The Joint Effect of Smoking and hOGG1 Genotype on Oral Cancer in
Taiwan
Chia-Wen Tsai
2,5,*, Ming-Hsui Tsai
1,2,*, Yung-An Tsou
1,2,*, Liang-Chun
Shih
1,2, Hsien-Chang Tseng
2, Wen-Shin Chang
2,5, Chien-Yi Ho
2,
Hong-Zin Lee
3and Da-Tian Bau
2,4,51
Department of Otolaryngology and
2Terry Fox Cancer Research Laboratory,
China Medical University Hospital, Taichung;
3School of Pharmacy, and
Graduate Institutes of
4Clinical Medical Science and
5Basic Medical Science,
China Medical University, Taichung, Taiwan, R.O.C.
* These authors contribute equally to this study
Correspondence to: Da-Tian Bau, Terry Fox Cancer Research Lab, China
Medical University Hospital, 2 Yuh-Der Road, Taichung, 404 Taiwan,
R.O.C. Tel: +886- 4-22053366 Ext 1523, Fax: +886 422053366 Ext 1543,
e-mail:
[email protected]
;
[email protected]
Running title: Tsai et al: hOGG1 genotypes in oral cancer
Key Words: hOGG1, single nucleotide polymorphism, oral cancer,
Abstract
This study aims at evaluating the association and interaction among hOGG1 genotypic polymorphism, smoking status and oral cancer risk in Taiwan. For this purpose, the well-known polymorphic variants of hOGG1, codon 326, was analyzed of its association with oral cancer susceptibility, and discussed of its joint effect with individual smoking habits on oral cancer susceptibility. In total, 620 patients with oral cancer and 620 healthy controls recruited from the China Medical Hospital were recruited and genotyped. The results showed that the hOGG1 codon 326 genotypes were differently distributed between the oral cancer and control groups (P=0.0266) and the C allele of hOGG1 codon 326 was significantly (P=0.0046) more frequently found in controls than in cancer patients. We have further analyzed the genetic-smoking joint effects on oral cancer risk and found an interaction between hOGG1 codon 326 genotypes and smoking status. The hOGG1 codon 326 genotypes were in association with oral cancer risk only in the smoker groups (P=0.0198), but not in the non-chewer group (P=0.8357). Our results provide the evidence that the C allele of
hOGG1 codon 326 may have a joint effect with smoking on the development of oral
Oral cancer is ranked the fourth most common cancer among Taiwan male, with the peak age at 55–59 years old, and is the leading type of cancer to cause death in the 40–50 age group. Three main factors, smoking, alcohol drinking, and betel quid chewing, are identified to be closely related to oral carcinogenesis. Smoking may generate reactive oxy-gen species (ROS) including superoxide anion radicals and hydrogen peroxide, which may further induce DNA single and double strand breaks. Sustained oxidative stress, such as smoking and betel quid exposure, induced oxidative DNA adducts to the human genome, and 8-hydroxy-2-deoxyguanine (8-OH-dG) seems to be the major form (1, 2). The 8-OH-dG is mutagenic which if not repaired on time, can cause severe transversions of GC to TA in several oncogenes and tumor suppressor genes and in turn lead to carcinogenesis (1, 2). Among the DNA repair pathways, 8-OH-dG and other oxidative DNA adducts are repaired by the base excision repair pathway (3). The human OGG1 (hOGG1) gene encodes a DNA glycosylase which catalyzes the cleavage of the glycosylic bond between the oxidized base and the sugar moiety, leaving an abasic apurinic/apyrumidinic site in DNA. The resulting apurinic/apyrumidinic site is then incised, and the repair is completed by successive actions of a phosphodiesterase, a DNA polymerase, and a DNA ligase (4).
Among the common single nucleotide polymorphisms (SNPs) of hOGG1 gene, the one located in the exon 7, resulting in an amino acid substitution of serine (Ser)
with cysteine (Cys) at codon 326 (Ser326Cys, rs1052133), has been demonstrated to affect the hOGG1 function (5). Those cells with CYS allele exhibited a reduced DNA repair activity (5), which has been reported to be associated with the risk of many types of cancers (6). In 2002, a study investigating the role of hOGG1 Ser326Cys genotypes in a Caucasian population was performed, but their sample size was rather small (case/control=169/338), and the samples of tumor sites are collected from the tonsil, tongue, floor of mouth, larynx and esophagus of the patients, not specific in oral cancer. In the present study, we aimed at analyzing the genetic polymorphisms of the hOGG1 Ser326Cys genotypes in a Taiwan oral cancer population (control/case=620/620), and investigated the interaction of hOGG1 Ser326Cys genotypes and smoking habits in a Taiwanese population.
