行政院國家科學委員會專題研究計畫 成果報告
台灣西部主要河口域及潟湖區橈足類多樣性及族群動態之
研究
計畫類別: 個別型計畫 計畫編號: NSC92-2313-B-110-002- 執行期間: 92 年 02 月 01 日至 93 年 01 月 31 日 執行單位: 國立中山大學海洋資源學系(所) 計畫主持人: 羅文增 共同主持人: 何汝諧,石長泰 計畫參與人員: 謝泓諺、徐培凱、余人堯 報告類型: 完整報告 處理方式: 本計畫可公開查詢中 華 民 國 93 年 7 月 13 日
行政院國家科學委員會補助專題研究計畫成果報告
台灣西部主要河口域及潟湖區橈足類多樣性及族群動態之研究
Biodiversity and population dynamics of copepods in the major
estuaries and lagoons in western Taiwan
計畫類別:■個別型計畫 □整合型計畫 計畫編號:NSC 92-2313-B-110-002 執行期間:92 年 02 月 01 日至 93 年 01 月 31 日止 計畫主持人:羅 文 增 共同主持人:石 長 泰 計畫參與人員:謝泓諺、徐培凱、余人堯 成果報告類型(依經費核定清單規定繳交):□精簡報告 ■完整報告 本成果報告包括以下應繳交之附件: □赴國外出差或研習心得報告一份 □赴大陸地區出差或研習心得報告一份 □出席國際學術會議心得報告及發表之論文各一份 □國際合作研究計畫國外研究報告書一份 處理方式:除產學合作研究計畫、提升產業技術及人才培育研究計畫、列管計畫 及下列情形者外,得立即公開查詢 □涉及專利或其他智慧財產權,□一年□二年後可公開查詢 執行單位:國立中山大學 海洋資源學系 中華民國 93 年 05 月 18 日
台灣西部主要河口域及潟湖區橈足類多樣性及族群動態之研究
—高屏溪河口域及大鵬灣浮游性橈足類種類組成及季節分布 執行單位:國立中山大學海洋資源學系 計畫編號:NSC 92-2313-B-110-002 計畫主持人:羅 文 增 共同主持人:石 長 泰 研究人員:謝泓諺、徐培凱、余人堯 聯絡方式:高雄市蓮海路 70 號 國立中山大學海洋資源學系 E-Mail: [email protected]
一、中文摘要
由 2003 年二月及八月於高屏溪河口域所採得樣品分析結果共發現浮游性橈 足類 23 科 46 屬 104 種,海水種類計有 17 科 27 屬 74 種及淡水種類則有 7 科 19 屬 30 種,總平均豐度為 1.3 ± 2.4 ×104 ind./m3 。高屏溪及東港溪的浮游橈足 類豐度明顯較高屏沿岸海域高出許多,不過橈足類種類數及種歧異度則以沿岸海 域相對較高。在優勢種方面,東港溪以橈足類無節幼生佔最大量,而高屏溪則以Pseudodiatomus marinus 及 Pseudodiatomus annandalei 居多,高屏沿岸海域
之優勢種類則為Temora turbinata及Acartia omorii。橈足類的豐度及優勢種
組成均有明顯的季節變化,二月海水及淡水種橈足類之平均豐度均明顯較八月高 出許多(分別達 73 及 17 倍);二月海水種以P. marinus數量最多(20.4%),淡 水種是以無節幼生數量最多(41.2%);八月海水種以 Corycaeus andrewsiz居多 (8.8%),淡水種以Mongolodiaptomus birulai數量最多(18.7%)。 大鵬灣之浮游橈足類共發現有 13 科 17 屬 55 種,平均豐度為 4.7 ± 6.4 × 104 ind./m3 。其中以Bestolina amyoensis的數量最多,佔總數量的 36.5%,而
Paracalanus crassirostris (17.7%)及Oithona oculata (17.0%)也甚為常見。
八月橈足類之平均豐度明顯較二月來的高。二月以 Acartia spp.及 Acartia sinjiensis 數量居多(分別佔總數量的 42.7%及 28.4%);八月以 B. amyoensis 為最優勢種(44.4%),而O. oculata 及P. crassirostris 亦不少。不同季節橈 足類豐度在測站間之分布趨向並不甚一致,但一般以靠近內灣的測站豐度較高, 而接近灣口的測站則相對較低。 高屏溪及東港溪之橈足類總豐度與溫度成顯著負相關,與鹽度及葉綠素a濃
度則無顯著相關性;前 6 個主要優勢種均與水文環境因子無顯著相關性。大鵬灣 之橈足類總豐度與鹽度成負相關,與溫度及葉綠素a濃度則無顯著相關性。主要 優勢種 B. amyoensis 及 P. crassirostris 分別與鹽度及葉綠素 a 呈顯著負相 關,O. oculata則與溫度呈顯著正相關與鹽度呈顯著負相關。 關鍵詞︰橈足類、高屏溪、東港溪、河口域、大鵬灣、季節分布、種類組成
Abstract
The species composition and distribution of planktonic copepods were investigated at 12 stations in Kaoping estuary and 6 stations in Tapong bay, southwestern Taiwan, in summer and winter of 2003. In all, 104 species of copepods belonging to 23 families and 46 genera were identified in Kaoping estuary, among these, 74 were seawater species and 30 belonged to freshwater species. The mean abundance of copepods was 1.3 ± 2.4 × 104
ind./m3
. Kaoping river and Tonkung river showed higher copepod abundance and lower species number and diversity than that in Kaoping coastal waters. Nauplii peaked in Tonkung river, Pseudodiatomus marinus and Pseudodiatomus annandalei dominated in Kaoping river, while Temora turbinate and Acartia omorii were abundant in coastal waters. The copepod abundance and species composition exhibited apparently seasonal variations, abundances of both freshwater and seawater species were up to 73 and 17 times higher in February than in August, respectively. In February, P. marinus (20.4%) was the most dominant seawater species and nauplii (41.2%) dominated in freshwater species; while in August, Corycaeus andrewsiz (8.8%) ranked first in seawater species and Mongolodiaptomus birulai (18.7%) was the most abundant freshwater species.
In Tapong bay, 55 species of copepods belonging to 13 families and 17 genera were identified, with an average abundance of 4.7 ± 6.4 ×104
ind./m3
. Among these species, Bestolina amyoensis was the most dominant taxon, contributing 37% to the numerical total of copepods. Other dominant copepods included Paracalanus crassirostris (17.7%) and Oithona oculata (17.0%). The average abundance of copepods in the bay was higher in August than in February. The species composition showed apparently seasonal difference, Acartia spp. (42.7%) and Acartia sinjiensiswere (28.4%) were very abundant in February, while B. amyoensis (44.4%), O.
oculata (20.6%) and P. crassirostris (18.9%) dominated in August.
Higher copepod abundances were generally found in the inner bay versus the lower abundance in the outer bay.
The copepod abundances of Kaoping estuary were significantly negative correlated with water temperature but showed no significant relationships with salinity and the concentration of chlorophyll a. Abundances of the 6 most dominant copepods showed no significant relationships with environmental variables. The copepod abundances of Tapong Bay were significantly negative correlated with salinity but showed no significant relationships with temperature and the concentration of chlorophyll a. Abundances of B. amyoensis and P. crassirostris were significantly negative correlated with salinity and the concentration of chlorophyll a, while O. oculata was significantly influenced by temperature and showed significantly negative relationship with salinity.
