行政院國家科學委員會專題研究計畫成果報告
葛根素對大鼠體溫調節作用及下視丘五羥色胺酸
濃度變化之影響
Effects of Puer ar in on Ther mor egulator y Response
and Hypothalamic Ser otonin Release in Rats
計畫編號:NSC 89-2314-B-039-005
執行期限:88 年 08 月 01 日至 89 年 07 月 31 日
主持人:闕甫
執行機構及單位名稱:中國醫藥學院中國藥學研究所
一、中文摘要 葛根素 (puerarin) 為野葛 (Pueraria lobata) 主 要 活 性 成 分 之 一 , 已 知 為 β -adren-ergic receptor 阻斷劑,具有解痙、抗 心率不整、降血壓等作用,此外還具有解 熱等作用,但有關 puerarin 降溫作用之機 轉至今仍並未見有研究報告提出;因此本 研究擬就 puerarin 對清醒大鼠之降溫作 用機轉進行探討。研究結果顯示,由側腦 室 給 予 puerarin (100µg/kg, intracerebro-ventricular injection; i.c.v.) 可引起室溫下(24±1℃) 正常清醒大鼠體溫降低作用,且
腹 腔 給 予 puerarin (5-30mg/kg, intraperi-toneal admi-nistration; i.p.) 可引起一具劑 量依存性之體溫降低作用,並同步降低大 鼠下視丘 serotonin (5-HT) 之濃度;此降 溫作用會因側腦室給予 serotonin neuro-toxin 5,7-dihydroxytryptamine (5,7-DHT, 200µg/10µl, i.c.v.) 或皮下給予5-HT1A 受 體之拮抗劑 (-)-pindolol (0.05, 0.5mg/kg, subcutaneously injection; s.c.)所減弱,但可 被 皮 下 給 予 5-HT1A 受 體 之 致 效 劑 8-hydroxy-dipropylaminotetralin (8-OH-DPAT; 0.05 mg/kg; s.c.) 所加強。此外,puerarin 誘 發之降溫作用亦會被5-HT2 受體之致效劑 (±)-2,5-dimethoxy-4-iodoamphetamine (DOI; 5, 10µg/10µl; i.c.v.; 0.5, 1mg/kg; i.p.) 、 quipazine (0.1, 1mg/kg; i.p.) 所拮抗,或為 5-HT2 受 體 之 拮 抗 劑 pirenperone (0.2mg/kg; s.c.)、ketanserin (1mg/kg; i.p.) 所 加強。由此可推知 puerarin 可能藉由降低 下 視 丘 serotonin 之 濃 度 , 並 作 用 於 postsynaptic serotonin 受體,致活 5-HT1A 受體及阻斷 5-HT2 受體,而達到降低體溫 之作用。 關鍵詞:葛根素、降溫作用、五羥色胺酸 Abstr act
Puerarin is an isoflavone compound isolated from Pueraria lobata. Puerarin, a β -adrenergic receptor blocker, possesses anti-convulsive, antiarrhythmic and antihyper-tension effects. It also reduces 2,4-dinitro-phenol-induced hyperthermia. However, the effects of puerarin on normal body tempe-rature are unknown. On this account, in the present study, experiments were carried out to assess the mechanism of puerarin induced-hypothermia in unanesthetized rats.
Puerarin (100µg/10µl, i.c.v.; 5-30mg/kg, i.p.) caused a dose-related fall in both colonic temperature and the 5-HT release in the hypothalamus at room temperature. Puerarin induced hypothermia was attenuated by pretreatment with 5,7-dihydroxytryptamine (5,7-DHT; a serotonin neurotoxin, 200
µg/10µl; i.c.v., one week ago), or (-)-pindolol (a 5-HTIA/β adrenoceptor antagonist; 0.05, 0.5mg/kg; s.c.) but potentiated by (±)-8– hydroxydiopropylaminoteralin ( 8-OH-DPAT; a 5-HT1A receptor agonist; 0.05mg/kg; s.c.).
