牛樟芝純化成分抑制幽門螺旋桿菌誘導人類胃上皮細胞的發炎作用; Antrodia camphorata constituents inhibit Helicobacter pylori- induced inflammation in human gastric epithelial cells
51
0
0
全文
(2) Content Abstract.......................................................................................................................III 中文摘要 (Abstract in Chinese) ................................................................................V Introduction..................................................................................................................1 Helicobacter pylori infection and inflammation responses ...............................1 Treatments of H. pylori infection ........................................................................2 Antrodia camphorata ............................................................................................5 Materials and methods ................................................................................................8 Bacterial strain and cell culture..........................................................................8 Determination of anti-bacterial activity.............................................................8 Cell survival assay (MTT method) .....................................................................9 Inhibition of H. pylori adherence to and invasion into epithelial cells ............9 Transient transfection of NF-κB and IL-8 reporter genes .............................10 Measurement of interleukin-8 expression........................................................11 Inhibition of macrophage activation ................................................................11 Western blot for assay of iNOS expression......................................................12 Cell survival assay (MTT method) in macrophages .......................................12 Statistical evaluation ..........................................................................................13 Results .........................................................................................................................14 Inhibition of H. pylori growth ...........................................................................14 Treatment of A. camphorata constituents AC-3, AC-10, and AC-12 does influence epithelial cell viability .......................................................................14 Inhibition of H. pylori adherence and invasion of gastric epithelial cells .....15 AC-3 inhibits H. pylori-induced inflammation of gastric epithelial cells......15 Treatment of A. camphorata constituents AC-3, AC-10, and AC-12 does influence macrophages viability .......................................................................17 A. camphorata constituents AC-3 and AC-10 inhibit LPS-mediated nitric oxide production in macrophages ....................................................................17 Western blot of inhibition of iNOS production ...............................................18 Discussion and conclusion .........................................................................................19 References...................................................................................................................22 Table 1 Molecular weight of A. camphorata constituents ...............................31 Table 2 Inhibitory effects of A. camphorata constituents ...............................32 Fig. 1. Chemiacl structures of A. camphorata constituents 1-11 ....................33 Fig. 2. Effect of A. camphorata constituents (AC-3, AC-10, and AC-12) and DMSO control on gastric epithelial cell viability. ...........................................36 Fig. 3. Effect of A. camphorata constituents (AC-3, AC-10, and AC-12) on H. pylori adhesion of gastric epithelial cells..........................................................37 I.
(3) Fig. 4. Effect of A. camphorata constituents (AC-3, AC-10, and AC-12) on H. pylori invasion of gastric epithelial cells...........................................................38 Fig. 5. Effect of A. camphorata constituents (AC-3, AC-10, and AC-12) on H. pylori-induced NF-κB activation in gastric epithelial cells.............................39 Fig. 6. Effect of A. camphorata constituents (AC-3, AC-10, and AC-12) on H. pylori-induced IL-8–luciferase activity in gastric epithelial cells. .................40 Fig. 7. Effect of A. camphorata constituents (AC-3, AC-10, and AC-12) and DMSO control in RAW 264.7 cells. ..................................................................41 Fig. 8. Inhibition by A. camphorata constituents (AC-3, AC-10, and AC-12) and DMSO control of LPS-induced NO production in RAW 264.7 cells......42 Fig. 9. Effects of A. camphorata constituents (AC-3 and AC-10) on the iNOS protein expression in RAW 264.7 cells. ............................................................43 Appendix 1 Therapy of H. pylori infection ......................................................44 Appendix 2 Drugs used in H. pylori therapy ...................................................45. II.
(4) Abstract Helicobacter pylori is a Gram-negative, micro-aerophilic bacterium that inhabits various areas of the stomach and duodenum. It causes a chronic inflammation of the stomach lining and strongly linked to the development of duodenal and gastric ulcers and stomach cancer. It has been well confirmed that infection with H. pylori is an etiological agent of peptic ulcer, and eradication of this organism can not only make the associated gastritis regression but cure these diseases. There are several treatments of H. pylori infection, most common treatment is triple therapy, including two antibiotics and a proton pump inhibitor. However, the increasing of antibiotic resistant rate was associated with the usage of antimicrobial agents in the treatment of H. pylori infection. Thus, the complementary therapeutic methods in eradication of H. pylori were needed. Antrodia camphorata (A. camphorata) is a rare treasure in Taiwan, the fruiting body of A. camphorata is well known by named niu-chang-chih or jang-jy. The miraculous efficacy in human body had been investigated and researched by many organizations in Taiwan. In this study, thirteen compounds were isolated from A. camphorata. Among them, compound 15α-Acetyldehydrosulphurenic acid (number 3) and Antcin C (number 10) had higher inhibition effects on H. pylori-induced inflammatory responses in gastric epithelial cells and mouse macrophage RAW 264.7 cells than other compounds. In addition, these constituents also inhibited the IL-8 protein expression and NF-κB signal III.
(5) activation in H. pylori-infected gastric epithelial cells and it might contribute to the potential role of A. camphorata as an anti-inflammatory action. Results from our study might bring a new method for treating of H. pylori infection.. Keywords: H. pylori, Antrodia camphorata, inflammation, NF-κB, IL-8 Abbreviations: NO, nitric oxide; DMSO, dimethylsulfoxide; NF-κB, nuclear factor (NF)-κB; IL-6, interleukin-6; IL-8, interleukin-8; iNOS, inducible nitric oxide; LPS, lipopolysaccharide. IV.
(6) 中文摘要 (Abstract in Chinese) 幽門螺旋菌 (H. pylori) 是一種微好氧螺旋形狀的革蘭氏陰性菌,會存在於 人類的胃以及十二指腸的表皮黏膜上。幽門螺旋菌會引起胃襯的慢性發炎並且與 十二指腸、胃潰瘍和以及胃癌的發展有密切關係。幽門螺旋菌對於胃腸道潰瘍的 發生有密切相關,一般認為滅除幽門螺旋菌可以使得胃發炎復原,亦可治療潰瘍 性疾病。現有多種治療幽門螺旋菌感染的療法,最為普遍使用的是三合一療法, 包含了兩種抗生素與一種胃酸分泌抑制劑。然而,在治療幽門螺旋菌感染上,抗 生素抗藥性的增加與抗微生物劑的使用有關。因此,根治幽門螺旋菌過程中需要 有輔助治療的方法。樟薄孔菌(Antrodia camphorata)是在台灣的一種稀有的藥 材,其子果實的名稱是眾所周知的牛樟芝或樟芝。在台灣,牛樟芝對人體的神奇 功效,被許多機構廣泛的調查與研究。在本篇論文中,從牛樟芝上分離出 13 種 萃取物。在 13 種萃取物中,不論是在胃上皮細胞與小鼠巨噬細胞上, 15α-Acetyldehydrosulphurenic acid (AC-3)與 Antcin C (AC-10)對於幽門螺旋菌擁 有較高的抑制效果。此外,對於 NF-κB 以及 IL-8 的抑制,也顯示出 AC 萃取物 的抗發炎功效。期望此實驗結果可以為治療幽門螺旋菌感染帶來新的方法。. . V.
