行政院國家科學委員會專題研究計畫 成果報告
Azotobacter vinelandii 對 tetracyanonicklate (TCN)
分解能力之探討
計畫類別: 個別型計畫 計畫編號: NSC91-2314-B-039-035- 執行期間: 91 年 08 月 01 日至 92 年 07 月 31 日 執行單位: 中國醫藥大學微生物學科 計畫主持人: 陳師慶 報告類型: 精簡報告 處理方式: 本計畫可公開查詢中 華 民 國 92 年 10 月 31 日
1
摘要
Azotobacter vinelandii ATCC 13705(A. vinelandii)是一株革蘭氏陰性好氧固 氮菌,在本研究中,A. vinelandii 被培養在 1、10 及 20 mM 三種不同濃度之四氰 化鎳(Tetracyanonickelate,簡稱 TCN)溶液中,以評估 TCN 對此菌的影響情形。 研究結果顯示菌株含 1 mM TCN 在對數生長期(19 小時)時,其 TCN 降解率最 大,比生長率最高,固氮酵素活性較高,氧呼吸率亦偏高,此現象與 nitrogenase 的呼吸保護作用符合。另外,本研究亦發現當含有其他氮源時,將造成菌株對 TCN 降解的抑制。如在 1 mM TCN 中添加 1、5 及 10 mM 銨離子時,TCN 降解 率為 28﹪(24 小時);添加 5 及 10 mM 亞硝酸根離子時,TCN 降解率為 16﹪(24 小時)。此外,在靜止細胞(resting cells)中降解 1 mM TCN 時,其降解率為 6 ﹪(16 小時);當加入 8﹪葡萄糖時則可加速 TCN 被降解的速率(8 小時為 43 ﹪)。此研究成果使我們對 A. vinelandii 分解四氰化鎳之機制有更深入的瞭解, 以期日後能夠將實驗參數條件應用於含氰化物廢水的實廠處理。 關鍵詞: Azotobacter vinelandii; TCN; 生物分解 英文摘要
In this study, Azotobacter vinelandii ATCC13705 (A. vinelandii), which is a free-living, nitrogen-fixing, gram-negative, and aerobic rod bacterium, was need to evaluate its ability to biodegrade tetracyanonickelate (TCN) under different conditions. Results show that A. vinelandii was able to biodegrade various concentrations of TCN (1, 10, and 20 mM) under aerobic conditions. Higher degradation rate of TCN, higher nitrogenase activity, higher oxygen consumption, and higher specific growth rates were also observed at log growth period. Results suggest that the hypothesis of respiratory protection of nitrogenase is supported. Moreover, the addition of ammonia (1, 5, and 10 mM) would cause the decrease of TCN degradation rate (28%) during a 24-hr incubation period. Inhibition of TCN degradation (degradation rate:16% for 24 hrs) was observed when nitrite (5 and 10 mM) was added into the growth medium. Furthermore, the addition of 8% of glucose would significantly enhance the TCN degradation by the resting cells (degradation rate:43% for 8 hrs) . Results from this study provide us insight into the characteristics and mechanisms of TCN conversion by A. vinelandii.
