ORIGINAL ARTICLE
Construction and expression of rabbit neutrophil peptide-1
gene in
Escherichia coli
Yu Ling Sun&Tang Ching Kuan&Yi Juain Lin& Yu Chi Chou&Chih Sheng Lin
Received: 4 February 2010 / Accepted: 22 March 2010 / Published online: 16 April 2010 # Springer-Verlag and the University of Milan 2010
Abstract Rabbit neutrophil peptide-1 (NP-1) is a proto-typic rabbit α-defensin with a broad antimicrobial spec-trum. The coding sequence of NP-1 was amplified and cloned into pET-31b(+) to construct an expression vector, pET31-NP1, which was transformed into E. coli BLR(DE3) pLysS for expressing the fusion NP-1 protein (fNP-1). The fNP-1 is downstream of a ketosteroid isomerase (KSI) and upstream of a (His)6-Tag, as KSI-NP1-His6. The optimal
condition, cultivation in enriched LB medium and induc-tion with 0.5 mM IPTG for 6 h, was determined for fNP-1 production. The fNP-1 was purified by Ni-NTA resin and cleaved by cyanogen bromide to release matured NP-1 peptide. The matured NP-1 peptide showed significant antimicrobial activities against clinical bacteria, Escherichia coli, Pseudomonas aeruginosa, Bacillus subtilis, and Staphylococcus aureus. The application of this expression approach represents a potential method to produce NP-1 by fusion protein expressed in E. coli without cytotoxicity. Keywords Antimicrobial activity . Fusion protein . Rabbit neutrophil peptide-1
Introduction
Defensins are endogenous, cysteine-rich, 3- to 4-kDa cationic peptides that exert antimicrobial activity through membrane permeabilization (Wimley et al. 1994; White et al. 1995; Gudmundsson and Agerberth 1999). Three subfamilies of defensins, including α-, β- and θ-defensin, exist among mammals (Ganz 2003). In humans, α-defensins are found in neutrophils (Lehrer et al. 1993), T and B lymphocytes (Agerberth et al.2000). In rabbits, but not in humans, α-defensins are also prominent in pulmo-nary alveolar macrophages (Ganz et al. 1985). Rabbit neutrophil peptide-1 (NP-1), an Arginine-rich and cationic peptide with 33 amino acids (Fuse et al. 1993), is a prototypic rabbit α-defensin that is abundant in rabbit granulocytes and rabbit alveolar macrophages (Patterson-Delafield et al. 1981; Selsted et al.1983), and it was first called MCP-1 (macrophage cationic peptide-1). Later sequencing proved that NP-1 from rabbit polymorphonu-clear leukocytes (Selsted et al. 1985) and MCP-1 were identical. Rabbit NP-1 exhibits growth inhibition activity in vitro against a broad spectrum of pathogenic microisms including Gram-positive and Gram-negative organ-isms, as well as some virus pathogens (Sinha et al.2003).
In order to investigate the possibility of pharmaceutical application of rabbit NP-1, a relatively large quantity of NP-1 needs to be produced economically. Recombinant technique has emerged as attractive cost-effective method and its application is expected to be widespread. In this study, we have been able to overexpress NP-1 fused to ketosteroid isomerase (KSI) by the plasmid pET-31b(+) in E. coli BLR(DE3)pLysS. The fusion strategy was employed in expression vector construction to avoid the toxic effect on host cell and produce the activated NP-1 after removing the fused proteins.
Y. L. Sun
:
Y. J. Lin:
Y. C. ChouDivision of Biotechnology, Animal Technology Institute Taiwan, P.O. Box 23, Chunan,
35099 Miaoli, Taiwan T. C. Kuan
:
C. S. Lin (*)Department of Biological Science and Technology, National Chiao Tung University,
No.75 Po-Ai Street, Hsinchu 30068, Taiwan e-mail: [email protected] DOI 10.1007/s13213-010-0046-z
Materials and methods Strains, vectors and reagents
Escherichia coli strain JM109 was used as the host for cloning and the propagation of plasmids. E. coli BLR(DE3) pLysS (Novagen, Madison, WI, USA) was used as the expression host. Plasmid pCR2.1-TOPO (Invitrogen, Carls-bad, CA, USA) was used for subcloning of the PCR product of NP-1 gene, and pET-31b(+) (Novagen) was chosen for fusion protein expression. All the restriction enzymes and T4 DNA ligase (Promega, Madison, WI, USA) were used for the construction of recombinant vector. All chemical reagents were purchased from Sigma (St. Louis, MO, USA) or Merck (Darmstadt, Germany). The recombinant DNA techniques were exploited as described by Sambrook et al. (1989).