Keywords: Copeods, Kaoping river, Tonkung river, Estuary, Tapong bay, Seasonal distribution, Species composition.
二、前言:
橈足類在海洋浮游動物群聚中經常是最優勢的類群,其在海洋生態食物鏈 中是屬於次級生產者的角色,大多數以浮游植物為食(Nybakken, 1997; Pechenik,
1991),浮游橈足類亦為其他海洋較高營養階動物之食物,故其在海域中之分布 情況亦會直接影響魚類資源動態。綜合以上所言,足可知浮游橈足類在海洋食物 鏈中對於物質與能量之傳輸,實占有不可或缺之地位。 海洋浮游性橈足類群聚組成之消長及分布易受環境因子變化之影響,尤其 是在陸海河交界水團變動頻繁之沿岸海域。當海域環境狀況或條件改變時,生物 群聚組成經常就會出現一定程度的變異;因此在不同的水團或環境條件下,自然 就會出現一些不同的種類組成,例如 Calanus finmarchicus是溫寒帶大洋常見
的種類(Colebrook, 1982),Acartia tonsa常在溫帶河口地區出現(Cervetto et
al., 1999);而Eucalanus bungii bungii則是北太平洋親潮的外洋冷水域中特
有種之一(Yamaji, 1991)。地區性氣壓變動所造成的季風亦會影響橈足類的分 布 , 例 如 Stephens et al. (1998) 對 於 北 大 西 洋 震 盪 (North Atlantic
Oscillation, NAO)與C. finmarchicus的分佈情形進行研究,並指出 NAO 的方
向會影響到海流流向,間接使得C. finmarchicus的分布擴散至某些海域。在國
內早期關於橈足類之研究十分缺乏,僅有 Tan (1967) 及 Tseng (1972)進行概略 性的研究;近幾年來才有較多的學者參與,例如,Shih and Young (1995) 首次 收集並編輯台灣附近海域橈足種類之區系名錄,其他有關橈足類之研究包括有: 日週性垂直遷移(Su, 1996)、與仔稚魚之相關性(Cheng, 1998)、種類多樣性與 水團的關係 (Shih and Chiu, 1998)、河口域之分布(Hsieh and Chiu, 1998) 及其與季風之關係(Chen, et al., 1998)等,均已陸續被發表。 台灣海峽北連東海大陸棚,南接南海大陸棚,為兩海域間唯一通道。海域內 之地形十分複雜,海峽南邊為水深 20 公尺之台灣淺灘,其南緣為大陸棚斜坡斜 入南海盆地;淺灘北側為澎湖群島,其與台灣本島間隔著一條南北走向之澎湖水 道,其平均寬度為 25 公里、深度由南向北遞減;澎湖水道向北延伸約 20 公里處, 則為一條東西走向之海底隆起(雲彰隆起,Yuen-Chang Ridge),水深約為 30 至 40 公尺。這些地形特徵與夏及冬季之季節風(西南季風及東北季風)的交互影 響下,主控海峽內台灣海峽水(或南海水)與大陸沿岸水兩種不同性質水團之推 移而呈現季節性勢力消長現象(Fan, 1982; Zhang, 2000; Jan et al., 1994、
1995 and 1998),春季時由於東北季風減弱且西南季風逐漸形成,使得黑潮支流
受西南季風推移沿台灣西部沿岸,經澎湖水道向北流動,至雲彰隆起處與大陸沿 岸流程對峙形態;夏季西南風盛行時,南海表層水受季風推移而沿著台灣西部海
岸線向北流至台灣東北部海域,而大陸沿岸流則退出海峽;秋季西南季風逐漸減 弱、東北季風逐漸形成時,大陸沿岸水及東海陸棚水受季風吹送而向南流動,致 使台灣海峽內的暖水逐漸向南退移;而冬季東北季風盛行時,黑潮支流由澎湖水 道進入海峽內與大陸沿岸留在於雲彰隆起附近相遇,並於隆起處形起湧昇現象 (郭,1994)。 台灣西南沿岸海域位於北回歸線以南,西北方有澎湖水道連接台灣海峽, 東南方有巴士海峽與太平洋連接,西南方為南海,高屏峽谷則是由高屏溪口向西 南延伸至南中國海盆,因此高屏海域的水文特性主要是受南海及附近沿岸河川淡 水所控制 (台南水工試驗所,1993)。大鵬灣為台灣西南部的大型潟湖之一,根 據劉(2000)的研究指出,大鵬灣屬於一淺水且半封閉型的潟湖,平均水深為 2.2 公尺,水體積為 9.7×106 立方公尺,面積為 4.4×106 平方公尺,其北邊有一出口 與外海連接且有一小的魚塭排水溝注入,在其東邊有牛埔溪支流注入,另外在其 東南亦有一大排水溝(主要收集林邊鄉及大鵬灣附近魚塭排放之廢水),均會對 大鵬灣內水質變化產生影響。 因此,本研究之目的除在了解不同季節淡水河口、高屏溪口和東港溪口及 大鵬灣之浮游動物種類組成及豐度的差異外,最主要的目的是要了解淡水沿岸海 域、高屏沿岸海域、淡水河、高屏溪、東港溪及大鵬灣浮游性橈足類之種類組成 及豐度在測站上或季節性的分佈狀況,並探討其與水文環境因子之相關性。由於 浮游動物及浮游橈足類的鑑種及計數工作相當繁重,在人手不足的情況之下,僅 完成浮游動物大類計數及部分浮游橈足類種類鑑定,所以目前僅就完成部分描述 其結果及加以討論。 三、採樣及研究方法: A. 浮游動物之採樣方法 本研究採樣區域有三個(Figure 1、Figure 2),茲分述如下:(1)2003 年四季(2、5、8、11 月)於台北縣淡水河 12 個測站(河口 6 個,河內 6 個) 進行採樣(2 月及 5 月因河道潮汐水淺因素,分別缺少 5 個及 1 個測站樣品), 共計採得 42 個樣品;(2)2003 年四季(2、5、8、11 月)於屏東縣高屏溪及東 港溪 12 個測站(河口 4 個,河內 8 個)進行採樣(五月及八月亦因河道水深因 素,分別缺少 1 個及 2 個樣品),共計採得 45 個樣品;(3)2003 年偶數月(2、
