Hunt et al., 2000)。利用 ALV-J 商業用酵素連結免疫吸附套組 (Enzyme-linked immunosorbent assay kit, ELISA kit, IDEXX, Westbook, Maine) 調查台灣北部 4 群雞群、中部 10 群雞群、南部 2 群雞群 ALV-J 抗體分佈情形,以了解 ALV-J 在台灣土雞發生情形。調查 結果發現北部雞群100% (4/4) 為抗體陽性,中部雞群則 80% (8/10) 為抗體陽性,南部雞群 則 0% (0/2) 為抗體陽性;其中公雞群與母雞群抗體陽性比率分別為 75% (6/8) 和 75% (6/8),兩者並沒有差異;種用雞群與肉用雞群的抗體陽性比率分別為 100% (10/10) 和 33.3% (2/6),種雞群抗體陽性比率較肉雞群為高。各雞群之抗 gp85 抗體之陽性率詳列於 Table 1.。 2004 年 2 月至 2004 年 4 月再次針對屠宰場來源雞隻進行 ALV-J 抗體檢測,一共調查 40 群有色雞,其中公雞 20 群,母雞 20 群。其中公雞群 15% (3/ 20)為抗體陽性,母雞群則 10 % (2/ 20) 為抗體陽性。抗體陽性率由 0%到 14.29%,結果詳見 Table 2.。 5.2 2002 年至 2004 年肉雞、蛋種雞與有色雞種雞之家禽白血病 J 亞群病毒分離情形 由2002 年至 2004 年一共自 11 個肉雞病例、8 個有色雞病例及 2 個蛋種雞病例分離出 家禽白血病J 亞群病毒,各病例之統整比較見 Table 4.,另詳述如下: 病例 3062 為 2002 年 11 月為台中縣一肉種雞場送檢病例,其雞隻品系為愛拔益加 (Arbor Acres, AA),飼養雞隻總數為 2600 隻,送檢時為 16 週齡,平均每天死亡或淘汰 5 至 6 隻雞隻。其雞隻採開放式飼養,據雞場獸醫師表示此批雞隻自 2002 年 8 月入雛,雞隻 10 週齡時解剖發現胸骨中央有白色腫瘤,自14 週齡起雞隻死亡數量開始增加,雞隻除了瘦弱 外,也有明顯蒼白,骨髓顏色變淡,其他無明顯病灶。此批雞隻也曾抽血送大成公司檢測 雞傳染性貧血 (CIA),此外同一雞場其他品種雞隻 (Avian) 也有相同問題,唯發病週齡愈 來愈早,損失的雞隻一批比一批多。此批雞隻於剖檢時見有腺胃及十二指腸有卡他樣滲出 液、腎臟萎縮褪色且有尿酸鹽沉積、肝臟亦有萎縮褪色、脾臟萎縮、胰臟萎縮、皮下有滲 出液、肌肉呈煮肉樣、骨骼脆弱、骨髓呈桃色。其送檢樣本為10 個公雞抗凝血液樣本,將 血漿同時以商用酵素連結免疫吸附分析套組檢測家禽白血病GSA (p27)抗原,以及進行病毒 分離工作。在ELISA 檢測 GSA (p27)抗原部份,10 個血漿樣本中有 5 個為抗原陽性 (5/ 10) ; 病毒分離部分每2 各血漿樣本 pool 成 1 個樣本進行病毒分離,經兩次繼代後成功分離出 1 株家禽白血病J 亞群病毒。 病例 3067、3068、3069 及 3070 為 2003 年 1 月,由苗栗同一飼主所送檢之病例。其 中3067 為安樂品系 10 週齡之種公雞,一共飼養 60 隻,以籠飼方式飼養,此次發病共淘汰 掉10% (6/ 60) 雞隻,據雞場主人表示雞隻主要發生跛腳為主,先腳弱而衰弱,最後死亡。 3068 則同為安樂品系 10 週齡之種母雞,總飼養隻數為 1800 隻,部份母雞採平飼,部份採