Synthesis and construction of a DNA fragment coding for NP-1
For the construction of a vector comprising a NP-1 gene encoding mature NP-1 peptides, the cDNA was prepared from white blood cells of rabbit as template DNA. A primer pair (NP1-F and NP1-R) were synthesized according to the sequence registered in GenBank (Accession number: M28883) and used to amplify the NP-1 gene fragment carrying restriction enzyme AlwNI recognized sites at its 5′ and 3′ termini. The amplified PCR DNA fragment was cloned to pCR2.1-TOPO, resulting in the vector, named pNP1-TOPO.
(1) NP1-F: 5′-CAG ATG CTG GTG GTC TGT GCG
TGC-3′ (underlined sequences indicate the AlwNI recognized site)
(2) NP1-R: 5′-CAG CAT CTG GCG GCG GCA GCA
CA-3′ (underlined sequences indicate the AlwNI recognized site)
Construction of NP-1 expression vector
The amplified NP-1 gene fragment was recovered from the pNP1-TOPO digested with AlwNI. Vector DNA, pET-31b (+), was also cleaved by the same restriction enzyme and the larger fragment was recovered. The recovered DNA fragments were ligated to construct the expression vector, pET31b-NP1. The complete sequence of the inserted NP-1 gene was confirmed by direct sequencing of the vector DNA.
Expression of the fusion protein
For expressing the fusion NP-1 protein (fNP-1), the expression vector pET31b-NP1 carrying the desired mature
NP-1 gene were purified and transformed into E. coli BLR (DE3)pLysS. A fresh bacterial clone of E. coli BLR(DE3) pLysS harboring pET31b-NP1, named BLR(DE3)/pET31b-NP1, was grown in enriched Luria-Bertani (LB) medium (1% Bacto-tryptone, 2% yeast extract and 0.5% NaCl, pH 7.2) supplemented with 100μg ml−1ampicillin, 34μg ml−1 chloramphenicol and 0.2% (w/v) glucose at 37°C. When the cells were cultured to OD600 at about 0.5, they were
inoculated into 600 ml fresh enriched LB medium with the volume ratio of 5% and the fusion protein expression was induced by the addition of isopropyl-β-D-l-thiogalatopyr-anoside (IPTG).
Purification and cleavage of the fusion protein
After additional 6 h culture by the induction of IPTG, the cultured cells of BLR(DE3)/pET31b-NP1 were harvested by centrifugation at 8,000g for 20 min, then resuspended in 20 ml cooled of binding buffer (20 mM Tris-HCl, 0.5 M NaCl and 5 mM imidazole, pH 7.9) and disrupted by sonication. After centrifugation at 12,000g for 20 min, the insoluble pellet (inclusion body) was dissolved in 10 ml of binding buffer containing 6 M guanidine-HCl. Following the centrifugation to remove residual matter, the fusion protein containing (His)6-Tag was purified using the
pre-charged nickel-nitrilotriacetic acid (Ni-NTA) agarose affin-ity chromatography (Invitrogen). The fusion protein bound to the resin was washed by washing buffer (20 mM Tris-HCl, 6 M guanidine-Tris-HCl, 0.5 M NaCl and 40 mM imidazole, pH 7.9), and then eluted with elution buffer (20 mM Tris-HCl, 6 M guanidine-HCl, 0.5 M NaCl and 1 M imidazole, pH 7.9). The elution of fusion protein was collected and dialyzed at 4°C against deionized water for 24 h. The purified fusion protein was analyzed by 15% SDS-polyacrylamide gel electrophoresis and western blot-ting. The concentration of protein was analyzed by BCA Protein Assay Kit (Pierce, Rockford, IL, USA).
The purified fusion protein was lyophilized and treated with 1 mg/ml CNBr (Sigma) fusion protein (in 70% formic acid) for 24 h at 25°C to release mature NP-1 peptide. The cleaved fusion protein was dissolved in 30 ml deionized water and rotated by rotary evaporator to remove the remaining CNBr. The released mature NP-1 peptide was analyzed by 4–NP-12% NuPAGE gel (Invitrogen) and quantitated.