4、6、8、10、12 月)於屏東縣大鵬灣 6 個測站漲、退潮時進行採樣,共計採得 72 個樣品。 採樣方法是以北太平洋標準網(網口直徑 45 ㎝,網長 180 ㎝,網目 200 μm)進行近水表層之水平拖網 3 分鐘,採得的樣品裝入 1 公升保存瓶,以 5%濃 度之中性福馬林保存,每次採樣前亦使用自攜式溫、鹽儀測定海水之溫度及鹽 度,並記錄天候海況資料;另外亦採取一公升表水樣品置入暗瓶冷藏,俾便進一 步測定水體葉綠素a之濃度。 B.浮游動物之鑑定及計數 浮游動物大類之鑑定及計數是先以分樣器將中性福馬林保存之浮游動物樣 品視量分成 1/2~1/8 之子樣品後,再置於解剖顯微鏡下計數各主要組成大類的數 量。橈足類的鑑定及計數則取約含 500 個橈足類個體之子樣品,置於光學顯微鏡 下進行鑑定,並儘可能鑑定至種或屬。 C.資料分析 本研究除了依目的進行基礎之圖表分析外,亦計算橈足類之種歧異度以及利 用複迴歸分析法來探討水體溫度、鹽度及葉綠素a等環境因子與橈足類豐度間的 相關性。 D. 葉綠素a濃度測定 水樣葉綠素a濃度測定,是以濾膜(孔徑 0.45μm,濾膜半徑 20mm)過濾 500ml 的海水後,將濾膜置入 5ml 之 90%的丙酮內,並放入-20℃之恆溫冰箱蔽光保存 24 小時,使其充分萃取葉綠素後,以離心機速率 3300rpm 離心 20 分鐘,取上層 液以螢光光度計於加酸前後(0.1N HCl) 測量吸光值,再計算得到葉綠素a濃度。 四、結果: 水文環境因子變化 淡水河河口水表層溫度有明顯的季節變化(Figure 3),一年四季中,以八 月之溫度最高,一般介於 28~31℃之間,二月最低,為 19~20℃,相差約 10℃。 各 測 站 間 以 八月 及 十一 月 之 變 動 較大 , 最高 低 溫 分 別 介於 27.2~32.3 ℃ 及 22.6~25.9℃之間,而在二月及五月各測站間則無明顯差異。在鹽度方面,淡水 河河口水表層鹽度無明顯季節變化情形,各測站的變化亦不甚一致。6 個河口測 站中,除了十一月之測站 C6(29.6‰)較低,及十一月之測站 C4(37.0‰)較
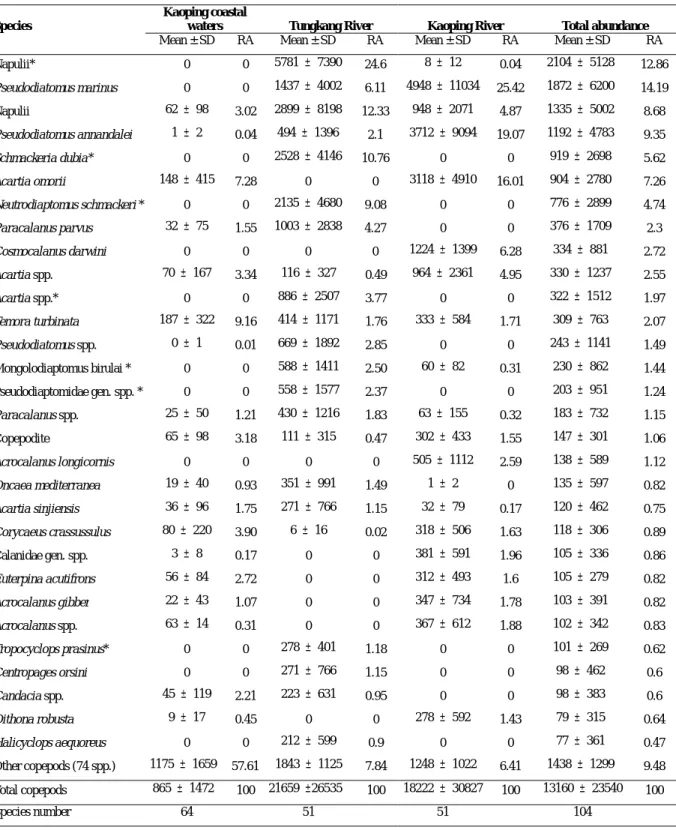
高外,大致介於 30~35‰之間;而 5 個河內測站除 2 月之測站 T1(24.0‰)及 T2(21.3‰)、五月之測站 T5(24.7‰)及八月之測站 T5(14.8‰)明顯較當月 低之外,其他幾個河內測站與河口測站之鹽度差異不大。 高屏溪及東港溪水文溫度及鹽度的變化情形,如 Figure 4 所示。溫度方面, 以八月最高(30℃左右)二月最低(25℃左右),同一季節各測站間的溫度差異 相對並不大。鹽度方面,二月及五月時,河口沿岸測站(C1~C4)與高屏溪河內 測站(K1~K3)的差異較小,而東港溪河內測站(T2~T5)則相對較低;八月時, 除河內測站明顯較低之外,河口測站 C3 及 C4 亦相對較低,分別為 25‰及 21‰; 十一月時,河口測站測站 C1 及 C2 較測站 C3 及 C4 來得高一些,而河內測站均明 顯較低。 大鵬灣的溫度有明顯的季節變化,表層海水平均溫度以八月最高(30.4℃), 十二月最低(22℃左右),相差約 8℃(Figure 5);但是同一月份各測站間的差 異並不大。鹽度方面以六月最低(平均 29.5‰),二月最高(34.5‰);八月測 站間的變動情形相對較其他月份大,四月各測站間的差異則相對最小。 浮游橈足類種類組成及季節變化 由於橈足種類鑒種分類計數工作十分繁重,以致淡水河樣品及部份之高屏溪 及大鵬灣樣品尚未完成,茲謹針對以分析完成的結果進行報告。由 2003 年二月 及八月於高屏溪及東港溪所採得樣品分析結果(Table 1),共發現浮游性橈足類 23 科 46 屬 104 種(包括鑑定至科或屬之未知種及幼生個體),總平均豐度為 1.3 ± 2.4 ×104 ind./m3 。其中,高屏溪及東港溪的浮游橈足類豐度分別為 1.8 ± 3.1 ×104 ind./ m3 及 2.2 ± 2.6 ×104 ind./ m3 ,明顯較高屏沿岸海域(0.9 ± 1.4 × 103 ind./m3 )高許多,相差約 20 倍左右。而三個水域之浮游橈足類優勢種類也 不甚相同,前 5 個主要優勢種中,分別於高屏溪或東港溪有大量出現的現象,如 第一(淡水 Napulii)、第三(海水 Napulii)及第五(Schmackeria dubia)優 勢種或幼生,分別於東港溪大量出現,其中第一優勢豐度的淡水種 Napulii 為 58±74 ×103 ind./m3 ,佔東港溪浮游橈足類總豐度的 24.6%,另兩個優勢種也分 別 佔 了 12.3% 及 10.8% ; 而 第 二 (Pseudodiatomus marinus ) 及 第 四 (Pseudodiatomus annandalei)優勢種,則多出現於高屏溪,其豐度分別為 4.9 ±11.0 ×103 ind./m3 (25.4%)及 3.7 ± 9.1 ×105 ind./m3 (19.1%)。高屏沿岸海
域之優勢種類為Temora turbinata(0.2 ± 0.3 ×103 ind./m3 , 9.2%)及Acartia omorii(0.1 ± 0.4 ×103 ind./m3 , 7.3%),但其數量相對於總平均豐度來說均不 高,僅分別名列總豐度的第 12 及第 6 優勢種。 季節變化方面,海水種橈足類二月之平均豐度為 1.7 ± 2.8 ×104 ind./m3 , 八月僅 0.2 ± 0.2 ×103 ind./m3 ,相差達 73 倍。其中,二月以Pseudodiatomus marinus 數量最多(3.4 ± 8.3 ×103 ind./m3 ),佔總數量的 20.4%,Napulii 居 次,佔 14.5%,其他為Pseudodiatomus annandalei(13.0%)、Acartia omorii (9.8%)、Paracalanus parvus(4.1%)及Cosmocalanus darwini(3.6%),這
前 6 個主要優勢種佔橈足類總量的 66%;八月以Corycaeus andrewsiz豐度最高,
為 0.2 ± 0.2 ×102
ind./m3
,佔總數量的 8.8%,其次為 Acrocalanus gibber,