In addition, the puerarin induced
hypothermia was attenuated by (±
)-2,5-dimethoxy-4-iodoamphetamine (DOI; 5-HT2
receptor agonist; 5, 10µg/10µl; i.c.v.; 0.5, 1mg/kg; i.p.) or quipazine (a 5-HT2 receptor agonist; 0.5, 1mg/kg; i.p.), but potentiated by
ketanserin (5-HT2 receptor antagonist;
1mg/kg; i.p.) or pirenperone (5-HT2 receptor antagonist; 0.2mg/kg; s.c.). These results
† 八十六年度及以前的一般 國科會專題計畫(不含產學 合作研究計畫)亦可選擇適 用,惟較特殊的計畫如國科 會規劃案等,請先洽得國科 會各學術處同意。
indicate that puerarin may act through 5-HT1A receptor activation or 5-HT2 receptor antagonism within the brain to induce its hypothermia.
Keywor ds: Puerarin, Hypothermia,
Hypothalamus, Serotonin 二、緣由與目的 葛根素 (puerarin) 為葛根之主要成分 之一,屬 isoflavonoids;近年來藥理研究 發現其為一β-adrenoreceptor 之阻斷劑(1), 對腦內之 benzodiazepine receptor 亦具致效 作用(2);此外 puerarin 還具有解酒(3)、解痙 (4) 、促進腦及心臟血流(5-8),降血壓(9,10)、抗 心率不整(11)、降低 2,4-dinitrophenol 誘發之 高溫(12)等作用。然有關 puerarin 解熱降溫 作用之機轉至今仍並未見有研究報告提 出,本實驗室於八十八年度年度國科會專 題研究計畫中,首先就 puerarin 對熱原性 發燒大鼠之解熱作用進行探討,初步研究 結果顯示室溫下 puerarin (20mg/kg, i.p.) 除 可降低細菌內毒素 lipoplysaccharide (LPS) 誘發高溫大鼠之直腸溫度(肛溫)外,亦 可明顯降低正常大鼠之直腸溫度。
1957 年 Brodie & Shore 與 1961 年 Von Euler 先後提出下視丘 可能在體溫調 節作用上扮演一控制的角色(13),及腦內單 胺 神 經 系 統 可 能 參 與 下 視 丘 體 溫 恆 定 (thermostat) 設定的機制後(14), 於 1980 年 Myers 等學者發現下視丘 serotonin (5-HT) 之 含 量 與 熱 的 產 生 有 關(15), 1991 年 Gorden 進一步報告指出當動物下視丘之 5-HT 活性增加時可使代謝產熱增加散熱減 少 (表皮血管收縮),反之,當動物下視丘 5-HT 活性降低會使代謝產熱減少散熱增 加 (表皮血管擴張,呼吸散熱率增加)(16), 可知下視丘之 serotoninergic system 在體 溫調節系統中扮演一重要之角色。因此本 研究擬繼續就 puerarin 對室溫下正常大 鼠 直 腸 溫 度 變 化 及 其 與 下 視 丘 serotoninergic system 之關係進行探討。 三、結果與討論 實驗結果發現腹腔給予 puerarin (5-30mg/kg) 對室溫下正常大鼠之體溫具明顯 之降溫作用,給藥後約 120 分鐘體溫降至 最低,其後則慢慢回復,持續時間則會隨劑 量 增 加 而 延 長 (Fig.1) , 而 側 腦 室 給 予 puerarin (100µg/rat) 對室溫下正常大鼠之 體溫也具明顯之降溫作用(Fig.2)。 為探討 puerarin 之降溫作用與體溫 調節中樞 (下視丘) serotoninergic system 間 之關係,本研究利用微透析法配合高壓液相 層析儀,對 puerarin 對於室溫下正常清醒 大鼠肛溫與下視丘 5-HT 濃度變化進行同 步偵測,實驗結果發現當 puerarin 降低大 鼠肛溫時,其下視丘 5-HT 之濃度亦有降低 之現象 (Table 1);故 puerarin 之降溫作用 可能與降低下視丘 5-HT 之濃度有關。 5,7-dihydroxytryptamine (5,7-DHT) 為 5-HT neurons之 neurotoxin(17-19),是神經科 學研究上常用之工具藥,由大鼠側腦室一次 給予 5,7-DHT,七天後可明顯降低下視丘 5-HT 濃度,但不會影響下視丘 dopamine 之濃度,且對大鼠之體溫並無明顯之影響 (20)。 由 於 puerarin 之 降 溫 作 用 , 會 因 5,7-DHT 破壞腦中 5-HT neurons 而減弱 (Table 2) ; 可 推 知 突 觸 後 serotoninergic receptors 亦可能參與 puerarin 降溫作用之 產生。 