行政院國家科學委員會專題研究計畫 期中進度報告
自然弒細胞發生學的雙系模式:白血球介質 15 及 21 分別導
引 CD56+Kir ±lectis+ 弒細胞 及 CD56±Kir lectin 自然弒
細胞(1/3)
計畫類別: 個別型計畫 計畫編號: NSC91-2314-B-002-179-執行期間: 91 年 08 月 01 日至 92 年 07 月 31 日 執行單位: 國立臺灣大學醫學院免疫學研究所 計畫主持人: 許世明 計畫參與人員: 林中梧、林鉦鎔、楊朝順、蘇哲俊、王坤藤 報告類型: 精簡報告 報告附件: 出席國際會議研究心得報告及發表論文 處理方式: 本計畫可公開查詢中
華
民
國 92 年 5 月 27 日
行政院國家科學委員會補助專題研究計畫
期中進
度報告
自然弒細胞發生學的雙系模式:白血球介質 15 及 21 分別導引
CD56+Kir±lectis+ 弒細胞 及 CD56±Kirlectin 自然弒細胞 (1/3)
計畫類別:■ 個別型計畫
□ 整合型計畫
計畫編號:NSC 91-2314-B-002-179
執行期間:91 年 08 月 01 日至 92 年 07 月 31 日
計畫主持人:許世明
共同主持人:
計畫參與人員:林中梧
、林鉦鎔、楊朝順、蘇哲俊、王坤藤
成果報告類型(依經費核定清單規定繳交):■精簡報告 □完整
報告
本成果報告包括以下應繳交之附件:
□赴國外出差或研習心得報告一份
□赴大陸地區出差或研習心得報告一份
□出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份
處理方式:除產學合作研究計畫、提升產業技術及人才培育研究
計畫、列管計畫及下列情形者外,得立即公開查詢
□涉及專利或其他智慧財產權,□一年□二年後可公開
查詢
附件一
執行單位:台大醫學院免疫所
中英文摘要
自然弒細胞發生學的雙系模式:白血球介質 15 及 21 導引 CD56+ Kir±lectis+ 第一 型 自然弒細胞及 CD56± Kir+lectin± 第二型自然弒細胞 關鍵詞: 發生學、自然弒細胞、介質 15、介質 21 免疫球蛋白受器、CD56、C-型糖類結合蛋白 自然弒細胞的發生須經由介質 15 作用,缺乏介質 15 的基因轉移老鼠完全沒 有自然弒細胞,但是人類的自然弒細胞和老鼠不同,其細胞表面有 CD56、C-type lectin 及 KIR,這些是老鼠所沒有的,更重要的是自然弒細胞可分泌 type I (IFN-r) 及 type II (IL-4)這兩種類型的自然弒細胞、和 CD56、C-type lectin 及 KIR 表現型的關係至今不明。 自然弒細胞和 T-細胞由同一來分化的 CD44+ CD25-CD161+ T/NK 先驅細胞 分化而來,這一 T/NK 細胞受介質 7 或轉錄因子 Notch、E2A 的作用分化為 T-細胞,相反的若受到介質 15 及轉錄因子 Id3 則分化為自然弒細胞,值得注意的 是,介質 15 及介質 7 它們有共同的受器 rc,因此我們可以利用流體細胞儀來分 離未分化的 CD44+ CD25-CD161+ rc+T/NK 先驅細胞,並將這細胞用來研究自然 弒細胞的分化及分生學,我們最有與趣的 3 個問題: 1. CD56+及 CD56±:人類的自然弒細胞可分為 CD56+及 CD56± 二 類,於 Nature am 的發表的分析顯示 CD56+NK cells 可經由介質 21 產生,但是其發生過程 詳細情形仍不清楚,和經由介質 15 所產生的的 CD56+ NK cell 的關係亦不明。 2. type I 及 type II:自然弒細胞和 F helper 細胞一樣可為根據其分泌的介質分為Type I 及 type Type I 主要分泌 IFN-r,type II 主要分泌 IL-4 而一般認為 type I 及 type II 是由同一個未成熟的 type O 細胞產生的,但是最近 Nature Immunol-py 21 所發表的報告顯示發生的過程是 type II-type-o-type-I,我們想 利用 T/NK 先驅細胞,來分別這 2 個模式。
3. C-lectin 及 KIR:人類的自然弒細胞和老鼠不同,不但有 C-lectin 還用 KIR 來 當做抗原受器(Trends Immunelop>001)我們認為 C-lectin 可能是發生學上較早 的抗原受器較為原始也較為不具特定性。
總結以上相對於過去學者所用的功能性分析,我們將以發生學的觀點來研究 自然弒細胞的發生以期建立一個人類自然弒細胞的雙系模式:
介質 15 及 21 分別促成 CD56+
Kir±lectin+等一型及 CD56±Kir+lectin±第二型自然弒 細胞的生成,這個模式可用於台灣常見的鼻咽淋巴癌的分類,因此也具有相當的 臨床意義。
A dual lineage model for the development of matur e human NK-cells: Inter leukin 15 and inter leukin 21 dr ive r espectively the development of
CD56br igthtKIRdimLectinbr ight type I (IFN-γ+) NK-cells and
CD56dimKIRbr ightLectindim type II (IL-13+) NK-cells
Keywords: Developmental biology, NK-cells, IL-15, IL-21, KIR, CD56, C-type lectin.
NK-cell development depends on interleukin 15, as shown by the complete absence of NK-cells in transgenic mice knocked out for either interleukin 15 or its receptor. However, mature human NK-cells in the peripheral blood differ from mouse NK-cells, with varying expression of surface NK-markers, such as CD56, C-type lectin, and KIR, and secretion of type I and type II cytokines. It is still unclear how the
expr ession patter ns of CD56, C-type lectins and KIRs ar e r elated to each other and to their functions, type I vs type II.
