Vol. 27: 131-139, 1996
1
DISEASES OF AQUATIC ORGANISMSDis Aquat O r g Published November 1 4
1 - p
Identification of white spot syndrome associated
baculovirus (WSBV) target organs in the shrimp
Penaeus monodon
by
in situ
hybridization
Poh-Shing changl, Chu-Fang
L O ~ ,Yu-Chi wang2, Guang-Hsiung
K O U ~ V * 'Department of Aquaculture, National Kaohsiung Institute of Marine Technology, Kaohsiung, Taiwan, ROC2 ~ e p a r t m e n t of Zoology. National Taiwan University, Taipei, Taiwan, ROC
ABSTRACT: Pendc'us monodon non-occluded baculo\ilrus 111 (PmNOB 111) is a strain of white spot syndrome associated baculovii-us (WSBV) isolated from diseased black tiger shrlrnp F! monodon. In order to identify the target organs of WSBV, the tissue sectlons of experimentally infected shrimp sampled at 0, 16, 22, 40, 52, and G4 h post-infection were h \ bridized in situ with PmNOB 111 DNA- specific probe labeled with digoxigenin. WSBV-positlve cells \yere initially observed at 16 h post- infection in the stomach, gill, cuticular epidermis a n d hepatopancreas. At 22 h post-infection, the lym- phoid organ, antenna1 gland, muscle tissue, hematopoietic tlssue, heart, midgut a n d hindgut w e r e found to b e WSBV-positive. T h e nervous tissue and compound eyes did not have WSBV-positive cells until 4 0 h post-infection. Thus various tissues from the mesoderm a n d ectoderm, such a s connective tissue. epithelium, nervous tissue and muscle, can all b e infected by WSBV. By 52 or 64 h post-infection, ~t was found that the stomach, 9111, cuticular epidermis, lymphoid organ, hematopoietic tissue a n d antennal gland w e r e all h ~ d v i l y Infected with WSBV a n d that these tissues had become necrotic. T h e whlte spot syndrome itself first appeared at 40 h post-infection, and the shrimp began to dle at 64 11 post-infection. The probe demonstrated no reactlon to LVSBV-negative tissue.
KEY WORDS: PmNOB I11 WSBV In situ hybridization . White spot baculovirus . Penaeid shrimp
INTRODUCTION
White spot syndrome associated non-occluded bac- ulovirus (WSBV) is the causative agent of a shrimp viral disease. The affected individuals always show white spots on their exoskeleton a n d this is the prin- cipal sign of the disease (Chou e t al. 1995, Wang e t al. 1995). Based on the published description, WSBV is also apparently related to Japan's rod-shaped nuclear virus of Penaeus japonicus (RV-PJ; Inouye et al. 1994, Momoyama et al. 1994, Nakano et al. 1994, Takahashi e t al. 1994), China's hypodermal hematopoietic necro- sis baculovirus (HHNBV; Cai e t al. 1995) a n d Thai- land's systemic ectodermal a n d mesodermal bac- ulovirus (SEMBV; Wongteerasupaya e t al. 1995).
'Addressee for correspondence. E-mail: [email protected]
WSBV 1s extremely virulent, has a wide host range a n d targets various tissues. The known hosts of WSBV include Penaeus japonicus, P monodon, l? chinensis a n d P penicillatus (Cai et al. 1995, Wang et al. 1995). According to a study by Momoyama et al. (1994) on the histopathology of RV-PJ-infected P japonicus, degen- erated cells with basophilic hypertrophied nuclei were observed in various tissues originating from both meso- derm a n d the ectoderm, such a s the cuticular epi- dermis, connective tissue, lymphoid organ, antennal gland, hematopoietic tissue a n d nervous tissue (Momo- yama et al. 1994). The virus can induce 100 % mortality in infected shrimp within 3 to 5 d (Nakano et al. 1994). Since 1993 the virus has been a significant pathogen in cultured shrimp in Asia.
The virions of Penaeus monodon non-occluded bac- ulovirus I11 (PmNOB 111 ), which is a strain of WSBV iso- lated from diseased black tiger shrimp l? monodon, O Inter-Research 1996
132 Dis Aquat Org 27
have been purified. In negatively stained preparations, the virion measures 70 to 150 nm at its broadest point and is 250 to 380 nm long, which is usually 10 % larger than In ultrathin sections (Wang et al. 1995). A PmNOB I11 genomic DNA library was also constructed and a set of specific primers was designed for the polymerase chain reaction (PCR) to detect PmNOB 111 infection in penaeid shrimp (Lo et al. 1996). However, the sites of early viral infection and the stages of the progression of WSBV infection through the shrimp remained un- known.