關鍵詞: Azotobacter vinelandii; TCN; biodegradation
Introduction
很多金屬表面處理廠通常為了增加金屬表面的光澤與美觀,常常會在電鍍製
水中常會含有或高或低的氰化物。而廢水中所含氰化物可分兩大類:複合氰化物 和單體氰化物。複合氰化物經水解或光解,可分解為 CN-和 HCN 兩種單體氰化 物型態。在酸性狀態下,對形成 HCN 較有利;反之若在鹼性狀態下,對形成 CN -較有利。兩種單體氰化物中,就毒性而言,HCN 遠高於 CN-,因此,廢水中含 有氰化物時,則需將廢水 pH 維持在鹼性狀態,切忌與酸混合,以防產生具有強 烈毒性的 HCN。 通常氰化物為含有劇毒性之高活性化學物,會抑制以金屬離子為活性中心的 酵素活性,如 hemoprotein 等,特別是位於有氧呼吸電子傳遞鏈上的胞色素氧化 酵素(Knowles, 1976),當氰化物進入人體後,會使此酵素失活,因而導致窒息 而死。對人體而言,致死量為 200~300 mg (Michael and Roberts, 1982),因此 我國現行放流水標準中,氰化物最大限值為 1.0 mg/L,美國同樣也是 1.0 mg/L,
但美國所管制的只有「可被氯氧化的氰化物」,而我國所管制的是「總氰化物」。
氰化物能抑制細胞內的許多酵素活性,而其在自然界中的來源除了人為工 業污染外,許多植物及微生物也會分泌氰化物到生態環境中(Knowles, 1976), 因此往往可由植物根部土壤篩選出對氰化物具有耐受力並可利用其當作氮源的 微生物(Mizushima and Arima, 1960;Arima and Oka, 1965;Skowronski and Strobel, 1969;Knowles, 1976;Harris and Knowles, 1983a;White et al., 1988;Silva-Avalos et al., 1990)。而具有氰化物合成能力的植物,包含樹薯(Cassava)、亞麻(Flax)、 高梁(Sorghum)、紫花苜蓿(Alfalfa)、桃樹(Peaches)、杏仁(Almonds)及多 種豆科植物(Beans)等農作類植物。這些植物通常由根部分泌氰化物到土壤中, 其合成氰化物途徑為先將氨基酸(Glycine)合成 Cyanogenic glycoside,之後再 將 Cyanogenic glycoside 分解產生氰化物。
除了上述植物之外,尚有許多原核生物(如 Chromobacterium violaceum、 Pseudomonas fluorescens、Pseudomonas aeruginosa 等)也會合成、分泌氰化物, 因此氰化物存在於自然界是一種普遍的現象。但其合成途徑與植物不同,是經由 氨基酸直接反應,反應式如下(Knowles, 1976;Knowles and Brunch, 1986):
自然界中存在許多具有可分解氰化物能力的微生物,而此能力是利用菌種 本身所擁有的酵素。不同種類的微生物會有不同類型的酵素,而各酵素對於氰化 物分解反應機制亦不相同。
本實驗探討A. vinelandii是否利用固氮酵素來降解 TCN,同時並檢測是否
還有其他的氰化物分解酵素。
3. Materials and Methods
實驗菌種
3
Azotobacter vinelandii ATCC 13705 購自食品工業發展研究所生物資源保存 及研究中心。
培養基
液體培養基為不含氮源之 Burk 培養基(nitrogen-free Burk’s medium),其成 分如下:CaCl2,0.64 mM;K2HPO4,3.7 mM;MgSO4,0.81 mM;Na2MoO4,
0.004 mM;KH2PO4,1.2 mM;FeSO4,0.01 mM;NaCl,1.3 mM,並再以 0.1 N
HCl 調其 pH 為 7.0 (Wong, 1993)。Glucose 經高溫高壓(120℃,AS-1080L 型) 滅菌後再加入前述之培養基中,其最終濃度為 0.8%。