NK-cells and T-cells arise from a common CD44+, CD25-, CD161+ bipotent T/NK precursor, which subsequently diverges into T-cells under interleukin 7, Notch and E2A, or
alternatively into NK-cells under interleukin 15 and Id3. Id3 and E2A are both helix-loop-helix transcription factors, whereas interleukins 7 and 15 have a 4-α-helical-bundle structure using a common γ-chain (γc) in their receptors. For the study of NK-cell development, we propose to isolate from bone marrow, thymus, or peripheral blood by flow cytometry the common bipotent CD44+, CD25-, CD161+,
γc +, T/NK cells. They will be positively selected for interleukin 15 receptor and Id3,
and negative selected for interleukin 7 receptor and E2A. We will then use a
developmental appr oach to support, clarify, or refute previous
functional-str uctur al studies on 3 issues.
(I) CD56br igtht vs CD56dim. Human NK-cells could be divided into a CD56brigtht subset and a CD56dim subset. Based on functional characterization (Natur e 2000) that showed expansion of CD56dim NK-cells under interleukin 21, we want to show that NK-cell precursors will develop into either the CD56brigtht subset in the presence of interleukin 15 or the CD56dim subset in the presence of interleukin 21.
(II) Type I vs Type II. In the classical dichotomy model, a common type zero NK
cell will give rise to either type I NK cells secreting predominantly IFN-γ and type II NK-cells secreting predominantly interleukin 4 and interleukin 13. However, functional analysis (Natur e Immunology 2001) seems to indicate a type II-type 0-type I sequence. We will clarify the dichotomy vs sequential controversy by tracing back to the common bipotent precursors.
(III) C-type Lectin vs KIR. Human NK-cells use both C-type lectins and KIRs as antigen receptors that recognize different structures of the MHC-1 molecules (Tr ends
Immunology, 2001). We want to show that C-type lectin might represent a more
primitive less specific antigen receptor earlier in the development of NK-cells.
Using a developmental approach, we want to integrate previous functional-structural data into a dual lineage model for the development of matur e
human NK-cells in which inter leukin 15 and inter leukin 21 dr ive r espectively the development of CD56br igthtKIRdimLectinbr ight type I (IFN-γ+) NK-cells and CD56dimKIRbr ightLectindim type II (IL-13+) NK-cells. This model can then be
applied to the study of sinonasal lymphoma, which is an EBV-associated NK-cell lymphoma rather common in Taiwan, and might contribute to the clinical management of these patients.
報告內容
We have found that peripheral NK cells are either CD94+ or CD94-. The CD94+ NK cells are probably more mature than the CD94- type. This distinction does have clinical implications, for example, we showed that the CD94+ phenotype is associated with a better prognosis in sinonasal lymphomas. We are currently investigating the clinical implications of the dual lineage model.
Expr ession of CD94 char acter izes a matur e subtype of sinonasal lymphoma with excellent pr ognosis Expression of NK-cell antigen receptors (NKRs), such as
CD94, NKG2, and killer immunoglobulin-like receptors (KIRs), is developmentally regulated and clonally distributed. We have shown a restricted KIR (rKIR) repertoire without monoclonal T-cell receptor (mTCR) rearrangement in sinonasal lymphomas (SNLs) of true NK-cell lineage. The lineage assignment, however, does not specify distinct developmental stages that might have clinical significance. CD94, often co-expressed with one of the NKG2 family, is the first NKR expressed by developing NK cells, followed by NKG2A, NKG2E, and finally NKG2C. This sequence suggests an immature CD94-and a mature CD94+ subtype of SNLs, and possibly a better prognosis of the mature type. Using a novel RT-PCR approach, we analyzed the NKR patterns in 25 SNLs, and we confirmed a true NK lineage in 19 cases. Among them, 8 of 10 CD94+ patients survived beyond 1 year, but 7 of 9 CD94- patients died within 1 year (median survival: 60 months vs 10 months by Meier-Kaplan survival analysis, p=0.026 by Cox-F test). The remaining 6 patients had an rKIR repertoire plus an mTCR rearrangement, suggesting mixed NK/T differentiation. They were CD94-, found predominantly in young females, and had a median survival of 35 months. Thus, based on the patterns of NKRs, a division of sinonasal lymphomas into CD94+, CD94-, and mixed NK/T types reflects a true biological divergence that manifests as differences in clinical behavior.
INTRODUCTION
Sinonasal lymphoma (SNL) is a rare disease that presents both diagnostic and therapeutic difficulties. The lymphoma may cause extensive ulceration and necrosis of the nasal cavity, often leaving little viable tissue for diagnosis. The lymphoma can pursue an aggressive course, sometimes terminating unexpectedly in fatal hemophagocytosis.1,2 At present, the only known prognostic factor for SNLs is the stage at diagnosis. However, patients with the same initial stage have greatly variable clinical courses.3 It is highly desirable to have molecular tests to confirm
the diagnosis and predict the outcome.
The cellular origin of SNL has been controversial because of the difficulty in separating the T lineage from the NK lineage, both being derived from the same bi-potential T/NK precursor.4,5 SNL is cytoplasmic CD3-positive, suggesting a T lineage, but, unlike other T-cell lymphomas, it is surface CD3-negative and does not have a monoclonal T-cell-receptor (TCR) rearrangement.6 Because cytoplasmic CD3 can also be found in fetal NK cells,7,8 and CD56, an NK-cell marker, is frequently expressed by SNLs, most authors now believe SNLs have an NK-cell origin.
