INTRODUCTION
Patients with diabetes mellitus (DM) are predisposed toward developing com-plications such as increased rates of skin infections and chronic foot ulcerations that frequently lead to amputation (1–4). The rate of nontraumatic lower extremity amputation for patients with DM is more than 40 times the rate for individuals who do not have DM (1). Immune dys-functions that affect leukocyte adherence, chemotaxis and phagocytosis occur under hyperglycemic conditions and may make DM patients more prone to infections (5–7). We recently demon-strated that the physiologic functions of keratinocytes, the immune-competent
cells that form the outermost layer of the skin, were also affected by high-glucose environments (8,9).
As the major constituents of the skin, keratinocytes generate a number of an-timicrobial peptides that are involved in innate immunity, providing the first de-fense against potential microbial invasion from the environment. The major antimi-crobial peptides found in human beings are defensins and cathelicidins (10). Human β-defensins (hBD) found in the skin were the focus of this study (10–12). Currently, four hBD of different charac-teristics have been identified (13–19). These defensins have demonstrated the capability to kill and/or inactivate a
spec-trum of bacteria, fungi and enveloped viruses in vitro (12). Results of previous studies have shown decreased BD levels in different tissues of diabetic rats (11); however, the possible alterations of hBD expression in the diabetic skin, especially during the wound-healing process, have not been thoroughly investigated.
In terms of antimicrobial activity, hBD1 and hBD2 have shown effects against Gram-negative bacteria. On the other hand, in addition to effects on Gram-negative bacteria, hBD3 has also shown antimicrobial effects against Gram-positive bacteria, including Staphy-lococcus aureus, the most common patho-gen found in diabetic wounds (20). Re-sults of previous studies demonstrated that hBD3 is the key molecule by which keratinocytes kill S. aureus (21). The cur-rent study was launched to investigate the biological and molecular effects of a high-glucose environment on epidermal hBD3 expression.
Reduces Human
β-3 Expression in Keratinocytes
Cheng-Che E. Lan,
1,2Ching-Shuang Wu,
3Shu-Mei Huang,
1,4Hsuan-Yu Kuo,
1I-Hui Wu,
1Chien-Hui Wen,
5Chee-Yin Chai,
5Ai-Hui Fang,
6and Gwo-Shing Chen
11Department of Dermatology, Kaohsiung Medical University Hospital, Department of Dermatology, College of Medicine, and Center
of Excellence of Environmental Medicine, Kaohsiung Medical University, Kaohsiung, Taiwan; 2Department of Dermatology, Kaohsiung Municipal Ta-Tung Hospital, Kaohsiung, Taiwan; 3Department of Medical Laboratory Science and Biotechnology,
Kaohsiung Medical University, Kaohsiung, Taiwan; 4Department of Biological Sciences, National Sun Yat-sen University, Kaohsiung, Taiwan; 5Department of Pathology and 6Department of Microbiology, College of Medicine, Kaohsiung Medical University,
Kaohsiung, Taiwan
Diabetes mellitus is characterized by elevated plasma glucose and increased rates of skin infections. Altered immune re-sponses have been suggested to contribute to this prevalent complication, which involves microbial invasion. In this study we explored the effects of a high-glucose environment on the innate immunity of keratinocytes by focusing on β defensin-3 (BD3) using in vivo and in vitro models. Our results demonstrated that the perilesional skins of diabetic rats failed to show enhanced BD3 expression after wounding. In addition, high-glucose treatment reduced human BD3 (hBD3) expression of cultured human keratinocytes. This pathogenic process involved inhibition of p38MAPK signaling, an event that resulted from increased forma-tion of advanced glycaforma-tion end products. On the other hand, toll-like receptor-2 expression and funcforma-tion of cultured keratino-cytes were not significantly affected by high-glucose treatment. In summary, high-glucose conditions inhibited the BD3 expres-sion of epidermal keratinocytes, which in turn contributed to the frequent occurrences of infection associated with diabetic wounding.
© 2011 The Feinstein Institute for Medical Research, www.feinsteininstitute.org Online address: http://www.molmed.org
doi: 10.2119/molmed.2010.00091
Address correspondence and reprint requests to Gwo-Shing Chen, 100 Shih-Chuan First Road, Kaohsiung, Taiwan. Phone: +8867 3121101 (Ext 6108); Fax: +8867 3216580; E-mail: [email protected]
Submitted June 28, 2010; Accepted for publication March 22, 2011; Epub (www.molmed.org) ahead of print March 22, 2011.
ANIMAL MODEL
STZ-Induced Diabetic Rats
Male Wistar rats (200–300 g; BioLASCO Taiwan, Taipei, Taiwan) were acclimatized for 7 d before the experimental procedures and had free access to standard laboratory chow and water. Diabetes was then in-duced by single 55-mg/kg intraperitoneal injection of streptozotocin (STZ; Sigma, St. Louis, MO, USA). The induction was con-sidered successful if the blood glucose lev-els measured were greater than 350 mg/dL 2 d after STZ injection.
Wound-Healing Model
Samples of rat skin were obtained under the protocols approved by the eth-ics committee of Kaohsiung Medical Uni-versity Hospital. Prior to skin injury, the rats were anesthetized by a single 50-mg/kg intraperitoneal injection of Zoletil (Virbac, Carros, France). After the dorsal hair was shaved and the exposed skin was cleaned with 75% ethanol, thickness wounds were made on the rat dorsum by using a 6-mm biopsy punch. Perilesional biopsies were performed im-mediately and at 1, 3 and 5 d after wounding. The specimens were then di-vided into two sections. The first section was preserved with 4% neutral buffered formalin and kept at 4°C overnight. The skin section was then preserved at –20°C for subsequent immunohistochemical (IHC) studies. For the second section, the dermal component was immediately re-moved. The epidermis was then incu-bated with RNAlater solution (Ambion, Austin, TX, USA) at 4°C for 24 h fol-lowed by preservation at –80°C for sub-sequent quantitative polymerase chain reaction (PCR) analyses.