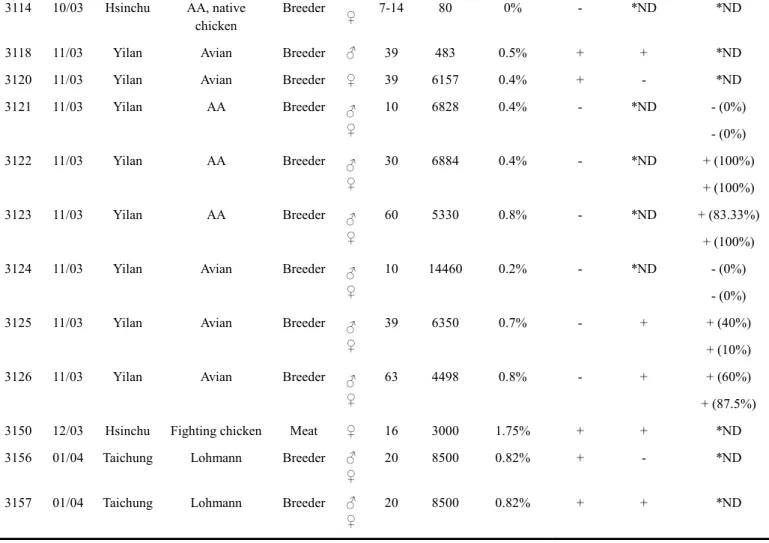
籠飼,平飼部分母雞有跛腳的情形,此次發病一共淘汰掉22% (400/ 1800) 雞隻,週死亡率 約2.2%,此批雞隻同樣發生軟腳、衰弱死亡的情形,另外雞場主人亦表示此批雞隻於 5 週 齡剖檢時,於肝臟見有腫瘤。3069 為 69 週齡之公珍珠雞種雞,一共飼養 100 隻,採籠飼, 此次發病淘汰掉10% (10/ 100) 雞隻,且其後代品質差,有千分之 3 至 5 的後代有跛腳,7 至8 週齡的大雞咳嗽不易治癒。3070 為 69 週齡之母珍珠雞種雞,一共飼養 4800 隻,採籠 飼,此次發病淘汰掉6.25% (300/ 4800) 雞隻,其週死亡率約 0.09%,此批雞隻發生產蛋率 下降 (自 70%產蛋率降至 50%),其後代品質不佳,尤其初產仔雞淘汰率達 15%,此後跛腳 雞隻很多,到7 至 8 週齡時,一有咳嗽就治不好。3067 共送檢 15 個抗凝血液樣本,其中 10 個為外觀正常之公雞,5 個為跛腳雞隻來源;3068 共送檢 60 個抗凝血樣本,其中 40 個 為採籠飼正常母雞隻樣本,20 個為採平飼且發生跛腳之母雞樣本;3069 則有 17 個抗凝血 樣本;3070 則為來自同一雞場 A、B、C 三區各 20 各抗凝血樣本。以商用酵素連結免疫吸 附分析套組檢測家禽白血病GSA (p27)抗原及家禽白血病 J 亞群特異性抗體結果見 Table 5. 及Table 6.,由抗原檢測結果看來有無跛腳或是不同棟別並無太大的差異。接著取 GSA (p27) 抗原 ELISA 檢測結果 OD 值最高者之血漿進行 RT-PCR 檢測,結果 3067 與 3068 為 H5/ H7 陽性,3069 與 3070 為 H5/ H7 陰性。雖然同時進行病毒分離,但未分離出病毒。 3102 與 3104 為 2003 年 10 月自宜蘭同一肉種雞場送檢之病例,3102 為艾維茵 (Avian) 品系,31 週齡雞隻,剖檢 3 隻雞隻於其中 2 隻見有腹膜炎,並於腸繫膜上見有脂肪結節; 3104 為愛拔益加品系(Arbor Acres),22 週齡雞隻,據雞場主人表示此批雞隻共有 18520 隻, 每日淘汰率約為0.16%。3102 一共送檢 2 個公雞血液樣本及 2 個母雞血液樣本,以及公雞 與母雞組織樣本各1 個,共 6 各樣本;3104 則送檢 2 個母雞組織樣本。結果自 3102 中 3 個公雞樣本分離出 3 株家禽白血病 J 亞群病毒,自 3 個母雞樣本分離出 2 株家禽白血病 J 亞群病毒;3104 則自 2 個樣本中分離出 1 株家禽白血病 J 亞群病毒。 3106、3107 與 3108 為 2003 年 10 月台中同一雞場送檢之病例。3106 為 5 個珍珠雞血 液樣本,3107 為 10 個公仿鬥雞血液樣本,3108 為 25 個母仿鬥雞血液樣本。據雞場主人表 示此批14 週齡雞隻共 5000 隻,分為 6 棟於荔枝園內開放飼養,於 8 月初開始發病,期間 死亡約1500 隻,淘汰率為 30% (1500/ 5000),主要見到的臨床症狀為削瘦、營養狀況不佳、 精神萎靡。