(7) Introduction Helicobacter pylori infection and inflammation responses Helicobacter pylori is a Gram-negative micro-aerophilic gastric pathogen that infects approximately half of the human population (Marshall, 2002; Parsonnet, 1998; Warren, 1983). Persistent colonization is associated with chronic inflammation, gastric atrophy, gastric adenocarcinoma and mucosa-associated lymphoid tissue (MALT) lymphoma (Parsonnet, 1998). The standard, recommended method of treating infected patients with severe symptoms was the combination of a proton pump inhibitor and two antibiotics: mainly clarithromycin, and amoxicillin or metronidazole (Hentschel et al., 1993; Logan et al., 1991). Indeed, an eradication rate of more than 90% was found in a number of reports based on this combination therapy (Gu et al., 2006; Hsu et al., 2005; Mirbagheri et al., 2006; Zullo et al., 2005). Given the widespread use of antibiotics in pass years, it was not unexpected that a few studies showed a relatively high failure rate (20% to 40%) (Bochenek et al., 2003; McMahon et al., 2003; Tankovic et al., 2001b). In parallel, antimicrobial resistance was found to be the main cause of therapy failure (Megraud and Lamouliatte, 2003; Poon et al., 2002; Tankovic et al., 2001a).. Chronic gastritis is induced by mononuclear cell infiltrates, included of lymphocyte and macrophages. In the study of macrophages, which are major source. 1.
(8) of the IL-6 present in H. pylori induced chronic gastritis, are known to play a critical role in the pathogenesis of mucosal inflammations (Sartor, 1994). It was also reported that macrophages secretion increased levels of IL-1 and TNF-α in H. pylori infected gastric tissues (Harris et al., 2000; Yamaoka et al., 1996). Another bactericidal agent of macrophages, nitric oxide (NO) is generated by inducible nitric oxide synthase (iNOS)-mediated conversion of L-arginine to L-citrulline. However, H. pylori can survive despite marked induction of inducible iNOS in macrophages (Gobert et al., 2001). In infected H. pylori on gastric epithelial cells, expression of CagA resulted in translocation of NF-κB into nucleus and activation of IL-8 transcription (Brandt et al., 2005). Accumulated reports have been found that functional CagA is capable not only of inducing actin rearrangement but also having their potential in induction of IL-8 secretion (Brandt et al., 2005; Kim et al., 2006), thus contributing to inflammation in H. pylori-induced gastric diseases. With experiments involving H. pylori-infection, IL-8 secretion from epithelial cells is directly dependent on the NF-κB signal pathway (Brandt et al., 2005; Nozawa et al., 2002). Treatments of H. pylori infection A number of studies have been reported that H. pylori was related to the induction of gastritis, peptic ulcer, and gastric cancer (Graham et al., 2009). Elimination of H. pylori can obviously reduce the relapse of ulceration, even cure peptic ulcer. The effective medicine for clinical use should meet the following 2.
(9) conditions: (1) According to the intention-to-treat analysis, it can reach at least 80% of eradication rate. (2) No serious side effects. (3) The lowest possibility of inducing drug resistance of bacteria. (4) The highest possibility of patients' following doctor's advice, for example, the shortest period of treatment to reach desired effect, or minimal drugs to take (Suerbaum and Michetti, 2002). Treatment of H. pylori infection was using a sole kind of antibiotic. However, the antimicrobial activity of antibiotics was ineffectiveness under gastric juices (Hacin et al., 2008). Thus the dual or triple therapy combines the antibiotics with bismuth citrate or proton pump inhibitor was used as a golden standard. Bismuth citrate can destroy bacteria's cell wall and protect the ulcerated parts from the corrosion caused by gastric juices. Proton pump inhibitor was used to suppress acidity in the host gastric environment. Both of those were used to elevate gastric pH and enhance the effectiveness of antibiotics (Chourasia et al., 2008). The recommendation in treatment of H .pylori infection of medicine selection along with the dosage has been established in Europe (Malfertheiner P et al.,1997) and Asia-Pacific regions (Lam and Talley, 1998). According to the principle of treatments from US, Europe and Asia, all take the triple therapy as the first line medication (Jodlowski et al., 2008). The improved triple therapy adds one more kind of antibiotics. There are various prescriptions of triple therapy; the elimination rates of H. pylori are still with high variation from region to region due to antibiotic 3.
(10) resistance. The Asia-Pacific H. pylori conference hold in 1997 recommended that the first line medication of triple therapy is as follows: bismuth citrate or proton pump inhibitor (1) omeprazole 20mg, lansoprazole 30mg, pantoprazole 40mg, or rabeprazole 20mg, twice daily for one week (2) ranitidine bismuth citrate 400mg, clarithromycin 500mg, amoxicillin 1000mg or metronidazole 400mg, twice daily for a week (Wong et al., 2002). The elimination rate of H. pylori may increase by 3% to 5% if medicating for two weeks, but side effects also increase. The common side effects of triple therapy are related to the usage of antibiotics, including nausea, diarrhea, constipation and skin rashes caused by amoxicillin, or vomiting, poor sense of taste and abdomen discomfort caused by clarithromycin (Filipec Kanizaj et al., 2009). However, with long-term usage of antibiotics to treat H. pylori, the drug resistance in bacteria has been to be increased. Therefore, the latest treatment is the quadruple therapy, including two antibiotics, bismuth citrate, and a proton pump inhibitor (de Boer et al., 1995). The comparison of treatments with H. pylori was shown in Appendix 1. The antibiotics for clinical commonly use to eliminate H. pylori include amoxicillin, clarithromycin, metronidazole and tetracycline. Optimal first-line regimens consist of a proton pump inhibitor in combination with clarithromycin and either amoxicillin or metronidazole (triple therapy) for at least 7 days, but preferably 10 days. Because the most effective second-line regimens contain metronidazole, it is 4.
(11) advisable to use amoxicillin instead of metronidazole as first-line therapy in order to optimize results should subsequent therapy be required (Peek, 2004). The comparison of mechanism how these medicines work was shown in Appendix 2. The effect of metronidazole test in vivo is almost the same as in vitro. Clarithromycin also shows very high confidence level on the sensitivity test in vitro when estimating its elimination effect. Therefore, the selection of medicine may depend on the antibiotic susceptibility test. However, that was difficult to estimate the elimination effect of amoxicillin because the result highly depends on gastric environment, such as pH. Among these antibiotics, there are few reports indicate drug resistance to amoxicillin and tetracycline. It's still not discovered that these two antibiotics have the drug resistance in Taiwan (Chan and Chan, 2003). Antrodia camphorata The fruiting body of Antrodia camphorata, it’s also called ‘National treasure of Taiwan’, which is a rare and very precious medical fungus and is well known in Taiwan by name niu-chang-chih or jang-jy (Shen et al., 2003). This fungus is known only in Taiwan and is restricted to Cinnamomum kanehirai Hay (Lauraceae). The basidiomes have been used for the treatment of food and drug intoxication, diarrhea, abdominal pain, hypertension, skin itching and cancer. It’s miraculous efficacies made in human body had been investigated and researched by many Taiwanese universities, scholars and hospitals, they try to find the important components inside A. 5.
(12) camphorata through repetitive research and analysis. It is well known as a Traditional Chinese Medicine (TCM), and Taiwanese aborigines used this species to treat liver diseases and food and drug intoxication. The compounds identified in A. camphorata are predominantly polysaccharides, triterpenoids, steroids, benzenoids and maleic/succinic acid derivatives. A. camphorata only grows inside the hollow trunk of the old-aged (over 100 years) Cinnamomum kanehirai trees. The Cinnamomum kanehirai trees spread on Taiwan Mountain of 450 ~ 2000 meters elevation. Due to the magic efficacies of A. camphorata to human being’s body, the Cinnamomum kanehirai trees had been seriously cut illegally to sell for great benefit, now the Cinnamomum kanehirai tree had been protected by Taiwan government, so the price of A. camphorata in Taiwan market is fairly high nowadays. The most efficacies of A. camphorata are in its fruit body. Although there are many private organizations in Taiwan put into production of A. camphorata, the technique to cultivate the fruit body is very costly and difficult, and it also takes long. Therefore, the current cultivations of A. camphorata in the market are with liquid type or solid type; the A. camphorata cultivated by liquid type is mycelium, not the fruit body, and the components of the fruit body cultivated by solid type is only similar to the components of wild A. camphorata. Recent research has revealed that A. camphorata possesses extensive biological activity, such as hepatoprotective, 6.