Immunohistochemical Staining
Three-micron paraffin sections were deparaffinized in xylene and rehydrated in graded alcohol dilutions. Endogenous peroxidase activity was blocked by incu-bation with 3% H2O2for 5 min. Antigen retrieval was performed by pressure cooking for 10 min (121°C, 1.2 kg/cm2) in 0.01 mol/L citrate buffer (pH 6.0). The
slides were then incubated with goat anti-BD3 antibody (1:50 dilution; Santa Cruz Biotechnology, Santa Cruz, CA, USA) at room temperature for 60 min. Antibody reactions were detected with bi-otinylated link antigoat antibody (Biocare Medical, Concord, CA, USA) for 20 min at room temperature followed by incubation with Trekavidin-HRP (Biocare Medical) for 20 min. The color was developed by using DAB substrate-chromogen solution (Biocare Medical). The slides were then counterstained with hematoxylin.
Total RNA Extraction and Quantitative Real-Time PCR Analyses
The total RNA of the epidermal speci-men was extracted using TRI Reagent (Invitrogen, Carlsbad, CA, USA). The total RNA was DNase treated by using TUBRO DNase (Applied Biosystems) for 25 min at 37°C. Two micrograms of TUBRO DNase-treated RNA were re-verse transcribed using High Capacity RNA-to-cDNA kit (Applied Biosystems, Foster City, CA, USA). The cDNA sam-ples were then diluted with nuclease-free water to a final concentration equivalent to 20 ng/μL RNA input; 6 μL of these di-lutions were then subjected to 40 cycles of quantitative real-time PCR analyses using the SYBR Green Master (Roche Ap-plied Science, Indianapolis, IN, USA) and the ABI Prism 7500 Sequence Detec-tion System (Applied Biosystems). Primers for rBD-3 (sense: 5′-CAATG GAGGA ATATGCTGGCTTA-3′; antisense: 5′-GAGCTGACGA GTGCCACAAC-3′) were tested alongside the normalizing gene β-actin (β-actin-F5′-TCTGTGTGGA TTGGTGGCTCT-3′; β-actin-R5′-GACTC ATCGTACTCCTGCTTGCT-3′). The data were analyzed by use of the Delta-Delta CT method (Applied Biosystems).
Cell Proliferation and Cell Morphology
Normal human keratinocytes were ob-tained from adult foreskin through rou-tine circumcision as previously de-scribed (22). Briefly, the skin specimens were washed with phosphate-buffered saline (PBS), cut into small pieces and incubated in medium containing 0.25%
trypsin (Gibco BRL, Gaithersburg, MD, USA) overnight at 4°C. The epidermal sheet was then lifted from the dermis by using fine forceps. The epidermal cells were then pelleted by centrifugation (500g, 10 min) and dispersed into indi-vidual cells by repeated aspiration using a pipette. The keratinocytes were then gently resuspended in 5 mL of keratino-cyte-SFM medium (Gibco BRL), which contained 25 ng/mL bovine pituitary ex-tract and 5 ng/mL recombinant human epidermal growth factor. The rationale for glucose levels used was described in our previous reports (8–9). Seven days after 6 mmol/L and 26 mmol/L D-glucose treatments, the keratinocytes were subjected to a 3-(4,5-dimethylthiazol- 2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium (MTS) cell proliferation assay for determination of viability. In addition, a deoxyuridine (BrdU)–enzyme-linked im-munosorbent assay (ELISA) was used to evaluate cell proliferation. The morphol-ogy of keratinocytes after D-glucose treatment was observed under inverted microscope. The MTS (Cell-Titer 96 aqueous proliferation assay kit; Promega, Madison, WI, USA) was per-formed according to the manufacturer’s instructions. The proliferation of kerati-nocytes after D-glucose treatment was assessed by a BrdU incorporation assay using a commercially available BrdU-ELISA cell proliferation assay kit (Roche, Basel, Switzerland). The treated kerati-nocytes were fixed and stained accord-ing to the manufacturer’s instructions. The optical density was measured at 450 nm (Model 450; Bio-Rad, Hercules, CA, USA).
Evaluation of Antibacterial Activity
The paper-disc agar diffusion method with slight modification described by Jar-boe et al. (23) was employed. Briefly, the concentration of S. aureus (BCRC No. 10781) was adjusted to approximate a 0.5 MacFarland standard by adding nutrient broth (Merck, Darmstadt, Germany). Sus-pensions of 0.5 MacFarlands were swabbed evenly onto the Mueller–Hinton
agar plates (DIFCO, Becton Dickinson, Sparks, MD, USA). Immediately after in-oculation, two paper discs impregnated with 20 μL of supernatants derived from keratinocytes treated with 6 mmol/L or 26 mmol/L D-glucose for 7 d were dropped onto both sides of the agar plate and incubated in ambient air at 35°C. Six-teen hours later, the diameters of the inhi-bition zones were measured.
Measurement of hBD3 in Keratinocytes
The treatments of keratinocytes in-cluded: (a) 6 mmol/L D-glucose; (b) 12 mmol/L D-glucose; (c) 26 mmol/L D-glucose; (d) incubation with 6 mmol/L or 26 mmol/L D-glucose, followed by a stimulation with a TLR-2 agonist [20 μg/mL Pam3 Cys-Ser-(Lys)4, hy-drochloride (Calbiochem, Merck KGaA, Darmstadt, Germany)]; (e) incubation with 6 mmol/L D-glucose followed by treatment with 10 μmol/L p38 inhibitor (SB203580, Calbiochem); (f) incubation with 26 mmol/L D-glucose followed by treatment with 10 μmol/L SB203580 (Calbiochem) for 2 h, then stimulated with TLR-2 agonist; (g) incubation with 6 mmol/L D-glucose with or without ad-vanced glycation endproduct–modified bovine serum albumin (AGE-BSA) for 2 d; and (h) incubation with 6 mmol/L D-glucose with 20 mmol/L mannitol. The supernatants were collected and stored at –20°C. The concentrations of hBD3 were determined by using a commer-cially available ELISA kit (PeproTech, Rocky Hill, NJ, USA) according to the manufacturer’s instructions. The detect-able concentration range of hBD-3 was 64–4000 pg/mL.