目前已知腦中至少存在有七種不同之 serotoninergic receptor (5-HT1A/B/D/E/F, 5-HT2/A/B/C, 5-HT3, 5-HT4, 5-HT5, 5-HT6 及 5-HT7)(21)。 相 關 研 究 發 現 serotoninergic receptors 與體溫調節作用之關係會因使用 動物品種、藥物劑量大小、給藥途徑及部位 之不同等而有所差異。(-)-pindolol 為 5-HT1A 受體及 β-adrenoceptor 之拮抗劑,可 拮抗 5-HT1A 受體之致效劑 (8-OH-DPAT, gepirone, (+)S-20499) 所 誘 發 之 降 溫 作 用 (22) ;8-OH-DPAT 為突觸前及突觸後 5-HT1A 受體之致效劑(23,24),腹腔或皮下給予可引起 明顯之降溫作用(25-28),並降低腦中 5-HT 之釋放(29);近來研究指出 8-OH-DPAT 降溫 作用之機轉大鼠與鼷鼠並不相同,在大鼠
8-OH-DPAT 之降溫作用不會因 5,7-DHT 前 處 理 破 壞 5-HT neurons 或 利 用 p-chlorophenylalanine (p-CPA; 5-HT synthesis
inhibitor) 排空腦內 5-HT 含量而消失,亦不 因給予選擇性之 5-HT uptake 抑制劑、5-HT precursor 或 5-抑制劑、5-HT releasing agent 提高 腦內 serotonin 之釋放所影響,故其對大鼠 降溫作用是藉由興奮突觸後 5-HT1A 受體 所達成(30,31);而鼷鼠則剛好相反是藉由興奮 突觸前 5-HT1A 受體所致(31,32)。實驗結果發 現 puerarin 誘發之降溫作用可被 5-HT1A 受 體 之 拮 抗 劑 (-)-pindolol 所 拮 抗 (Fig.3) , 或 為 5-HT1A 受 體 之 致 效 劑 8-OH-DPAT 所 加 強 (Fig.4) ; 故 可 推 知 puerarin 對室溫下正常大鼠之降溫作用可 能是藉由致效突觸後 5-HT1A 受體所致。 其次,全身性給予 5-HT2A/C 受體之致 效劑 DOI、MK-212 或 quipazine 等均會 引起體溫上升之作用(27,33-35),且此升溫作用 可 為 事 先 給 予 5-HT2A/C 受 體 之 拮 抗 劑 ketanserin、LY53857、mianserin、ritanserin 或 5-HT1A/2 及 D2 受 體 之 拮 抗 劑 spiperone 所阻斷(36,37),而 5-HT2 受體之拮 抗劑 ketanserin 或 pirenperone 單獨給藥 時則具降溫之作用(38),顯示 5-HT2 受體在 體溫調節作用上扮演一與 5-HT1A 受體相反 之升溫的角色;進一步的研究發現 DOI 不 論中樞或周邊給予均可誘發大鼠之升溫作 用(20),且其升溫作用不受 5-HT 1A/1B/2C 受體 及 β-adrenoceptor 之 拮 抗 劑 propranolol 與 5-HT3 受 體之 拮 抗劑 MDL-72222 或 ondansetron 所影響,可知 DOI 是藉由興奮 突觸後 5-HT2A receptor 誘發大鼠之升溫作 用(37);而 quipazine 不僅為 5-HT1B/1C/2 受 體之致效劑亦為 5-HT3 受體之拮抗劑 (39) 與 5-HT-uptake 之抑制劑(40),可增加突觸 間 serotonin 之濃度(41-43),其誘發之升溫作 用亦是藉由興奮突觸後 5-HT2 受體所致 (44,45) 。實驗結果發現 puerarin 誘發之降溫 作用可被腹腔給予 pirenperone、ketanserin 之 5-HT2 受體拮抗劑所加強 (Fig.5,6),或 為 腹 腔 或 側 腦 室 給 予 DOI 及 腹 腔 給 予 quipazine 之 5-HT2 受 體 致 效 劑 所 拮 抗 (Fig.7-9);可知 puerarin 對室溫下正常大鼠 之降溫作用可能與阻斷突觸後 5-HT2 受體 有關。 綜合上述實驗結果,可知 puerarin 可 能藉由降低下視丘 5-HT 之濃度,並作用 於 postsynaptic serotonin recepter,致活
5-HT1A 受體及阻斷 5-HT2 受體,而達到降 低體溫之作用。 四、計畫成果自評 本研究計畫案計畫書中擬進行之各項 實驗除部分藥物給藥方式與劑量,依實際 實驗狀況略有修正更動外,均已完成。 五、參考文獻
[1] Lu XR., Gao E., Xu LZ., Li HZ., Kang B., Chen WN., Chen SM., Chai XS.: Blocking effect of puerarin on beta-adrenoceptors of isolated organs and the whole animal. Acta Pharmacologica Sinica. 1986, 7(6):537-9.