Western blotting analysis
Protein sample was resolved by electrophoresis and electro-transferred onto PVDF membrane (Amersham Biosciences, Piscataway, NJ, USA). The membrane was blocked with 5% (w/v) non-fat milk in PBS and incubated with rat anti-NP1 antisera in 1:1,000 dilution for 2 h. Following washing
with PBS containing 0.1% Tween 20, the blot was reacted with alkaline phosphate-conjugated anti-rat IgG (diluted 1:2,500). The protein bound enzymes on the membrane were visualized with BCIP/NBT Substrate Solution (PerkinElmer, Boston, MA, USA).
Assay of antimicrobial activity
Antimicrobial activity of the NP-1 was analyzed using radial diffusion assay as described previously with slight modifica-tions (Lehrer et al.1991). Clinical isolates, E. coli (ATCC 8739), P. aeruginosa (ATCC 10145), B. subtilis (ATCC 6633) and S. aureus (ATCC 6538P), were obtained from the American Type Culture Collection (Manassa, VA, USA) and used in this study. All bacteria were incubated overnight and diluted in 3% trypticase soy broth (TSB), and grown for 2.5– 3.5 h (mid-exponential growth phase; 0.2–0.25 bacterial OD620, around 4–8×10
7
CFU ml−1) at 37°C with shaking. Following centrifugation (6,000 g, for 10 min), the bacterial pellets were washed with 10 mM PBS (pH 7.4) and resuspended in 10 ml of the same buffer. A volume containing approximate 1×106bacterial CFU was added to 10 ml PBS that contained 3 mg of powdered TSB, 1% w/v of agar, and 0.02% Tween 20. Bacteria-containing medium was poured into a 100-mm Petri dish to form a uniform layer 1 mm deep. A gel punch (3 mm inside diameter) was used to make evenly spaced wells. Samples of 5μl of the CNBr-treated NP-1 or the control solvent was added to the wells. The plates were incubated for 3 h at 37°C and then overlaid with 10 ml of sterile agar consisting of 6% w/v solution of TSB and 1% agar. After incubation at 37°C for 10–16 h, the diameters of the clearance zones were measured.
Minimum inhibitory dose (MID), i.e., the minimum dose of NP-1 forming a detectable clear zone larger than the size of the central well (3 mm) under our experimental conditions, was determined from serial dilutions of the CNBr-treated NP-1 by plotting the known doses of a peptide that was added to the well versus the respective clearance zones. The regression equation was then used to determine the MID for each tested microorganism.
Results and discussion
Construction of expression vector
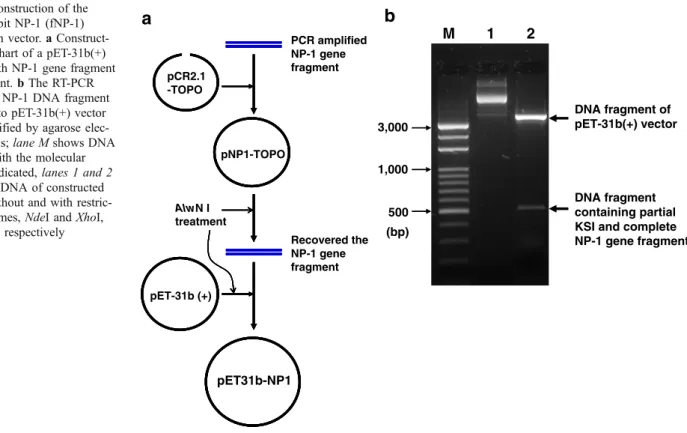
The NP-1 gene suitable for E. coli expression was cloned into pET-31b(+) successfully to construct the expression vector, NP1. The construction strategy of pET31b-NP1 was shown in Fig. 1a and the inserted NP-1 DNA fragment within plasmid pET-31b(+) was verified (Fig.1b). The constructed NP-1 gene was under the control of T7
promoter and fused with a downstream of ketosteroid isomerase (KSI) and an upstream of (His)6-Tag to generate
the fusion NP-1 protein as KSI-NP1-His6(Fig. 2) and the
expressed fNP-1 is 18.8 kDa. There is (His)6-Tag at the
C-terminum of fNP-1, which serves as the later purification of fusion protein. There are also AlwNI recognized sites at the 5’ and 3’termini of NP-1 gene. By the AlwNI, methionine residue could be translated and provided cyanogen bromide (CNBr) cleavage sites for releasing mature NP-1 peptide from the fusion protein.
