BRAIN NOCICEPTIVE IMAGING IN RATS USING
18F-FLUORODEOXYGLUCOSE SMALL-ANIMAL POSITRON
EMISSION TOMOGRAPHY
Y.-Y. I. SHIH,a Y.-C. CHIANG,a,b1 J.-C. CHEN,c,d C.-H. HUANG,a,b Y.-Y. CHEN,e R.-S. LIU,f C. CHANGa* AND F.-S. JAWb*
aInstitute of Biomedical Sciences, Academia Sinica, Taipei, Taiwan, Republic of China
bInstitute of Biomedical Engineering, National Taiwan University, Tai-pei, Taiwan, Republic of China
c
Department of Biomedical Imaging and Radiological Sciences, Na-tional Yang-Ming University, Taipei, Taiwan, Republic of China d
Department of Education and Research, Taipei City Hospital, Taipei, Taiwan, Republic of China
e
Department of Electrical and Control Engineering, National Chiao-Tung University, Hsinchu, Taiwan, Republic of China
f
School of Medicine, National Yang-Ming University, Taipei, Taiwan, Republic of China
Abstract—Preclinical exploration of pain processing in the brain as well as evaluating pain-relief drugs in small ani-mals embodies the potential biophysical effects in hu-mans. However, it is difficult to measure nociception-re-lated cerebral metabolic changes in vivo, especially in unanesthetized animals. The present study used 18 F-flu-orodeoxyglucose small-animal positron emission tomog-raphy to produce cerebral metabolic maps associated with formalin-induced nociception. Anesthesia was not applied during the uptake period so as to reduce possible con-founding effects on pain processing in the brain. The for-malin stimulation at the hind paw of rats resulted in signif-icant metabolic increases in the bilateral cingulate cortex, motor cortex, primary somatosensory cortex, secondary somatosensory cortex, insular cortex, visual cortex, cau-date putamen, hippocampus, periaqueductal gray, amyg-dala, thalamus, and hypothalamus. Among the measured areas, clear lateralization was only evident in the primary somatosensory cortex and hypothalamus. In addition, pre-treatment with lidocaine (4 mg/kg, i.v.) and morphine (10 mg/kg, i.v.) significantly suppressed formalin-induced cerebral metabolic increases in these areas. The present protocol allowed identification of the brain areas involved in pain processing, and should be useful in further
evalu-ations of the effects of new drugs and preclinical therapies for pain. © 2008 IBRO. Published by Elsevier Ltd. All rights reserved.
Key words: PET, pain, morphine, lidocaine, rat.
Novel pain-relief drugs and therapeutic techniques have been developed in recent years to alleviate human suffer-ing from pain. However, the mechanisms and circuits un-derlying pain processing are extremely complex, involving not only sensory responses to noxious stimuli, but also cognitive and emotional factors (McMahon and Koltzen-burg, 2005). This makes it difficult to conclusively identify the brain areas that specifically process nociceptive stim-uli. Imaging approaches such as positron emission tomog-raphy (PET) and functional magnetic resonance imaging (fMRI) allow functional mapping of the intact brain and measurement of the responses in multiple areas simulta-neously, thus bringing the study of pain into a deep level (Phelps et al., 1979; Ogawa et al., 1990a,b; Phelps, 2000). Preclinical verification for evaluating the efficacy of pain-alleviation strategies is usually essential. Of the nu-merous pain-testing models in animals, formalin test is one of the most commonly used techniques for generating nociception since it evokes inflammatory pain responses without influencing other sensory modalities; furthermore, the associated behavioral responses have been well in-vestigated (Tjolsen et al., 1992). Our recent animal fMRI studies have shown that formalin stimulation of the rat hind paw significantly activates the cingulate cortex (Cg), motor cortex (M), primary somatosensory cortex (S1), secondary somatosensory cortex (S2), insular cortex (IC), visual cor-tex (VC), caudate putamen (CPu), hippocampus (HIP), periaqueductal gray (PAG), thalamus (Th), and hypothal-amus (HT) (Shih et al., in press, 2008b). However, it is usually essential to apply anesthesia during fMRI experi-ments in order to sedate the animal and reduce motion artifacts. This tackles a very difficult technical obstacle when imaging the representation of pain in anesthetized animals. Small-animal positron emission tomography (mi-croPET) might be more suitable for imaging brain activa-tion in the conscious animals (Schiffer et al., 2007; Ohashi et al., 2008). An animal can react to stimuli outside the scanner without stresses during the uptake of radionuclide, and then be lightly anesthetized for imaging the accumu-lated responses, thus minimizing possible confounding variables that could influence central nociceptive process-ing.
1
Equal contribution with first author.