[2] Shen XL., Witt MR., Nielsen M., Sterner O.: Inhibition of [3H] flunitrazepam binding to rat brain membranes in vitro by puerarin and daidzein. Acta Pharmaceutica Sinica. 1996, 31(1):59-62.
[3] Overstreet DH., Lee YW., Rezvani AH., Pei YH., Criswell HE., Janowsky DS.: Suppression of alcohol intake after administration of the Chinese herbal medicine, NPI-028, and its derivatives. Alcoholism, Clinical & Experimental Research. 1996, 20(2):221-7.
[4] Wang LY., Zhao AP., Chai XS.: Effects of puerarin on cat vascular smooth muscle in vitro. Acta Pharmacologica Sinica. 1994, 15(2):180-2. [5] Fan LL., Sun LH., Li J., Yue XH., Yu HX.,
Wang SY.: The protective effect of puerarin against myocardial reperfusion injury. Study on cardiac function. Chinese Medical Journal. 1992, 105(1):11-7.
[6] Fan LL., Sun LH., Li J., Yue XH., Yu HX., Wang SY., Dong SQ.: Protective effect of puerarin against myocardial reperfusion injury. Myocardial metabolism and ultrastructure. Chinese Medical Journal. 1992, 105(6):451-6. [7] Jiang XL., Xu LN.: Beneficial effect of puerarin
on experimental microcircu-latory disturbance in mice. Acta Pharmaceutica Sinica. 1989,
24(4):251-4.
[8] Fan LL., O'Keefe DD., Powell WW Jr.:
Pharmacologic studies on radix puerariae: effect of puerarin on regional myocardial blood flow and cardiac hemodynamics in dogs with acute
myocardial ischemia.. Chinese Medical Journal. 1985, 98(11):821-32.29.
[9] Kang RX.: The intraocular pressure depressive effect of puerarin. Chung-Hua Yen Ko Tsa Chih [Chinese Journal of Ophthalmology]. 1993, 29(6):336-9.
[10] Song XP., Chen PP., Chai XS.: Effects of puerarin on blood pressure and plasma renin activity in spontaneously hypertensive rats. Acta Pharmacologica Sinica. 1988, 9(1):55-8. [11] Chai XS., Wang ZX., Chen PP., Wang LY., Lu
XR., Kang B.: Anti-arrhythmic action of puerarin. Acta Pharmacologica Sinica. 1985, 6(3):166-8. [12] Zhou Y., Su X., Cheng B., Jiang J., Chen H.:
Comparative study on pharmacological effects of various species of Pueraria. China Journal of Chinese Materia Medica. 1995, 20(10):619-21, 640.
[13] Brodie BB, Shore AA. The concept for a role of serotonin and norepinephrine chemical mediators in the brain. Ann NY Acad Sci. 1957; 66 : 631-42.
[14] Von EC. Physiology and pharmacology of temperature regulation. Pharmacol Rev. 1961; 13 : 361-98.
[15] Myers RD. Hypothalamic control of
thermoregulation.Neurochenical mechanisms. In Morgane PJ, Panksepp J (eds). Handbook of the Hypothalamus. Dekker. New York. 1980; 3 : 83-212.
[16] Gorden CJ. Thermal biology of the laboratory rats. Physiol Behav. 1991; 47 : 963-91.
[17] Lin MT. Effects of brain monoamine depletions on thermoregulation in rabbits. Am J Physiol. 1980; 364-71.
[18] Fernadez GA, Lopez RC, Perez UJ, Castaneda HG. Evidence for a postsynaptic action of the serotoninergic anxiolytics:ipsapirone, indoranate and buspirone. Brain Res Bull 1992; 28: 497-501.
[19] Frankfurt M, Mendelson SD, Mckittrick CR, Mcewen BS. Alterations of serotonin receptor binding in the hypothalamus following acute denervation. Brain Res 1993; 601: 349-52. [20] Hsieh MT, Chueh FY, Lin MT. Magnolol
decreases body temperature by reducing 5-hydroxytryptamine release in the rat hypothalamus. Clin Exp Pharmacol Physiol. 1998; 25: 813-7.