佔了 7.7%,其他幾個優勢種類為Farranula concina(6.1%)、Farranula gibbula (5.2%)、Oncaea venusta(4.4%)及 Napulii(3.8%)。淡水種橈足類二月之平 均豐度為 0.9 ± 1.6 ×104 ind./m3 ,八月為 0.5 ± 0.7 ×103 ind./m3 ,相差了 17 倍。其中,二月是以 Napulii 數量最多(3.8 ± 6.6 ×103 ind./m3 ),佔總數量的 41.2%,Schmackeria dubia 次之,佔總數量的 18.1%,此外 Neutrodiaptomus
schmackeri亦不少,佔 15.3%,其次為Acartia spp.(6.4%)、Pseudodiaptomidae
gen. spp.(4.0%)及 Mongolodiaptomus birulai(3.6%),這前 6 個主要優勢
種佔橈足類總量的 89%;八月以Mongolodiaptomus birulai數量最高,為 100 ±
129 ind./m3
,佔總數量的 18.7%,依次為Paracyclopina nana(8.2%)、Mesocyclops leuckarti(7.7%)、Diaptominae gen. spp.(7.6%)、Paracyclopina fimbriatus (7.2%)及 Napulii(6.8%)。 測站變化方面,海水種前 6 優勢種類於各測站的變化情形由 Figure 6 可以 發現,多以高屏溪測站測站 K3 及東港溪測站測站 T1 的豐度較高,如第一、第三 及第四優勢種均以測站 K3 數量最多,而第二及第五優勢種則是分別於測站 T1 數 量最高,第六優勢種僅出現於高屏溪河內測佔,其中以測佔 K2 最多。淡水種橈 足類變化情形方面(Figure 7),前 6 個主要優勢種大多於二月時出現於東港溪 測站,高屏溪測站則甚少或無。其中第一優勢種 Nauplii 以測站 T3 數量最多, 而測站 T2、T4 及 T5 亦不少;第二優勢種亦有類似情形,除了測站 T3 外,均只 在二月出現於東港溪測站;第三、第四及第六優勢種,均僅於二月時出現於測站 T2 或 T3,其他測站則沒有發現;第五優勢種二月時雖然僅出現於測站 T3,但其
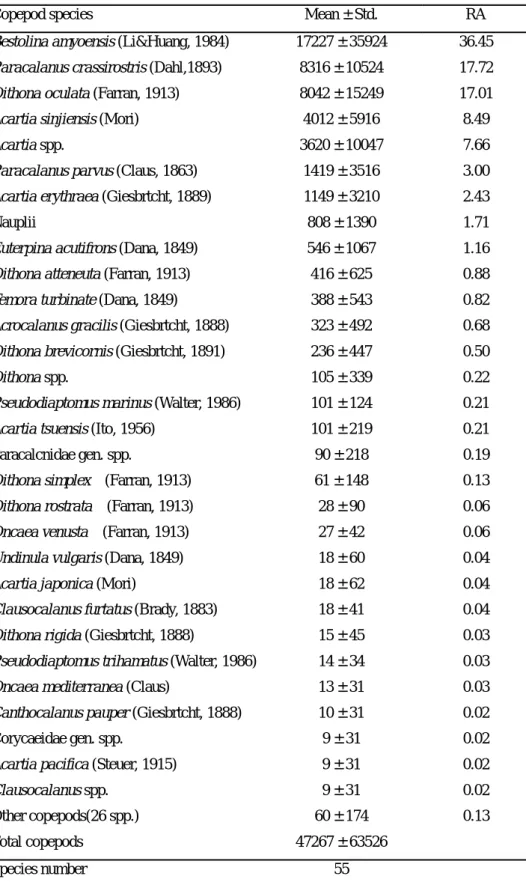
豐度明顯較八月時出現的幾個測站來的高。 在不同測站之豐度、種類數及種歧異度的變化情形(Figure 8),海水種橈 足類二月之豐度變化以測站 K3 最高,測站 T1 次之,分別為 8.1 ×103 ind./m3 及 7.1 ×103 ind./m3 ,明顯較其他測站高許多;八月之變動情形則不大,但以沿 岸測站的豐度相對較高。種類數方面,二月時沿岸 4 個測站差異不大(15~20 種), 其中以測站 C3 及 C4 種類數最多(20 種),而東港溪測站種類相對較少(0~14 種), 其中測站 T3、T4 及 T5 均沒有發現;八月之變動情形較大,以測站 C2 及 C4 種類 最多(23 種),測站 T3 最少(沒有發現)。種歧異度方面,二月之變化情形與種 類數變化情形相類似,沿岸 4 個測站相差不大,介於 3.5~4.0 之間,東港溪測站 則較低;8 月時以測站 C2 的種歧異度最高,為 4.1,測站 T2 及 T3 最低,為 0。 淡水種橈足類部分,二月時僅出現於東港溪測站,沿岸測站及高屏溪測站均沒有 發現,其中以測站 T3 豐度最高,為 5.1 ×103 ind./m3 ;八月除測站 C1 及 C2 外 均有發現,其中以東港溪測站 T2 最高,而高屏溪測站則有愈往上游豐度愈高的 現象。種類數方面,二月在 5 個有淡水種橈足類出現的東港溪測站中,種類數有 愈往上游愈高的趨勢;八月之變化情形則不甚一致,在有淡水種橈足類出現的測 站中,以測站 T3 的種類數最多(11 種)。種歧異度方面,二月時東港溪 5 個測 站,測站 T1 的種歧異度為 0,測站 T2~T4 的差異則不大,介於 2.0~2.2 之間; 八月時以測站 T2 及 T3 的種歧異度最高,為 3.3。 大鵬灣之浮游橈足類部份,由 2003 年偶數月採得的樣品分析結果(Table 2),共發現浮游性橈足類 13 科 17 屬 55 種(包括鑑定至科或屬之未知種及幼生 個體),總平均豐度分別為 4.7±6.4 ×104 ind./m3 。常見的優勢種類中以Bestolina amyoensis 的數量最多,為 1.7 ± 3.6 ×104 ind./m3 ,佔總數量的 36.5%,
Paracalanus crassirostris 居次,佔總數量的 17.7%,此外 Oithona oculata
(17.0%)、Acartia sinjiensis(8.5%)、Acartia spp.(7.7%)及Paracalanus parvus(3.0%)也較為常見,這前 6 個主要優勢種佔橈足類總量的 90%。八月時 橈足類之平均豐度(7.8 ± 7.8 ×104 ind./m3 )明顯較 2 月(1.7 ± 2.4 ×104 ind./m3 ) 來的高。二月以Acartia spp.數量最多(0.7 ± 1.4 104 ind./m3 ),佔總數量的
42.7%,其他幾個優勢種類依序為Acartia sinjiensis (28.4%)、Paracalanus
crassirostris (12.3%)、Nauplii(6.3%)、Acartia erythraea(1.6%)及Temora
amyoensis 豐度最高,為 3.5 ± 4.6 ×104
ind./m3
,佔總數量的 44.4%,其次為
Oithona oculata(20.6%),Paracalanus crassirostris(18.9% )、Acartia
sinjiensis(4.1%)、Paracalanus parvus(3.6%)及Acartia erythraea(2.6%) 亦較為常見,合計佔該月總豐度的 94%。
前 6 個優勢種類橈足類豐度在不同季節及測站間變化方面(Figure 9),B.