This paper reports on the use of a PmNOB 111 DNA- specific probe to Identify by in situ hybridization the target organs and tlssu.es of WSBV in experimentally infected shrimp. With this technique, we could identify the major tissues which were sites of initial viral infec- tion a s well as the preferred attack sites of WSBV. We were also able to follow the stages of the progression of WSBV infection in shrimp. Tne results of our current studies will provide important insights into the patho- genic mechanism, early diagnosis a n d disease control.
MATERIALS AND METHODS
Experimental infection. Samples of Penaeus mono- don were obtained from a shrimp farm at Kaohsiung in Taiwan. They were originally determined to be not infected by PmNOB I11 by PCR detection. Detection of the virus by PCR followed the methods of Lo et al. (1996). Three groups of about 30 shrimps weighing 0.35 to 0.45 g were kept in 50 l plastic tanks containing salinity 20 ppt, filtered, aerated sea water at ambient temperature (approximately 28 to 30°C). Two of these groups were fed wlth severely PmNOB 111-infected
P monodon for one meal and then with artificial diet
for the subsequent days. The third group of shrimp served as a negative control and tvere fed with arti- ficial diet throughout the study.
Sampling procedure and treatment. Three shrimp from each group were sampled at 0, 16, 22, 40, 52 and 64 h post-infection (pi). These shrimp were fixed in buffered neutral formalin solution (10"i, formalin, 33 m M NaE12P0,, 45 m.M Na2HP0,). The fixed tlssues were dehyd.rated, embedded in paraffin wax and sec- tioned a t approximately 4 to 5 pm thickness on a rotary microtome. The sections were stained with modified hematoxylin and eosin (H&E stain) for normal histo- logy and the sections from the same tissue blocks were also placed onto 2 % silane pretreated slides for further in situ hybridization (Nuovo et al. 1991).
DNA probe for in situ hybridization. The recombi- nant plasmid pms146 selected from the PmNOBtII Sal I genomic library (Lo et al. 1996) was used for the pre- paration of the DNA probe. The Insert of pms146 was
gel purified and nonradioactively labeled wlth digoxi- genin-dUTP using a random priming method available from Boehringer Mannhelm Biochemical, Bedford, England.
In situ hybridization procedure. The tissue sections were deparaffinized in xylene and rehydrated by a series of graded alcohols (absolute to 50 %) and finally w ~ t h distilled water The sections were soaked briefly in phosphate-buffered saline (PBS; 0.137 M NaC1, 0.0027 M KC1, 0.0043 M Na2HP0,
.
2 H 2 0 , 0.0014 M KH2P0,, pH 7.4) and then placed for 10 min in 0.02 N HCl. After washing twice for 5 min in PBS at room temperature (RT), the slides were treated for 30 min with 100 1-19 ml-' proteinase K in PBS at 37°C. Follow- ing the proteolytic treatment, slides were washed twice for 5 min in PBS containing 0.2% glycine. Post- fixation was carried out by 4 % paraformaldehyde in PBS for 10 min at RT, and then the sections tvere washed in 0.2 % glycine in PBS for 10 min followed by a wash in 2x standard saline citrate (SSC; l x SSC = 0.15 M NaCl, 0.015 M tri-sodium citrate, pH 7.0). These sections were pre-hybridized in 500 p1 hybridization solution (50% formamide, 0.2% Ficoll 400, 0.2% poly- vinylpyrrol.i.done, 0.2 % bovine serum albumin (BSA), 5 x SSC, 50 mM Tris-HC1, pH 8.0, 1 mM EDTA) for 1.5 h at 37OC. Digoxigenin-labeled DNA probes were diluted to 10 ng m1'
in hybridization solution and 100 p1 was applied to each slide. A cover glass was put on top and the slides were placed on a 95°C hot plate for 5 min. The slides were then transferred to a humid chamber a n d incubated at 37°C for 16 to 20 h. After hybridization, the slides were uncovered and washed by 2x SSC for 30 min at RT, followed by l x SSC for 30 min at RT, 0 . 5 ~ SSC for 30 min at 37°C and then 0 . 