(13) anti-cancer, anti-fatigue, liver protection, blood circulation improvement, immunity enhancement, antihypertensive, anti-hyperlipidemic, immuno-modulatory, anti-inflammatory and antioxidant activities (Ao et al., 2009; Chen et al., 2007; Chien et al., 2008; Hseu et al., 2008; Hsiao et al., 2003; Yang et al., 2006). The fruiting bodies and fermented products of A. camphorata have been reported to exhibit activity when treating liver diseases, such as preventing ethanol-, CCl4- and cytokine-induced liver injury, inhibiting the hepatitis B virus, ameliorating fatty liver and liver fibrosis, and inhibiting liver cancer cells (Ao et al., 2009). Despite combination of antibiotic therapy was powerful for eradication of H. pylori infection. However, the bacterial resistant rate was accompanied with increasing abuse of antibiotics in clinical. In this investigation, we sought to investigate whether chemical compounds extracted from traditional medicine-A. camphorata have the ability to inhibit H. pylori infection of gastric epithelial cells. We also investigated the effects of those isolated constituents on H. pylori-induced NF-κB activation and inflammation mediator IL-8 secretion from epithelial cells. In addition, the same time, the effects of those isolated constituents on LPS-induced nitric oxide in macrophage have been investigated.. 7.
(14) Materials and methods Bacterial strain and cell culture H. pylori 26695 (ATCC700392) reference strain was recovered from frozen stocks on Brucella agar plates (Becton Dickinson) containing 10 % sheep blood, 6 μg/ml vancomycin and 2 μg/ml amphotericin B under microaerophilic conditions for 2–3 days as described previously (Lai et al., 2006). Human gastric adenocarcinoma epithelial cell line─AGS cells (ATCC CRL 1739) were cultured in F12 (Hyclone) supplemented with 10% de-complement FBS (Hyclone). Penicillin and streptomycin (GIBCO BRL) were also added if needed. In the bacterial adherence assay, invading assay, luciferase activity assay and induction of IL-8 secretion, the cell culture medium was not supplemented with antibiotics. Determination of anti-bacterial activity The in vitro anti-bacterial activities of the constituents [dissolved in dimethylsulfoxide (DMSO, Sigma-Aldrich)] were determined by disk agar diffusion method (Lai et al., 2008b). Briefly, a total volume of 100 µl of H. pylori suspension (1 × 108 colony forming units (CFUs) /ml) was spread onto Mueller Hinton agar plates (BBL) containing 10% sheep blood. Sterile paper disks (6 mm, BBL) were placed on the agar surface with 20 µl of constituents (5mM) individually. DMSO was used as negative control and antibiotics amoxicillin (AMX, 0.05 mg/ml), clarithromycin (CLR, 0.05 mg/ml), and metronidazole (MTZ, 0.8 mg/ml) were used as positive 8.
(15) control as described previously (Lai et al., 2008b). After 72 h of incubation at 37 °C under the microaerophilic condition with humidity, the inhibition zone was determined in diameter. Cell survival assay (MTT method) The MTT assay was used to measure the ability of A. camphorata constituents AC-3, AC-10, and AC-12 to induce death of gastric epithelial cells (Rao et al., 2007). The AGS cells were exposed to various concentrations of the constituents during a 16 h incubation period. Cell viability was measured by the ability of viable cells to reduce MTT (Sigma-Aldrich) to formazan based on the ability of living cells to utilize Thiazolyl Blue and convert it into purple formazan, which absorbs light at 570 nm and could be analyzed spectrophotometrically. Measurement was performed in triplicate. The absorbance was measured using the spectrophotometer (BioRad). Inhibition of H. pylori adherence to and invasion into epithelial cells H. pylori adhesion and invasion of cultured AGS cells was done using a standard gentamicin assay as previous described (Lai et al., 2008a). A. camphorata constituents AC-3, AC-10, and AC-12 and DMSO diluted in cell culture medium were added (to reach the indicated dilutions) directly to the cell culture medium for 10 min prior to inoculation of wells with H. pylori in log-phase. AGS cells were added with H. pylori at a MOI of 50 and incubated at 37°C for 6 h. To determine the number of cell-adhesion bacteria, infected cells were washed three times to remove unbound 9.
(16) bacteria and then lysed with distilled water for 10 min. Lysates were diluted in PBS, plated onto Brucella blood agar plates and cultured for 4–5 days, after which the CFUs were counted. To determine the number of viable intracellular bacteria, infected cells were washed three times in PBS and incubated with 100 µg/ml of the membrane-impermeable antibiotic gentamicin (Sigma-Aldrich) for 1.5 h at 37 °C to remove extracellular bacteria, followed by the same procedures as above to obtain CFUs. The adhesion or invasion activity was determined as the mean of at least six experiments performed in duplicate. The controls contained H. pylori infected AGS cells without test samples were used to establish 100% adhesion or invasion. The results were expressed as the percentage of relative inhibition of H. pylori adhesion or invasion, as compared with the controls. Transient transfection of NF-κB and IL-8 reporter genes NF-κB-Luc and IL-8 (AP1+NF-κB)-Luc reporter plasmid was given from Dr. Chih-Hsin Tang (Department of Pharmacology, China Medical University) (Fong et al., 2008; Tang et al., 2007). AGS epithelium was grown to 90% confluence in 12 well–plate and transfected using Lipofectamine 2000 (Invitrogen). After 24 h incubation, transfection was complete, and cells were incubated with various concentrations of AC-3, AC-10, AC-12 and DMSO (to reach the indicated dilutions) and then infected with H. pylori strains for 6 h. To prepare cell lysates, 100 μl of reporter lysis buffer (Promega) was added to each well, and cells were scraped from 10.
(17) dishes. An equal volume of luciferase substrate was added to all samples, and luminescence was measured in a microplate luminometer. The value of luciferase activity was normalized to transfection efficiency monitored by the co-transfected β-galactosidase expression vector obtained from Promega. Measurement of interleukin-8 expression To detect interleukin–8 (IL-8) expression by gastric epithelial cells during H. pylori infection, the levels of IL-8 was measured. AGS cells were added with various concentrations of DMSO, AC-3, AC-10, and AC-12 (to reach the indicated dilutions) in cell culture medium before H. pylori infection. The treated cells then infected with H. pylori strain 26695 at a MOI of 1:100 in the presence of these agents. The supernatants were collected and stored at −80°C before analysis. The level of IL-8 in supernatants from AGS cell cultures was determined by using a sandwich enzyme-linked immunosorbent assay (ELISA) kit (R&D systems), according to the manufacturer's instruction (Lai et al., 2008a). Inhibition of macrophage activation To investigate the inhibitory effect of nitric oxide (NO) released from macrophage. Mouse macrophage RAW 264.7 cells were incubated with various concentrations of AC-3, AC-10, AC-12 and DMSO, and supplemented with lipopolysaccharide (LPS) (20μg/ml). After incubation for 20 h, equal volumes of culture supernatant and Griess reagent were mixed and incubated for 15 min at room temperature. The absorbance was 11.
(18) measured at 570 nm on a spectrophotometer (BioTek), and referred to a nitrite standard curve to determine the nitrate concentration in supernatants. Western blot for assay of iNOS expression Mouse macrophage RAW 264.7 cells were incubated with various concentrations of AC-3, AC-10 and supplemented with or without LPS (20μg/ml). Then infected cells were washed three times with PBS and added RIPA buffer (150mM NaCl , 50mM Tris base pH7.5, 1mM EDTA, 1% NP-40, 0.25% Deoxylates) on ice. After 15 min, transferred to microcentrifuge tube and centrifuged (SORVALL) for 10 min at 13000 rpm. Tweenty microliter of supernatants were took into dry bath boiling for 10 min. The samples were then subjected to 10% SDS-PAGE and transferred onto polyvinylidene difluoride membranes (Millipore). The membranes were blocked with 5% skim milk in PBS buffer containing 0.1% Tween 20 at room temperature for 1 h and then incubated overnight with iNOS antibody (Santa Cruz Biotechnology) (dilution, 1:2,000) at 4°C. The membranes were washed with PBS containing 0.1% Tween 20 and then incubated with HRP-conjugated anti-rabbit antibody (Santa Cruz Biotechnology) at a dilution of 1:5,000. The proteins of interest were visualized by using the ECLTM Western Blotting Detection Reagents and were detected by exposure the autoradiograph to X-ray films. Cell survival assay (MTT method) in macrophages The MTT assay was used to measure the ability of A. camphorata constituents AC-3, AC-10, and AC-12 to induce death of RAW 264.7 cells. The RAW 264.7 cells 12.