*Corresponding author. Tel:⫹886-2-2789-9027; fax: ⫹886-2-2788-7641 (C. Chang); Tel: ⫹886-2-3366-5266; fax: ⫹886-2-2394-0049 (F.-S. Jaw).
E-mail address: [email protected] (C. Chang), jaw@ntu. edu.tw (F.-S. Jaw).
Abbreviations: Amyg, amygdala; Cg, cingulate cortex; CPu, caudate putamen; fMRI, functional magnetic resonance imaging; HIP, hip-pocampus; HT, hypothalamus; IC, insular cortex; M, motor cortex; MAP, maximum a posteriori; microPET, small-animal positron emis-sion tomography; PAG, periaqueductal gray; PET, positron emisemis-sion tomography; S1, primary somatosensory cortex; S2, secondary so-matosensory cortex; Th, thalamus; VC, visual cortex;18
F-FDG,18
F-fluorodeoxyglucose; %ID/g, percentage injected dose per gram. 0306-4522/08 © 2008 IBRO. Published by Elsevier Ltd. All rights reserved. doi:10.1016/j.neuroscience.2008.07.013
The present study aimed to elucidate formalin-induced nociceptive responses by using 18
F-fluorodeoxyglucose (18
F-FDG) microPET with the aid of a maximum a poste-riori (MAP) reconstruction algorithm to further improve the spatial resolution (Qi et al., 1998). In addition, MRI and a rat brain atlas were used to anatomically align the rat brain (Paxinos and Watson, 1998; Shih et al., 2007). The new information provided by the current study is that nocicep-tion-induced metabolic maps can be imaged in the con-scious rat brain, in contrast to previous studies only reveal-ing indirect hemodynamic responses durreveal-ing anesthesia (Morrow et al., 1998; Tuor et al., 2000; Shah et al., 2005; Shih et al., in press, 2008b). The effects of pain-relief drugs such as morphine and lidocaine were also examined. This pain-measuring protocol can be used to evaluate the ef-fectiveness of new pain-relief drugs and therapeutic tech-niques at the preclinical stage.
EXPERIMENTAL PROCEDURES
SubjectsNineteen adult male Wistar rats (8 –10 weeks old; weighing ap-proximately 250 –300 g; National Laboratory Animal Center, Taipai, Taiwan, Republic of China) were used in the present study. The animals were housed in a well-controlled environment with a 12-h light/dark cycle and constant humidity and temperature. Rats were housed in plastic cages at three animals per cage with free access to food and water. All experimental procedures were ap-proved by the Institutional Animal Care and Use Committee, Na-tional Taiwan University, College of Medicine. All experiments conformed to international guidelines on the ethical use of ani-mals. All efforts were made to minimize the number of animals used and their suffering.
Imaging experiments
Seven rats were used to produce the formalin-induced nociceptive maps. The18F-FDG was used as a radiotracer to reveal brain
glucose metabolism. Each rat was initially lightly anesthetized using ether, and 0.5 ml of18F-FDG with an activity of 1.0 –1.2 mCi
was administered i.v. via the tail lateral vein, after which the rat was returned to its cage in a quiet environment for 45 min uptake in the conscious state. Following the uptake, the rat was lightly anesthetized using 1.5% isoflurane and fixed in a custom-built stereotaxic head holder by two ear bars and an incisor fixer so as to minimize motion artifacts (Shih et al., 2007). The body temper-ature was maintained using a warming lamp whose light field was restricted to avoid additional visual stimulation. MicroPET imaging
(R4, Concorde Microsystems/Siemens, Knoxville, TN, USA) was performed for 30 min, with the images reconstructed using the MAP algorithm (Qi et al., 1998). After 1 week, 50l of 5% formalin was injected into the left hind paw prior to18F-FDG injection. The
imaging procedures were identical to those described above. Another two groups containing six rats each were used to evaluate the effects of lidocaine and morphine. The drugs were given prior to the formalin stimulation, followed by an identical imaging protocol. Rats were i.v. injected with 4 mg/kg lidocaine in one group and 10 mg/kg morphine in the other group.
MRI anatomical images were captured using a 4.7-T Biospec 47/40 spectrometer to define the brain margin. A 72-mm volume coil was used as the RF transmitter, and a 2-cm quadrature surface coil placed on the head was used as the receiver. A T2-weighted scout image was taken in the mid-sagittal plane to
localize the anatomical position by identifying the anterior com-missure (bregma⫺0.8 mm). T2-weighted template images were
then acquired using RARE sequence with a repetition time of 4000 ms, echo time of 80 ms, field of view of 2.56 cm, slice thickness of 1.2 mm, number of excitation of 2, and an acquisition matrix of 256⫻128 (zero-filled to 256⫻256).