Western Blotting Analysis
The treatments of keratinocytes in-cluded (a) 6 mmol/L D-glucose, (b) 12 mmol/L D-glucose, (c) 26 mmol/L D-glucose, (d) 6 mmol/L or 26 mmol/L D-glucose followed by stimulation with a toll-like receptor 2 (TLR2) agonist and (e) incubation with 26 mmol/L D-glucose and 2 mmol/L aminoguanidine (AG) for 7 d. Total cellular proteins were extracted
with lysis buffer (0.1% sodium dodecyl sulfate [SDS], 20 mmol/L Tris-HCl, 5 mmol/L EDTA, 2 mmol/L EGTA, and 1% Triton X-100, pH7.4) containing protease inhibitor cocktail (Roche, Mannheim, Germany). Proteins (50 μg) were loaded into 10% SDS–polyacry-lamide gel electrophoresis and trans-ferred onto a nitrocellulose membrane. After being blocked and washed, the membranes were incubated with first an-tibodies including transglutaminase-1 (Santa Cruz Biotechnology), involucrin (Santa Cruz Biotechnology), TLR-2 (Im-genex, San Diego, CA, USA), phosphory-lated p38MAPK (pp38MAPK) (Cell Sig-naling, Beverly, MA, USA), and
glyceraldehyde-3-phosphate dehydroge-nase (GAPDH) (Millipore, Billerica, MA, USA). Subsequently, the membranes were incubated with horseradish peroxi-dase (HRP)-labeled secondary antibody (Millipore) and developed in the Immo-bilonTW Western Chemiluminescent HRP Substrate (Millipore). The blots were then analyzed by digital imaging system (α Imager 2000; α Innotech, San Leandro, CA, USA).
Real-Time Quantitative PCR Detection for hBD3 and TLR2 mRNA
The treatments of keratinocytes in-cluded (a) 6 mmol/L D-glucose, (b) 12 mmol/L D-glucose, (c) 26 mmol/L D-glucose, and (d) incubation with 6 mmol/L or 26 mmol/L D-glucose, fol-lowed by stimulation with TLR2 agonist. Total RNA was extracted by using the Trizol method (Gibco BRL) and
processed as recommended by the man-ufacturer. As the PCR template 5 μg of RNA was reverse-transcribed to cDNA. The target genes analyzed were hBD3 and GAPDH. The primers used for hBD-3 were 5′-TAGCAGCTATGAGGA TCCA-3′ (forward) and 5′-CTTCGGCAGC ATTTTC GG-3′ (reverse); the primers for TLR2 were: 5′-TCTCCCATTTCCGTC TTTTT-3′ (forward) and 5′-GGTCTTGGTG TTCATTATCTTC-3′ (reverse); and for GAPDH were 5′-CCACCCATGGCAAAT TCC-3′ (forward) and 5′-GGGATTTCCA TTGATGACA-3′ (reverse). Amplification
and detection were performed with an ABI Prism 7500 sequence detection sys-tem (Applied Biosyssys-tems). The data were analyzed as described above.
Small Interfering RNA Experiment
The cultured keratinocytes were trans-fected with Lipofectamine 2000 (Invitro-gen) according to the manufacturer’s in-structions. Briefly, 1 μL of hBD3 siRNA or control small interfering RNA (siRNA) and 1 μL of Lipofectamine 2000 were each diluted with 50 μL of Opti-MEM®
I reduced serum medium (Invitrogen) without supplements and incubated at room temperature for 5 min. The se-quences of double-strand siRNA directed against HBD3 were A: 5′-CUUACGACGA CAGCACUUACGACCA-3′, and B: 5′-ACUUGCCGAUCUGUUCCUCC UUUGG-3′ (Invitrogen). The Stealth RNAi duplexes (Invitrogen) were used as a negative control. Subsequently, they were mixed and incubated at room tem-perature for 20 min. One milliliter of the mixture (oligomer-Lipofectamine 2000 complex) was added to each well and in-cubated at 37°C for 6 h. The reaction me-dium was changed into keratinocyte cul-ture medium and incubated at 37°C for 72 h. Quantitative PCR was performed to confirm the success of transfection. The supernatants were then collected and subjected to the antimicrobial activity ex-periment as described previously.
Statistical Analysis
For each experiment, at least three in-dependent experiments were performed. Quantitative results are expressed as mean ± SD. Statistical analysis was de-termined using the Student t test. A P value less than 0.05 was considered as significant.