amyoensis、P. crassirostris、O. oculata及P. parvus於八月之豐度明顯高
於二月,而 A. sinjiensis 及 Acartia spp.則以二月出現數量較多。不同種類
於各測站的豐度變化情形不甚一致,但除了P. crassirostris及P. parvus外,
大致呈現出內灣測站(4、5、6)高於接近灣口測站(1、2、3)的現象。其中,
B. amyoensis及O. oculata在 8 月時多出現於內灣測站,二月時則甚少或無;
P. crassirostris 於八月時出現於接近灣口及灣中測站,二月之數量相對較低
並有由灣口向內灣遞減的趨向;A. sinjiensis及Acartia spp.於二月時多出現
於測站內灣測站,且均以測站 5 之豐度最高;P. parvus多於八月出現,且以灣 中測站豐度最高。 二月及八月份橈足類在不同測站之豐度分布情形並不一致(Figure 10), 二月時以測站 5 的豐度最高,測站 3 最低,兩者相差了 23 倍之多;八月則以測 站 4 數量最多,測站 5 及測站 6 數量亦不少,而測站 1、2 則明顯相對較低。種 類數方面,二月時除了於測站 3(32 種)種類數明顯較多之外,其他幾個測站差 異不大(12~18 種);八月時各測站之種類數差不多,介於 13~17 種之間。種歧 異度方面,兩個月份均以測站 3 及測站 4 的種歧異度較高,但各測站間的差異不 大,分別介於 1.4~2.7 及 1.1~2.2 之間。 浮游橈足類豐度與環境因子相關性 由複迴歸分析結果(Table 3)得知,高屏溪、東港溪之橈足類總豐度與溫 度成顯著負相關(P<0.05),與鹽度及葉綠素a濃度則無顯著相關性。前 6 個主 要優勢種,均與水文環境因子無顯著相關性。 大鵬灣之橈足類總豐度與鹽度成顯著負相關(P<0.05),與溫度及葉綠素 a 濃度則無顯著相關性(Table 3)。前 6 個主要優勢種中,B. amyoensis 與鹽度 呈顯著負相關, P. crassirostris與葉綠素a濃度呈顯著負相關,而O. oculata 與溫度呈顯著正相關與鹽度呈顯著負相關,其他 3 個優勢種類則與三種環境因子
無顯著相關性。
五、討論: 高屏溪及東港溪
高屏海域水文特性主要受到黑潮支流(高溫高鹽)及南海水(高溫低鹽)季 節性消長以及季風強弱作用所影響(Fan and Yu, 1981; Fan, 1982;王和陳, 1987; Shaw, 1989;吳,1996)。夏季時,西南季風盛行,推動高溫低鹽的南海 表層水進入台灣海峽;秋末(九月後)由於東北季風逐漸增強,致推動高屏沿岸 流往南移動,此時高溫高鹽黑潮支流經由台灣海峽入侵高屏海域,並在冬季時達 到最盛期;直到春季(二月後)東北季風逐漸緩和,黑潮支流入侵漸停止,海峽 東南部暖水團才有機會往北移動。高屏海域除受到季節性水團變動影響外,氣候 亦是影響因素之一。每年十月至隔年四月平均月降雨量經常低於 50 mm,故被稱 為乾季;而五月至九月平均月下雨量大多高於 100 mm,故稱為濕季。濕季來臨 時,西南季風逐漸增強經常帶來充沛降雨量,亦帶動低鹽南海表層水團進入高屏 海域一帶,使得濕季時該海域經常有低鹽高溫的水團特徵(詹,1995;吳,1996; 張,2000)。 由高屏沿岸海域二月(冬季)及八月(夏季)所得到的結果發現,浮游橈足 類豐度於二月較高,八月較低(Table 1、Figure 8),而羅(2002)在高屏海域 研究所得結果是以夏秋季豐度較高,冬季時較低,與本次研究呈現相反的情形。 但根據朱等(1993)在福建東山灣浮游動物之分佈研究發現,在冬季及夏季均有 高豐度值,並以橈足類所佔比例最高,而此浮游動物豐度之所以出現雙高峰,可 能因該地區不同季節沿岸高低溫水團的變化,造成夏季及冬季浮游生物族群消長 所致,此現象或可解釋高屏海域浮游性橈足類豐度變化在不同季節或年間的差 異。 除此之外,本次的研究尚包含高屏溪及東港溪的測站,根據結果發現,高屏 溪及東港溪之橈足類豐度明顯高於高屏沿岸海域,在橈足類總豐度前 5 個主要優 勢種中(Table 1),分別於高屏溪或東港溪有大量繁生的現象,進一步分析發現, 這 5 個主要優勢種類多出現於接近河口的測站,包含了高屏溪測站 K3、東港溪 測站 T1 及測站 T3。而由藍等(2002)在淡水河鄰近海域的橈足類研究中發現,
橈足類豐度以淡水河口附近測站最高,並推測可能是受淡水河注入影響,使河口 水域含有高量營養鹽及基礎生產量,並造成浮游動物大量繁生,這個結果與本研 究相類似。然而,由複迴歸分析結果顯示葉綠素a濃度與浮游橈足類豐度之間無 顯著相關性,此或許是浮游橈足類的分布亦同時受到多項水文環境因子交互作用 的影響,但仍待進一步的探討。 大鵬灣是個高度優養化的亞熱帶半封閉型潟湖,之前由於水產養殖過度地開 發利用,加上灣內外水體的交換僅靠單一潮口進出,以致於底質沈積物裡的有機 物含量明顯由潮口向灣內遞增。根據洪(2000)的研究報告(蚵架拆除前)顯示, 大鵬灣高度優養化及高基礎生產力的重要原因可能是水域系統磷過剩所造成,但 由於灣內水體面積不大,其水文及水質狀態易受到海水潮汐、降雨量及廢水排放 等影響,而有明顯的季節性及測站間的變化,本研究期間(蚵架已於 2002 年 10 月拆除)採樣分析結果亦印證了上述的論調。由海水溫、鹽度變化情形可知 (Figure 5),大鵬灣除了冬季外,一般水溫都能達 26℃以上,這可能是位於亞 熱帶地區及受到黑潮支流及南海水所彙集的沿岸水團影響所致,而各測站間溫度 差異大致呈現出內灣測站略高於外側(近灣口)測站的現象。此外,根據中央氣 象局以往的降雨資料顯示(交通部中央氣象局, 2000),大鵬灣地區有明顯的乾 濕季節,在夏季(六~九月)時之平均降雨量及天數都較其他月份高,而本研究結 果亦顯示大鵬灣在夏季時鹽度較其他月份為低。大鵬灣這種明顯的季節性差異, 可能是造成灣內水域橈足類群聚組成及數量在測站間及季節性消長的重要原因 之一。 由浮游橈足類於各測站的分布情形來看,不論是二月或八月均呈現出內灣測 站(測站 4、5、6)高於外側測站(測站 1、2、3)的現象。其中,二月時之所 以會有內側測站高於外側測站的現象,主要是因為同為 Acartia 屬的 Acartia
sinjiensis 及 Acartia spp.大量出現所導致,而八月時則是因為 Bestolina
amyoensis 及 Oithona oculata 大量繁生所造成。將本次研究之優勢種類與羅
(2000)在同一海域中所發現的優勢種類比較後發現,其主要優勢種類卻略有所 不同,在羅(2000)的研究中,是以A. tsuensis、Acartiidae 幼生體、Paracalanus
crassirostris、Paracalanidae 幼生體及 O. dissimilis 為主要優勢種類。而
本次研究與羅(2000)的研究除在測站方面稍有不同之外,最大的差異在於蚵架 的有無,因此蚵架拆除是否是造成本次研究與先前研究差異的主要原因,仍須在
未來的研究中作更進一步的探討。
參考文獻
Cervetto, G., Gaudy R. and Pagano, M. (1999). Influence of salinity on the distribution of Acartia tonsa (Copepoda, Calanoida). J. Exp. Mar. Biol. Ecol., 239: 33-45.