四氰化鎳(TCN)濃度標準曲線之製備
TCN 濃度之測定係採用紫外光光譜分析法,已知 TCN 在波長 267 nm 處有 最高吸收光譜,因此利用分光光度計(HITACHI,U 2000 型)對已知一系列 TCN 濃度之溶液於波長 267 nm 附近進行波長掃描,即可得一與各 TCN 濃度相對之吸 光值標準曲線(Rollinson et al., 1987;Silva-Avalos et al., 1990)。
銨離子濃度之測定
利用分析銨離子濃度的變化,以檢測氰化物分解產物中是否有銨離子的存 在。在 eppendrof 中加入 400μL 樣品及 200μL 的 Sodium phenate (取 0.312 M 之 NaOH 溶液 100 mL ,加入 2.5 g 之 phenol 配製而成)、200μL Sodium nitroprusside (0.01%)、200μL Sodium hypochlorite(6%),加蓋混合均勻,置於 室溫反應 30 分鐘,再以分光光度計(HITACHI,U 2000 型)測量波長 630 nm 之吸光值(Fawcett and Scott, 1960;Rollinson et al., 1987)。
甲醯胺(Formamide)濃度之測定
利用分析甲醯胺濃度的變化,以檢測氰化物分解產物中是否有甲醯胺的存 在。在 eppendrof 中分別加入 0.2 mL 樣品、0.2 mL 3.5 N 之 NaOH、0.2 mL 之 2.3 M Hydroxylamine hydrochloride,置於 60℃水浴槽中 10 分鐘,再於室溫下加 入 0.2 mL 之 4 N HCl 及 0.2 mL 之 1.23 M FeCl3,靜置 5 分鐘後,以分光光度計
(HITACHI,U 2000 型)測量波長 540 nm 之吸光值(Kunz et al., 1992)。 甲酸(Fromate)濃度之測定
利用分析甲酸濃度的變化,以檢測氰化物分解產物中是否有甲酸的存在。 在 eppendrof 中分別加入 0.02 mL NAD solution(0.05 Mβ-NAD)、0.16 mL 樣 品、0.80 mL phosphate buffer(50 mM, pH 7.0),以分光光度計(HITACHI,U 2000 型)測量波長 340 nm 之吸光值 E1,之後再加入 0.02 mL formate dehydrogenase
(20 U / mL),混合均勻後靜置 2 ~ 4 分鐘,再以分光光度計(Hitachi U 2000) 測量波長 340 nm 之吸光值(Hopner and Knappe, 1974)。
結果與討論
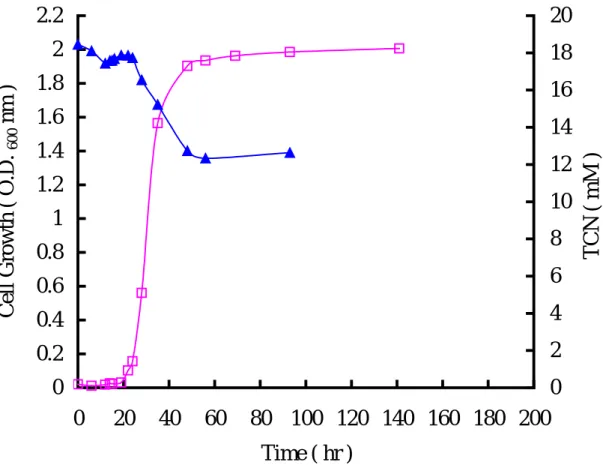
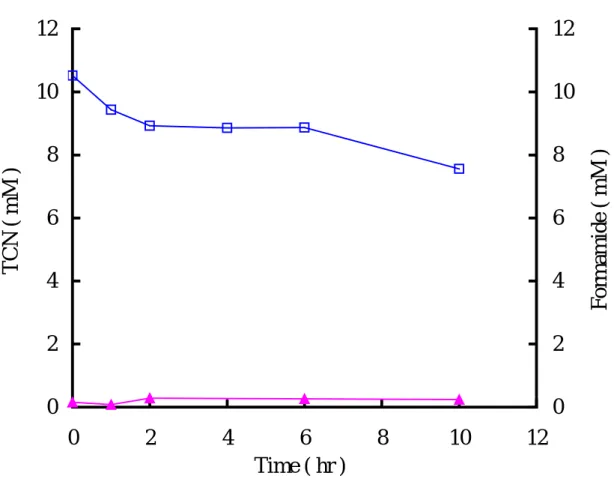
圖 一顯示,當細菌處於遲滯期(0 ~ 19 小時)時,四氰化鎳並無明顯降解 的現象,而隨著菌量的增加,在第 19 小時(對數生長期)可發現四氰化鎳濃度 亦隨之下降,直到 93 小時後將 20 mM 四氰化鎳降解至 12.6 mM。推測此菌可利 用 TCN 作為氮源以供生長•相關文獻(Harris and Knowles, 1983b;Ingvorsen et al., 1991;Kunz et al., 1992;Barclay et al., 1998b;Kwon et al., 2002)指出氰化物具 有相當多的酵素降解途徑,而且不同酵素會產生不同的降解產物及降解途徑。目 前已證實氰化物被降解後,所產生的產物包含甲醯胺、甲酸、氨、二氧化碳和甲 烷等,但並不是每一株菌降解氰化物都會產生以上所述的產物,因為每株菌本身 所具有的酵素皆不同,就會有不同的產物產生。依據 Kunz 等人(1992)的研究, P. fluorescens NCIMB11764 在降解氰化物後可檢測到甲醯胺(formamide)和甲 酸(formate);另外 1998 年 Watanabe 等人的研究中也指出 P. stutzeri Ak61 降解 氰化物後會產生甲酸。