Materials and Methods
Study population and sample collection. Six hundred and twenty cancer patients
diagnosed with oral cancer were recruited at the outpatient clinics of general surgery during 1998-2010 at China Medical University Hospital in central Taiwan. The clinical characteristics of patients including histological details were all graded and defined by expert surgeons (Dr. Tsai, Tsou, Shih and Tseng). All patients voluntarily participated and provided their peripheral blood for this research. The same amounts
of non-oral cancer healthy volunteers as the controls were selected by matching for their age, gender and habits after initial random sampling from the Health Examination Cohort of the hospital. The exclusion criteria of the control group included previous malignancy, metastasized cancer from other or unknown origin, and any familial diseases. Both groups completed a well-informed questionnaire which included the individual habits. Smokers were defined as daily or almost daily smokers, who had smoked at least five packs of cigarettes in their lifetime. Smokers were asked for the age of initiation, whether they were currently smoking or had already quit, and if so, when they had quit, and on average, how many cigarettes they smoked or had smoked daily. Our study was approved by the Institutional Review Board of the China Medical University Hospital and written-informed consent was obtained from all participants.
Genotyping assays. Genomic DNA was prepared from peripheral blood leukocytes
using a QIAamp Blood Mini Kit (Blossom, Taipei, Taiwan) and further processed according to previous studies (7-16). The polymerase chain reaction (PCR) cycling conditions were: one cycle at 94oC for 5 min; 35 cycles of 94oC for 30 sec, 55oC for
30 sec, and 72oC for 30 sec, and a final extension at 72oC for 10 min. Pairs of PCR
Statistical analyses. Only those with both genotypic and clinical data
(control/case=620/620) were included for the final analysis. To ensure that the controls used were representative of the general population and to exclude the possibility of genotyping error, the deviation of the genotype frequencies of hOGG1 codon 326 in the controls from those expected under the Hardy-Weinberg equilibrium was examined by the goodness-of-fit test. Pearson’s Chi-square test was performed to compare the distributions of the genotypes between case and control groups. Data were recognized as statistically significant when individual P-value was less than 0.05.
Results
The demographic distribution of characters of all the oral cancer patients and healthy controls are summarized in Table II. These characteristics of patients and controls are all well matched and none of the distribution for age, gender, or indulgence between the two groups was statistically significant (P>0.05) (Table II).
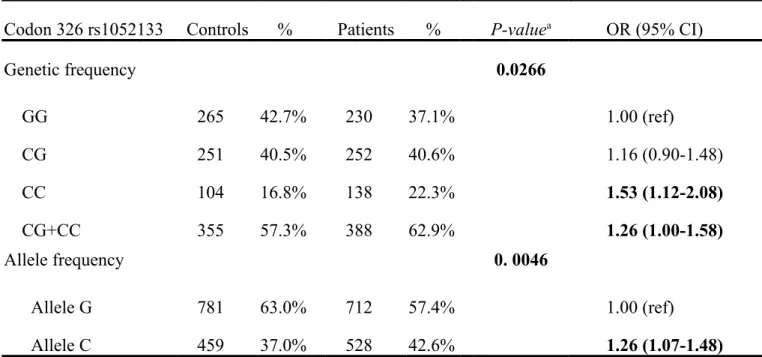
The frequencies of the genotypes for hOGG1 codon 326 in controls and oral cancer patients are listed and analyzed in Table III. The genotype distributions of
hOGG1 codon 326 were significantly different between oral cancer and control
controls and oral cancer patients are also shown in Table III, and the trend is more obvious. The data showed that the C allele of the hOGG1 codon 326 polymorphism was significantly associated with higher oral cancer risk (P=0.0046). It is more convincing to provide the results from multiple approaches so we have also performed the analysis of odds ratio for oral cancer risk among the variant genotypes. The odds ratio analysis showed that those who carry homologous CC have 1.53-fold higher oral cancer risk (95%CI=1.12-2.08), compared with those with homologous GG. A combination of CC+CG vs GG has similar higher risk compared with those with homologous GG (odds ratio=1.26, 95%CI=1.00-1.58). The conclusive finding deduced from the data in Tables III and IV is that the C allele of hOGG1 codon 326 seems to be associated with higher risk for oral cancer in Taiwan.