Chen, Q. C., Hwang, J. S. and Wong, C. K. (1998). The characteristics of zooplankton in the northern coastal water of Taiwan during the initial period of NE monsoon. J. Taiwan Strait (Taiwan Haixia) 17: 87-90 (in Chinese. With English summary).
Cheng C. C. (1998). The distributional correlation between planktonic copepods and ichthyoplankton in the western North Pacific. MSc thesis, National Taiwan Univ., Taipei.
Colebrook, J. M. (1982). Continuous plankton records: seasonal variations in the distribution and abundance of plankton in the North Atlantic Ocean and North Sea. J. Plankton Res., 4: 435-462.
Fan, K. L. (1982). A study of water masses in Taiwan Strait. Acta Oceanogr. Taiwanica, 13: 140-153.
Fan, K. L. and Yu, C. Y. (1981). A study of water masses in the seas of southernmost Taiwan. Acta Oceanogr. Taiwanica, 12: 94 -111. Hsieh, C. H. and Chiu, T. S. (1998). Copepod abundance and species
composition in Tanshui River estuary and adjacent waters. Acta Zoologica Taiwanica, 9: 1-9.
Jan, S., Chern, C. S. and Wang, J. (1994). Influences of sea surface wind stress on summertime flow pattern in the Taiwan Strait. Acta Oceanographica Taiwanica, 33: 63-80. (In Chinese)
Jan, S., Chern, C. S. and Wang, J. (1995). A numerical study on currents in Taiwan Strait during summer time. La mer, 33: 23-40.
Jan, S., Chern, C. S. and Wang, J. (1998). A numerical study of currents in the Taiwan Strait during winter. Terrestrial Atmosphere Oceanic
Science, 9: 615-642.
Nybakken, J. W. (1997). Marine Biology: An Ecological Approach. Adision-Wesley Education Publishers Inc., 41-43.
Pechenik, J. A. (1991). Biology of the Invertebrates. Wm. C. Browm Publishers., 375-381.
Shaw, P. T. (1989) The intrusion of water masses into the sea southern of Taiwan. J. Geophys. Res., 94(C12): 18213-18226.
Shih, C. T. and Chiu, T. S. (1998) Copepod diversity in the water masses of the southern East China Sea north of Taiwan. J. mar. Systems, 15: 533-542.
Shih, C. T. and Young, S. S. (1995) A checklist of free-living copepods, including those associated with invertebrates, reported from the adjacent seas of Taiwan. Acta Zool. Taiwanica, 6: 65-81.
Stephens, J. A., Jorden, M. B., Taylor, A. H. and Proctor, R. (1998). The effects of fluctuations in North Sea flows on zooplankton abundance. J. Plankton Res., 20: 943-956.
Su, C. H. (1996). Vertical migration of five dominant copepods in the northern South China Sea. M.Sc. Thesis, National Taiwan Ocean University.
Tan, T. H. (1967). Distribution of biomass and abundance of zooplankton in water surrounding Taiwan. Acta Oceanogr. Taiwanica, 1: 127-136. Tseng, W. Y., (1972). On Copepoda of the family Candaciidae in the
northeast sea-waters of Taiwan. In: Sugawara, K. (ed.). The Kuroshio II. Proceedings of the Second Symposium on the Results of the Cooperative Study of the Kuroshio and Adjacent Regions. Tokyo. September 28-October 1, 1970. Saikon Publishing Comp. Ltd. Tokyo, pp. 245-259.
Yamaji, I. (1991). Illustrations of the marine plankton of Japan. Hoikusha, Pub. Co. Ltd., 538pp.