Expression of fusion NP-1 protein
The constructed expression vector, pET31b-NP1, was transformed into E. coli strain, BLR(DE3)pLysS, to obtain a cloned E. coli strain, named BLR(DE3)/pET31b-NP1, to produce the fusion protein, fNP-1. The E. coli BLR(DE3)/ pET31b-NP1 was seeded and cultured in enriched LB medium. After 1 mM IPTG induction and SDS-PAGE analyses, there was an obvious foreign protein band with a molecular weight of 18.8 kDa, which is consistent with the predicted fusion protein, fNP-1 (KSI-NP1-His6) (Fig. 3a).
The fusion protein could be purified by Ni-NTA agarose resin and further confirmed by reacting with rat anti-NP1 antisera (Fig. 3b). Introduction of artificial fusion protein, such as KSI fusion partner (enhancing the formation of inclusion bodies), at the N- or C-terminus of fusion protein, is a popular way to facilitate purification of recombinantly produced proteins (Cipáková et al.2006).
Induction effect of IPTG
For the expression vector, pET31b-NP1, the foreign protein expression was triggered by adding IPTG into the culture medium. Final IPTG concentration and induction time should be optimized because of its great contribution to fusion protein expression (Xu et al. 2006). In this work, IPTG concentration was examined from 0.3 to 1 mM and the expression of fNP-1 was analyzed in Fig. 4a. At 0.5 mM, the fNP-1 expression was the highest; therefore, further studies were all carried out under this condition.
In the process of fusion protein production in E. coli BLR(DE3)/pET31b-NP1, IPTG induction is the turning point between cell growth and fusion protein synthesis. Therefore, the effect of induction timing was evaluated by adding IPTG at different stages of growth phase. The production of fNP-1 was compared as indicated in Fig.4b. With different induction timing, the concentration of protein varied in a wide range from 0.14 to 1.31 g l−1, with the highest value observed when induced at 6 h, i.e., at the middle stage in exponential growth of cloned E. coli. Hence, induction at the middle stage of exponential growth was adopted.
Antimicrobial activity of the fusion protein
The E. coli BLR(DE3)/pET31b-NP1 was cultured and induced by 0.5 mM IPTG for 6 h. The fNP-1 accounted for around 35% of total protein. Using KSI as a fusion partner, the fusion protein was expressed as insoluble protein and accumulated in inclusion bodies in E. coli (Park et al.2009). Therefore, the inclusion bodies of E. coli cells were harvested for the purification of fNP-1. The fNP-1 was purified with Ni-NTA Resin. The purities of eluted fNP-1 were about 80%. Typically, the yield of fusion protein was 10–20 mg from 10 g of wet pelleted cells. After dialysis against deionized H2O, the
purified KSI-NP1-His6fusion protein was cleaved with CNBr
and the mature NP-1 was released.