同時也已知甲醯胺和甲酸是由 cyanide hydratase 和 cyanidase 降解氰化物所產生的,因此若可檢測到甲醯胺和甲酸,則 cyanide hydratase 和 cyanidase 就極有可能為其氰化物降解酵素。而甲烷和氨則是固氮酵 素降解氰化物之產物(Li et al., 1982;Kao et al., 2003)。故本實驗測定四氰化鎳 被菌種降解後之產物,以間接推測何種酵素可能參與降解作用。Figure 2 為靜止 細胞(resting cells)分解 10 mM 四氰化鎳過程中,檢測是否有甲醯胺的生成。 在 10 小時內四氰化鎳由原濃度 10.5 mM 降解至 7.6 mM,而甲醯胺濃度並未隨四
氰化鎳濃度降低而增加(每隔 2 小時檢測一次 TCN 及甲醯胺濃度)。雖然甲醯胺
在 TCN 降解過程中,無法測得其存在,但亦不能排除其為 TCN 降解後之可能性 產物。另外甲酸(formate)為 cyanidase 降解 TCN 之產物(Watanabe et al., 1998; Barclay et al., 1998b;Kwon et al., 2002),本實驗亦未測得甲酸之產生(data not
shown)。此外,發現此菌株並不會利用甲酸(data not shown)。因此可推測 A.
vinelandii 極有可能不具有 cyanidase,因為當四氰化鎳被降解時,並未測得甲酸 的存在。由以上結果可排除 cyanidase 參與 TCN 降解之可能性。
銨離子為四氰化鎳被降解後之最終產物(Yanase et al., 2000;Kwon et al., 2002;Kao et al., 2003),Figure 3 顯示四氰化鎳在靜止細胞(resting cells)情況 下,其降解能力較為緩慢,但仍可測得銨離子的產生。在第 14 小時銨離子產量 最高,為 0.1 mM,之後銨離子生成曲線便呈現遞減趨勢,直到第 24 小時便檢測 不到銨離子濃度;而四氰化鎳則由原濃度 1.0 mM 降解為 0.8 mM。由此可推知四 氰化鎳的降解會導致銨離子的生成,且在其它研究中亦有提及氰化物可被降解成 銨離子以供細胞利用(White et al., 1988;Fallon et al., 1991;Kunz et al., 1992;
Watanabe et al., 1998)。但在生長情況下卻無法測得銨離子的產量,此現象可能
是因為銨離子迅速被利用掉,在 Kunz 等人(1992)之研究中 P. fluorescens NCIMB 11764 亦有相同情形,當氰化物濃度較低時(0.25 ~ 1 mM)並無法測得銨離子的 濃度。
5
當培養基中缺乏葡萄糖時,其 TCN 降解情形十分緩慢,但加入葡萄糖卻可 促進 resting cells 對四氰化鎳快速的降解,Figure 4 為降解過程中加入葡萄糖後, 四氰化鎳快速被降解之情形。結果顯示,在 16 小時前未加入葡萄糖時,四氰化 鎳降解速度較緩慢,由原濃度 1.0 mM 降到 0.9 mM,但在第 16 小時加入葡萄糖 後,四氰化鎳便在 8 小時內快速降解至 0.5 mM。同時也檢測不到銨離子濃度。 推測葡萄糖在此過程中是提供菌種碳源以利能分解 TCN,消耗能量所需。Kwon 等人(2002)的研究中就指出 Cryptococcus humicolus MCN2 有相同的現象,假 如提供足夠的碳源(例如,葡萄糖)便能夠快速降解高濃度的金屬氰化物。
Fig. 1. The growth curve of A. vinelandii and TCN degradation in Burk’s
medium contained 20 mM TCN.(
□)cells growth curve;(▲)consumption of 20 mM TCN.