[21] Emile Z, Gilles F. 5-hydroxytryptamine receptor. Am Pharmacol Exp Therap. 1992; 44 (3) : 401-58.
[22] Scott PA, Chou JM, Tang H, Frazer A. Differntial induction of 5-HT1A-mediated
responses in vivo by three chemically dissimilar 5-HT1Aagonists. J Pharmacol Exp Therap. 1994;
270 (1) : 198-208.
[23] Hjorth S, Carlsson A, Lindberg P, Sanchez D, Wikstrom H, Arvidsson L-E, Hacksell U, Nilsson JLG.
8-Hydroxy-2-(di-n-propylamino)tetralin, 8-OH-DPAT, a potentand selective simplified ergot congener with central 5-HT-receptor stimulating activity. J Neural Transm. 1982; 55: 169-88.
[24] Middlemiss DN, Fozard JR. 8-Hydroxy-2-(di-n-propylamino)tetralin discriminates between subtypes of the 5-HT1 recognition site. Eur J Pharmacol. 1983; 90: 151-3.
[25] Goodwin GM, De Souza RJ, Green AR. The pharmacology of the hypothermic response in mice to 8-hydroxy-2-(di-n-propylamino)tetralin, (8-OH-DPAT): a model of presynaptic 5-HT1
function. Neuropharmacology. 1985; 24: 1187-94.
[26] Gudelsky GA, Koenig JI, Meltzer HY.
Thermoregulatory responses to serotonin (5-HT) receptor stimulation in the rats.
Neuropharmacology. 1986; 25: 1307-13. [27] Hjorth S. Hypothermia in the rat induced by the
potent serotoninergic agent 8-OH-DPAT. J Neural Transm. 1985; 61: 131-5.
[28] Hypothermia inducecd by the putative 5-HT1A
agonist LY 165163 and 8-OH-DPAT is not prevented by 5-HT depletion. Eur J Pharmacol. 1987; 143: 221-8.
[29] Cooper JR, Bloom FE, Roth RH. The
biochemical Basis of Neuropharmacology. 1996; 353-407.
[30] Martin KF, Phillips I, Hearson M, Prow MR, Heal DJ. Charaterization of 8-OH-DPAT-induced hypothermia in mice as a 5-HT1A
autoreceptor response and its evaluation as a model to selectively identtify antidepressions. Br J Pharmacol. 1992; 107: 15-21.
[31] Millan MJ, Rivet JM, Canton H, Le S, Girardon M, Goe.A. Induction of hypothermia as a Model of 5-Hydroxytryptamine 5-HT1A
receptor-mediated activity in the rats: A pharmacological characterization of the actions of novel agonists and antagonists. J Pharmacol Exp Therap. 1993; 264 (3) : 1364-76.
[32] Bill DJ, Knight M, Forster EA, Fletcher A. Direct evidence for an important species difference in the mechanism of 8-OH-DPAT-induced hypothermia. Br J Pharmacol. 1991; 103: 1857-64.
[33] Nash JF, Meltzer HY, Gudelsky GA.
Antagonism of receptor mediated neuroendocrine and temperature responses by atypical
neuroleptics in the rat. Eur J Pharmacol. 1988; 151: 463-9.
[34] Salmi P, Karlsson T, Ahlenius S. Antagonism by SCH 23390 of clozapine-induced hypothermia in the rat. Eur J Pharmicol. 1994; 253: 67-73. [35] Yamanaki S, Lai H, Horita A. Dopaminergic and
mediation of thermal effects of apomorphine and dopamine. J Pharmacol Exp Therap. 1983; 227: 383-8.
[36] Mazzolai-Pomietto P, Aulakh CS, Wozniak KM, Hill JL, Murphy DL. Evidence that 1-(2,5-dimethoxy-4-iodophenyl)-2-aminopropane (DOI)-induced hyperthermia in rats is mediated by stimulation of 5-HT2A receptors.
Psychopharmacology. 1995; 117: 193-9. [37] Peter S, Sven A. Evidence for function
interactions between 5-HT1A and 5-HT2A
receptor in rats thermoregulatory mechanisms. Pharmacol Toxicol. 1998; 82: 122-7.