RESULTS
Diabetic Rats Showed Delayed Wound Healing and Poor BD3 Induction Compared with Control Rats
As expected, the wounds of diabetic rats (n = 9) showed significantly delayed healing time compared with the wounds
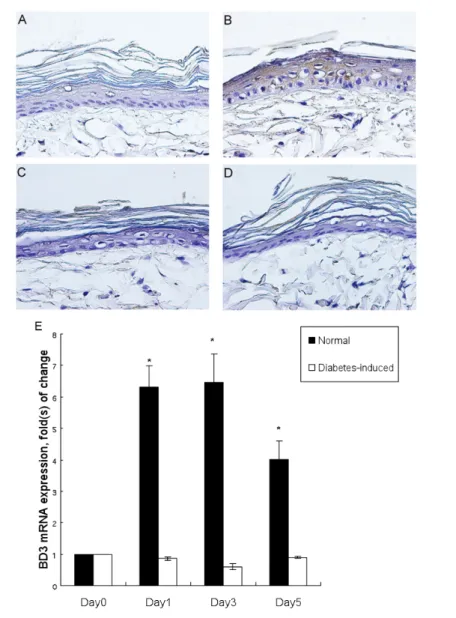
of control rats (n = 9). More specifically, the healing times for the control and dia-betic rats were 9.3 ± 0.8 d and 11.8 ± 1.9 d, respectively. Because the wounding pro-cess has been reported to induce BD ex-pression (24), the IHC staining was per-formed on the perilesional skin in a sequential manner. The expressions of BD3 appeared 1 d after skin wounding on the upper layers of the perilesional skin of the control rats, whereas the per-ilesional specimen obtained from the dia-betic rats demonstrated negligible BD3 expression even after wounding. The BD3 staining of the skin obtained from the control rats became more intense 3 d after wounding (Figure 1A–D). On the other hand, throughout the experiment, the BD3 expression was not observed on the perilesional diabetic rat skin. Corrobo-rating with the results of IHC staining, the results of quantitative PCR analyses of the control perilesional skin showed signifi-cant increases of BD3 mRNA expression (an approximately six-fold increase at day 1 and day 3 compared with day 0) after wounding, whereas no significant increase was observed in the diabetic rat skin (Figure 1E).
Keratinocytes Cultivated under Normal- and High-Glucose Conditions Showed Comparable Viability, Growth and Differentiation after 7 Days of Treatment
Our previously reported results showed that prolonged cultivation of ke-ratinocytes under high-glucose environ-mental conditions resulted in decreased cellular survival, possibly due to deple-tion of growth supplements and accumu-lation of harmful metabolic byproducts. Because we intended to evaluate the ef-fects of a high-glucose environment on keratinocytes for a longer term, the cul-ture medium was changed every other day. The viability of keratinocytes cul-tured at various glucose levels was not significantly different, as determined by the MTS assay (data not shown). In addi-tion, cell proliferaaddi-tion, as determined by the BrdU-ELISA assay, showed no signif-icant difference between the normal- and
high-glucose cultivated groups. More-over, microscope analysis revealed no significant differences in cellular mor-phology among the keratinocytes treated with different levels of glucose. Involu-crine and transglutaminase-1 are two im-portant markers for characterizing the differentiation status of keratinocytes. High- glucose treatment did not signifi-cantly alter the expression of these pro-teins (Figure 2). These results indicated that the cellular physiologic status was comparable between the normo- and
high-glucose groups under our experi-mental conditions.
Keratinocytes Cultured at High-Glucose Settings Showed Decreased hBD3 at Both mRNA and Protein Levels
Because our experimental conditions did not significantly affect the growth and the differentiation status of kerati-nocytes, we next evaluated the effects of high-glucose treatment on keratinocytes in terms of hBD3 production. The levels of hBD3 in the culture supernatants
ob-Figure 1. Expression of BD3 in wounded rat skin. (A–D) Immunohistochemical BD3 staining (brown) of the perilesional skin of the control (A, B) and the diabetic rat (C, D) at day 0 and 3 after wounding respectively. Original magnification 400×. (E) The BD3 mRNA levels of perilesional rat skin at different time points after wounding *P <0.05 compared with day 0.
tained from the keratinocytes treated with 12 mmol/L and 26 mmol/L glu-cose were significantly lower (approxi-mately 67% and 60%, respectively) than their normo-glucose (6 mmol/L) coun-terpart. Corroborating with the protein expression, the hBD3 mRNA expression of the keratinocytes cultivated in 12 mmol/L and 26 mmol/L glucose were also significantly lower (approxi-mately 70% and 54%, respectively) com-pared with the 6 mmol/L control group (Figure 3). Treating cultured keratino-cytes with the p38 inhibitor SB203580 significantly decreased the levels of hBD3 to 56.5% ± 4.1% of the control. The osmolarity control treatment group showed an insignificant reduction of hBD3 from 583.5 ± 15.0 pg/mL to 483.7 ± 35.1 pg/mL, indicating that the changes in glucose and not osmolarity levels are responsible for reduction of hBD3 expression.
Culture Supernatants from
Keratinocytes Cultivated in Normal Glucose Concentrations Showed Better Anti–S. Aureus Activity than Keratinocytes Cultivated in High Glucose
Results of a previous study showed that hBD3 is the key molecule with which keratinocytes kill S. aureus (21). Therefore, we examined the anti–S au-reus activity of the culture supernatants derived from different culture condi-tions. The supernatants from keratino-cytes cultured with high glucose exhib-ited only approximately 80% of the anti–S. aureus capacity of keratinocytes cultured with normal glucose (Figure 4). This result indicates that high-glucose treatment rendered keratinocytes less capable of fending off S. aureus. To con-firm the functional role of hBD3 on
anti–S. aureus capacity in our experi-ment, we used gene silencing of hBD3 by siRNA. Accordingly, the super-natants from the siRNA-treated kerati-nocytes showed only 70% ± 8.0% capac-ity against S. aureus growth compared with the control vehicle. This result in-dicated that hBD3 was at least partially responsible for inhibiting S. aureus growth in our experiments. The remain-ing antimicrobial activity noted in the experiment was likely due to antibiotics present in the medium we used for cell culture and possibly due to other an-timicrobial peptides released by kerati-nocytes. To confirm this hypothesis, su-pernatants were collected from
keratinocytes cultured without the addi-tion of antibiotics, and similar experi-ments were repeated. However, none of the supernatants obtained under these conditions demonstrated significant an-timicrobial activity, regardless of the glucose levels during culture.