5 ~ SSC for 30 min at RT.Hybridization detection. The slides were washed briefly in Buffer I (100 mM Tris-HC1, 150 mM NaCl, pH 7.5) and then blocked with Buffer I containing 1 % blocking reagent (Boerhrhger) (Buffer 11) for 30 min at 37°C. The alkaline phosphatase-con.jugated anti- digoxigenin was diluted 1:2500 in Buffer 11. The slides were covered with 300 p1 diluted conjugate and incu- bated for 30 min at 37OC in a dark, humid chamber After washing twice for 15 min with Buffer I containing 0.3 % Tween-20, the slides were equilibrated for 5 min in Buffer I11 (100 mM Tris-HC1, 100 mM NaC1. 50 mM MgCl,, pH 9.5) at RT The slides were covered with 500 1-11 of freshly prcpared color developer [ 4 5 p1 nitro- blue tetrazolium (90 mM) and 35 p1 5-bromo-4-chloro- 3-indoyl phosphate (120 mM) in 10 m1 Buffer 1111 and were then incubated for l h or overnight at RT After incubation, the slides were washed with l x TE buffer for 5 min at RT. Neutral red was used to counterstain the slides for 10 mln an.d then they were dehydrated with a series of graded al.cohols (80'!;) to absolute),
(:hang et al.: WSBV target organs In shrimp 133
before being finally immersed in xylene. Each slide subsequently with sampling time. No experimental was then mounted with a plastic lid by using a histo- shrimp died until 64 h pi, at which time the mortality of logical mounting medium and examined under an the remaining shrimp was 33%. Shrimp in the control Olympus B>;-50 bright field light microscope. group did not show any positive signal in their tissues b y in s i t u hybridization or by PCR, nor did they have any white spots on their carapaces All the shrimp in RESULTS the control gro.up were healthy and survived during
the experimental period (data not shown). Infectivity and mortality of the experimental infection
- -
In situ hybridization" 0 ' ' 33 % 66%) 10OUA, 100";. 100i'4,
White spot syndrome" - -
+
++ +++Mortality C 0/30 0/27 0/24 0/21 0/18 5/15
The percentages of sampled shrimp with positive in Detection of WSBV by in situ hybridization situ hybridization a r e shown in Table 1. At 16 h pi,
33 % of the shrimp showed a blue to purple precipitate The degree of WSBV infection in the tissues was in their tissues. At 22 h pi, 66 % of the shrimp produced rated by the number of cells with blue-purple precipi- a positive signal and by 40 h pi all the sampled shrimps tate, the intensity of coloration and the integrity of cells were found to be WSBV-positive by the in situ hybrid- on a scale of
+ to
++++.
For those tissues that were intact ization method. These percentages were similar to and stained a light blue to purple in the nuclei, the those of shrimp found positive by PCR. The white spot rating of+ was used. This corresponds to an early stage
syndrome itself was first seen in the shrimp sampled at of infection. Intact tissues that were stained a bright 40 h pi. The size and number of white spots increased blue and contained hypertrophied nuclei were given ratings of ++. Tissues in which the Table l . Detection of white spot syndrome associated non-occluded baculovirus membrane had disintegrated in tissues of exper~mentally infected Penaeus monodon by in sit11 hybridization and blue to purple coloration extended pi: post-lnfectlon to the cytoplasm were rated+++. The
tissues that were stained bright purple"Percentage of positive shrimps in each sampled detected by In situ hybndization
'Relative amount and size of white spots on the carapace: (-) no white spot detected, (+) few and small, (++) many and small, (+++) many and large CMortal~ty of remaining shrimps at each sampllng
0 h pi 16 h p1 22 h pi 40 h pi 52 h p] 64 h p1
Table 2 . Chronological appearance and staining intensity of white spot syn- drome associated non-occluded baculovirus by in situ hybridization in tissues of experimentally infected Penaeus monodon. pi: post-infection. Relative inten- sity of positive staining and integrity of cell: (+) light blue, intact cell with normal nuclei; (++) bright blue, intact cell with hypertrophied nuclei; (+++) blue to pur- ple, cell with disintegrated n~lclear membrane, (++++) bright purple, cell lysed;
(-) no colored precipitate detected
and showed obvlous cytopathological
Tissue O h p i 1 6 h p i 2 2 h p i 4 0 h p i 5 2 h p i 6 4 h p i Stomach -
+
+++++
++++ ++++ Gill - +++
++
+++++++
Cuticular epidermis -+
+++
+++
++++++++
Hepatopancreas - ++
+
++++
Heart -+
++
++
+++
changes were given a rating of
++++.