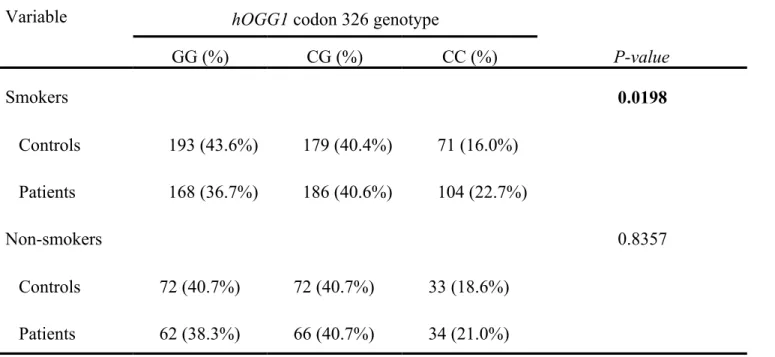
The interaction of genotype of hOGG1 codon 326 and the smoking habits was of our interest. The genotype distribution of various genetic polymorphisms of hOGG1 codon 326 was significantly different between oral cancer and control groups who have smoking habit (P=0.0198), while that for hOGG1 codon 326 was not significant (P=0.8357) (Table IV). Consistent with the findings in Table III, the C allele frequency was still significantly higher in cancer patients who have smoking habit than in smoking controls. There was no such difference in the non-smoking groups.
Discussion
In order to examine the role of hOGG1 in oral cancer, in this study, we selected the most important SNP of the hOGG1 gene, the codon 326 and clarify its association with the susceptibility for the oral cancer population of Taiwan, where the oral cancer density is the highest worldwide. We found that the C variant genotypes of hOGG1 codon 326 were significantly associated with a higher susceptibility for oral cancer (Tables III). As we supposed, the effects of the hOGG1 codon 326 on oral carcinogenesis are complex, exerting either an adverse effect or an advantageous influence on determining oral cancer risk.
Previous studies have implicated the hOGG1 codon 326 polymorphism in risk for smoking- and/or alcohol-related orolaryngeal cancer in USA (17). Significant increases in risk were found for the homozygous G/G genotype and orolaryngeal cancer in the Caucasian study (17), which not the same as ours. In addition, the risky G allele had obvious joint effects with both smoking and alcohol drinking on orolaryngeal carcinogenesis (17). Further, a significant positive association between
hOGG1 genotype and cancer risk was also observed foresophageal cancer (18). To compare the difference between Caucasian and oriental populations, we have also analyzed the association between hOGG1 codon 326 genotypes and oral cancer risk in patients and controls who have a smoking habit in Taiwan, an oriental population
(Table IV). Interestingly, the interaction between hOGG1 codon 326 and smoking habit is obvious, and the C allele is indeed the risky one (Table IV). Compared with the previous study, we have the larger population of both controls and cases for the further stratification with higher analysis power. In addition, during the sampling work we have recruited almost the same proportion of recruited cases and controls taking their individual habits into consideration (Table I). Furthermore, all the subjects investigated in this study are Taiwanese, much more genetically conservative than the one collected in USA. However, the worldwide conclusion about oral carcinogenesis may be concealed by variations among ethnics with different levels of exposure to environmental factors, such as smoking, and multi-institutional studies including different ethnic groups with more careful matching between cases and controls should be performed in the near future. We have also stratified the population by gender, and the findings in overall population were consistent in Taiwan males, but no significance was found in Taiwan females (data not shown). This may be due to the gender ratio is about nine to one for male to female, and the female sample size was limited.
We propose that the different genotypes of codon 326 may affect the hOGG1 enzymatic activity, slightly altering its normal function. Generally speaking, oxidative insults to genome DNA are continuously happening, which may be caused by both
endogenous oxidative stress and exogenous carcinogens. If the hOGG1 does not work well, the DNA adducts could be left unrepaired, leading to mutations or even carcinogenesis. In Taiwanese population, as these people with the C allele(s) get older, the alterations towards carcinogenesis may accumulate via the decreasing functions of hOGG1. There are several studies suggested our idea that the amino acid change in hOGG1 may affect the catalytic properties of the enzyme (19, 20). Indirect explanation for the functional relevance of the polymorphism is that the variant allele may be linked to other functional polymorphisms in hOGG1 involved in the removal of oxidative DNA damage. Direct explanation is that the variant genotype may be deficient in repair of oxidative DNA damage only under conditions of excessive cellular oxidative stress (19). However, both of the proposed hypotheses need to be further confirmed in the patient tissue cells.