No significant differences in the ex-pression of TLR-2 were observed be-tween the normal- and
high-glucose–cultivated groups. TLR-2 is an important regulator of hBD3 expression. High-glucose treatment may reduce TLR-2 expression, which can lead to re-duced hBD3 production. Therefore, we evaluated the expression of TLR-2 in ke-ratinocytes under high-glucose cultiva-tion. As demonstrated by the data pre-sented in Figure 5, high-glucose treatment did not significantly alter the expression of TLR-2 at the protein and mRNA levels.
Figure 2. Effect of high-glucose treatment on human cultured keratinocyte differen-tiation. (A) The effect of high-glucose treatment on transglutaminase-1 and in-volucrin expressions of keratinocytes. The figure shown represents one experiment repeated 3x with similar results. (B) Densit-omeric analysis of (A), ratio of differentia-tion markers/GAPDH, with 6 mmol/L glu-cose treatment representing 100%.
Figure 3. Effects of different glucose levels on hBD3 production of cultured human keratinocytes. (A) The protein expressions of hBD3 in the supernatants of keratino-cytes cultivated under different levels of glucose. (B) The hBD3 mRNA expressions of the keratinocytes cultivated under differ-ent glucose levels. The expression of hBD3 for keratinocytes cultivated with 6 mmol/L glucose representing 100% in the figure. *P < 0.05.
Figure 4. The anti–S. aureus activity of the supernatants obtained from keratinocytes cultivated under different glucose condi-tions The diameters of the inhibition zones were shown. *P < 0.05.
109.83 pg/mL and 341.72 ± 7.68 pg/mL to 627.39 ± 107.21 pg/mL, respectively. However, after TLR-2 stimulation the ab-solute levels of hBD3 from the group cul-tivated in normal glucose remained sig-nificantly higher than the levels of their high-glucose counterpart. These results indicated that the keratinocyte TLR-2 re-mained functional after high-glucose treatment, and a certain constitutive in-hibitory effect was imparted by the high-glucose treatment that resulted in re-duced hBD3 production.
Phosphorylated p38MAPK Expression Was Significantly Reduced in
Keratinocytes Cultivated in High Glucose
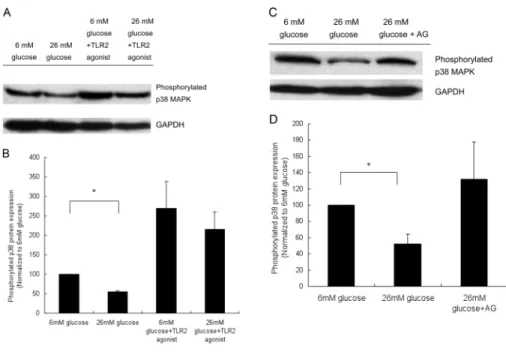
Treatment with a TLR2 agonist signifi-cantly increased the expression of phos-phorylated p38MAPK. Both p38MAPK and nuclear factor κB (NFκB) signaling are known to regulate hBD3 expressions. As demonstrated in Figure 6, both 12-mmol/L and 26-mmol/L glucose treatment significantly decreased the constitutive expression of phosphorylated p38MAPK in cultured keratinocytes. On the other hand, the NFκB-binding activities of keratinocytes treated with 26 mmol/L glucose were 1.12 ± .19–fold greater than those of the 6-mmol/L glucose group, with no significant differences noted be-tween the two groups. Treating cultured keratinocytes with a TLR-2 agonist re-sulted in a clear increase of phosphory-lated p38MAPK expression, even in the high-glucose–cultivated keratinocytes, as shown in Figure 7A.
SB203580, a p38MAPK Inhibitor, Abrogated the hBD3-Enhancing Effect of TLR2 on High-Glucose–Treated Keratinocytes
Because the TLR-2 agonist increased phosphorylated p38MAPK expression and hBD3 production of the glucose–cultivated keratinocytes, SB203580 was added to determine the functional role of enhanced phosphory-lated p38MAPK in TLR-2 induced hBD3 increase. As expected, treating the high-glucose–cultivated keratinocytes with
SB203580 abrogated the enhancing effect of TLR-2 agonist on hBD3 expressions. More specifically, the hBD3 levels de-creased from 564.5 ± 1.2 pg/mL to 330.4 ± 2.3 pg/mL (approximately 60%), con-firming the importance of p38MAPK sig-naling in TLR-2–induced hBD3 response and validating the integrity of TLR-2 function under the experimental condi-tions of our study.
AGE-BSA Significantly Decreased the hBD3 and the Phosphorylated p38MAPK Expressions of Cultured Keratinocytes
High-glucose treatments are known to induce AGE accumulation. However, the effects of AGE on hBD3 production have not been documented. Treating keratino-cytes with BSA and AGE-BSA resulted in hBD3 levels of 763.3 ± 16.6 pg/mL and 556.6 ± 1.3 pg/mL, respectively,
indicat-Figure 5. Effect of high glucose cultivation on TLR-2 expressions of keratinocytes. (A) The protein expressions of TLR-2 in keratino-cytes cultivated under different levels of glucose. A representative blot from three independent experiments with similar re-sult is shown. (B) Densitometric analysis of (A), ratio of TLR2/GAPDH, with 6 mmol/L glucose treatment representing 100%. (C) The TLR-2 mRNA expressions of the kerati-nocytes cultivated under different levels of glucose with keratinocytes cultivated with 6 mmol/L glucose representing 100% in the figure. *P < 0.05.
Figure 6. The phosphorylated p38MAPK expression of cultured human keratino-cytes after indicated treatment condition. (A) The protein expressions of phosphory-lated p38MAPK in keratinocytes cultivated with 6 mmol/L, 12 mmol/L and 26 mmol/L glucose. A representative blot from three independent experiments with similar re-sults is shown. (B) The densitometric analy-sis of (A), ratio of phosporylated p38MAPK/ GAPDH, with 6 mmol/L glucose treatment representing 100%. *P < 0.05.