Intact tissues in which no pigmentation was detected were given a rating of -. The sequential progression of the virus to the organs is shown in Table 2 . The light blue to purple precipitate was present in the nuclei of the columnar cuticular epithelia1 cells and spongy connective tissue of the stom- ach demonstrated at 16 h pi (Fig. 1A). These tissues were still intact and most of the cells did not show any cyto- pathological changes. The percentage of positive cells was not more than 10% at this time. At 22 h pi, the number of positive cells increased, the coloration turned deeper and hyper- trophied cell nuclei began to appear (Fig. 1B). The degree of infection be- came more severe at 40 h pi (Fig. 1C). Even in the sections stained with H&E, it was easy to observe the hypertro- phied nuclei (Fig. 1E). Some cell nucleiLymphojd organ -
-
+
++
were enlarged to more than twice theHematopoietic tissue - -
+
+++++
+++ ++++*+*+
I
diameter of a normal nucleus. AfterAntenna1 gland -
-
+ ++ +++++++
Mid- and h ~ n d g u t -
-
+
+
t + +++Compound eye -
-
-+
++
++
Nervous system - - -
+
+++ +
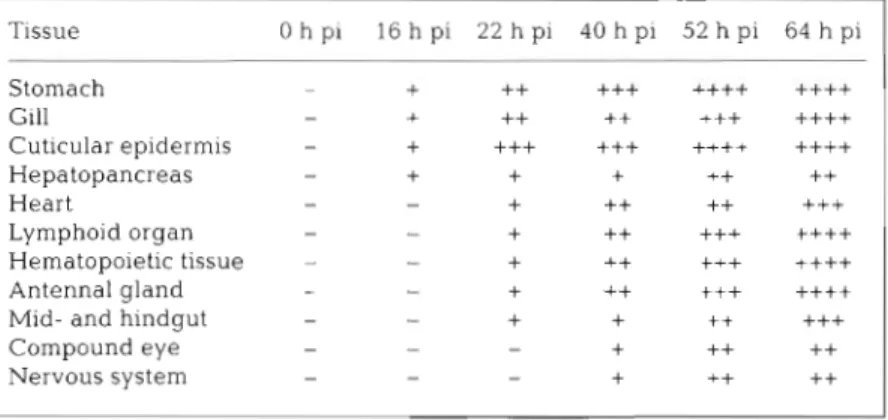
52 h pi, the epithelium layer and connective tissue of stomach were strongly positive and many cells were lysed (Fig. I D ) .
134 Dis Aquat Org 27 131-139, 1996 ' , :?.,
'
,
.
F*W.TiL
. v . . .&-+9 - - - . p
..
-
-,
.
*
-
.
,,
*..
.:;
.(
.
..R.'
.
-f. . . c.
-.
+ b . . ..
*
.
-
3 ' ~ , : is: . r1 ' .
-
.
,
P'.
**' .'Sc
.-a
.
.
.- i .' 4'
,-, \-.- +.- , I, .
-
-:.
C
. -
:-
.
.
C r . . W - *In the gill, viral D N A was first seen at 16 h pi in the epithelium of the primary and secondary filaments The number of positive cells was small and the degree of infection was low initially (Fig. 2A). The color of the precipitate became d e e p blue and cytopathological changes began to occur a t 52 h pi (Fig 2B). At 64 h pi, more than half of the epithelial cells were virus- positive, contamed hypertrophied nuclei, and had be- come degenerate. The organ structure wa.s also disinte- grated a s the area of the lacunaes were increased (Fig. 2C). Cells with hvpertrophied nuclei were observed in the sections stained with H&E at this time (Fig. 2D).
The cuticular epidermis (including the epithelium of the eye stal.k, appendages and the epithe11.um under the carapace) were also first observed to have purple precipitate at 16 h pi. These epithelial cells became severely degenerative and lysed after 52 h pi (Fig. 3 ) .