seawater from the Taiwan Strait. M. Sc. Thesis, National Sun Yat-Sen University, 100pp. (In Chinese)
中央氣象局 (2000)。氣候資料年報。交通部中央氣象局。 王胄及陳慶生 (1987)。南海北部暖心渦流。台灣海洋學刊。18:92-113。 朱長壽、萬傳龍、陳栩、蘇鳴及林元燒 (1993)。福建東山灣浮游動物的種類組 成和分佈特點。熱帶海洋。12(3):10-15。 吳德泰 (1996)。高屏峽谷水文特性之調查及研究。國立中山大學海洋資源研究 所碩士論文。79 頁。 洪佳章 (2000)。高屏海域生地化作用與通量研究:碳、營養鹽及微量元素(I)。 2000 年國科會永續會永續發展研討會論文集摘要。 郭智聖 (1994)。台灣海峽潮汐數值模式。國立台灣海洋大學海洋科學研究所碩 士論文,64 頁。 張育嘉 (2000)。利用船碇式 ADCP 觀測高屏峽谷之流場特性。國立中山大學海洋 資源研究所碩士論文。96 頁。 台南水工試驗所 (1993)。高雄海域海氣象調查研究: 國立成功大學台南水工試 驗所,研究試驗報告第 140 號。 詹森 (1995)。台灣海峽流場季節變化之研究。國立台灣大學海洋研究所博士論 文。2-6。 劉祖乾 (2000)。高屏海底峽谷大鵬灣交互影響之近岸作用及懸浮沈積物傳輸研 究。2000 年國科會永續會永續發展研討會論文集摘要。 藍揚麒、李明安及石長泰 (2002)。淡水河鄰近海域橈腳類之種類多樣性與其分 布之研究。2002 年「台灣鄰近海域海洋科學研討會」。 羅文增 (2000)。高屏海域陸海交互作用(I)-子計畫二:屏東大鵬灣浮游橈足 類群聚之攝食及對物質傳輸重要性之研究。2000 年國科會永續會永續發展 研討會論文集摘要。 羅文增 (2002)。高屏海域陸海交互作用(II)-高屏附近海域浮游橈足類群聚 之分布及攝食之研究。行政院國家科學委員會補助專題研究計畫成果報告。
Table 1. 2003 年 2 月及 8 月於高屏沿岸海域、東港溪及高屏溪橈足類前 30 個優勢種
類平均豐度(ind./m3)及百分比(RA, %)(*代表淡水種)
Species
Kaoping coastal
waters Tungkang River Kaoping River Total abundance
Mean ± SD RA Mean ± SD RA Mean ± SD RA Mean ± SD RA Napulii* 0 0 5781 ± 7390 24.6 8 ± 12 0.04 2104 ± 5128 12.86 Pseudodiatomus marinus 0 0 1437 ± 4002 6.11 4948 ± 11034 25.42 1872 ± 6200 14.19 Napulii 62 ± 98 3.02 2899 ± 8198 12.33 948 ± 2071 4.87 1335 ± 5002 8.68 Pseudodiatomus annandalei 1 ± 2 0.04 494 ± 1396 2.1 3712 ± 9094 19.07 1192 ± 4783 9.35 Schmackeria dubia* 0 0 2528 ± 4146 10.76 0 0 919 ± 2698 5.62 Acartia omorii 148 ± 415 7.28 0 0 3118 ± 4910 16.01 904 ± 2780 7.26 Neutrodiaptomus schmackeri * 0 0 2135 ± 4680 9.08 0 0 776 ± 2899 4.74 Paracalanus parvus 32 ± 75 1.55 1003 ± 2838 4.27 0 0 376 ± 1709 2.3 Cosmocalanus darwini 0 0 0 0 1224 ± 1399 6.28 334 ± 881 2.72 Acartia spp. 70 ± 167 3.34 116 ± 327 0.49 964 ± 2361 4.95 330 ± 1237 2.55 Acartia spp.* 0 0 886 ± 2507 3.77 0 0 322 ± 1512 1.97 Temora turbinata 187 ± 322 9.16 414 ± 1171 1.76 333 ± 584 1.71 309 ± 763 2.07 Pseudodiatomus spp. 0 ± 1 0.01 669 ± 1892 2.85 0 0 243 ± 1141 1.49 Mongolodiaptomus birulai * 0 0 588 ± 1411 2.50 60 ± 82 0.31 230 ± 862 1.44 Pseudodiaptomidae gen. spp. * 0 0 558 ± 1577 2.37 0 0 203 ± 951 1.24 Paracalanus spp. 25 ± 50 1.21 430 ± 1216 1.83 63 ± 155 0.32 183 ± 732 1.15 Copepodite 65 ± 98 3.18 111 ± 315 0.47 302 ± 433 1.55 147 ± 301 1.06 Acrocalanus longicornis 0 0 0 0 505 ± 1112 2.59 138 ± 589 1.12 Oncaea mediterranea 19 ± 40 0.93 351 ± 991 1.49 1 ± 2 0 135 ± 597 0.82 Acartia sinjiensis 36 ± 96 1.75 271 ± 766 1.15 32 ± 79 0.17 120 ± 462 0.75 Corycaeus crassussulus 80 ± 220 3.90 6 ± 16 0.02 318 ± 506 1.63 118 ± 306 0.89 Calanidae gen. spp. 3 ± 8 0.17 0 0 381 ± 591 1.96 105 ± 336 0.86 Euterpina acutifrons 56 ± 84 2.72 0 0 312 ± 493 1.6 105 ± 279 0.82 Acrocalanus gibber 22 ± 43 1.07 0 0 347 ± 734 1.78 103 ± 391 0.82 Acrocalanus spp. 63 ± 14 0.31 0 0 367 ± 612 1.88 102 ± 342 0.83 Tropocyclops prasinus* 0 0 278 ± 401 1.18 0 0 101 ± 269 0.62 Centropages orsini 0 0 271 ± 766 1.15 0 0 98 ± 462 0.6 Candacia spp. 45 ± 119 2.21 223 ± 631 0.95 0 0 98 ± 383 0.6 Oithona robusta 9 ± 17 0.45 0 0 278 ± 592 1.43 79 ± 315 0.64 Halicyclops aequoreus 0 0 212 ± 599 0.9 0 0 77 ± 361 0.47 Other copepods (74 spp.) 1175 ± 1659 57.61 1843 ± 1125 7.84 1248 ± 1022 6.41 1438 ± 1299 9.48 Total copepods 865 ± 1472 100 21659 ±26535 100 18222 ± 30827 100 13160 ± 23540 100 Species number 64 51 51 104 Table 2. 2003 年 2 月及 8 月大鵬灣橈足類前 30 個優勢種類平均豐度(ind./ m3)及 百分比(RA, %)
Table 3. 2003 年 2 月及 8 月高屏溪、東港溪及大鵬灣浮游性橈足類總豐度及前 6 優勢種與三種環境因子(溫度、鹽度、葉綠素 a 濃度)之複迴歸分析(註*者為
Copepod species Mean ± Std. RA
Bestolina amyoensis (Li&Huang, 1984) 17227 ± 35924 36.45
Paracalanus crassirostris (Dahl,1893) 8316 ± 10524 17.72
Oithona oculata (Farran, 1913) 8042 ± 15249 17.01
Acartia sinjiensis (Mori) 4012 ± 5916 8.49
Acartia spp. 3620 ± 10047 7.66
Paracalanus parvus (Claus, 1863) 1419 ± 3516 3.00
Acartia erythraea (Giesbrtcht, 1889) 1149 ± 3210 2.43
Nauplii 808 ± 1390 1.71
Euterpina acutifrons (Dana, 1849) 546 ± 1067 1.16
Oithona atteneuta (Farran, 1913) 416 ± 625 0.88
Temora turbinate (Dana, 1849) 388 ± 543 0.82
Acrocalanus gracilis (Giesbrtcht, 1888) 323 ± 492 0.68
Oithona brevicornis (Giesbrtcht, 1891) 236 ± 447 0.50
Oithona spp. 105 ± 339 0.22
Pseudodiaptomus marinus (Walter, 1986) 101 ± 124 0.21
Acartia tsuensis (Ito, 1956) 101 ± 219 0.21
Paracalcnidae gen. spp. 90 ± 218 0.19
Oithona simplex (Farran, 1913) 61 ± 148 0.13
Oithona rostrata (Farran, 1913) 28 ± 90 0.06
Oncaea venusta (Farran, 1913) 27 ± 42 0.06
Undinula vulgaris (Dana, 1849) 18 ± 60 0.04
Acartia japonica (Mori) 18 ± 62 0.04
Clausocalanus furtatus (Brady, 1883) 18 ± 41 0.04
Oithona rigida (Giesbrtcht, 1888) 15 ± 45 0.03
Pseudodiaptomus trihamatus (Walter, 1986) 14 ± 34 0.03
Oncaea mediterranea (Claus) 13 ± 31 0.03
Canthocalanus pauper (Giesbrtcht, 1888) 10 ± 31 0.02
Corycaeidae gen. spp. 9 ± 31 0.02
Acartia pacifica (Steuer, 1915) 9 ± 31 0.02
Clausocalanus spp. 9 ± 31 0.02
Other copepods(26 spp.) 60 ± 174 0.13
Total copepods 47267 ± 63526
淡水種) * = P<0.05, ** = P<0.01, *** = P<0.001 高屏溪、東港溪
Temperature Salinity Chl.a
Nauplii* -0.822 0.492 -0.284 Pseudodiatomus marinus -2.733 0.866 -0.540 Napulii -1.410 0.712 -0.418 Pseudodiatomus annandalei -1.973 0.544 -0.365 Schmackeria dubia* -4.409 3.036 -0.858 Acartia omorii 3.481 1.455 0.629 Total Copepods -4.860* 2.165 -1.046 大鵬灣
Temperature Salinity Chl.a
Bestolina amyoensis 3.829 -8.520* -2.760 Paracalanus crassirostris 4.580 -4.779 -6.322* Oithona oculata 5.061* -6.048* -3.275 Acartia sinjiensis -0.005 -0.042 0.156 Acartia spp. -0.838 0.977 0.808 Paracalanus parvus 1.850 -2.836 -1.754 Total Copepods 4.168 -7.890* -3.126
Figure 1. 2003 年於台北淡水河口域之採樣測站圖
Figure 2. 2003 年於高屏溪及東港溪河口域以及大鵬灣之 採樣測站圖
15 20 25 30 35 Tem p er at ur ( ) 慢 C1 C2 C3 C4 C5 C6 T1 T2 T3 T4 T5 F M A N 10 15 20 25 30 35 40 S a lin ity (? C1 C2 C3 C4 C5 C6 T1 T2 T3 T4 T5 Station Figure 3. 2003 年四季於淡水河口域各測站海水表層之溫度及鹽度分布圖。 F: February; M: May; A: August; N: November.