Like TCRs or immunoglobulins, which are the antigen receptors for T cells or B cells, respectively, NK cells use CD94, NKG2, and killer immunoglobulin-like receptors (KIRs) as NK-cell antigen receptors (NKRs).9 Furthermore, the expression of NKRs is developmentally regulated and clonally distributed.10 Because a monoclonal TCR rearrangement is the gold standard for T-cell lymphoma and light-chain restriction is characteristic of B-cell lymphoma, the patterns of NKRs might be used for establishing monoclonality for an SNL of the NK lineage and further identifies developmental stages within the NK lineage.
The KIRs have a consensus sequence composed of 3 immunoglobulin-like domains, D0, D1, and D2.11-14 Alternative splicing of the consensus sequence results in 3 groups, KIR2DL4 (D0-D2), KIR2D (D1-D2), or KIR3D (D0-D1-D2). A polyclonal NK-cell population is expected to express all 3 groups, but a monoclonal NK-cell population would have a KIR repertoire restricted to one or 2 groups. As expected, we have shown that a restricted KIR repertoire is typically found in SNLs of NK-cell origin.15 The demonstration of a restricted KIR repertoire, however, could not be used further to specify distinct developmental stages that might predict outcomes.16
Unlike KIRs, CD94 and NKG2 are C-type lectins.17 The NKG2 family includes 2A, 2B, 2C, 2D, 2E, and 2F. During NK-cell development, CD94 is expressed first, followed by 2A, 2E, and finally 2C.18-21 Usually, the expressed CD94 forms heterodimers with NKG2, usually 2A, but sometimes with 2E or 2C.22 NKG2B is a splicing variant of 2A. NKG2F is highly homologous to 2C and 2F, and is a 3’-truncated variant. NKG2D is divergent from the other members, and is expressed as homodimer. This developmental regulation implies an immature CD94-and a mature CD94+ NK-cell lymphoma, and, possibly, a better clinical outcome associated with the mature phenotype.23
To prove the hypothesis that expression of CD94/NKG2 receptors is of prognostic value, we developed a novel RT-PCR approach to characterize the expression of
CD94 and the NKG2 family in 25 SNLs. In conjunction with the patterns of KIRs, we hope to identify mature and immature subsets, and to correlate the phenotype and genotype with the clinical course of SNLs.
Mater ials and Methods Tissue samples
Tissue sections and medical records of all sinonasal lymphomas (SNLs) at the Department of Pathology, National Taiwan University Hospital, in the period from 1995 to 2002, were reviewed by 2 independent hematopathologists. The diagnosis was made by a combination of morphology, immunohistochemistry, and clinical features according to the WHO classification. Twenty-five cases with sufficient tissue for studies were collected. All of the cases were CD3-positive by immunohistochemistry and EBER (EB virus encoded RNA) positive by in situ hybridization. Parts of the KIR and TCR data have been published in 2 previous papers.14,15 Nine consecutive lymphoblastic lymphomas involving the mediastinum or the lymph nodes were used in a search for possible NK differentiation at the immature stage. These lymphomas were CD3 and TdT (terminal deoxynucleotidyl transferase). Ten randomly chosen peripheral T-cell lymphomas were used as negative controls. All 10 cases had a monoclonal TCR rearrangement.24
RT-PCR for CD94
The cDNA sequence of CD94 was taken from Genbank U30610.25 For reverse
transcription, an antisense primer,
5’-acaaactgtgttcactagcACCCATTTTTCTTGGCAA GAACAGCA-3’ (281-256) was used. The capital letters correspond to the antisense sequence of CD94 cDNA from position 271 to 251. The small letters constitute a random sequence for increased PCR specificity and incorporation of fluorescent labels.
Specifically, reverse transcription was done in a 20-µl reaction mixture at 40o
C for 50 minutes. The mixture included 0.1 µg of RNA purified from formalin-fixed, paraffin-embedded tissue blocks, 0.5 µM antisense primer, 200 units of reverse transcriptase, 50 mM Tris-HCl at pH 8.3, 75 mM KCl, 3 mM MgCl2, 10 mM DTT,
and 200 µM of each dNTP. A reaction for actin was run as a positive control, and a reaction under the same condition except for the omission of reverse transcriptase was run as a negative control. The cDNA from 2 µl of the RT reaction mixture was
then PCR-amplified by a forward primer,
5’-TTGTTGAAAAATTCTTTTACTAAACT-3’ (172-197), and a fluorescence-labeled primer of the random sequence,
5’-tet-cttctgacaaactgtgttcactagc-3’. Specifically, a 20 µl PCR mixture included cDNA, 0.3 M of each primer, 15 mM Tris-HCl at pH 8.0, 1.5 mM MgCl2, 50 mM
KCl, 200 µM of each dNTP, and 1 unit of Taq polymerase. The reaction mixture was subjected to 35 cycles of PCR. Each cycle consisted of denaturation at 94 C for 45 seconds, annealing at 45C for 45 seconds, and extension at 72C for 45 seconds. At the end of 35 cycles, a portion of the PCR product was loaded on and separated by a high-resolution electrophoretic system (ABI377 equipped with GeneScan software, Perkin-Elmer, Foster City, CA). The size for the RT-PCR product of CD94 is 135.
Sequence alignment and RT-PCR for the NKG2 family
The NKG2 family, with Genbank accession numbers in parentheses, includes NKG2A (X54867) and its splicing variant 2B, NKG2C (X54869), NKG2D (X54870), NKG2E (L14542), and NKG2F (AJ001683).26-30 Multiple sequence alignment was performed with the MAP program from Baylor College of Medicine, Houston, Texas.
As shown in the top part of Figure 1, the consensus structure of the NKG2 family consists of a 5’ untranslated region, an intracytoplasmic domain (cyt), a transmembrane domain (TM), an extracellular carbohydrate recognizing domain (CRD) and a 3’ untranslated region.