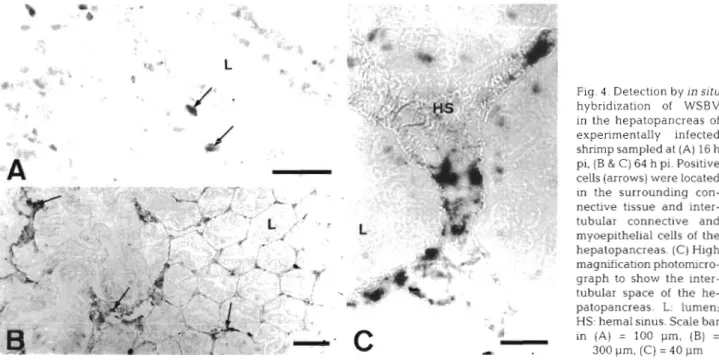
In the hepatopancreas, the connective tissues and myoepithelial cells of the hepatopancreas sheath and in the intertubular spaces were positive at 3.6 h p1 but the epithelium of the tubules was not (Fig. 4A). Cyto-
- ?- " L . *l A
P
'
.
A :.
F,'
Fig. 1 Detection by in sltu hy- bridization of white spot syn- drome associated non-occluded baculov~rus (WSBV) in the stom- ach of experimentally infected shrimp sampled at different times: (A) 16 h post-infection (pi), (B1 22 h pi, (('I 40 h pi, (D) 52 h pi. Pos~tive c ~ . l l s (arrows) were located in the cuticular epithe- lium (CE) and spongy connectlve tissue (SC) but not in the smooth musclc laycr (SM). Scale bar = 100 pm. (E) Severely dffected epithelium in the stomach (H&E staln; 40 h PI) The nuclei of the CE crlls and the SC are hyper- trophied (arrows). Scale bar =
50 p m
pathological changes in the hepatopancreas occurred at 40 h pi. After 52 h pi, most of the cells in the intertubular region were strongly posi.ti.ve but the tubular epithelium was still almost completely nega- tive (Fig. 4B, C ) .
The virus was first seen in the heart, muscle tissue, lymphoid organ, hematopoietic tissue, antenna.1 gland, midyut and hindgut at 22 h pi. In the heart, positive cells were located in muscle fibers and connectlve tissue. After 64 h pi, about 40% of the muscle fibers and connective tissue yielded a deep blue precipitate a n d contained hypertrophied nuclei (Fig. 5A) In the muscle tiss'ue, purple precipitate was present in the nuclei of the muscle f ~ b e r s . At 64 h pi, the nuclel became hypertrophied and stained deep purple but the muscle fibers were still intact (Fig. 5B). In the lymphoid organ, many stromal matrix cells surround- ing thc! lumen and the fibrous connective tissue were positive [Fig. 6 A ) . These cells became lysed and the sheaths had degenerated by 64 h pi. Thc stem cells of the hematopoietic tissues were virus-posltive (Fig. 6B).
Chang et al.: WSBV target organs In shnmp 135
Fig. 2. Detection by in situ
hybridization of W S B V in
the gill of experimentally
infected shrimp sampled at different times: (A) 16 h pi,
(B) 52 h pi, (C) 64 h pi. Posi-
tive cells (arrows) were lo- cated in the epithelium of primary and secondary fila- ments. LA: lacunae (arrow
head). Scale bar = 100 pm.
(D) Severely affected epithe-
lium in the gill (H&E stain;
64 h pi). The nuclei of the
epithelia1 cells (arrows) are
hypertrophied. Scale bar =
50 pm
In the late infection stage, hematopoietic tissue was The fasciculated zone and lamina ganglionaris of the
severely necrotic because most cells were lysed. In the compo.und eyes and the glial cells of the ganglia were antenna1 gland, the positive cells were located in the initially viral DNA positive at 40 h pi. After 64 h pi, epithelium surrounding the gland tubules. The struc- these tissues still showed only medium levels of infec- ture of the tubules disintegrated during the late infec- tion (Fig. 7A, B).