22 24 26 28 30 32 T e m p eratur ( ) 慢 C1 C2 C3 C4 K1 K2 K3 T1 T2 T3 T4 T5 Station F M A N 0 10 20 30 40 S a linity (? C1 C2 C3 C4 K1 K2 K3 T1 T2 T3 T4 T5 Station F M A N Figure 4. 2003 年四季於高屏溪及東港溪河口域各測站表層海水之溫度及鹽度 分布圖。F: February; M: May; A: August; N: November.
20 25 30 35 Tem p erat ur ( ) 慢
Feb. Apr. Jun. Aug. Oct. Dec.
25 30 35 S alin ity ( ?
Feb. Apr. Jun. Aug. Oct. Dec.
Month St. 6 St. 5 St. 4 St. 3 St. 2 St. 1 Figure 5. 2003 年於大鵬灣各測站海水表層溫度及鹽度之季節分布。
0 50 100 150 200 Abundance ( ind./ 100 m 3) C1 C2 C3 C4 K1 K2 K3 T1 T2 T3 T4 T5 Station Paracalanus parvus 0 200 400 600 Abundance ( ind./ 100 m 3) C1 C2 C3 C4 K1 K2 K3 T1 T2 T3 T4 T5 Pseudodiatomus marinus 2744 0 200 400 600 C1 C2 C3 C4 K1 K2 K3 T1 T2 T3 T4 T5 Napulii Aug. Feb. 1133 2319 0 200 400 600 Abundance ( ind./ 100 m 3) C1 C2 C3 C4 K1 K2 K3 T1 T2 T3 T4 T5 Pseudodiatomus annandalei 0 200 400 600 C1 C2 C3 C4 K1 K2 K3 T1 T2 T3 T4 T5 Acartia omorii 1268 2227 803 0 100 200 300 C1 C2 C3 C4 K1 K2 K3 T1 T2 T3 T4 T5 Station Cosmocalanus darwini Figure 6. 2003 年 2 月及 8 月於高屏溪及東港溪河口域各測站前 6 個優勢 海水種橈足類之豐度分布圖
0 250 500 750 1000 Abundance (ind./100 m 3) C1 C2 C3 C4 K1 K2 K3 T1 T2 T3 T4 T5 Napulii 0 100 200 300 400 500 C1 C2 C3 C4 K1 K2 K3 T1 T2 T3 T4 T5 Schmackeria dubia Aug. Feb. 0 100 200 300 400 500 Abundance (ind./100 m 3) C1 C2 C3 C4 K1 K2 K3 T1 T2 T3 T4 T5 Neutrodiaptomus schmackeri 0 100 200 300 400 500 C1 C2 C3 C4 K1 K2 K3 T1 T2 T3 T4 T5 Acartia spp. 0 10 20 30 40 50 Abundance (ind./100 m 3) C1 C2 C3 C4 K1 K2 K3 T1 T2 T3 T4 T5 Station Mongolodiaptomus birulai 0 10 20 30 40 50 C1 C2 C3 C4 K1 K2 K3 T1 T2 T3 T4 T5 Station Pseudodiaptomidae gen. spp. 2040 1225 1323 709 406 446 1125 Figure 7. 2003 年 2 月及 8 月於高屏溪及東港溪河口域各測站前 6 個優勢淡水 種橈足類之豐度分布圖
0 300 600 900 Abundance (x10 3 ind./100 m 3) C1 C2 C3 C4 K1 K2 K3 T1 T2 T3 T4 T5 February 0 5 10 15 20 C1 C2 C3 C4 K1 K2 K3 T1 T2 T3 T4 T5 August FW spp. SW spp. 0 5 10 15 20 25 S p eci es num ber C1 C2 C3 C4 K1 K2 K3 T1 T2 T3 T4 T5 0 5 10 15 20 25 C1 C2 C3 C4 K1 K2 K3 T1 T2 T3 T4 T5 0 1 2 3 4 5 .H' C1 C2 C3 C4 K1 K2 K3 T1 T2 T3 T4 T5 Station 0 1 2 3 4 5 C1 C2 C3 C4 K1 K2 K3 T1 T2 T3 T4 T5 Station nd nd nd Figure 8. 2003 年 2 月及 8 月於高屏溪及東港溪河口域各測站橈足類豐度、種 類數及種歧異度分布圖
0 500 1000 1500 1 2 3 4 5 6 Bestolina amyoensis Aug. Feb. 0 100 200 300 400 1 2 3 4 5 6 Paracalanus crassirostris 0 100 200 300 400 500 Abundance (x10 4 ind./100 m 3) 1 2 3 4 5 6 Oithona oculata 0 50 100 150 200 1 2 3 4 5 6 Acartia sinjiensis 0 100 200 300 400 1 2 3 4 5 6 Staion Acartia spp. 0 50 100 150 1 2 3 4 5 6 Staion Paracalanus parvus Figure 9. 2003 年 2 月及 8 月於大鵬灣各測站前 6 優勢種橈足類之豐度分布圖
0 50 100 150 200 250 A b undance (x10 5 ind./100 m 3 ) 1 2 3 4 5 6 Aug. Feb. 0 10 20 30 40 Species number 1 2 3 4 5 6 0.0 0.5 1.0 1.5 2.0 2.5 3.0 H' 1 2 3 4 5 6 Staion Figure 10. 2003 年 2 月及 8 月於大鵬灣各測站之橈足類豐度、種類數 及種歧異度分布