Below the consensus sequence, in the middle part of Figure 1, individual cDNAs were aligned with exon boundaries indicated by the vertical bars. Note the correspondence between the domains and the exon structure. NKG2B is a splicing variant of NKG2A without the consensus TM-CRD junction. NKG2C and NKG2E are highly homologous and have identical TM-CRD junctions. NKG2F also has homology with C and E, and is a truncated variant at the 3’ end. Finally, NKG2D is only distantly related to the others.
In the lower part of Figure 1, the highly homologous sequence at the TM-CRD junction is shown. This portion is used for the design of an efficient RT-PCR approach by a consensus reverse primer and 4 specific forward primers. The
consensus reverse primer is
5’-acaaactgtgttcactagcATCCACTTTTCAGGACAAGGGCCACA-3’. The specific forward primers, and their cDNA positions in parentheses, were 5’-TTGT
TATTCCCTCTACATTAA-3’ for 2A/2B (436-456);
5’-ATAGTTCTTATTCCTTTCCT -3’ for 2C/2E (278-297 of 2C or 316-335 of 2E); 5’-GAGTGCTGTATTCCTAAACT-3’ for 2D (560-579); and 5’-AATAGTTCTTATTCCTTGTATTGG-3’ for 2F (452-475).
The conditions for RT-PCR were the same as that for CD94, except for the primers. The PCR products were also separated by the GeneScan system. The expected size
for the RT-PCR products are 125 for A, 120 for C/E, 115 for D, and 130 for F. NKG2B, which does not have the correct TM-CRD junction, is not amplified. Illustrative patterns of the CD94/NKG2 analysis are shown in Figure 2.
Gr oup specific RT-PCR for KIRs
We have used a group-specific RT-PCR to demonstrate a restricted KIR repertoire as a clonality test for NK-cell lymphomas.15 Briefly, the KIRs can be put into one of 3 splicing groups, KIR2DL4, KIR2D, or KIR3D. Three pairs of group-specific primers were designed to target the unique junctions created by splicing. They are
5’-GCTGAGAGAGAAGGTTCTCATAT-3’ and
5’-CACTCCCCCACTGGGTGGTCGGC -3’ for KIR 2DL4, 5’-TGGGTGGGCCAGAGGAAGGTTT-3’ and 5’-CATGGCGTG
TGTTGGGTTCTTCTTG-3’ for KIR2D, and
5’-TGGGTGGGCCAGGAGGAAGGTTT-3’ and 5’-CACTCC
CCCCACTGGGTGGTCGGC-3’ for KIR3D.
The RT-PCR conditions were the same as those for CD94 and the NKG2 family, except that fluorescence-labeled dCTP was used at a concentration of 2 µM[TAMRA]-dCTP during the PCRs. The sizes for the PCR products were 83, 85, and 90 for KIR2DL4, KIR2D, and KIR3D, respectively. Two common patterns of KIR repertoires are shown in Figure 3.
T-cell-receptor gene rear r angement
PCR-based tests were done according to a protocol developed in our laboratory.16,24
Immunohistochemistr y
IHC for CD3 and CD56 was done for all cases on formalin-fixed, paraffin-embedded tissue sections. Immunohistochemistry for CD94 and NKG2A was done, when available, on fresh frozen tissue fixed with acetone. All antibodies were used according to the manufacturer’s recommendation: CD3 (Dako SA, Glostrup, Denmark), CD56 (Novocastra Lab LTD, Newcastle, UK), CD94 (BD PharMingen, San Diego, CA, USA), and NKG2A (Santa Cruz Biotechnology, Santa Cruz, CA, USA). Illustrative staining patterns are shown in Figure 4.
Sur vival analysis
Kaplan-Meier survival analysis based on the product-limit method was done by the statistical software Statistica (StatSoft, Tulsa, OK, USA). Two-sample comparison between the CD94+
vs CD94
subtypes was done by the Cox-F test. Survival curves for the CD94+
and CD94
subtypes are shown in Figure 5.
RESULTS
Lineage assignment
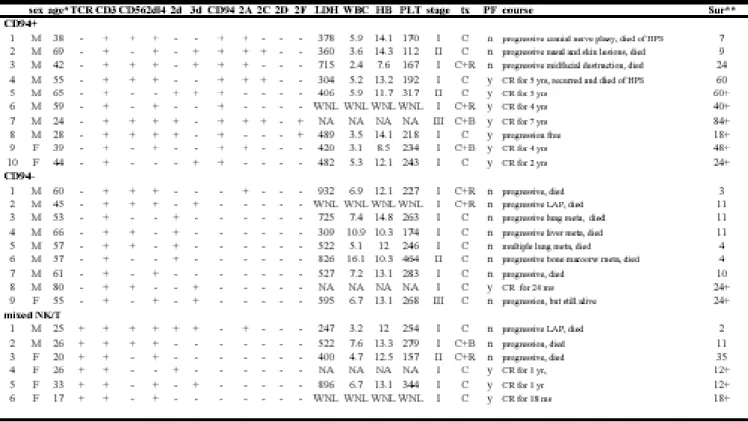
We have used a restricted KIR repertoire without a monoclonal TCR rearrangement as a criterion for the NK lineage,15 a monoclonal TCR rearrangement without a restricted KIR repertoire as a criterion for the T lineage, and a restricted KIR repertoire plus a monoclonal TCR rearrangement as a criterion for mixed NK/T differentiation.16 This assigned the 25 SNLs to 19 cases of the NK lineage (NK-SNL) and 6 cases to the mixed NK/T lineage (NK/T-SNL), as shown in Table I.