tion stage (Fig. 6C). Unlike the stomach, the positive
cells were located in the muscle and underlying con-
nective tissue region but not in the epithelium of the DISCUSSION
rnidgut and hindgut. However, it is hard to determine
whether the positive signal was in the muscle cells or The results demonstrated that a 1461 bp DNA frag- in the connective tissue or both. At 64 h pi, the nuclei of ment from the genornic library of PmNOB 111 can be
positive cells became hypertrophied and stained deep used as a specific probe to detect WSBV in paraffin-
purple (Fig. 6D). embedded, fixed tissues of infected shrimp by in situ hybridization. The absence of any precipitate in tissues not infected by WSBV suggests that this probe does not
. L+&-
--
- cross-react with shrimp tissue. Recently, in situ hybrid-ization techniques have been developed for some
W
-
shrimp virus diagnosis, such as infectious hypodermaland hematopoietic necrosis virus (IHHNV; Mari et
a 1 1993). Penaeus nionodon-type baculovirus (MBV;
Poulos et al. 1994), Baculovil-us penaei (BP; Bruce et
al. 1993) and hepatopancreatic parvo-like virus (HPV;
Man et al. 1995). The occluded baculovirus can be detected in the tissues of infected shrimp before the occlusion bodies occur by in situ hybridization (Bruce Fig. 3. Detection by in situ hybridization of WSBV in the
pleopod of experimentally infected shrimp sampled at 52 h pi. et l g g 4 ) . It is advantageous use jn ' j t u ~ o s t of the epithelia] cells were strongly positive and had hybridization to detect viral DNA in shrimp tissue than
136 Dis Aquat Org 27: 131-139, 1996
PCR. Although the electron microscopy technique can
definitely provide the information about the sites of viral distribution and the state of viral maturation a t the cellular level, much time is required to obtain the results and expensive equipment is also required. In
s i t u hybridization can accurately provide the precise location of viral DNA present in tissue sections as a result of the highly specific interaction between the probe a n d the target sequence of viral DNA. This property makes in situ hybridization superior to PCR, which cannot provide information about the target cell type within a given organ or tissue. Although WSBV does not form occlusion bodies, it can be easily de- tected by histological staining in late stages of infec- tion. However, this may be more difficult in the lightly infected stage when cytopathological phenomena are still not obvious. In situ hybridization can overcome this problem.
Fig. 4 . Detection by In situ hybridization of WSBV in the hepatopancreas of experimentally infected shrimp sampled at (A) 16 h pi, (B & C) 64 h pi. Positive cells (arrows) were located in the surrounding con- nective tissue and inter- tubular connective and myoepithelial cells of the hepatopancreas. (C) High magnification photomicro- graph to show the inter- tubular space of the he- patopancreas. L: lumen; HS: hemal sinus. Scale bar in (A) = 100 pm, (B) = 300 pm, (C) = 40 p m
The results of this study suggest that the cuticular epithelium from every part of the body of the shrimp is one of the main target tissues for the WSBV. The con- nective tissues of some organs are also infected by WSBV In other tissues such a s nervous tissue, muscle tissue, lyrnphoid tissue and hematopoietic tissue, WSBV DNA can also be observed. In terms of the organs, WSBV does severe damage to the stomach, gills, hematopoietic tissue, lymphoid organ, antenna1 gland and cuticular epidermis of the shrimp. During the late stages of infection, these organs were de- stroyed and many cells were lysed. The organs which were more lightly infected by WSBV included the hepatopancreas, nerve node, compound eye, the muscle and connective tissue of midgut and hindgut. The degree of infection of these organs did not increase and they maintained organ integrity up to the late infection stage, although a few cells had cyto-
Fig. 5. Detection by in situ
hybridization of WSBV in (A) the heart and (B) the muscle tissue of experi- mentally infected shrimp at 64 h pi Nuclei of muscle cells show a positive signal. HE: heart; HP: hepatopan- creas. Scale bar in (A) =
Chang et al.: WSBV target organs in shrimp 137
Fig. 6. Detection by in sjtu hybrid- lzation of WSBV in (A) the lymphoid organ, (B) the hematopoietic tissue, (C) the antenna1 gland, and (D) midgut of experimentally infected shrimp sampled at (A, B, C) 52 h pi and at (D) 64 h pi. Positive cells indi- cated by arrows. FC: fibrous connec- tive tissue; E: epithelium; L: antennel gland lumen: LM: midgut lumen; MS: muscle; S: stromal matrix cell. Scale bar in (A) & (B) = 50 pm, in
(C) = 25 pm, in (D) = l00 pm
Fig. 7. Detection by in situ
hybridization of WSBV in nervous tissues of experi- mentally infected shrimp sampled at 64 h pi. (A) compound eye. The posi- tive cells (arrows) were located in the fasciculated zone (F) and lamina gan- glionaris (LG). CR: crys- talline tract. Scale bar = 100 p m . (B) Ganglion. The glial cells of ganglion were positlve (arrows). Scale
138 DIS Aquat Org 25
pathological signs and were lysed. Thus, it can be concluded that WSBV can infect shrimp t ~ s s u e s that originate from the ectoderm or the mesoderm. This distinguishes WSBV from other shrimp baculoviruses such a s MBV, BP and baculoviral midgut gland necro- sls virus (BMNV) which only infect the epithelia1 cells of the hepatopancreas tubules and the midgut in shrimp (Momoyama 1983, Chang et al. 1992, 1993, Bruce et al. 1993, 1994).