剖檢4 隻削瘦雞隻中,2 隻見有肝臟腫大 (2/ 4) 且表面有乾酪樣滲出物與胸壁 腹膜粘連 (2/ 4),腹膜與小腸的漿膜呈現混濁有黃色肥厚的纖維素被覆(2/ 4),其他器官則 無可視之病變。將3106 的 5 個樣本、3107 的 10 個樣本、3108 的 25 個樣本進行病毒分離。 接著以商用酵素連結免疫吸附分析套組檢測第一代病毒分離之細胞上清液中之家禽白血病 GSA (p27)抗原時,發現 3106 的 5 個樣本皆為陰性 (0/ 5);3107 的 10 個樣本中有 1 個為抗 原陽性 (1/ 10);3108 中的 5 個樣本中有一個為抗原陽性 (1/ 5),剩下的 20 個病毒分離樣 本則以RT-PCR 檢測,結果有 5 個樣本為 RT-PCR H5/ H7 陽性 (5/ 20),經第 2 次病毒分離 後,3106 未分離出家禽白血病 J 亞群病毒 (0/ 5);3107 則分離出 1 株家禽白血病 J 亞群病 毒 (1/ 10);3108 也分離出 1 株家禽白血病 J 亞群病毒 (1/ 25)。 3114 為 2003 年 10 月新竹 KI 公司所送檢的病例,此批雞隻為育種用雞群,包括愛拔 益加 (Arbor Acres) 品系白肉雞與購自肉雞場之台灣北部大型土雞,少部分為進口土雞,共 送檢80 個血液樣本,其中 16 個為白肉雞血清,64 個為土雞血清,有部分血清形成不良, 雞隻年齡為7 週齡至 14 週齡。由於雞隻並未發病只是一般送檢,且血清樣本已經過一段時
間冷藏不適合進行病毒分離,因此只進行ALV GSA (p27) 抗原檢測,檢測結果詳見 Table 7.。 不同週齡之血液樣本各取GSA (p27) 抗原 ELISA 吸光值最高之 2 個樣本進一步萃取 DNA,
以primer H5/ H7 進行 PCR 檢測,結果 12 週齡的 2 個樣本皆為 H5/ H7 陽性,其餘皆為陰 性。因此進一步將H5/ H7 陽性之樣本以 ev1/ ev2 (阮, 2000),增幅出 3114 provirus 之 gp85 基因片段,並以yT & A TA cloning kit 將此片段選殖出來並定序。
3118、3120 為 2003 年 11 月宜蘭同一肉種雞場送檢之病例。3118 為艾維茵 (Avian) 品 系,39 週齡肉種雞,一共送檢 2 個公雞臟器乳劑與 1 個母雞 (自公雞群抓出) 臟器乳劑; 3120 同樣為艾維茵 (Avian) 品系,39 週齡肉種雞,僅送 1 個母雞臟器檢測。經 2 代病毒分 離後自3118 的 2 個公雞臟器乳劑中各分離出 1 株家禽白血病 J 亞群病毒 (2/ 3);3120 則未 分離出病毒。 3121、3122、3123、3124、3125 與 3126 為 2003 年 11 月宜蘭同一肉種雞場送檢之病 例。其中3121、3122 與 3123 分別為 10 週齡、30 週龄以及 60 週齡之愛拔益加 (Arbor Acres) 品系隻肉種雞;3124、3125 與 3126 則分別為 10 週齡、39 週龄以及 63 週齡之艾維茵 (Avian) 品系隻肉種雞。分別以商用酵素連結免疫吸附分析套組 (Enzyme-linked immunosorbent assay kit, ELISA kit, IDEXX, Westbook, U.S.A.) 