Lineage assignment or patter ns of KIR reper toires do not affect sur vival
The survival time, in terms of months, of the 25 cases was shown in the last column of Table I. The 1-year survival rate was 0.53 (10/19) for the 19 NK-SNLs and 0.67 (4/6) for the 6 NK/T-SNLs. The difference was not significant (p=0.69). For the 19 NK-SNLs, the presence or absence of KIR2DL4, KIR2D, and KIR3D also did not affect survival significantly either. For example, the difference between the 1-year survival rate for the KIR2DL4 NK-SNLs (7/12 or 53%) and that of the KIR2DL4 NK-SNLs (3/7 or 43%) was not statistically significant (p=0.32). Similarly, there was no statistical significance in the comparisons: KIR2D (3/7 or 43%) vs KIR2D (7/12 or 58%) at p=0.53, or KIR3D (5/7 or 71%) vs KIR3D (5/12 or 42%) at p=0.24. This showed that the patterns of KIR repertoires or the lineage assignment were not predictive of clinical outcomes.
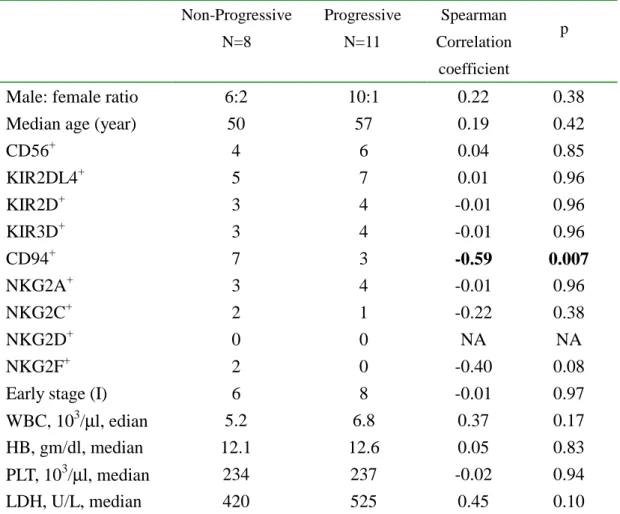
Disease progression in SNLs has the str ongest association with CD94 expression
To find the factors that might determine clinical outcomes, we assigned the 19 NK-SNLs to 2 groups, depending on the response after treatment: non-progressive (N=8) and progressive (N=11). The non-progressive group consisted of patients with complete remission, partial remission, or a stable lesion after treatments. The progressive group consisted of patients with lymphoma progression, midfacial destruction, cranial nerve palsy, regional and/or systemic lymphadenopathy, or distant metastasis. Among various clinical and pathologic features evaluated in Table II, we noticed that only CD94 had a strong association with disease progression, with a Spearman correlation coefficient of -0.59 at p=0.007. It is of interest to note again that the presence or absence of KIRs had almost no correlation with disease progression (p=0.96). Among the traditional prognostic indices, such as stage, LDH, and blood counts, only the level of LDH reached marginal significance at p=0.10.
Distinct patter ns of CD94/NKG2 receptor s were identified in SNLs
We separated the 19 NK-SNLs into 10 CD94+ and 9 CD94- cases (Table III). Among the 10 CD94+ cases, three expressed CD94 only, and the remaining 7 cases expressed one or more NKG2 members, mostly 2A, followed by 2C and 2F. Among the 9 CD94- cases, one case unexpectedly expressed NKG2A, and the remaining 8 cases did not express any members of the NKG2 family. For the 6 NK/T-SNLs, only one case expressed NKG2A, and CD94 was not expressed at all. Thus, CD94 was not only predictive of non-progression for the NK-SNLs, but it was also found more frequently than NKG2 or any other members of the NKG2 family, and it was rarely associated with NK/T-SNLs.
Cor r elation with mor phology and immunohistochemistr y (IHC)
We have performed IHC for CD94 and NKG2A on 4 NK-SNLs that were CD94+/NKG2A+by RT-PCR, 3 peripheral T-cell lymphomas, 3 tonsils, 3 spleens, and 3 peripheral-blood smears. IHC showed strong and diffuse staining for CD94 and NKG2A (Figure 4) only for the 4 NK-SNLs. The finding not only confirmed the data from the RT-PCR approach, but was also significant in that all of the controls were either negative or showed only sparse positive cells. As shown in Table III, we compared 4 morphologic parameters: nuclear grade, clear-cell change, angiocentric necrosis, and reactive infiltrates of eosinophils and plasma cells. None of these had statistical significance among the 3 subtypes. The staining pattern of CD56, usually considered to be the characteristic marker of NK-cell differentiation, did not differ significantly among the 3 subtypes either (p=0.72).
Clinical cor r elation
The NK-SNLs, both CD94+ and CD94- types, were associated with middle-aged males (median ages 43 and 57 years, male to female sex ratio 8:2 and 8:1), whereas the NK/T-SNLs were associated with younger females (median age 26 years, male to female sex ratio 2:4). The differences among median ages and sex ratios were both statistically significant, p<0.001 and 0.047, respectively (Table III).
The 3 subtypes had comparable initial clinical presentations, in terms of stage, the level of LDH, and peripheral blood cell counts. However, the clinical outcomes differed significantly in terms of disease progression and 1-year survival (p=0.031 for both statistics). This was mainly due to the rapid progression and poor survival of the CD94- subtype.
To confirm further the significance of the 1-year survival rate, we constructed the Kaplan-Meier survival curves. Two-sample comparison between CD94+ and CD94 -subtypes showed a highly significant difference between the curves (p= 0.026;
Figure 5). The median survival times for CD94+ and CD94- subtypes were 60 months and 10 months, respectively. In contrast, the survival curve for the mixed NK/T subtype was between CD94+ and CD94- subtypes. The difference does not reach statistical significance, in part because of the small sample size. The median survival for the NK/T type was 35 months.
Patients with SNLs might sometimes terminate in a hemophagocytotic syndrome, defined clinically as fever, pancytopenia, hepatosplenomegaly, jaundice, and hemophagocytosis on bone marrow aspiration smears. We found only 2 CD94+ patients with this fatal outcome. However, no tissue materials were available for documenting hemophagocytosis. Thus, the occurrence of hemophagocytosis in SNLs is possible, but was rare in our series.