According to the results of this study, although WSBV can also infect the hepatopancreas, it is mainly confined to the myoepithelial cells of the hepatopan- creatic sheath and the fibroblasts of the connective tissue; there was little evidence, if any, that it infected the epithelium of the tubules. Some parts of the hepa- topancreas were lysed in the late infection stage but no viruses were detected. This lysls was evidently not due directly to virus infection, but was probably due to the generally unhealthy state of the infected shrimp a n d resulted from autolysis. As for the enteric tissue, the time at which midgut and hindgut were first detected to be infected with WSBV was later than for the stom- ach and the degree of infection was also lighter than the degree of infection in the stomach.
The results also show that the virus initially appears in the shrimp in the stomach, gill, cuticular epidermis and connective tissue of the hepatopancreas. Chou et al. (1.995) indicated that under experimental conditions the virus can infect the shrimp via water and oral ino- culation. From this study, some shrimp sampled a t 16 h showed virus in the stomach but not in the gills, while others sampled at the same time showed virus in the gills but not in the stomach (data not shown). T h ~ s sug- gested that the virus infection could be either via the oral pathway or via water to the gill or cuticular epider- mis. Since virus-infected shrimp tissues were given to the experimental shrimp as feed, both oral and water- borne infections were possible. In some sections, a few pos~tive signals could be detected in the lumen of the stomach at the immediate onset of infection. Specifi- cally, these signals were in the cuticle layer above the epithelium. This finding can be explained if the virus- containing feed had just entered the stomach lumen and some of it had adhered to the setal grooves of the cuticle layer. It may also result from a non-specific bind- ing to chitin that has been noted by Lightner (1996).
WSBV can also infect the heart, eyes and nervous tissue of shrimp The most severely infected site of the eyes was the cuticular epidermis of the eye stalk. Some nerve cells of the compound eyes and glial cells of the ganglia were also lightly infected by WSBV in the late stage. Changes found in WSBV-infected shrimps can be distinguished easily from the white eye syndrome in Tai- wan In 1991 in terms of brown fibrous nodules in the fas- ciculated zone of the compound eyes (Chen et a1 1991).
In conclusion, a sensitive, specific and nonradio- active nucle~c a c ~ d probe can be used for the in situ
detection of the WSBV genome. Viral infection was detected at a n early stage, the degree of infection was determined, and target tissues were identified. The route of WSBV entry and progression of the disease caused by WSBV was also established.
Acknowledgements. This work was supported by the National Science Council under grant NSC 85-2321-B-022- 001 and the Council of Agriculture under grant No. 85-AST- 1 1-FAD-49(21)A.