[38] Charanjit SA, Pascale MP, Krystyna MW, James LH, Dennis LM. Evidence that
1-(2,5- dimethoxy-4-methylphenyl)-2-amino-propane-induced hypophagia and hyperthermia in rats is mediated by serotonin-2A receptors. J Pharmacol Exp Ther. 1994; 270: 127-32.
[39] Hoyer D, Schoeffter P. 5-HT receptors: Subtypes and second messengers. J Recet Res. 1991; 11: 197-214.
[40] Alhaider AA. New hybrids of quipazine and trazodone as selective inhibitors of uptake of 5-hydroxytryptamine. J Pharm Sci. 1992; 81 (1) : 99-103.
[41] Apud JA, Grayson DR, De Erausquin E, Costa E. Pharmacological characterization of regulation
of phosphorinositide metabolism by recombinant 5-HT2 receptors in rat. Neuropharmacology.
1992; 31: 1-8.
[42] Conn PJ, Sanders-Bush E. Serotonin-stimulated phosphoinositide turnover: mediation by the S2
binding sites in rat cerebral cortex but not in subcortical regions. J Pharmacol Exp Ther. 1985; 234: 195-203.
[43] Cory RN, Rouot B, Guillon G, Sladeczek F, Balestre MN, Bockaert J. The
5-hydroxytrptamine (5-HT2) receptor stimulates
inositol phosphate formation in intact and broken WRK1 cells: Determination of occupancy-response relationships for 5-HT agonists. J Pharmacol Exp Ther. 1987; 241: 258-67. [44] Glennon RA & Luncki I. Behavioral models of
serotonin receptor activation. In : the Serotonin Receptors, ed. by E Sanders-Bush. pp. 253-93, Humana Press Clifton, 1988..
[45] Pawlowski L. Amitriptyline and femoxetine, but not clomipramine or citalpram, antagonize hyperthermia induced by directly acting 5-hydroxytryptamine-like drugs in heat adapted rats. J Pharm Pharmacol. 1984; 36: 197-9.
Fig. 1 Time course of the effects of intraperitoneal administration of puerarin on colonic
temperature in rats. Puerarin (1, 5, 10, 30mg/kg, i.p.) was injected at 0 min. The colonic temperature of vehicle-injected rats was 37.82±0.20°C at time 0 min. ∆, denote the difference between the control value before injected and exchange after injected. The value are mean±SEM of 8-12 rats per group.*P<0.05, **P<0.01, significantly different from corresponding control value (vehicle group), ANOVA.
T im e ( m i n ) - 6 0 0 6 0 1 2 0 1 8 0 2 4 0 3 0 0 3 6 0 ∆ Tc o ( o C ) - 2 .0 - 1 .5 - 1 .0 - 0 .5 0 .0 0 .5 V e h ic le P u e r a r in e 1 m g /k g i.p . P u e r a r in e 5 m g /k g i.p . P u e r a r in e 1 0 m g /k g i.p . P u e r a r in e 3 0 m g /k g i.p . * * * * * * * * * * * * * * * * * * * * * * * * * * * *
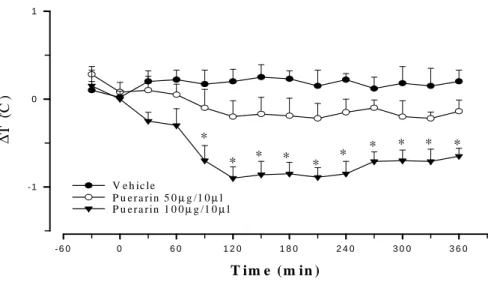
Fig. 2 Time course of the effects of intracerebroventricular administration of puerarin on
colonic temperature in rats. Puerarin (50, 100µg/kg, i.c.v.) was injected at 0 min. The colonic temperature of vehicle-injected rats was 37.14±0.15°C at time 0 min. ∆, denote the difference between the control value before injected and exchange after injected. The value are mean±SEM of 8 rats per group.*P<0.05, significantly different from corresponding control value (vehicle group), ANOVA.