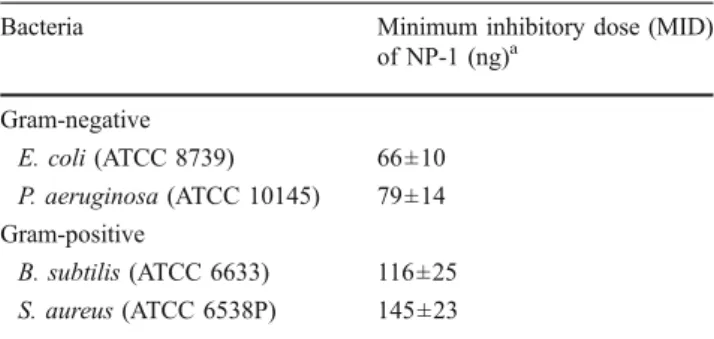
To show that the expression approach used retaining the antimicrobial activity of the prepared NP-1, a radial diffusion assay was performed. The elution samples were administered to a panel of bacterial organisms. As shown in Table 1, the purified NP-1 provided high, dose-dependent antimicrobial effect against Gram-negative bacteria (E. coli and P. aeruginosa) and Gram-positive bacteria (B. subtilis and S. aureus). An antimicrobial peptide, rabbit NP-1, was successfully expressed in bacteria E. coli as a fusion protein in the present study. However, further work should be done AlwN I treatment pET-31b (+) PCR amplified NP-1 gene fragment pNP1-TOPO pCR2.1 -TOPO pET31b-NP1 Recovered the NP-1 gene fragment
a
b
M 1 2 500 (bp) 1,000 3,000 DNA fragment containing partial KSI and complete NP-1 gene fragment DNA fragment of pET-31b(+) vector Fig. 1 Construction of thefused rabbit NP-1 (fNP-1) expression vector. a Construct-ing flowchart of a pET-31b(+) vector with NP-1 gene fragment was present. b The RT-PCR amplified NP-1 DNA fragment cloned into pET-31b(+) vector was identified by agarose elec-trophoresis; lane M shows DNA marker with the molecular weight indicated, lanes 1 and 2 show the DNA of constructed vector without and with restric-tion enzymes, NdeI and XhoI, treatment, respectively
(His(6)-Tag)
KSI (125 aa) NP1 (99 nt; 33 aa)
(His(6)-Tag) (AlwN I)
(AlwN I)
Fig. 2 Map and sequences of the fusion rabbit NP-1 (fNP-1) within pET31b-NP1 vector. In the expression vector of pET31b-NP1, NP-1 coding sequence was placed downstream of ketosteroid isomerase (KSI) gene and upstream of (His)6-Tag sequence, KSI-NP1-His6. Use
of the AlwNI sites enabled unidirectional insertion of coding sequence
adjacent to a methionine residue and provided the cyanogen bromide (CNBr) cleavage site for releasing mature NP-1 peptide from the fusion protein. The fNP-1 gene was under the expressing regulation of T7 promoter. aa Amino acid residues, nt nucleotides
to elevate the expression level of the NP-1 and to establish an effective purification process. Moreover, it is also worth testing the capacity of produced antimicrobial peptides against to drug-resistant strains (Cipáková et al.2006).
Conclusion
In summary, this work presents the expression method that can be applied for the production of rabbit NP-1 in bacteria.
The above results showed that KSI-NP1-His6fusion protein
expression system was able to circumvent the toxicity of NP-1 on the host bacterial cells. Since produced mature NP-1 exhibited strong antimicrobial activity against tested microorganisms, we could conclude that this expression approach of producing fusion NP-1 provides a powerful tool for production of sufficient quantities of biologically active NP-1 for its further biological study.
Acknowledgements The authors gratefully acknowledge Dr. Shih-Rong Wang at the Animal Technology Institute Taiwan for the gift of rat anti-NP1 antiserum. This work was supported by the grant of 96-EC-17-A-17-R7-0454 from the Ministry of Economic Affairs of Executive Yuan, Taiwan.
References
Agerberth B, Charo J, Werr J, Olsson B, Idali F, Lindbom L, Kiessling R, Jörnvall H, Wigzell H, Gudmundsson GH (2000) The human antimicrobial and chemotactic peptides LL-37 and alpha-defensins are expressed by specific lymphocyte and monocyte populations. Blood 96:3086–3093
Cipáková I, Gasperík J, Hostinová E (2006) Expression and purification of human antimicrobial peptide, dermcidin, in Escherichia coli. Protein Expr Purif 45:269–274
Fuse N, Hayashi Y, Fukata J, Tominaga T, Ebisui O, Satoh Y, Isohara T, Uno I, Imura H (1993) Purification and characterization of new anti-adrenocorticotropin rabbit neutrophil peptides (defen-sins). Eur J Biochem 216:653–659
Ganz T (2003) Defensins: antimicrobial peptides of innate immunity. Nat Rev Immunol 3:710–720