Treating Keratinocytes with a TLR-2 Agonist Significantly Increased hBD3 Expression
Because TLR-2 is an important regula-tor of hBD3 expression and its expression was not affected by high-glucose treat-ment, we next evaluated the functional-ity of TLR-2 on keratinocytes cultivated under high-glucose conditions. Treating keratinocytes with a TLR-2 agonist in-creased the protein and mRNA expres-sion of hBD3 more than two-fold regard-less of the glucose treatment. More specifically, the hBD3 levels of the kerati-nocytes cultivated in 6 mmol/L and 26 mmol/L glucose increased from 583.48 ± 76.40 pg/mL to 1148.24 ±
ing that AGE significantly reduced the hBD3 production of keratinocytes. To de-termine if AGE formation was involved in suppressing constitutive phosphory-lated p38MAPK expression in the high-glucose–treated keratinocytes, AG, an in-hibitor of AGE formation, was added to the high-glucose–treated keratinocyte culture. Restoration of phosphorylated p38MAPK expression was found in the high-glucose–treated keratinocytes after introduction of AG (Figure 7C).
DISCUSSION
It has been shown that wounding of the skin initiates various innate immune responses, including induction of hBD3 expression via transactivation of the epi-dermal growth factor receptor (EGFR) (24). Because it is impractical to perform sequential repeated biopsies on human diabetic patients, an animal model was
employed to study the effect of hypergly-cemia on epidermal BD expression. In parallel with previous findings, our rat model showed enhanced BD3 expression at both mRNA and protein levels after skin wounding. On the other hand, the perilesional skin of the diabetic rat wound showed negligible BD3 induction even after wounding, indicating that BD3 response was dysfunctional under hyper-glycemic conditions. To mimic diabetic conditions in human beings, additional in vitro studies were performed using cul-tured human keratinocytes under various glucose-treatment conditions.
As demonstrated by our results, high-glucose cultivation reduced the constitu-tive hBD3 expression of human keratino-cytes, both at mRNA and protein levels. In addition, the supernatants obtained from keratinocytes grown in cultures containing high glucose concentrations
showed decreased bactericidal capacity toward S. aureus compared with kerati-nocytes grown in cultures containing normal glucose concentrations. These data strongly suggest that hBD3 expres-sion and function of keratinocytes were suboptimal at hyperglycemic settings, and this particular dysfunction at least contributed to the high infection rates as-sociated with diabetic wounds. It should be noted that the osmolarity control treatment using mannitol did not signifi-cantly affect the concentrations of hBD3. This result further validated and strengthened the notion that high glu-cose levels rather than elevated osmolar-ity were responsible for the reduced hBD3 levels observed. It should be noted that the supernatants obtained from ke-ratinocytes cultured without antibiotics did not demonstrate significant antimi-crobial activity in our experimental con-ditions. It has previously been shown that certain synergistic antimicrobial ef-fects can be found when suboptimal doses of antibiotic and hBD3 are com-bined (25). Therefore, it is likely that the concentrations of hBD3 in our experi-mental conditions were not adequate to induce significant antimicrobial effects alone, but when combined with exoge-nous antibiotic, the levels of hBD3 pres-ent were able to induce a synergistic effect.
TLRs played an important regulatory role in hBD synthesis. Human skin ex-pressed both TLR-2 and TLR-4, whereas cultured primary normal human kerati-nocytes expressed functional TLR-2, but not TLR-4 (26). TLR-2 is considered the major receptor for Gram-positive bacte-ria by virtue of its ability to recognize bacterial cell –wall components (27). When cultured under high-glucose con-ditions, TLR-2 expression on keratino-cytes was not significantly altered. More-over, addition of TLR-2 agonist was able to increase the hBD3 levels of both nor-mal- and high-glucose–treated groups, although the absolute hBD3 levels were still significantly lower in the glucose–cultivated group. These results indicate that at clinical settings, the
ab-Figure 7. The phosphorylated p38MAPK expression of cultured human keratinocytes after indicated treatment condition. (A) The protein expressions of phosphorylated p38MAPK in keratinocytes with or without TLR-2–agonist stimulation were evaluated. A representative blot from three independent experiments with similar results is shown. (B) The densitomet-ric analysis of (A), ratio of phosporylated p38MAPK/GAPDH, with 6 mmol/L glucose treat-ment representing 100%. (C) The effects of AG, an inhibitor of advanced glycation end-product, on protein expression of phosphorylated p38MAPK expression of high glucose – cultivated keratinocyte. The figure shown is representative of blots from three indepen-dent experiments with similar results. (D) Densitometric analysis of (C), ratio phosphory-lated p38/GAPDH, with 6 mmol/L glucose treatment representing 100%. *P < 0.05.
solute hBD3 levels may not be ade-quately expressed in response to normal physiological stress such as skin wound-ing or infection at hyperglycemic set-tings. In addition, these results also demonstrated that the TLR-2 signaling of the high-glucose–treated keratinocytes remained functional.
Induction of defensin synthesis may involve several cascades, including p38MAPK and NFκB pathways (28–31). As demonstrated by our results, high-glucose conditions reduced constitutive phosphorylated p38MAPK expression but has a limited impact on active NFκB binding. Therefore, the constitutively downregulated p38MAPK signaling con-tributed to the reduced hBD3 expressions under high-glucose environmental con-ditions. Intriguingly, that TLR-2 agonist was able to increase the expression of phosphorylated p38MAPK, resulting in restoration of hBD3 levels. These results indicated that TLR-2 signaling remained intact in a high-glucose environment and that the TLR-2 agonist may be clinically useful for promoting better diabetic wound healing through upregulation of hBD3, an effective antimicrobial peptide and important component of innate immunity.