LITERATURE CITED
Bruce LD, Lightner DV, Redman RM (1994) Comparison of traditional and molecular detection methods for baculo- virus penaei ~nfections in larval Penaeus vannamei. J Aquat Anim Health 6:355-359
Bruce LD, Redman RM, Lightner DV, Bonami JR (1993) Appli- cation of gene probes to detect a penaeid shrimp baculo- virus in fixed tissue using in situ hybridization. Dis Aquat Org 17:215-221
Cai S, Huan.9 J , Wang C, Song X , Sun X, Yu J , Zhang Y, Yang C (1995) Epidemiological studies on the explosive epi- demic disease of prawn in 1993-1994. J Fish China 19: 112-117
Chang PS, Lo CF. Kou GH, Lu CC, Chen SN (1993) Purifica- tion and amplification of DNA from Penaeus monod0.n- type baculovirus (MBV). J Invertebr Pathol 62.116-120 Chang PS, Wang YC, Lo CF, Kou GH, Chen SN (1992) Pu.rifi-
cation and biochemical characteristics of occlusion body of Penaeus monodon-type baculovirus (MBV). Fish Pathol 27:127-130
Chen SN, Chang PS, Kou GS (1991) Surveys on the diseases of cultured Penaeus japonicus in Taiwan. Environmental survey and its Improvement on shrimp grow-out pond. COA (Counc Agric) Fish Ser 28.123-132
Chou HY, Huang CY, Wang CH. Chiang HC, Lo CF (1995) Pathogenicity of a baculovirus infection causing white spot syndrome in cultured penaeid shrimp in Taiwan. Dis Aquat Org 23:165-173
Inouye K , Miwa S, Oseko N, Nakano H, Kirnura T (1994) Mass mortalities of cultured kuruma shnmp, Penaeusjaponicus, in Japan in 1993: electron m~croscopic evidence of the causative virus. Fish Pathol 29:149-158 ( ~ n Japanese) Lightner (1996) A handbook of pathology and diagnostic
procedures for dlseases of penaeid shrimp. World Aqua- culture Society, Baton Rouge, LA
Lo CF, Leu J H , Ho CH, Chen CH, Peng SE, Chen YT, Chou CM, Yeh PY, Huang CJ, Chou HY, Wang Cfl, Kou GH (1996) Detection of baculovirus associated with white spot syndrome (\YSBV) in penaeid shrimps using polymerase chain reaction Dis Aquat Orq 25:133-141
Man J , Bonami JR, Lightner DV (1993) Partial cloning of the genome of infectious hypodermal and hematopoietic necrosis virus, a n unusual parvovirus pathogenic for penaeid shrimps; diagnosis of the disease using a specific probe. J Gen Virol74:2637-2643
Mari J , Lightner DV, Poulos BT, Bomani JR (1995) Partial cloning of the genome of an unusual shrimp parvovirus (HPV): use of gene probes in disease diagnosis. Dis Aquat Org 22:129-134
Chang et al.: WSBV target organs in shrlmp
Momoyama K (1983) Studies on baculoviral mid-gut gland
necrosis of kuruma s h r l n ~ p (Penaeus japonlcus) 111. Fish
Path01 17 263-268
Momoyarna K. Hiraoka M, Nakano H, Koube H, lnouye K,
Oseka N (19941 Mass inortdllties of cultured kuruma
shrimp, Penaeus japonicus, in Japan in 1993: histopatho- logical study. Fish Pathol 29:141-148 (in Japanese)
Nakano H. Koube H, Umezaea S, Momoyama K. Hiraoka M.
Inouye K , Oseko N (1994) Mass mortalities of cultured
kurulna shrimp, Penaeirs]aponicus, In Japan In 1993: epi-
zoot~olog~cal survey a n d infection trlals. Fish Pathol 29:
135-139 (in Japanese)
Nuovo GJ. Darfler MM. Imparaim CC, Bromley SE (1991)
Occurrence of multiple types of human papillomdvirus
in genital tract lesions: analysis by in situ hybridization and the polymerase chain reaction. Am J Pathol 58,
51 8-523
Poulos BT, Mari J , Bonaini JR, Redman R, Lightner D V (1994)
Use of non-radioactively labeled DNA probes for the
Responsible Sublect Editor; J. E. Stewart, Dartmouth, Nova
Scotia, Canada
detection of a baculovirus from Penaeus monodon
[PmSNPV = MBV) by In sltu hybrid~zatlon on fixed tissue.
J Virol Meth 49:187-194
Takahashi Y, ltami T, Kondom M, .Maeda M , Fujii R, Tonlo-
naga S, Supamatlaya K, Boonyaratpalln S (1994) Elec-
tron microscopic evidence of barilliforrn virus infection
in Kuruma shrimp (Penaeus japonicus). Fish Pathol 29:
121-125
Wany C H , Lo CF, Leu J H , Chou CM, Yeh PV, Chou HY, Tung MC, Chang CF, Su I\4S, Kou G11 (1995) Purification and
yenomic dnalysis of baculovirus associated with w h ~ t e
spot syndrome (WSBV) of Pendeus monodon. Dis Aquat O r g 23:239-242
Wongteerasupaya C, Vickers JE, Sriurairatana S, Nash GL,
Akarajamorn A, Boonsaeng V, Panyim S, Tassanakajon A, Withyachumnarnkul B, Flegel TW (1995) A non-occluded,
systemic baculovlrus that occurs in cells of ectodermal and mesodermal origin and causes high mortality in the black tiger prawn Penaeus monodon. Dis Aquat O r g 21:69-77
A4anuscript first received: J a n u a r y 24, 1996 Revised version accepted: M a y 2, 1996