(19) were exposed to various concentrations of the constituents during a 16 h incubation period. Cell viability was measured by the ability of viable cells to reduce MTT (Sigma-Aldrich) to formazan based on the ability of living cells to utilize Thiazolyl Blue and convert it into purple formazan, which absorbs light at 570 nm and could be analyzed spectrophotometrically. Measurement was performed in triplicate. The absorbance was measured using the BioRad spectrophotometer. Statistical evaluation The correlation of AC-3, AC-10, AC-12 anti-bacterial adherence, invasion activity, and IL-8 secretion of treated AGS epithelial cells relative to DMSO treated control cells was determined by Student’s t-test. *P < 0.05, **P < 0.01.. 13.
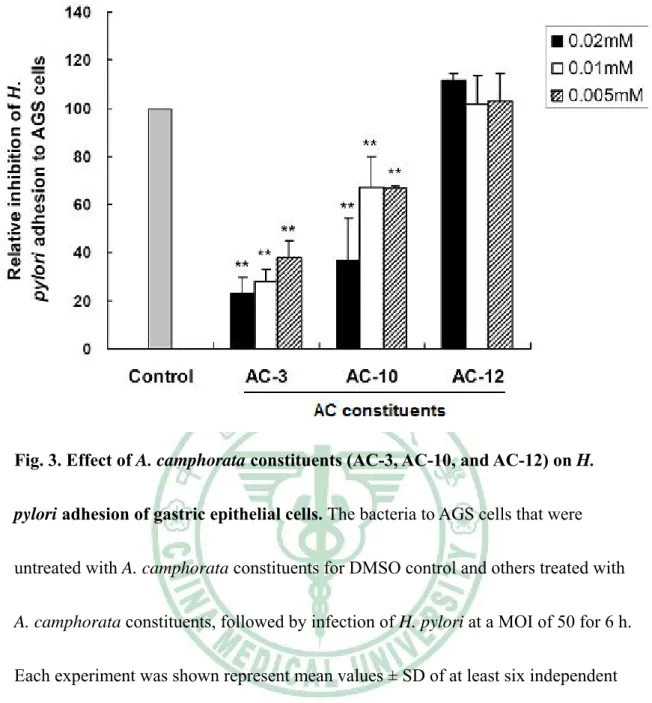
(20) Results Inhibition of H. pylori growth The anti-H. pylori activities of isolated compounds (AC-1 to 13) based on the disk agar diffusion method were shown in Table 2. DMSO used as a negative control, the constituents of AC-3, AC-10, and AC-12 were most active and inhibited H. pylori with inhibition zone of 9 to 10 mm at the concentration of 0.2mM. This effect was nearly similar to the standard drug metronidazole (MTZ), which is one of the most commonly used antibiotics for treatment of H. pylori infection (8 mm of inhibition zone at 0.8 mg/ml). This result indicates that isolation of pure constituents from the A. camphorata constituents may lead to an increase in the H. pylori inhibitory effect as expected. Therefore, the constituents of AC-3, AC-10, and AC-12 are chosen for further experiments.. Treatment of A. camphorata constituents AC-3, AC-10, and AC-12 does influence epithelial cell viability. We then analyzed how AC constituents AC-3, AC-10 and AC-12 influence the viability of gastric epithelium─AGS cells. Assays were performed to assess the cell viability using MTT method of untreated as well as test sample-treated AGS cells. As shown in Figure 2, at the minimum concentration (0.005mM) tested the constituents AC-3, AC-10 and AC-12 have no significant effect upon cell viability. This result 14.
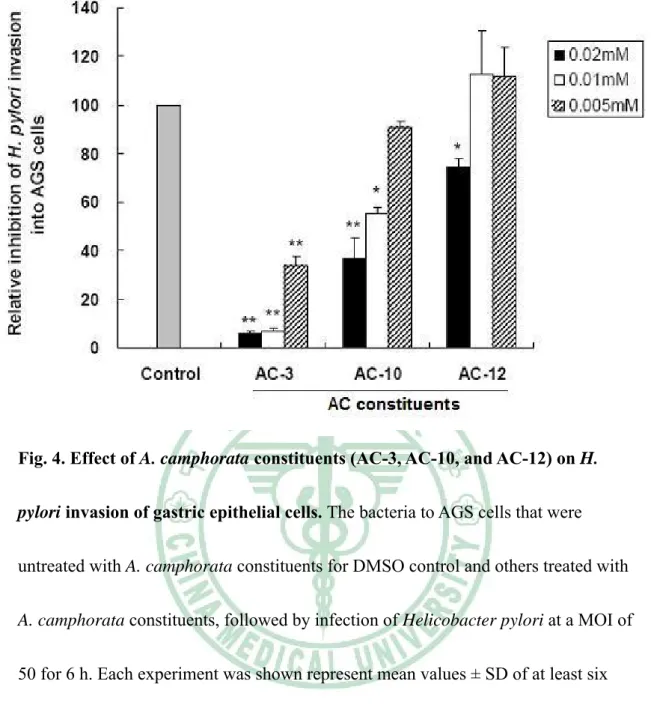
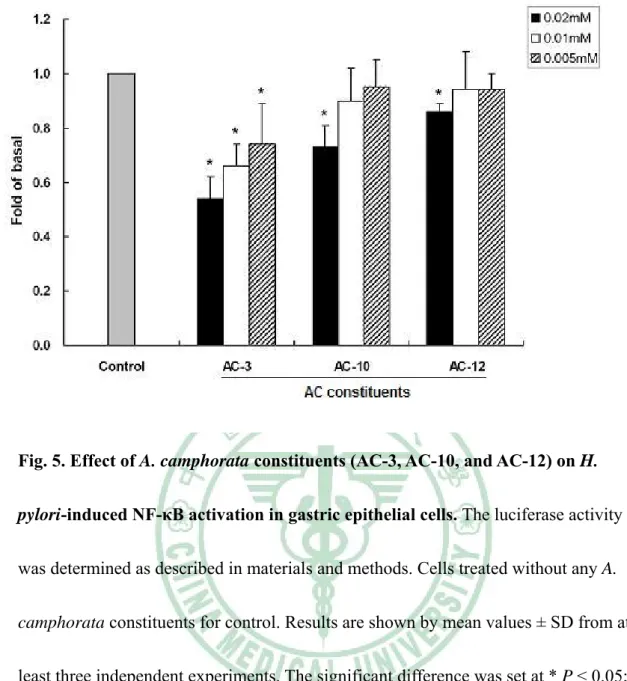
(21) indicated that constituents of AC-3, AC-10, AC-12 as well as control dissolved DMSO, have barely effected at the indicated dilutions.. Inhibition of H. pylori adherence and invasion of gastric epithelial cells Since the constituents AC-3, AC-10 and AC-12 determined to inhibit H. pylori growth at an effective level. Those constituents were further assayed with regard to their ability to inhibit the adhesion of H. pylori to AGS cells. AC-3 and AC-10 were shown to exhibit marked anti-adhesion activity against H. pylori with a concentration range from 0.005mM to 0.02mM (P < 0.01 compare to each indicated concentration of DMSO), while AC-12 was not (Figure 3). A significant inhibition of bacterial invasion into AGS cells by using AC-3 was also found with a dramatic reduction of more than 60% in a range of concentrations from 0.005mM to 0.02mM (Figure 4). Consistently, after treatment of AC-10, the bacterial invasion activity was reduced more than 40% in a range of concentration of 0.005 to 0.02mM. However, AC-12 showed no effect on inhibition of H. pylori invasion into cells. Results from this study demonstrate that AC-3 and AC-10 have their ability to inhibit bacterial adhesion and invasion of gastric epithelial cells.. AC-3 inhibits H. pylori-induced inflammation of gastric epithelial cells. Recent reports had demonstrated that H. pylori-induced IL-8 released from gastric epithelial cells is mediated activation of nuclear transcription factor NF-κB 15.