Increased levels of advanced glycation endproducts (AGE) have been implicated in complications associated with diabetes (32). It has been recognized that glucose can react directly with protein amino groups (glycation) to form labile Schiff bases which subsequently rearranged to more stable Amadori product. In time, glycated protein yielded reactive dicar-bonyl compounds that can form AGE. Under physiologic conditions, AGEs are formed constantly in the body. The rate of formation is prominently increased in the presence of high glucose and/or in-creased oxidative stress. Previous studies have shown that AGE imparted detri-mental effects on fibroblasts and endothe-lial cells (33,34), the important partici-pants of wound healing process. It has also been shown that AGE accumulation inside the cell is a rapid process as Gia-rdino et al. (35) had shown that 7 days of
high-glucose (30 mmol/L) treatment re-sulted in a 13.8-fold increase of AGE ac-cumulation compared with cells cultured in normal (5 mmol/L) glucose. AG, a nu-cleophic hydrazine compound, inhibits the formation of AGE and has been shown to prevent impaired wound heal-ing in diabetic rats (36,37). Therefore, the effects of AGE on keratinocytes in terms of hBD3 production and p38MAPK sig-naling were examined. Treating keratino-cytes with AGE significantly decreased the levels of hBD3 production. In addi-tion, treating the high-glucose–cultivated keratinocytes with AG showed reversal of the reduced phosphorylated
p38MAPK expression, indicating that the increased formation of AGE contributed to the reduced phosphorylated
p38MAPK expression observed in kerati-nocytes cultivated at high glucose levels. Recently reported data demonstrate that inhibition of p38MAPK activity during wounding blocked EGFR internalization (38), an event crucial for hBD3 induction. Therefore, downregulation of p38MAPK signaling by AGE may be the early event responsible for suboptimal hBD3 expres-sion found in diabetic wounds. It should be noted that previous reports have shown that an AGE- induced transient in-crease of p38MAPK signaling that re-turned to the basal level within 45 min (39,40). Our current results suggested that after initial transient elevation, sustained AGE exposure inhibited the activity of the p38MAPK cascade. Further studies clarifying the differences between the short- and long-term effects of AGE on cellular physiology are warranted.
In summary, we demonstrated that a high-glucose environment reduced hBD3 expression in keratinocytes. This patho-genic process involved inhibition of p38MAPK signaling, an event that re-sulted from increased AGE formation. In addition, the TLR-2 agonist was able to elevate the hBD3 expressions in glucose–cultivated keratinocytes. Recently reported data suggested that enhanced hBD3 expression promoted wound heal-ing in infected diabetic wounds (41). Our current findings provide further support
for this approach. Although hyper-glycemic conditions inhibited constitutive hBD3 expression on keratinocytes and re-sulted in inadequate hBD3 upregulation upon stimulation, the signaling via TLR-2 appeared to be intact. The topical TLR (TLR7 and TLR8) agonist imiquimod has already been used for treatment of vari-ous dermatologic conditions, and topical TLR2 agonists treatment for managing di-abetic wounds is a strategy worthy of in-vestigation in the near future.
ACKNOWLEDGMENTS
This study was supported by National Science Council of Taiwan (NSC 98-2314-B-037-016-MY3; NSC 98-2314-B-037-017-MY3).
DISCLOSURE
The authors declare that they have no competing interests as defined by Molec-ular Medicine, or other interests that might be perceived to influence the re-sults and discussion reported in this paper.
REFERENCES
1. Abdulrazak A, Bitar ZI, Al-Shamali AA, Mobasher LA. (2005) Bacteriological study of di-abetic foot infections. J. Diabetes Complications. 19:138–41.
2. Calvet HM, Yoshikawa TT. (2001) Infections in diabetes. Infect. Dis. Clin. North. Am. 15:407–21. 3. Muller LM, et al. (2005) Increased risk of
com-mon infections in patients with type 1 and type 2 diabetes mellitus. Clin. Infect. Dis. 41:281–8. 4. Shah BR, Hux JE. (2003) Quantifying the risk of
infectious diseases for people with diabetes.
Dia-betes Care. 26:510–3.
5. Delamaire M, et al. (1997) Impaired leucocyte func-tions in diabetic patients. Diabet. Med. 14:29–34. 6. Gallacher SJ, et al. (1995) Neutrophil bactericidal
function in diabetes mellitus: evidence for associ-ation with blood glucose control. Diabet. Med. 12:916–20.
7. Sheetz MJ, King GL. (2002) Molecular under-standing of hyperglycemia’s adverse effects for diabetic complications. JAMA. 288:2579–88. 8. Lan CC, Liu IH, Fang AH, Wen CH, Wu CS.
(2008) Hyperglycaemic conditions decrease cul-tured keratinocyte mobility: implications for im-paired wound healing in patients with diabetes.
Br. J. Dermatol. 159:1103–15.
9. Lan CC, Wu CS, Kuo HY, Huang SM, Chen GS. (2009) Hyperglycaemic conditions hamper ke-ratinocyte locomotion via sequential inhibition of distinct pathways: new insights on poor wound
closure in patients with diabetes. Br. J. Dermatol. 160:1206–14.
10. Lehrer RI, Ganz T. (2002) Defensins of vertebrate animals. Curr. Opin. Immunol. 14:96–102. 11. Froy O, Hananel A, Chapnik N, Madar Z. (2005)
Differential expression of rat beta-defensins.
IUBMB Life. 57:41–3.
12. Yang D, Biragyn A, Kwak LW, Oppenheim JJ. (2002) Mammalian defensins in immunity: more than just microbicidal. Trends Immunol. 23:291–6. 13. Ali RS, et al. (2001) Expression of the peptide an-tibiotics human beta defensin-1 and human beta defensin-2 in normal human skin. J. Invest.
Der-matol. 117:106–11.
14. Becker MN, Diamond G, Verghese MW, Randell SH. (2000) CD14-dependent lipopolysaccharide-induced beta-defensin-2 expression in human tracheobronchial epithelium. J. Biol. Chem. 275:29731–6.