Lymphoblastic lymphoma may also expr ess KIRs, CD94, and NKG2
Lymphoblastic lymphoma (LBL) usually belongs to the T-lineage, but a true NK-lineage was also mentioned in the literature.31 This possibility should not be overlooked, because primitive hematologic neoplasms are often lineage-promiscuous. Lineage promiscuity is made even more likely because a common bi-potent precursor in the thymus gives rise to both T cells and NK cells.4 We therefore analyzed the patterns of NKRs in LBLs for comparison with the significance of NKRs in SNLs (Table IV).
We collected 9 consecutive cases, presenting with either a mediastinal mass or lymphadenopathy, with or without bone marrow involvement. All of these cases were CD3+ and TdT+. We found that 4 cases had a polyclonal pattern of TCR rearrangement and a restricted KIR repertoire, suggesting an NK lineage (NK-LBL). The remaining 5 cases had a monoclonal TCR rearrangement, suggesting a T lineage (T-LBL).
Unlike NK-SNLs, where the expression of CD94 could be confirmed by IHC, IHC for CD94 in 2 NK-LBLs that were CD94 by RT-PCR was negative. Furthermore, for LBLs, the lineage assignment to the T or NK lineage or the status of CD94 expression did not have a prognostic impact in this small series. Taken together, these differences strengthen the significance of NKR expression in SNLs.
Per ipher al T-cell lymphomas r ar ely expr ess KIRs, CD94 or NKG2
We also used 10 cases of peripheral T-cell lymphomas (PTCLs) for comparison with SNLs. Only one case had a detectable KIR repertoire, and another case had CD94 expression. This shows that NK differentiation in PTCLs is rare.
Rearrangement of T-cell receptor (TCR) or immunoglobulin (Ig) genes is important for the development of lymphocytes.32,33 TCR γ chain rearrangement takes place
prior to chain rearrangement, and Ig heavy chain rearrangement takes place prior to light chain rearrangement. This sequence of rearrangement can be used for
distinguishing immature from mature T or B cells. Despite the important role of antigen receptors, data on NK-cell antigen receptors, such as KIRs, CD94, and NKG2, are quite limited,34 in part due to the rarity of NK-cell lymphomas. We have collected 25 SNLs, one of the largest series, for molecular analysis.
Diagnosis of SNLs is traditionally based on the presence of pleomorphic CD56+ lymphoid cells in an angiocentric pattern and/or in association with EBV infection. The diagnosis, however, could be difficult when a low-grade lymphoid infiltrate mixed with reactive cells is found in a young patient, or when extensive necrosis leaves little viable tissue for diagnosis. To resolve these difficulties, we have advocated a restricted pattern of KIRs as a diagnostic tool. Most SNLs belong to the true NK lineage (NK-SNL). They are characterized by the presence of a restricted KIR repertoire without a monoclonal TCR rearrangement.15 Some SNLs belong to a mixed NK/T lineage (NK/T-SNL). They are characterized by the presence of both a monoclonal TCR rearrangement and a restricted KIR repertoire.16 In spite of its diagnostic value, the pattern of the KIR repertoire has little prognostic value.
To understand better the behavior of SNLs, we have turned to the analysis of the other NK-cell antigen receptors, CD94 and NKG2. In contrast to the KIRs, which belong to the immunoglobulin super-family, CD94/NKG2 are C-type lectins. More importantly, the developmental regulation of CD94/NKG2 is better defined than that of KIRs; CD94 was expressed first, followed by NKG2A, 2E, and 2C. This strongly suggests that the differential pattern of CD94 expression might subdivide NK-SNLs into a mature CD94+ subtype and an immature CD94- subtype.
Our data support the hypothesis that a division into the CD94+ and the CD94- subtypes has clinical significance, whether by the criteria of disease progression, 1-year survival, or survival curves (Table III and Figure 5). On the other hand, a correlation between outcomes and treatments is difficult to establish in a retrospective study of this sample size. The therapeutic regimens are usually adriamycin-based protocols such as CHOP or its variation, with or without local radiotherapy. Investigations into how the biological divisions into the CD94+ and the CD94- subtypes might influence the choice of therapeutic regimens are certainly of great clinical significance.
them died of rapid progression of the disease, often with distant metastases. In contrast, only 4 of the 10 CD94+ patients died; none of them had metastasis, and 2 of them died of hemophagocytic syndrome. Because recent data suggest that the hemophagocytic syndrome is often a reflection of underlying defect of NK cells,37 further investigation is required to determine whether essays of NK-cell function might be useful in predicting and preventing this fatal outcome.
In conclusion, we showed that CD94 is the most important prognostic factor in SNLs. This emphasizes again the importance of antigen receptors in diagnostic hematopathology. A CD94+ phenotype is consistent with that of a mature NK cell and implies an excellent prognosis. Finally, we showed that the occurrence of hemophagocytosis is unusual even in SNLs.
FIGURE LEGENDS
Figure 1. Str ucture of the NKG2 family and the RT-PCR str ategy The upper
part shows the consensus structure of the NKG2 family, including a 5’ untranslated region, an intracytoplasmic domain (cyt), a transmembrane domain (TM), an extracellular carbohydrate recognizing domain (CRD), and a 3’ untranslated region. The middle part shows alignment of the cDNA sequences, where the exon boundaries are marked by the vertical bars. The lower part shows sequences of the TM-CRD junction used for RT-PCR. The exon encompassing the TM-CRD junction spans nucleotides 448-501 in A, 294-338 in C, 580-619 in D, 332-376 in E, and 469-522 in F. The sequences used for the forward primers are in italicized letters. The consensus sequence, 5’-TGTGGCCCTTGTCCTGAA AAGTGGAT-3’, is used for the binding of reverse primer. The figure is drawn to show relative position only but is not proportional to exact sizes.