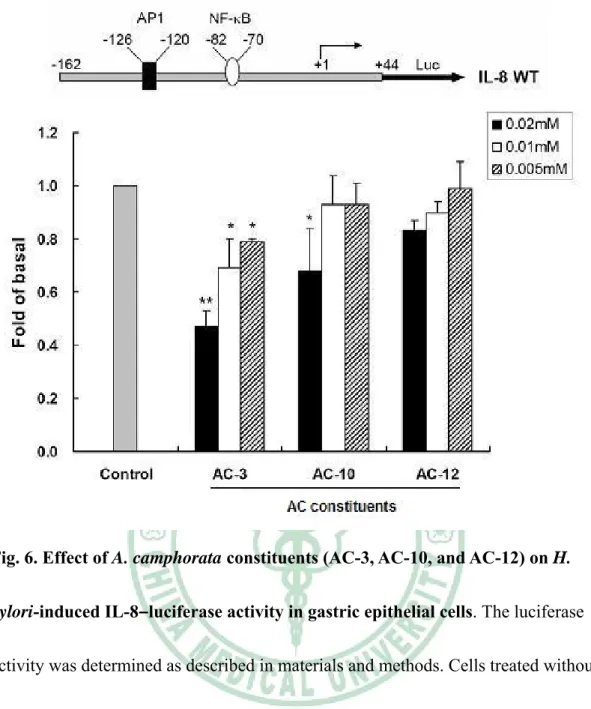
(22) (Brandt et al., 2005). Translocation of NF-κB into cell nucleus and induce activation of IL-8 transcription (Brandt et al., 2005). To examine whether the AC-3, AC-10 and AC-12 have their ability to inhibit inflammatory mediators in responses to H. pylori infection, NF-κB–luciferase and IL-8–luciferase constructs were used to determined luciferase expression following pretreatment of chemical compounds and infection with H. pylori. When AGS cells were transiently transfected with NF-κB luciferase construct, pretreatment of cells with AC-3 prior to H. pylori infection led to a dose-dependent reduction in the stimulation of luciferase activity, while only with 30% inhibition of treatment with AC-10 at a maximum concentration of 0.02mM (Figure 5). Consistent with NF-κB luciferase assay, the level of IL-8–luciferase activity by AGS cells with H. pylori infection also decreased with a dose-dependent manner after treatment of AC-3. However, only 30% reduction was found at a higher concentration of 0.02mM after treatment of AC-10 (Figure 6). In light of these results, AC-3 was demonstrated profound inhibition against H. pylori-induced NF-κB activation and down-regulation of the stimulation of IL-8 gene transcription. Results from this study might indicate that AC-3 has its ability to inhibit H. pylori infection of epithelial cells and diminished the secretion of bacterial-induced inflammatory cytokine. Thus, our results indicate that AC-3 extracted from traditional herb, A. camphorata may be developed to a new potent drug for anti-H. pylori infection of cells. 16.
(23) Treatment of A. camphorata constituents AC-3, AC-10, and AC-12 does influence macrophages viability. We then analyzed how A. camphorata constituents AC-3, AC-10 and AC-12 influence the viability of RAW 264.7 cells. Assays were performed to assess the cell viability using MTT methods of untreated as well as test sample-treated RAW 264.7 cells. As shown in Figure 7 at their minimum concentration (0.005mM) tested the constituents AC-3, AC-10 and AC-12 have no significant effect upon cell viability. This result indicated that constituents of AC-3, AC-10, AC-12, and control DMSO, have barely effected at the indicated dilutions.. A. camphorata constituents AC-3 and AC-10 inhibit LPS-mediated nitric oxide production in macrophages. To assess whether A. camphorata constituents can inhibit LPS-induced nitric oxide production in macrophages, mouse macrophage RAW 264.7 cells were cultured with LPS (20μg/ml) and A. camphorata constituents for 16h. When compare with control group, the production of nitric oxide in LPS-induced RAW 264.7 cells was suppressed by A. camphorata constituents (Figure 8). Among them, both AC-10 and AC-12 constituents showed the inhibition activity at higher concentration, while AC-3 has better inhibition effect with all concentrations. Thus, these results demonstrated that AC-3 isolate might inhibit LPS-induced nitric oxide production in mouse 17.
(24) macrophage RAW 264.7 cells.. Western blot of inhibition of iNOS production. To investigate the effects of A. camphorata constituents on the cytosolic protein levels of iNOS, RAW 264.7 cells were treated with or without LPS and various concentrations of A. camphorata constituents for 16h and the protein levels of iNOS were analyzed by Western blot. As shown in Figure 9, AC-3 and AC-10 constituents show inhibition effects of iNOS protein expression. Only AC-3 isolate shows higher inhibition effect at treated with maximus concentration (0.02mM). This result indicated that A. camphorata constituents might inhibit LPS-induced nitric oxide production in macrophages.. 18.
(25) Discussion and conclusion There are many kinds of A. camphorata products in market, and in this study we use pure chemical constituents of A. camphorata. Our results showed that A. camphorata constituents have their effects on the inhibition of H. pylori-induced inflammation. The effects of A. camphorata constituents on anti-bacteria activity were similar to the standard drug metronidazole (MTZ). In addition, the A. camphorata constituents showed no cytotoxic effect on both gastric epithelial cells and macrophage RAW 264.7 cells.. Among lots of A. camphorata commodities in the world, most of them are used for liver protection, for instance, in vitro or in vivo treatment of ethanol-induced hepatis or hepatitis B virus infection in a murine model system (Lee et al., 2002; Lu et al., 2007). Using A. camphorata compounds for treatment to bacteria is unusual, however, it seems to be a good direction for eradication of bacterial infection. In this study, our results show that constituents of AC-3 and AC-10 have better inhibition effects on H. pylori-induced inflammation of gastric epithelial cells. This was consistent with previous study of anti-inflammatory and anti-cancer properties of A. camphorata (Rao et al., 2007).. In a macrophage RAW 264.7 cell system, A. camphorata constituents show their inhibitory effects by decreasing the production of nitric oxide. Although all the A. 19.
(26) camphorata constituents have the same trends, AC-3 shows the best effect than others. Our data also showed that iNOS expression in LPS-induced macrophage was inhibited by AC-3. In addition, MTT assay shows that all A. camphorata constituents have no cytotoxic in macrophage RAW 264.7 cells. These results maybe indicated that A. camphorata constituents would be a good candidate for therapy of inflammatory diseases. In this study, we demonstrated that A. camphorata constituents displayed potential anti-H. pylori activity. The constituents of AC-3 and AC-10 inhibit the H. pylori adhesion and invasion to AGS cells. In addition, these constituents also inhibit the IL-8 protein secretion and NF-κB signal activation in H. pylori-infected gastric epithelial cells. This inhibition of NF-κB activation and IL-8 secretion by AC-3 and AC-10 might contribute to the potential role of A. camphorata as an anti-inflammatory action in H. pylori induced gastric epithelial cells damage. Future studies are needed to clarify the molecular mechanisms by which A. camphorata inhibits H. pylori-mediated activation of NF-κB expression in AGS cells and to identify additional targets in gene regulation.. For the future, first we can do T cell experiment like test spleen cells by ex vivo mouse model system to examine if AC constituents can inhibit mitogen (conA)-induce T cell activation or not. Otherwise detect AC constituents inhibit TCR. 20.