15. Fulton C, Anderson GM, Zasloff M, Bull R, Quinn AG. (1997) Expression of natural peptide antibiotics in human skin. Lancet. 350:1750–1. 16. Jang BC, et al. (2004) Up-regulation of human beta-defensin 2 by interleukin-1beta in A549 cells: involvement of PI3K, PKC, p38 MAPK, JNK, and NF-kappaB. Biochem. Biophys. Res.
Com-mun. 320:1026–33.
17. Jia HP, et al. (2001) Discovery of new human beta-defensins using a genomics-based approach.
Gene. 263:211–8.
18. Singh PK, et al. (1998) Production of defensins by human airway epithelia. Proc. Natl.
Acad. Sci. U. S. A. 95:14961–6.
19. Sorensen OE, et al. (2005) Differential regulation of beta-defensin expression in human skin by mi-crobial stimuli. J. Immunol. 174:4870–9.
20. Goldstein EJ, Citron DM, Nesbit CA. (1996) Dia-betic foot infections. Bacteriology and activity of 10 oral antimicrobial agents against bacteria isolated from consecutive cases. Diabetes Care. 19:638–41. 21. Kisich KO, et al. (2007) The constitutive capacity
of human keratinocytes to kill Staphylococcus aureus is dependent on beta-defensin 3. J. Invest.
Dermatol. 127:2368–80.
22. Lan CC, et al. (2005) FK506 promotes melanocyte and melanoblast growth and creates a favorable milieu for cell migration via keratinocytes: Possi-ble mechanisms of how tacrolimus ointment in-duces repigmentation in patients with vitiligo.
Br. J. Dermatol. 153:498–505.
23. Jarboe E, et al. (1998) Evaluation of a disk diffu-sion method for determining susceptibility of My-cobacterium avium complex to clarithromycin.
Diagn. Microbiol. Infect. Dis. 30:197–203.
24. Sorensen OE, et al. (2006) Injury-induced innate immune response in human skin mediated by transactivation of the epidermal growth factor re-ceptor. J. Clin. Invest. 116:1878–85.
25. Midorikawa K, et al. (2003) Infect. Immun. 71:3730–9. 26. Kawai K, Shimura H, Minagawa M, Ito A,
Tomiyama K, Ito M. (2002) Expression of func-tional Toll-like receptor 2 on human epidermal keratinocytes. J. Dermatol. Sci. 30:185–94. 27. Schroder NW, et al. (2003) Lipoteichoic acid (LTA)
of Streptococcus pneumoniae and Staphylococcus aureus activates immune cells via Toll-like recep-tor (TLR)-2, lipopolysaccharide-binding protein (LBP), and CD14, whereas TLR-4 and MD-2 are not involved. J. Biol. Chem. 278:15587–94. 28. McDermott AM, et al. (2003) Defensin expression
by the cornea: multiple signalling pathways me-diate IL-1beta stimulation of hBD-2 expression by human corneal epithelial cells. Invest.
Ophthal-mol. Vis. Sci. 44:1859–65.
29. Ogushi K, et al. (2001) Salmonella enteritidis FliC (flagella filament protein) induces human beta-defensin-2 mRNA production by Caco-2 cells. J. Biol. Chem. 276:30521–6.
30. Wada A, et al. (2001) Helicobacter pylori-mediated transcriptional regulation of the human defensin 2 gene requires NF-kappaB. Cell.
Micro-biol. 3:115–23.
31. Whitmarsh AJ, Davis RJ. (1996) Transcription fac-tor AP-1 regulation by mitogen-activated protein kinase signal transduction pathways. J. Mol. Med. 74:589–607.
32. Brownlee M. (1994) Lilly Lecture 1993. Glycation and diabetic complications. Diabetes. 43:836–41. 33. Alikhani M, et al. (2007) Advanced glycation end
products induce apoptosis in fibroblasts through activation of ROS, MAP kinases, and the FOXO1 transcription factor. Am. J. Physiol. Cell. Physiol. 292:C850–6.
34. Duraisamy Y, et al. (2003) Aminosalicylic acid re-duces the antiproliferative effect of hypercaemia, advanced glycation endproducts and gly-cated basic fibroblast growth factor in cultured bovine aortic endothelial cells: comparison with aminoguanidine. Mol. Cell. Biochem. 246:143–53. 35. Giardino I, Fard AK, Hatchell DL, Brownlee M. (1998) Aminoguanidine inhibits reactive oxygen species formation, lipid peroxidation, and oxi-dant-induced apoptosis. Diabetes. 47:1114–20. 36. Teixeira AS, Caliari MV, Rocha OA, Machado RD, Andrade SP. (1999) Aminoguanidine pre-vents impaired healing and deficient angiogene-sis in diabetic rats. Inflammation. 23:569–581 37. Yavuz D, et al. (2005) Collagen ultrastructure and
TGF-beta1 expression preserved with aminoguani-dine during wound healing in diabetic rats. Endocr.
Res. 31:229–43.
38. Lambert S, Frankart A, Poumay Y. (2010) p38 MAPK-regulated EGFR internalization takes place in keratinocyte monolayer during stress conditions. Arch. Dermatol. Res. 302:229–33. 39. Lander HM, et al. (1997) Activation of the receptor
for advanced glycation end products triggers a p21(ras)-dependent mitogen-activated protein ki-nase pathway regulated by oxidant stress. J. Biol.
Chem. 272:17810–4.
40. Yeh CH, et al. (2001) Requirement for p38 and p44/p42 mitogen-activated protein kinases in RAGE-mediated nuclear factor-kappaB transcrip-tional activation and cytokine secretion. Diabetes. 50:1495–504.
41. Hirsch T, et al. (2009) Human beta-defensin-3 promotes wound healing in infected diabetic wounds. J. Gene. Med. 11:220–8.