In conclusion, oral cancer is high smoking-related and this is so far the largest study which focuses on the codon 326 of hOGG1 and their joint effects with smoking habit on oral cancer risk all over the world. Our results indicate that the C allele of
hOGG1 codon 326, interacts with smoking habits, may play an important role in the
oral carcinogenesis.
Acknowledgements
Yi-Shan Juan and the Tissue Bank at the China Medical University for their technical assistance. This study was supported by research grants from the Terry Fox Cancer Research Foundation, National Science Council (NSC98-2320-B-039-010-MY3), Taiwan Department of Health, China Medical University Hospital Cancer Research Center of Excellence (DOH101-TD-C-111-005) and China Medical University and Hospital (DMR-101-031).
References
1. Chen L, Elahi A, Pow-Sang J, Lazarus P and Park J: Association between polymorphism of human oxoguanine glycosylase 1 and risk of prostate cancer. J Urol 170: 2471-2474, 2003.
2. Xu J, Zheng SL, Turner A, Isaacs SD, Wiley KE, Hawkins GA, Chang BL, Bleecker ER, Walsh PC and Meyers DA: Associations between hOGG1 sequence variants and prostate cancer susceptibility. Cancer Res 62: 2253-2257, 2002. 3. Goode EL, Ulrich CM and Potter JD: Polymorphisms in DNA repair genes and
associations with cancer risk. Cancer Epidemiol Biomarkers Prev 11: 1513-1530, 2002.
4. Dianov GL, Souza-Pinto N, Nyaga SG, Thybo T, Stevnsner T and Bohr VA: Base excision repair in nuclear and mitochondrial DNA. Prog Nucleic Acid Res Mol Biol 68: 285-297, 2001.
5. Kohno T, Shinmura K, Tosaka M, Tani M, Kim SR, Sugimura H, Nohmi T, Kasai H and Yokota J: Genetic polymorphisms and alternative splicing of the hOGG1 gene, that is involved in the repair of 8-hydroxyguanine in damaged DNA. Oncogene 16: 3219-3225, 1998.
6. Weiss JM, Goode EL, Ladiges WC and Ulrich CM: Polymorphic variation in hOGG1 and risk of cancer: a review of the functional and epidemiologic literature. Mol Carcinog 42: 127-141, 2005.
7. Bau DT, Tsai MH, Lo YL, Hsu CM, Tsai Y, Lee CC and Tsai FJ: Association of p53 and p21(CDKN1A/WAF1/CIP1) Polymorphisms with Oral Cancer in Taiwan Patients. Anticancer Res 27: 1559-1564, 2007.
8. Chang CH, Chang CL, Tsai CW, Wu HC, Chiu CF, Wang RF, Liu CS, Lin CC and Bau DT: Significant association of an XRCC4 single nucleotide polymorphism with bladder cancer susceptibility in Taiwan. Anticancer Res 29: 1777-1782, 2009. 9. Chang CH, Chiu CF, Liang SY, Wu HC, Chang CL, Tsai CW, Wang HC, Lee HZ and Bau DT: Significant association of Ku80 single nucleotide polymorphisms with bladder cancer susceptibility in Taiwan. Anticancer Res 29: 1275-1279, 2009. 10. Chiu CF, Tsai MH, Tseng HC, Wang CL, Wang CH, Wu CN, Lin CC and Bau DT: A novel single nucleotide polymorphism in XRCC4 gene is associated with oral cancer susceptibility in Taiwanese patients. Oral Oncol 44: 898-902, 2008. 11. Chiu CF, Wang CH, Wang CL, Lin CC, Hsu NY, Weng JR and Bau DT: A novel
single nucleotide polymorphism in XRCC4 gene is associated with gastric cancer susceptibility in Taiwan. Ann Surg Oncol 15: 514-518, 2008.
12. Chiu CF, Wang HC, Wang CH, Wang CL, Lin CC, Shen CY, Chiang SY and Bau DT: A new single nucleotide polymorphism in XRCC4 gene is associated with
breast cancer susceptibility in Taiwanese patients. Anticancer Res 28: 267-270, 2008.
13. Hsu CF, Tseng HC, Chiu CF, Liang SY, Tsai CW, Tsai MH and Bau DT: Association between DNA double strand break gene Ku80 polymorphisms and oral cancer susceptibility. Oral Oncol 45: 789-793, 2009.
14. Hsu NY, Wang HC, Wang CH, Chiu CF, Tseng HC, Liang SY, Tsai CW, Lin CC and Bau DT: Lung cancer susceptibility and genetic polymorphisms of Exo1 gene in Taiwan. Anticancer Res 29: 725-730, 2009.