(27) (T cell receptor) induced by Jurkat cells activation in using Flow cytometry and cell cycle assay. 21.
(28) References Ao, Z.H., Xu, Z.H., Lu, Z.M., Xu, H.Y., Zhang, X.M. and Dou, W.F., 2009. Niuchangchih (Antrodia camphorata) and its potential in treating liver diseases. J Ethnopharmacol 121, 194-212. Bochenek, W.J., Peters, S., Fraga, P.D., Wang, W., Mack, M.E., Osato, M.S., El-Zimaity, H.M., Davis, K.D. and Graham, D.Y., 2003. Eradication of Helicobacter pylori by 7-day triple-therapy regimens combining pantoprazole with clarithromycin, metronidazole, or amoxicillin in patients with peptic ulcer disease: results of two double-blind, randomized studies. Helicobacter 8, 626-642. Brandt, S., Kwok, T., Hartig, R., Konig, W. and Backert, S., 2005. NF-kappaB activation and potentiation of proinflammatory responses by the Helicobacter pylori CagA protein. Proc Natl Acad Sci U S A 102, 9300-9305. Chan, Y. and Chan, C.H., 2003. Antibiotic resistance of pathogenic bacteria from odontogenic infections in Taiwan. J Microbiol Immunol Infect 36, 105-110. Chen, K.C., Peng, C.C., Peng, R.Y., Su, C.H., Chiang, H.S., Yan, J.H. and Hsieh-Li, H.M., 2007. Unique formosan mushroom Antrodia camphorata differentially inhibits androgen-responsive LNCaP and -independent PC-3 prostate cancer cells. Nutr Cancer 57, 111-121. Chien, S.C., Chen, M.L., Kuo, H.T., Tsai, Y.C., Lin, B.F. and Kuo, Y.H., 2008. 22.
(29) Anti-inflammatory activities of new succinic and maleic derivatives from the fruiting body of Antrodia camphorata. J Agric Food Chem 56, 7017-7022. Chourasia, D., Misra, A., Pandey, R. and Ghoshal, U.C., 2008. Gastric atrophy and intestinal metaplasia in a patient on long-term proton pump inhibitor therapy. Trop Gastroenterol 29, 172-174. de Boer, W.A., Driessen, W.M., Jansz, A.R. and Tytgat, G.N., 1995. Quadruple therapy compared with dual therapy for eradication of Helicobacter pylori in ulcer patients: results of a randomized prospective single-centre study. Eur J Gastroenterol Hepatol 7, 1189-1194. de Boer, W.A., Driessen, W.M., Potters, V.P. and Tytgat, G.N., 1994. Randomized study comparing 1 with 2 weeks of quadruple therapy for eradicating Helicobacter pylori. Am J Gastroenterol 89, 1993-1997. Filipec Kanizaj, T., Katicic, M., Skurla, B., Ticak, M., Plecko, V. and Kalenic, S., 2009. Helicobacter pylori eradication therapy success regarding different treatment period based on clarithromycin or metronidazole triple-therapy regimens. Helicobacter 14, 29-35. Fong, Y.C., Maa, M.C., Tsai, F.J., Chen, W.C., Lin, J.G., Jeng, L.B., Yang, R.S., Fu, W.M. and Tang, C.H., 2008. Osteoblast-derived TGF-beta1 stimulates IL-8 release through AP-1 and NF-kappaB in human cancer cells. J Bone Miner Res 23, 961-970. 23.
(30) Gobert, A.P., McGee, D.J., Akhtar, M., Mendz, G.L., Newton, J.C., Cheng, Y., Mobley, H.L. and Wilson, K.T., 2001. Helicobacter pylori arginase inhibits nitric oxide production by eukaryotic cells: a strategy for bacterial survival. Proc Natl Acad Sci U S A 98, 13844-13849. Graham, D.Y., Lu, H. and Yamaoka, Y., 2009. African, Asian or Indian enigma, the East Asian Helicobacter pylori: facts or medical myths. J Dig Dis 10, 77-84. Gu, Q., Xia, H.H., Wang, J.D., Wong, W.M., Chan, A.O., Lai, K.C., Chan, C.K., Yuen, M.F., Fung, F.M., Wong, K.W., Lam, S.K. and Wong, B.C., 2006. Update on clarithromycin resistance in Helicobacter pylori in Hong Kong and its effect on clarithromycin-based triple therapy. Digestion 73, 101-106. Hacin, B., Rogelj, I. and Matijasic, B.B., 2008. Lactobacillus isolates from weaned piglets' mucosa with inhibitory activity against common porcine pathogens. Folia Microbiol (Praha) 53, 569-576. Harris, P.R., Smythies, L.E., Smith, P.D. and Dubois, A., 2000. Inflammatory cytokine mRNA expression during early and persistent Helicobacter pylori infection in nonhuman primates. J Infect Dis 181, 783-786. Hentschel, E., Brandstatter, G., Dragosics, B., Hirschl, A.M., Nemec, H., Schutze, K., Taufer, M. and Wurzer, H., 1993. Effect of ranitidine and amoxicillin plus metronidazole on the eradication of Helicobacter pylori and the recurrence of duodenal ulcer. N Engl J Med 328, 308-312. 24.
(31) Hseu, Y.C., Chen, S.C., Yech, Y.J., Wang, L. and Yang, H.L., 2008. Antioxidant activity of Antrodia camphorata on free radical-induced endothelial cell damage. J Ethnopharmacol 118, 237-245. Hsiao, G., Shen, M.Y., Lin, K.H., Lan, M.H., Wu, L.Y., Chou, D.S., Lin, C.H., Su, C.H. and Sheu, J.R., 2003. Antioxidative and hepatoprotective effects of Antrodia camphorata extract. J Agric Food Chem 51, 3302-3308. Hsu, P.I., Lai, K.H., Lin, C.K., Chen, W.C., Yu, H.C., Cheng, J.S., Tsay, F.W., Wu, C.J., Lo, C.C., Tseng, H.H., Yamaoka, Y., Chen, J.L. and Lo, G.H., 2005. A prospective randomized trial of esomeprazole- versus pantoprazole-based triple therapy for Helicobacter pylori eradication. Am J Gastroenterol 100, 2387-2392. Jodlowski, T.Z., Lam, S. and Ashby, C.R., Jr., 2008. Emerging therapies for the treatment of Helicobacter pylori infections. Ann Pharmacother 42, 1621-1639. Kim, S.Y., Lee, Y.C., Kim, H.K. and Blaser, M.J., 2006. Helicobacter pylori CagA transfection of gastric epithelial cells induces interleukin-8. Cell Microbiol 8, 97-106. Lai, C.H., Chang, Y.C., Du, S.Y., Wang, H.J., Kuo, C.H., Fang, S.H., Fu, H.W., Lin, H.H., Chiang, A.S. and Wang, W.C., 2008a. Cholesterol depletion reduces Helicobacter pylori CagA translocation and CagA-induced responses in AGS cells. Infect Immun 76, 3293-3303. Lai, C.H., Fang, S.H., Rao, Y.K., Geethangili, M., Tang, C.H., Lin, Y.J., Hung, C.H., 25.