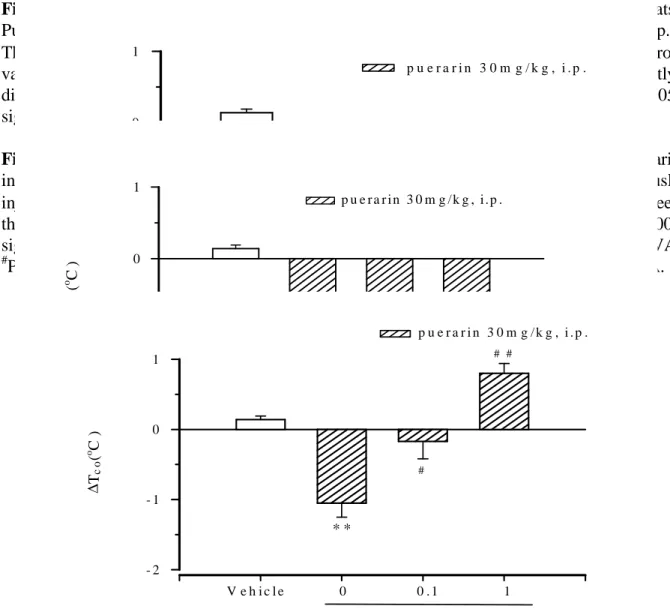
Fig. 3 Effects of (-)-pindolol (0.05, 0.5mg/kg; s.c.) on the hypothermic response of puerarin in
rats. Puerarin was injected 30 mins before (-)-pindolol (0.05, 0.5mg/kg) subcutaneously injected (s.c.) The value are mean±SEM of 8 rats per group. ∆, denote the difference between the control value before 120 mins after the start of puerarin injected. **P<0.01, significantly different from the corresponding control value (vehicle group), ANOVA. #P<0.05, ##P<0.01 significantly different from corresponding control value (puerarin group), ANOVA.
T im e (m in ) - 6 0 0 6 0 1 2 0 1 8 0 2 4 0 3 0 0 3 6 0 ∆T ( 0 C ) - 1 0 1 V e h ic le P u e r a r in 5 0µg /1 0µl P u e r a r in 1 0 0µg /1 0µl * * * * * * * * * * V e h ic le 0 0 .0 5 0 .5 (-)-p in d o lo l m g /k g , s.c . ∆ Tc o( o C ) -2 -1 0 1 p u e ra rin 3 0 m g /k g , i.p . * * # # #
7
Fig. 4 Effects of 8-OH-DPAT (0.01, 0.05mg/kg, s.c.) on the hypothermic response of puerarin
in rats. Puerarin was injected 100 mins before 8-OH-DPAT (0.01, 0.05mg/kg) subcutaneously injected (s.c.) The value are mean±SEM of 8 rats per group. ∆, denote the difference between the control value before 120 mins after the start of puerarin injected. **P<0.01, ***P<0.001 significantly different from the corresponding control value (vehicle group), ANOVA. #
P<0.05 significantly different from corresponding control value (puerarin group), ANOVA.
Fig. 5 Effects of ketanserin (0.5, 1mg/kg, i.p.) on the hypothermic response of puerarin in rats.
Puerarin was injected 60 mins before ketanserin (0.5, 1mg/kg) intraperitoneal injected (i.p.) The value are mean±SEM of 8 rats per group. ∆, denote the difference between the control value before 120 mins after the start of puerarin injected. **P<0.01, ***P<0.001 significantly different from the corresponding control value (vehicle group), ANOVA. #P<0.05 significantly different from corresponding control value (puerarin group), ANOVA.
Fig. 6. Effects of pirenperone (0.05, 0.2mg/kg, s.c.) on the hypothermic response of puerarin
in rats Puerarin was injected 60 mins before pirenperone (0.05, 0.2mg/kg) subcutaneously injected (s.c.) The value are mean±SEM of 8 rats per group. ∆, denote the difference between the control value before 120 mins after the start of puerarin injected. **P<0.01, ***P<0.001 significantly different from the corresponding control value (vehicle group), ANOVA. #
P<0.05 significantly different from corresponding control value (puerarin group), ANOVA.
V e h i c l e 0 0 .0 1 0 . 0 5 8 - O H - D P A T m g / k g , s . c . ∆T c o ( o C ) - 2 - 1 0 1 p u e r a r i n 3 0 m g / k g , i . p . * * * * * * * # V e h i c l e 0 0 . 5 1 k e t a n s e r i n m g / k g , i . p . ∆T c o ( o C ) - 2 - 1 0 1 p u e r a r i n 3 0 m g / k g , i . p . * * * * * * * # V e h i c l e 0 0 . 0 5 0 .2 p i r e n p e r o n e m g /k g , s .c . ∆T c o ( o C ) - 2 - 1 0 1 p u e r a r i n 3 0 m g / k g , i . p . * * * * * * * # ∆T c o ( o C ) - 2 - 1 0 1 p u e r a r i n 3 0 m g / k g , i . p . * * # # #
Fig. 7 Effects of quipazine (0.1, 1mg/kg, i.p.) on the hypothermic response of puerarin in rats.