15. T seng HC, saiT MH, C hiu CF, W ang CH, C hang NW, H uang CY, T sai CW, L
iang SY, Wang CL and Bau DT: Association of XRCC4 Codon 247 Polymorphism with Oral Cancer Susceptibility in Taiwan. Anticancer Res 28: 1687-1691, 2008.
16. Yang MD, Hsu YM, Kuo YS, Chen HS, Chang CL, Wu CN, Chang CH, Liao YM, Wang HC, Wang MF and Bau DT: Significant Association of Ku80 Single Nucleotide Polymorphisms with Colorectal Cancer Susceptibility in Central Taiwan. Anticancer Res 29: 2239-2242, 2009.
17. Elahi A, Zheng Z, Park J, Eyring K, McCaffrey T and Lazarus P: The human OGG1 DNA repair enzyme and its association with orolaryngeal cancer risk. Carcinogenesis 23: 1229-1234, 2002.
18. Xing DY, Tan W, Song N and Lin DX: Ser326Cys polymorphism in hOGG1 gene and risk of esophageal cancer in a Chinese population. Int J Cancer 95: 140-143. 2001.
19. Lee AJ, Hodges NJ and Chipman JK: Interindividual variability in response to sodium dichromate-induced oxidative DNA damage: role of the Ser326Cys polymorphism in the DNA-repair protein of 8-oxo-7,8-dihydro-2'-deoxyguanosine DNA glycosylase 1. Cancer Epidemiol Biomarkers Prev 14: 497-505, 2005.
20. Yamane A, Kohno T, Ito K, Sunaga N, Aoki K, Yoshimura K, Murakami H, Nojima Y and Yokota J: Differential ability of polymorphic OGG1 proteins to suppress mutagenesis induced by 8-hydroxyguanine in human cell in vivo. Carcinogenesis 25: 1689-1694, 2004.
Table I. The primer sequences, polymerase chain reaction and restriction fragment length polymorphism (PCR-RFLP) conditions for hOGG1 gene polymorphisms.
Polymorphism (location)
Primers sequences (5’->3’) Restriction enzyme SNP sequence DNA fragment size (bp) Codon 326 (rs1052133) F: ACTGTCACTAGTCTCACCAG R: GGAAGGTGGGAAGGTG Fnu4HI 37 oC for 2 h C (Ser) G (Cys) 200 100 + 100 *F and R indicate forward and reverse primers, respectively.
Table II. Characteristics of oral cancer patients and controls
Characteristics Controls (n = 620) Patients (n = 620) Pa
n % Mean (SD) n % Mean (SD) Age (y) 51.3 (7.4) 52.4 (7.2) 0.78 Gender 1.00 Male 586 94.5% 586 94.5% Female 34 5.5% 34 5.5% Indulgence Cigarette smokers 443 71.5% 458 73.9% 0.37
Betel quid chewers 382 61.6% 399 64.4% 0.35
Alcohol drinkers 413 66.6% 441 71.1% 0.10
Site of main tumor
Tongue 301 48.5% Buccal mucosa 180 29% Mouth floor 36 5.8% Retromolar trigone 26 4.2% Alveolar ridge 17 2.7% Palate 14 2.3% Lip 12 1.9% Others 34 5.5%
Table III. Distribution of hOGG1 codon 326 genetic and allelic frequencies among oral cancer patient and control groups.
Codon 326 rs1052133 Controls % Patients % P-valuea OR (95% CI)
Genetic frequency 0.0266 GG 265 42.7% 230 37.1% 1.00 (ref) CG 251 40.5% 252 40.6% 1.16 (0.90-1.48) CC 104 16.8% 138 22.3% 1.53 (1.12-2.08) CG+CC 355 57.3% 388 62.9% 1.26 (1.00-1.58) Allele frequency 0. 0046 Allele G 781 63.0% 712 57.4% 1.00 (ref) Allele C 459 37.0% 528 42.6% 1.26 (1.07-1.48)
Table IV. Distribution of hOGG1 codon 326 genotypes in oral cancer patients after stratification by individual smoking habits.
Variable hOGG1 codon 326 genotype
GG (%) CG (%) CC (%) P-value Smokers 0.0198 Controls 193 (43.6%) 179 (40.4%) 71 (16.0%) Patients 168 (36.7%) 186 (40.6%) 104 (22.7%) Non-smokers 0.8357 Controls 72 (40.7%) 72 (40.7%) 33 (18.6%) Patients 62 (38.3%) 66 (40.7%) 34 (21.0%)