(32) Wang, W.C. and Tzeng, Y.M., 2008b. Inhibition of Helicobacter pylori-induced inflammation in human gastric epithelial AGS cells by Phyllanthus urinaria extracts. J Ethnopharmacol 118, 522-526. Lai, C.H., Kuo, C.H., Chen, P.Y., Poon, S.K., Chang, C.S. and Wang, W.C., 2006. Association of antibiotic resistance and higher internalization activity in resistant Helicobacter pylori isolates. J Antimicrob Chemother 57, 466-471. Lam, S.K. and Talley, N.J., 1998. Report of the 1997 Asia Pacific Consensus Conference on the management of Helicobacter pylori infection. J Gastroenterol Hepatol 13, 1-12. Lee, I.H., Huang, R.L., Chen, C.T., Chen, H.C., Hsu, W.C. and Lu, M.K., 2002. Antrodia camphorata polysaccharides exhibit anti-hepatitis B virus effects. FEMS Microbiol Lett 209, 63-67. Logan, R.P., Gummett, P.A., Misiewicz, J.J., Karim, Q.N., Walker, M.M. and Baron, J.H., 1991. One week eradication regimen for Helicobacter pylori. Lancet 338, 1249-1252. Lu, Z.M., Tao, W.Y., Zou, X.L., Fu, H.Z. and Ao, Z.H., 2007. Protective effects of mycelia of Antrodia camphorata and Armillariella tabescens in submerged culture against ethanol-induced hepatic toxicity in rats. J Ethnopharmacol 110, 160-164. Marshall, B., 2002. Helicobacter pylori: 20 years on. Clin Med 2, 147-152. 26.
(33) McMahon, B.J., Hennessy, T.W., Bensler, J.M., Bruden, D.L., Parkinson, A.J., Morris, J.M., Reasonover, A.L., Hurlburt, D.A., Bruce, M.G., Sacco, F. and Butler, J.C., 2003. The relationship among previous antimicrobial use, antimicrobial resistance, and treatment outcomes for Helicobacter pylori infections. Ann Intern Med 139, 463-469. Megraud, F. and Lamouliatte, H., 2003. Review article: the treatment of refractory Helicobacter pylori infection. Aliment Pharmacol Ther 17, 1333-1343. Mirbagheri, S.A., Hasibi, M., Abouzari, M. and Rashidi, A., 2006. Triple, standard quadruple and ampicillin-sulbactam-based quadruple therapies for H. pylori eradication: a comparative three-armed randomized clinical trial. World J Gastroenterol 12, 4888-4891. Nozawa, Y., Nishihara, K., Peek, R.M., Nakano, M., Uji, T., Ajioka, H., Matsuura, N. and Miyake, H., 2002. Identification of a signaling cascade for interleukin-8 production by Helicobacter pylori in human gastric epithelial cells. Biochem Pharmacol 64, 21-30. Parsonnet, J., 1998. Helicobacter pylori. Infect Dis Clin North Am 12, 185-197. Peek, R.M., 2004. Helicobacter pylori and Gastroesophageal Reflux Disease. Curr Treat Options Gastroenterol 7, 59-70. Poon, S.K., Chang, C.S., Su, J., Lai, C.H., Yang, C.C., Chen, G.H. and Wang, W.C., 2002. Primary resistance to antibiotics and its clinical impact on the efficacy of 27.
(34) Helicobacter pylori lansoprazole-based triple therapies. Aliment Pharmacol Ther 16, 291-296. Rao, Y.K., Fang, S.H. and Tzeng, Y.M., 2007. Evaluation of the anti-inflammatory and anti-proliferation tumoral cells activities of Antrodia camphorata, Cordyceps sinensis, and Cinnamomum osmophloeum bark extracts. J Ethnopharmacol 114, 78-85. Salcedo, J.A. and Al-Kawas, F., 1998. Treatment of Helicobacter pylori infection. Arch Intern Med 158, 842-851. Sartor, R.B., 1994. Cytokines in intestinal inflammation: pathophysiological and clinical considerations. Gastroenterology 106, 533-539. Shen, C.C., Kuo, Y.C. and Huang, R.L., 2003. New ergostane and lanostane from Antrodia camphorata. J Chin Med 14, 247-258. Suerbaum, S. and Michetti, P., 2002. Helicobacter pylori infection. N Engl J Med 347, 1175-1186. Tang, C.H., Yang, R.S., Chen, Y.F. and Fu, W.M., 2007. Basic fibroblast growth factor stimulates fibronectin expression through phospholipase C gamma, protein kinase C alpha, c-Src, NF-kappaB, and p300 pathway in osteoblasts. J Cell Physiol 211, 45-55. Tankovic, J., Lamarque, D., Lascols, C., Soussy, C.J. and Delchier, J.C., 2001a. Clarithromycin resistance of Helicobacter pylori has a major impact on the 28.
(35) efficacy of the omeprazole-amoxicillin-clarithromycin therapy. Pathol Biol (Paris) 49, 528-533. Tankovic, J., Lamarque, D., Lascols, C., Soussy, C.J. and Delchier, J.C., 2001b. Impact of Helicobacter pylori resistance to clarithromycin on the efficacy of the omeprazole-amoxicillin-clarithromycin therapy. Aliment Pharmacol Ther 15, 707-713. Walsh, C., 2000. Molecular mechanisms that confer antibacterial drug resistance. Nature 406, 775-781. Walsh, J.H. and Peterson, W.L., 1995. The treatment of Helicobacter pylori infection in the management of peptic ulcer disease. N Engl J Med 333, 984-991. Warren, J.R., 1983. Unidentified curved bacilli on gastric epithelium in active chronic gastritis. Lancet i, 1273. Wong, W.M., Wong, B.C., Lu, H., Gu, Q., Yin, Y., Wang, W.H., Fung, F.M., Lai, K.C., Xia, H.H., Xiao, S.D. and Lam, S.K., 2002. One-week omeprazole, furazolidone and amoxicillin rescue therapy after failure of Helicobacter pylori eradication with standard triple therapies. Aliment Pharmacol Ther 16, 793-798. Yamaoka, Y., Kita, M., Kodama, T., Sawai, N. and Imanishi, J., 1996. Helicobacter pylori cagA gene and expression of cytokine messenger RNA in gastric mucosa. Gastroenterology 110, 1744-1752. Yang, H.L., Chen, C.S., Chang, W.H., Lu, F.J., Lai, Y.C., Chen, C.C., Hseu, T.H., Kuo, 29.
(36) C.T. and Hseu, Y.C., 2006. Growth inhibition and induction of apoptosis in MCF-7 breast cancer cells by Antrodia camphorata. Cancer Lett 231, 215-227. Zullo, A., Gatta, L., De Francesco, V., Hassan, C., Ricci, C., Bernabucci, V., Cavina, M., Ierardi, E., Morini, S. and Vaira, D., 2005. High rate of Helicobacter pylori eradication with sequential therapy in elderly patients with peptic ulcer: a prospective controlled study. Aliment Pharmacol Ther 21, 1419-1424.. 30.
(37) Table 1 Molecular weight of A. camphorata constituents Compound. Molecular weight (M.W). AC-1. 482. AC-2. 468. AC-3. 526. AC-4. 470. AC-5. 468. AC-6. 484. AC-7. 486. AC-8. 466. AC-9. 454. AC-10. 470. 31.
(38) Table 2 Inhibitory effects of A. camphorata constituents. †. Compounds. Inhibition zone (mm). AC-1. -. AC-2. 8. AC-3. 10. AC-4. -. AC-5. -. AC-6. -. AC-7. -. AC-8. -. AC-9. 7. AC-10. 9. AC-11. -. AC-12. 10. AC-13. -. AMX†. 22. CLR. 28. MTZ. 12. DMSO was used as negative control, and the antibiotics amoxicillin (AMX,. 0.05 mg/ml), clarithromycin (CLR, 0.05 mg/ml), and metronidazole (MTZ, 0.8 mg/ml) were used as positive control.. 32.