Figure 2. Two representative patter ns of RT-PCR analysis of CD94 and NKG2 family A CD94+ SNL expresses NKG2A strongly, and NKG2C and 2F weakly
(upper panel); a second SNL expresses CD94 only (lower panel).
Figure 3. Examples of KIR r eper toire by RT-PCR A KIR2DL4+ KIR2D- KIR3D- SNL is shown on the left side, another KIR2DL4- KIR2D+ KIR3D+ SNL is shown on the right side. The expected sizes of the PCR products for KIR2DL4, KIR2D, KIR3D, and actin are 83, 86, 90, and 96. The solid line is RT-PCR with reverse transcriptase; the dashed line is RT-PCR without reverse transcriptase, as negative control.
Figure 4. H/E sections and immunohistochemistr y of SNL Representative
examples of grade I (A) and grade III (B) nuclear morphologies. Note the smaller nuclei with condensed chromatin in A, and the larger nuclei with open chromatin in B. Not shown here, an intermediate morphology is grade II. An NK-SNL was
CD94 (C) and NKG2A (D) by immunohistochemistry. All at 1000X, H/E on paraffin sections (A and B) or immunoperoxidase stains on frozen sections (C and D).
Figure 5. Kaplan-Meier Sur vival analysis Survival curves for the CD94+ and CD94- subtypes are shown. The median survival times for the 2 groups were 60 months vs 10 months (p=0.026). An open circle indicates complete follow-up; a star indicates a censored case.
Table III. Clinical and pathologic compar ison among the 3
subtypes
Non-Progressive N=8 Progressive N=11 Spearman Correlation coefficient pMale: female ratio 6:2 10:1 0.22 0.38
Median age (year) 50 57 0.19 0.42
CD56+ 4 6 0.04 0.85 KIR2DL4+ 5 7 0.01 0.96 KIR2D+ 3 4 -0.01 0.96 KIR3D+ 3 4 -0.01 0.96 CD94+ 7 3 -0.59 0.007 NKG2A+ 3 4 -0.01 0.96 NKG2C+ 2 1 -0.22 0.38 NKG2D+ 0 0 NA NA NKG2F+ 2 0 -0.40 0.08
Early stage (I) 6 8 -0.01 0.97
WBC, 103/µl, edian 5.2 6.8 0.37 0.17
HB, gm/dl, median 12.1 12.6 0.05 0.83
PLT, 103/µl, median 234 237 -0.02 0.94
CD94+ N=10 CD94 -N=9 Mixed NK/T N=6 P (ANOVA) Sex (M: F) 8:2 8:1 2:4 0.047
Median age, year 43 57 26 <0.001
Nuclear grade I:II:III 6:1:3 1:5:3 3:1:2 0.42
Clear-cell change 3 3 0 0.31
Angiocentric necrosis 6 4 2 0.59
Reactive infiltrates 3 0 1 0.22
CD56+ 5 5 2 0.72
Early stage (I) 7 7 5 0.78
WBC, 103/µl, median 4.4 7.2 5.7 0.83 HB, gm/dl, median 12.7 12.1 12.8 0.83 PLT, 103/µl, median 205 263 276 0.51 LDH, U/L, median 413 660 461 0.45 Progression free 7 1 3 0.031 Survival >1 yr 8 2 4 0.031 Hemophagocytic syndrome 2 0 0 0.21
參考文獻(Reference)
1. Jaffe ES, Harris NL, Stein H, et al. World Health Organization classification of tumors. Pathology and genetics of tumors of haematopoietic and lymphoid tissues. IARC press, Lyon, France, 2001.
2. Jaffe ES. Classification of natural killer (NK) cell and NK-like T-cell malignancies. Blood. 1996;87:1207-1210.
3. Kwong YL, Chan AC, Liang R, et al. CD56+ NK lymphoma: clinicopathological features and prognosis. Brit J Haemat. 1997;97:821-829
4. Sanchez MJ, Spits H, Lanier LL, et al. Human natural killer cell committed thymocytes and their relation to the T cell lineage. J Exp Med. 1993;178:1857-1866.
5. Sanchez MJ, Muench MO, Roncarolo MG, et al. Identification of a common T/natural killer cell progenitor in human fetal thymus. J Exp Med. 1994;180:569-576.
6. Suzumiya J, Takeshita M, Kimura N, et al. Expression of adult and fetal natural killer cell markers in sinonasal lymphomas. Blood. 1994;83:2255-2260.
7. Phillips JH, Hori T, Nagler A, et al. Ontogeny of human natural killer cells: Fetal NK cells mediate cytolytic function and express cytoplasmic CD3e proteins. J Exp Med. 1982;175:1055-1066.
8. Lanier LL, Chang C, Spits H, et al. Expression of cytoplasmic CD3 ctivated human proteins in a
adult NK cells and CD3 complexes in fetal NK cells: Implications for the relationship of NK and T lymphocytes. J Immunol. 1992;14:1876-1880.
9. Natarajan K, Dimasi N, Wang J, et al. Structure and function of natural killer cell receptors: Multiple molecular solutions to self, nonself discrimination. Annu Rev Immunol. 2002;20:853-885.
10. Raulet DH, Vance RE, McMahon CW. Regulation of the natural killer cell receptor repertoire. Annu Rev Immunol. 2001;19:291-330.
11. Vilches C, Parham P. KIR: diverse, rapidly evolving receptors of innate and adaptive immunity. Ann Rev Immunol. 2002;20:217-251.