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
2
2.2
0
20
40
60
80
100 120 140 160 180 200
Time ( hr )
Cel
l Growt
h
( O.D.
600nm )
0
2
4
6
8
10
12
14
16
18
20
TCN ( mM )
0
2
4
6
8
10
12
0
2
4
6
8
10
12
Time ( hr )
TCN ( mM )
0
2
4
6
8
10
12
Formami
d
e ( mM )
Fig. 2. The degradation of TCN and the formation of formamide by
A.-
vinelandii resting cells(O.D.
600nm=1.00, cell mass=0.5 mg /
7
0
0.2
0.4
0.6
0.8
1
1.2
0
10
20
30
40
50
60
70
80
Time ( hr )
TCN ( mM )
0
0.2
0.4
0.6
0.8
1
1.2
Ammo
n
ia ( mM )
Fig. 3. The degradation of 1mM TCN and production of ammonia by
A.-
vinelandii resting cells in Burk’s medium.(
◆)1 mM TCN;(
○)
amounts of ammonia.
0
0.2
0.4
0.6
0.8
1
1.2
0
20
40
60
80
Time ( hr )
TCN ( mM )
0
0.2
0.4
0.6
0.8
1
1.2
Ammonia ( mM )
0 .8 ﹪gluco seFig. 4. The degradation of 1mM TCN and production of ammonia by
A.-
vinelandii resting cells in Burk’s medium with the addition of 0.8%
glucose into 16 hours cultures.(◆)1 mM TCN;
(○)amounts of
ammonia.
Reference
Arima, K., and Oka, T.(1965)Cyanide resistance in Achromobacter. I. Induced
formation of cytochrome a and role in cyanide-resistant respiration. Jounral of Bacteriology 90:734-743.
Burris, R. H. and Wilson, P. W.(1946)Ammonia as an intermediate in nitrogen
fixation by Azotobacter. Journal of Bacteriology 9:505-512.
Chena, S. C., and Liu, J. K.(1999)The respiratory responses to cyanide of a
cyanide-resistant Klebsiella oxytoca bacterial strain. FEMS Microbiology Letters 175:37-43.
Fawcett, J. K., and Scott, J. E.(1960)A rapid and precise method for the
determination of urea. Journal of Clinical Pathology 13:156-160.
Haddock, B. A., and Jones, C. W.(1977)Bacterial respiration. Bacteriological Reviews 41:47-99.
Harris, R. E., and Knowles, C. J.(1983a)The conversion of cyanide to ammonia by
extracts of a strain of Pseudomonas fluorescens that utilizes cyanide as a source of nitrogen of growth. FEMS Microbiology Letters 20:337-341.
Hopner, Y., and Knappe, J. ( 1974 ) Formate : determination with formate
dehydrogenase. Methods of Enzymatic Analysis 3:1551-1555.
Jones, R., Woodley, P., and Robson, R.(1984)Cloning and organization of some
genes for nitrogen fixation from Azotobacter chroococcum and their expression in Klebsiella pneumoniae. Molecular & General Genetics 197:318-327.
9
Klugkist, J., and Haaker, H.(1984)Inhibition of nitrogenase activity by ammonium
chloride in Azotobacter vinelandii. Journal of Bacteriology 157:148-151.
Knowles, C. J.(1976)Microorganisms and cyanide. Bacteriological Reviews 40:
652-680.
Knowles, C. J., and Bunch, A. W.(1986)Microbial cyanide metabolism. Advances in Microbial Physiology 27:73-111.
Knowles, C. J.(1988)Cyanide utilization and degradation by microorganisms. CIBA Foundation Symposium 140:3-15.
KoKormondy, E. J.(1996)Concepts of Ecology. Fourth edition:128-140.