(39) 28. O. COOCH3 25. 24 17. 11 14. O 3. O. 6. Methyl antcinate B (1). 28 HOOC. 25. 24 17. 11 29. HO 3. 6. Dehydroeburicoic acid (2). 28 HOOC. 25. 17. 11 29. HO 3. 24. OAc. 6. 15α-Acetyldehydrosulphurenic acid (3). HOOC. 25. 17. 11. HO 3. 24. 28. OH. 6. 3β,15α-Dihydroxylanosta-7,9(11),24triene-21-oic acid (4). Fig. 1. Chemiacl structures of A. camphorata constituents 1-11. 33.
(40) 28. O. 24. 20 17. 11. 26 COOH 25. 14 O 3. O. 6. Antcin B (or) Zhankuic acid A (5). 28 HOOC. 25. 17. 11. HO 3. 24. 29. OH. 6. Dehydrosulphurenic acid (6). 28. OH O 11 HO 3. 6. 24. COOH. 17. O. Zhankuic acid C (7). 28. HOOC 11. 24 17. OH HO 3. 6. Sulphurenic acid (8). Fig. 1. Chemiacl structures of A. camphorata constituents 1-11 (continuous). 34.
(41) 28. O 1. 22. 21 18. 17 16. 19 11 9. 3. COOH. 7. O. Antcin A (9). 28 21 18. O 1 3. 19 11 9. O. 7. 26. 22. COOH. 17 16. OH. Antcin C (10). 28. O. 21 18. 19 11 1 9 3 7. O. 22. COOCH3. 17 16. Methyl antcinate A (11). Fig. 1. Chemiacl structures of A. camphorata constituents 1-11 (continuous). 35.
(42) Fig. 2. Effect of A. camphorata constituents (AC-3, AC-10, and AC-12) and DMSO control on gastric epithelial cell viability. The AGS cells were exposed to various concentrations of the constituents during a 20 h incubation period and treated with no constituents for control. Cell viability was assessed by MTT assay. Results represent mean values ± SD from at least triplicate independent experiments.. 36.
(43) Fig. 3. Effect of A. camphorata constituents (AC-3, AC-10, and AC-12) on H. pylori adhesion of gastric epithelial cells. The bacteria to AGS cells that were untreated with A. camphorata constituents for DMSO control and others treated with A. camphorata constituents, followed by infection of H. pylori at a MOI of 50 for 6 h. Each experiment was shown represent mean values ± SD of at least six independent experiments. The significant difference was set at * P < 0.05; ** P < 0.01.. 37.
(44) Fig. 4. Effect of A. camphorata constituents (AC-3, AC-10, and AC-12) on H. pylori invasion of gastric epithelial cells. The bacteria to AGS cells that were untreated with A. camphorata constituents for DMSO control and others treated with A. camphorata constituents, followed by infection of Helicobacter pylori at a MOI of 50 for 6 h. Each experiment was shown represent mean values ± SD of at least six independent experiments. The significant difference was set at * P < 0.05; ** P < 0.01.. 38.
(45) Fig. 5. Effect of A. camphorata constituents (AC-3, AC-10, and AC-12) on H. pylori-induced NF-κB activation in gastric epithelial cells. The luciferase activity was determined as described in materials and methods. Cells treated without any A. camphorata constituents for control. Results are shown by mean values ± SD from at least three independent experiments. The significant difference was set at * P < 0.05; ** P < 0.01.. 39.
(46) Fig. 6. Effect of A. camphorata constituents (AC-3, AC-10, and AC-12) on H. pylori-induced IL-8–luciferase activity in gastric epithelial cells. The luciferase activity was determined as described in materials and methods. Cells treated without any A. camphorata constituents for control. Results are shown by mean values ± SD from at least three independent experiments. The significant difference was set at * P < 0.05; ** P < 0.01.. 40.
(47) Fig. 7. Effect of A. camphorata constituents (AC-3, AC-10, and AC-12) and DMSO control in RAW 264.7 cells. The RAW 264.7 cells were treated with or without LPS (20μg/ml) and others treated with both LPS and A. camphorata constituents or DMSO control during a 20 h incubation period. Cell viability was assessed by MTT assay. Results represent mean values ± SD from at least triplicate independent experiments.. 41.
(48) Fig. 8. Inhibition by A. camphorata constituents (AC-3, AC-10, and AC-12) and DMSO control of LPS-induced NO production in RAW 264.7 cells. Cells were treated with or without LPS (20μg/ml) and others treated with both LPS and A. camphorata constituents or DMSO control. After incubation over 20 h, the culture supernatant of RAW 264.7 cell line was then collected for the assay of nitric oxide production, respectively. Each experiment was shown represent mean values ± SD of at least six independent experiments. The significant difference was set at * P < 0.05; ** P < 0.01.. 42.
(49) Fig. 9. Effects of A. camphorata constituents (AC-3 and AC-10) on the iNOS protein expression in RAW 264.7 cells. Cells were treated with or without LPS (20μg/ml) and others treated with both LPS and A. camphorata constituents. After 20 h, cells were washed with PBS and homogenized. Each lane contained protein 20μg constituents. Electrophoresis was performed using 10% SDS-PAGE and detected with anti-iNOS and anti-β-actin antibodies, respectively.. 43.
(50) Appendix 1 Therapy of H. pylori infection Therapy Single therapy. Dose Use bismuth or single antibiotic. Illustrate Single antibiotic does not have good effect for their antibiotic. Dual therapy. (1) Bismuth and one antibiotic. Omeprazole can make. (2) Proton pump inhibitor and amoxicillin. the pH value of stomach. or clarithromycin. increase and then enhance the effect of antibiotic. Triple therapy. (1) Bismuth with one of tetracycline and amoxicillin , and one of metronidazole. therapy has higher side. and clarithromycin. effect, and proton pump. (2) Proton pump inhibitor and two antibiotics Quadruple therapy. Bismuth-based triple. inhibitor-based triple therapy is more popular. Proton pump inhibitor , bismuth and two. Best effect. antibiotics (de Boer et al., 1994). 44.
(51) Appendix 2 Drugs used in H. pylori therapy Drugs Amoxicillin. Classify. Mechanism. Penicillin. To interfere compound, inhibit bacteria cell wall compound. Clarithromycin. Macrolide. To form a reversible complex with 50S ribosomal subunits, and inhibit bacteria to synthesize protein. Metronidazole. Nitroimidazole. To inhibit bacteria’s nitroreductase. Bismuth. To destroy the completeness of bacteria cell wall, interfere bacteria to adhere to gastric epithelium cell and inhibit bacteria’s urease and phospholipase. Proton pump. To inhibit host cell secrete stomach acid. inhibitor (Salcedo and Al-Kawas, 1998; Walsh, 2000; Walsh and Peterson, 1995). 45.
(52)
數據



+7
相關文件
The functional fall of the salivary secretion observed at 180 days post irradiation was not only associated with a reduction of gland mass but also to an alteration of the
(E) Varying numbers of tumour cells expressing cancer antigen 125 are present in the lining of tubules (objective magnification ¥10).. The arrowed cells show strong nuclear
Histologically VX is distinguished from other lesions by presence of large numbers of foam cells, and limited to the connective tissue papillae.. The foam cells on
pylori in pathogenesis of oral mucosal lesions or ulcerations is still unclear, it seems that patients with oral lesions as leukoplakia and oral lichen planus, and concurrent
-- the tumor odontogenic epithelial cells usually formed small nests and thin strands that were positive for keratin and AE1 plus AE3. -- 5 cases showed mild to moderate
Stain (-) because the main body of the tumor consisted of epithelioid or epithelial cells that contained few positive smooth muscle cells. No clear ductal structures or
This is the first reported case of successful treatment after curative radiation therapy for maxillary AC.. Ó 2018 American Association of Oral and Maxillofacial Surgeons J
Higher immunoexpression of HIF-1 a, NOTCH1, ADAM-12, and heparin-binding epidermal growth factor like growth factor (HB-EGF) in epidermoid cells in compari- son with mucous cells