Ganz T, Sherman MP, Selsted ME, Lehrer RI (1985) Newborn rabbit alveolar macrophages are deficient in two microbicidal cationic peptides, MCP-1 and MCP-2. Am Rev Respir Dis 132:901–904
Gudmundsson GH, Agerberth B (1999) Neutrophil antibacterial peptides, multifunctional effector molecules in the mammalian immune system. J Immunol Methods 232:45–54
Lehrer RI, Rosenman M, Harwig SS, Jackson R, Eisenhauer P (1991) Ultrasensitive assays for endogenous antimicrobial polypeptides. J Immunol Methods 137:167–173
Lehrer RI, Lichtenstein AK, Ganz T (1993) Defensins: antimicrobial and cytotoxic peptides of mammalian cells. Annu Rev Immunol 11:105–128 M 1 2 3 M 1 2 3 26 17 (kDa) 18.8 KDa fNP-1 (KSI-NP1-His6) 1 2 3
a
b
Fig. 3 Expression and verification of the expressed fNP-1. a SDS-PAGE analysis of the expressed fNP-1 was shown; lane M shows protein marker with the molecular weight indicated, lanes 1 and 2 show the total protein profile from the E. coli BLR(DE3)/pET3b-NP1, which were cultured in the enriched LB medium without and with IPTG induction, respectively. Lane 3 shows the purified fNP1 by Ni-NTA resin. b Western blotting analysis of the expressed fNP-1. The samples were resolved on SDS-PAGE, transformed to PVDF membrane, incubated with rat anti-NP1 antisera, probed with alkaline phosphate-conjugated anti-rat IgG, and then visualized with BCIP/ NBT treatment. The recognized fNP-1 is indicated by the arrow
Table 1 Antibacterial activity of NP-1
Bacteria Minimum inhibitory dose (MID)
of NP-1 (ng)a Gram-negative E. coli (ATCC 8739) 66±10 P. aeruginosa (ATCC 10145) 79±14 Gram-positive B. subtilis (ATCC 6633) 116±25 S. aureus (ATCC 6538P) 145±23
a Experiments were repeated five times and the values presented are
means ± SEM
a
b
1 2 3 4 5 6 7 8 fNP-1 (KSI-NP1-His6) 1 2 3 4 fNP-1 (KSI-NP1-His6)Fig. 4 Effect of IPTG concentration and IPTG inducing time on the fNP-1 expression level of cloned BLR(DE3)/pET31b-NP1. a Lanes 1–4 show the total protein profiles of the cultures treated with 0, 0.3, 0.5, and 1.0 mM of IPTG for 12 h. b Lanes 1–8 show total protein profiles of the cultures at 0, 1, 2, 3, 4, 5, 6, and 12 h after 0.5 mM IPTG induction. The expressed fNP-1 is indicated by the arrows
Park TJ, Kim JS, Choi SS, Kim Y (2009) Cloning, expression, isotope labeling, purification, and characterization of bovine antimicro-bial peptide, lactophoricin in Escherichia coli. Protein Expr Purif 65:23–29
Patterson-Delafield J, Szklarek D, Martine RJ, Lehrer RI (1981) Microbicidal cationic proteins of rabbit alveolar macrophages: amino acid composition and functional attributes. Infect Immun 31:723–731
Sambrook J, Fritsch EF, Maniatis T (1989) Molecular cloning: a laboratory manual, 2nd edn. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY
Selsted ME, Brown DM, DeLang RJ, Lehrer RI (1983) Primary structures of MCP-1 and MCP-2, natural peptide antibiotics of rabbit lung macrophages. J Biol Chem 258:14485–14489
Selsted ME, Brown DM, DeLange RJ, Harwig SS, Lehrer RI (1985) Primary structures of six antimicrobial peptides of rabbit peritoneal neutrophils. J Biol Chem 260:4579–4584
Sinha S, Cheshenko N, Lehrer RI, Herold BC (2003) NP-1, a rabbit alpha-defensin, prevents the entry and intercellular spread of herpes simplex virus type 2. Antimicrob Agents Chemother 47:494–500
White SH, Wimley WC, Selsted ME (1995) Structure, function, and membrane integration of defensins. Curr Opin Struct Biol 5:521–527 Wimley WC, Selsted ME, White SH (1994) Interactions between human defensins and lipid bilayers: evidence for formation of multimeric pores. Protein Sci 3:1362–1373
Xu Z, Zhong Z, Huang L, Peng L, Wang F, Cen P (2006) High-level production of bioactive human beta-defensin-4 in Escherichia coli by soluble fusion expression. Appl Microbiol Biotechnol 72:471–479