Krol, A. J. M., Hontelez, J. G. J., Roozendall, B., and Kammen, A. van.(1982)On the operon structure of the nitrogenase genes of Rhizobium leguminosarum and Azotobacter vinelandii. Nucleic Acids Research 10:4147-4157.
Kunz, D. A., Nagappan, O., Silva-avalos, J., and Delong, G. T.(1992)Utilization of
cyanide as a nitrogenous substrate by Pseudomonas fluorescens NCIMB 11764: evidence for multiple pathways of metabolic conversion. Applied and Environmental Microbiology 58:2022-2029.
Kunz, D. A., Wang, C. S., and Chen, J. L.(1994)Alternative routes of enzymic
cyanide metabolism in Pseudomonas fluorescens NCIMB 11764. Microbiology 140: 1705-1712.
Liu, J. K., Liu, C. H., and Lin, C. S.(1997)The role of nitrogenase in a
cyanide-degrading Klebsiella oxytoca strain. Proceedings of the National Science Council B, ROC 21:37-42.
Lucinski, R., Polcyn, W., and Ratajczak, L.(2002)Nitrare reduction and nitrogen
fixation in symbiotic association Rhizobium legumes. Acta Biochimica Polonica 49: 537-546.
Manhart, J. R., and Wong, P. P.(1979)Nitrate reductase activities of Rhizobia and
the correlation between nitrate reduction and nitrogen fixation. Canadian Journal of Microbiology 25:1169-1174.
Michael, I. G., and Roberts, J. R. ( 1982 ) Cyanide poisonings. Emergency Medicine:112-113.
Mizushima, S., and Arima, K. ( 1960 ) Mechanism of cyanide resistance in Achromobacter. I. Adaptive formation of cyanide resistant respiratory system in growing cells. The Journal of Biochemistry 47:351-360.
Nester, E. W., Roberts, C. E., and Nester, M. T.(1995)Metabolism:The generation
of energy and synthesis of small molecules. Microbiology. Wm. C. Brown Communications, Inc.
Perry, J. J., and Staley, J. T.(1997)Microbiology:Dynamics & Diversity:442-444.
Schmitz, R. A., Klopprogge, K., and Grabbe, R.(2002)Regulation of nitrogen
fixation in Klebsiella pneumoniae and Azotobacter vinelandii:NifL, transducing two environmental signals to the nif transcriptional activator NifA. Journal of Molecular Microbiology and Biotechnology 4:235-242.
Silva-Avalos, J., Richmond, M. G., Nagappan, O., and Kunz, D. A.(1990)
Degradation of the metal-cyano complex tetracyanonicklate(Ⅱ)by cyanide-utilizing bacterial isolates. Applied and Environmental Microbiology 56:3364-3670.
Skowronski, B., and Strobel, G. A.(1969)Cyanide trsistance and cyanide utilization
by a strain of Bacillus pumilus. Canadian Journal of Microbiology 15:93-98.
Vaughn, S. A., and Burgess, B. K.(1989)Nitrite, a new substrate for nitrogenase. Biochemistry 28:419-424.
Watanabe, A., Yano, K., Ikebukuro, K., and Karube, I.(1998)Cyanide hydrolysis
in a cyanide-degrading bacterium, Pseudomonas stutzeri AK61, by cyanidase. Microbiology 144:1677-1682.
Way, J. L.(1983)Mechanism of cyanide intoxication and its antagonism:introduction.
Fundamental and Applied Toxicology : Official Journal of the Society of Toxicology 3:369.
White, J. M., Jones, D. D., Huang, D., and Gauthier, J. J. (1988) Conversion of
cyanide to formate and ammonia by a pseudomonad obtained from industrial wastewater. Journal of Industrial Microbiology 3:263-272.
11
Wong, T. Y.(1993)Effects of calcium on sugar transport in Azotobacter vinelandii. Applied and Environmental Microbiology 59:89-92.
Yates, M. G.(1970)Control of respiration and nitrogen fixation by oxygen and
adenine nucleotide in N2-grown Azotibacter chroococeca. Journal of General