檢測家禽白血病 GSA (p27)抗原及家禽白血 病J 亞群特異性抗體。其中 3125 及 3126 進ㄧ步進行病毒分離,經 2 次病毒分離後,自 3125 的5 個公雞血液中分離出 2 株家禽白血病 J 亞群病毒 (2/ 5),並自 8 個母雞血液樣本中分離 出1 株家禽白血病 J 亞群病毒 (1/ 8),3126 則未分離出病毒 (公:0/5,母:0/8)。 病例3150 為 2003 年 12 月由新竹送檢之病例。此病例為 16 週齡之母鬥雞,飼養總數 為3000 隻,採平飼,自 8 到 9 週齡開始發病,每日死亡 7 到 8 隻,雞隻生長發育參差不齊 且見有精神沉鬱,臨床上見有灰白色軟便、雞隻削瘦、有逆毛嗜睡狀況、肉冠呈貧血狀, 剖檢時於肺臟見有結節,消化器官見有寄生蟲,腎臟見有結節及尿酸鹽沉積,肝臟及脾臟 亦見有腫大與結節,另外在心臟也見有結節,華氏囊有腫大的情形。自心、肝、脾釣菌培 養於Blood agar 並未分離出有意義的細菌。3150 共送檢一個新鮮組織乳劑 (由多隻雞隻臟 器混合製成),以及 11 個血清樣本,由於此批血清樣本存放於 4℃下約 1 個月,且已見有溶 血的情形,故僅以唯一的組織乳劑樣本進行病毒分離,血清樣本則以自行塗鍍兔子抗GSA (p27) 抗原抗血清之 ELISA 平盤檢測樣本中之抗原。在進行病毒分離前先抽取臟器 DNA 進 行PCR 檢測,結果發現為家禽白血病 J 亞群病毒陽性 (H5/ H7 陽性) 且為家禽白血病 A 亞 群病毒陽性 (PA1/ PA2 陽性);同時也以馬立克病毒的引子 MD1r/ MD1f (Zhu et al., 1992) 進行PCR 檢測,結果為陰性。另外血清樣本經 ELISA 檢測後發現有 10 個血清樣本 GSA (p27) 抗原陽性 (10/ 11)。而組織乳劑樣本經 2 次病毒分離後,僅自樣本中分離出 1 株家禽白血病 J 亞群病毒而無家禽白血病 A 亞群病毒。 病例 3156 及 3157 為 2004 年 1 月,台中同一蛋種雞場前後送檢病例,此批雞隻為羅 曼品系(Lohmann),共 8500 隻,137 日齡,依雞場主人表示,該場蛋種雞先前均由德國進 口,現因德國為高病原性禽流感疫區而改由西班牙輸入,此批雞隻從輸入後,雛雞明顯發 育不良且大腸桿菌感染嚴重,僅有85%育成率,自 120 日齡起雞隻陸續發生死亡,雞群飼 料攝取量下降 (80 克/ 隻),平均每隻少 20 克,且有產蛋上升率減緩的趨勢,每日約死亡 10 隻,經剖檢見有肝臟腫大、腫瘍及脾臟腫大。臟器經組織切片檢查後,於肝臟見有骨髓 球細胞的浸潤。為進一步做馬立克病與家禽白血病之鑑別診斷,於是萃取臟器 DNA 進行 PCR 診斷,結果以馬立克病毒的引子 MD1r/ MD1f (Zhu et al., 1992) 增幅出 300 bp 左右的 片段,但PCR 產物定序後證實非馬力克病毒序列,另外也以本實驗室設計的家禽白血病引
子ev1/ ev2 檢測臟器 DNA (阮, 2000),結果增幅出 998 bp 的白血病病毒片段。同時間以雞 隻血液樣本每3 個血漿 pool 成一管,公母各 5 各樣本進行病毒分離,經二次繼代後成功自