Puerarin was injected 60 mins before quipazine (0.1, 1mg/kg) intraperitoneal injected (i.p.) The value are mean±SEM of 8 rats per group. ∆, denote the difference between the control value before 120 mins after the start of puerarin injected. **P<0.01 significantly different from the corresponding control value (vehicle group), ANOVA. #P<0.05, ##P<0.01 significantly different from corresponding control value (puerarin group), ANOVA.
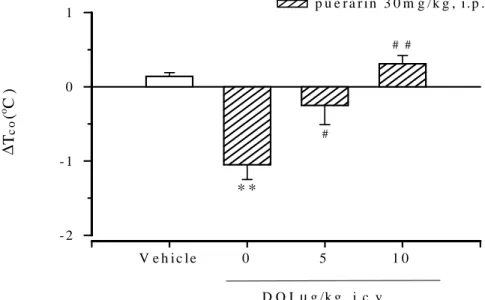
Fig. 8 Effects of DOI (0.5, 1mg/kg, i.p.) on the hypothermic response of puerarin in rats
Puerarin was injected 90 mins before DOI (0.5, 1mg/kg) intraperitoneal injected (i.p.) The value are mean±SEM of 8 rats per group. ∆, denote the difference between the control value 120 mins after the start of puerarin injected. *P<0.05, **P<0.01, significantly different from the corresponding control value (vehicle group), ANOVA. #P<0.05, ##P<0.01 significantly different from corresponding control value (puerarin group), ANOVA
Fig. 9 Effects of DOI (5, 10µg/10µl, i.c.v.) on the hypothermic response of puerarin in rats. Puerarin was injected 90 mins before DOI (5,10µg/10µl) intracrerbroventricular injected (i.c.v.) The value are mean±SEM of 8 rats per group. ∆, denote the difference between the control value 120 mins after the start of puerarin injected. *P<0.05, **P<0.01, significantly different from the corresponding control value (vehicle group), ANOVA. #P<0.05, ##P<0.01 significantly different from corresponding control value (puerarin group), ANOVA
V e h i c l e 0 5 1 0 D O I µg /k g , i.c .v . ∆ Tc o ( o C ) - 2 - 1 0 1 p u e r a r i n 3 0 m g /k g , i . p . * * # # # V e h i c l e 0 0 . 5 1 D O I m g / k g , i . p . ∆T c o ( o C ) - 2 - 1 0 1 p u e r a r i n 3 0 m g / k g , i .p . * * * # # #
Table 1. The effect of puerarin on the hypothalamic serotonin (5-HT) release in the normal rats.
Treatments
(i.p.)
∆
Colonic temperature
(
°C)
Hypothalamic 5-HT
release (% baseline)
Vehicle
0.14
±
0.05
99.12
±
34.25
Puerarin
10mg/kg
-0.78
±
0.21*
52.14
±
35.23*
30mg/kg
-1.05±0.20**
25.12±31.22**
The value are mean±SEM of 5 rats per group. The vehicle-treated control value for extracellular 5-HT release in the hypothalamus are 1.58±0.65pg/18µl/30 mins. ∆, denote the difference between the control value before 120 mins after the start of puerarin injected. *P<0.05, **P<0.01 significantly different from the corresponding control value (vehicle group), ANOVA.
Table 2. The effects of puerarin on the colonic temperature in rats treated by the 5,7-DHT.
Changes in colonic temperature (
∆
℃)
Treatment
Normal
5,7-DHT
Vehicle
0.14±0.05
0.20±0.14
puerarin
10mg/kg (i.p.)
-0.78±0.21*
-0.21±0.11
#30mg/kg (i.p.)
-1.05
±
0.20**
-0.53
±
0.13*
#The value are mean±SEM from 8 rats per group. The control value for colonic temperature are 37.8±0.12℃ and 38.1±0.14℃ for normal and 5,7-DHT-treated rats, respectively. ∆, denote the difference between the control value before 120 mins after the start of puerarin injected. *P<0.05, **P<0.01 significantly different from corresponding control value (vehicle group), ANOVA. #P<0.05 significantly different from corresponding control value (normal group), ANOVA.