12. Long EO, Rajagopalan S. HLA class I recognition by killer cell Ig-like receptors. Semin Immunol. 2000;12:101-108.
13. Wilson MJ, Torkar M, Trowsdale J. Genomic organization of a human killer cell inhibitory receptor gene. Tissue Antigens. 1997;49:574-579.
14. Steffens U, Vyas Y, Dupont B, et al. Nucleotide and amino acid sequence alignment for human killer cell inhibitory receptors (KIR), 1998. Tissue Antigens. 1998;51:386-413.
15. Lin CW, Lee WH, Chang CL, et al. Restricted killer cell immunoglobulin-like receptor repertoire without T-cell receptor rearrangement supports a true NK-cell lineage in a subset of sinonasal lymphomas. Am J Path. 2001;159:1671-1679.
16. Lin CW, Yang JY, Chuang YC, et al. Presence of restricted killer immunoglobulin-like receptor repertoire and monoclonal T-cell receptor rearrangement as evidence of mixed NK/T-cell differentiation in a subset of sinonasal lymphomas. Lab Invest. 2003;83:55-64.
17. Lopez-Botet M, Perez-Villar JJ, Carreterro M, et al. Structure and function of the CD94 C-type lectin receptor complex involved in recognition of HLA class I molecules. Immunol Rev. 1997;155:165-174.
18. Takei F, McQueen KL, Maeda M, et al. Ly49 and CD94/NKG2A: developmentally regulated expression and evolution. Immunol Rev. 2001;181:90-103.
19. Lian RH, Maeda M, Lohwasser S, et al. Orderly and nonstochastic acquisition of CD94/NKG2 receptors by developing NK cells derived from embryonic stem cells in vitro. J Immunol. 2002;168:4980-4987.
20. Williams NS, Kubota A, Bennett M, et al. Clonal analysis of NK cell development from bone marrow progenitors in vitro: orderly acquisition of receptor gene expression. Eur J Immunol. 2000;30:2074-2082.
21. Kubota A, Kubota S, Lohwasswe S, et al. Diversity of NK cell receptor repertoire in adult and neonatal mice. J Immunol. 1999;163:212-216.
22. Perez-Villar JJ, Carretero M, Navarro F, et al. Biological and serological evidence for the existence of functionally distinct forms of CD94 NK cell receptors. J Immunol. 1996;157:5367-5374.
23. Mori KL, Egashira M, Oshimi K. Differentiation stage of natural killer cell-lineage lymphoproliferative disorders based on phenotypic analysis. Brit J Haemat. 2001;115:225-228. 24. Lin CW, Chang CL, Li CC, et al. Spontaneous regression of Kikuchi’s lymphadenopathy with
oligoclonal T-cell populations favors a benign immune reaction over a T-cell lymphoma. Am J Clin Path. 2002;117:627-635.
25. Chang C, Rodriguez A, Carretero M, et al. Molecular characterization of human CD94: a type II membrane glycoprotein related to the C-type lectin superfamily. Eur J Immunol. 1995;25:2433-2437. 26. Rodriguez A, Carretero M, Glienke J, et al. Structure of the human CD94 C-type lectin gene.
Immunogenetics. 1998;47:305-309.
27. Houchins JP, Yabe T, McSherry C, et al. DNA sequence analysis of NKG2, a family of related cDNA clones encoding type II integral membrane proteins in human natural killer cells. J Exp Med. 1991;173:1017-1020.
28. Plougastel B, Jones T, Trowsdale J. Genomic structure, chromosome location, and alternative splicing of the human NKG2A gene. Immunogenetics. 1996;44:286-291.
29. Adamkiewicz TV, McSherry C, Bach FH, et al. Natural killer lectin-like receptors have divergent carboxy-termini, distinct from C-type lectins. Immunogenetics. 1994;39:218-218.
30. Glienke J, Sobanov Y, Brostjan C, et al. The genomic organization of NKG2C, E, F, and D receptors in the human natural killer gene complex. Immunogenetics. 1998;48:163-173.
31. Nakamura F, Tatsumi E, Kawano S, et al. Acute lymphoblastic leukemia/lymphoblastic lymphoma of natural killer (NK) lineage: Quest for another NK-lineage neoplasm. Blood. 1997;89:4665-4666. 32. van Oers NSC. T cell receptor-mediated signs and signals governing T cell development. Seminar
Immunol. 1999;11:227-237.
33. Niro H, Clark EA. Regulation of B-cell fate by antigen-receptor signals. Nature Rev Immunol. 2002;2:945-956.
34. Haedicke W, Ho FCS, Chott A, et al. Expression of CD94/NKG2A and killer immunoglobulin-like receptors in NK cells and a subset of extranodal cytotoxic T-cell lymphomas. Blood.
2000;95:3628-3630.
35. Rajagopalan S, Long EO. A human histocompatibility leukocyte antigen (HLA)-G-specific receptor expressed on all natural killer cells. J Exp Med. 1999;189:1093-1099.
36. Miller JS, McCullar V. Human natural killer cells with polyclonal lectin and immunoglobulinlike receptors develop from single hematopoietic stem cells with preferential expression of NKG2A and KIR2DL2/L3/S2. Blood. 2001;98:705-713.
37. Stepp SE. Dufourcq-Lagelouse R. Le Deist F. et al. Perforin gene defects in familial hemophagocytic lymphohistiocytosis. Science. 1999;286:1957-1959.
計畫成果自評
請就研究內容與原計畫相符程度、達成預期目標情況
We have not yet established a dual lineage model. However, it turns out the presence or absence of CD94 is more important clinically. This finding, in a sense, does support the dual lineage model. But we have to modify and present the model in terms of CD94, rather than KIRs.
研究成果之學術或應用價值、是否適合在學術期刊發表或申請專利、主要發現或 其他有關價值等,作一綜合評估
A revised manuscript for the model is currently submitted to Blood.
可供推廣之研發成果資料表 無