Data analysis
Images were analyzed using PMOD (PMOD Technologies, Adliswil, Switzerland) and a custom-built ISPMER system (Shih et al., 2007). MicroPET images were initially coregistered among the subjects using a mutual-information algorithm and then averaged to generate the incidence images. A pixel value in incidence images represents the averaged percentage injected dose per gram (%ID/g) of an experimental group, where a higher pixel value indicates a greater number of rat responses consistent with the given task.
The statistical analysis was based on the %ID/g values sam-pled from different brain structures of each rat. Repeated-mea-sures ANOVAs were used to examine whether formalin stimula-tion induced metabolic changes in the corresponding brain re-gions in both hemispheres, with the significance level set at
P⬍0.05. Factorial ANOVAs were used to assess differences in
18F-FDG uptake among the groups with formalin stimulation
alone, formalin stimulation with lidocaine pretreatment, and for-malin stimulation with morphine pretreatment, with P⬍0.05 again considered to be significant. Fisher’s post hoc tests were used to assess differences between groups.
RESULTS
Formalin-induced nociceptive mapsThe present study used18
F-FDG microPET to elucidate the nociception-induced glucose metabolic changes in the brains of conscious rats. In order to further improve the
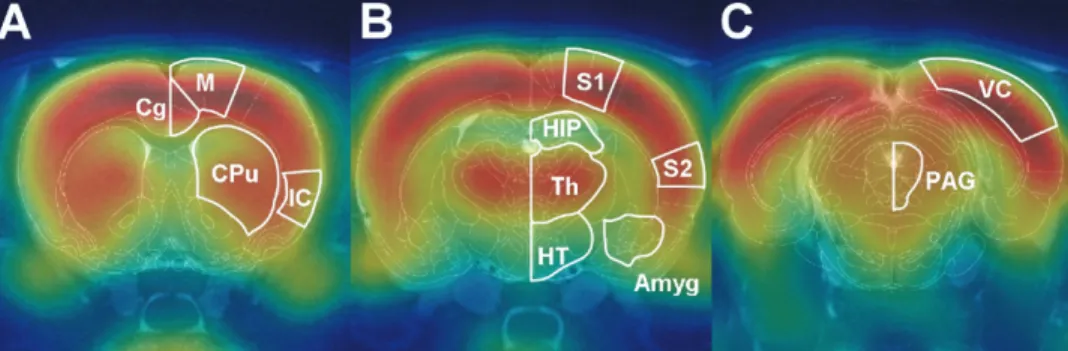
Fig. 1. Fusion of the rat brain atlas,18
F-FDG microPET images, and T2-weighted MRI images. These images provide anatomical alignment and
accuracy with which anatomical locations were deter-mined, microPET and MRI images were coregistered and fused with a digital atlas of the rat brain (Paxinos and Watson, 1998). This method allowed regions of interest to be selected based on clear spatial references (Fig. 1). Averaged formalin-induced metabolic maps overlaid on the MRI images are shown in Fig. 2A. The averaged
18
F-FDG uptake in the hind-limb area of the S1 (S1HL) was higher on the contralateral side than on the ipsilateral side. Statistical comparisons of the18
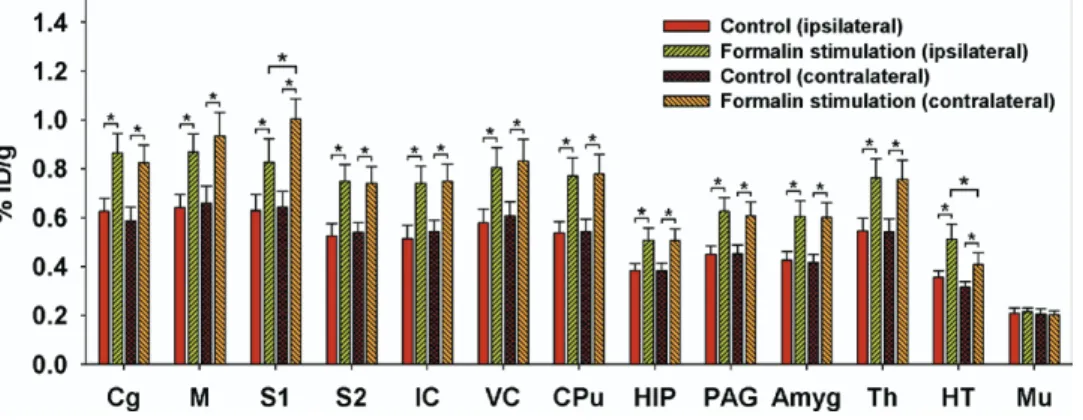
F-FDG uptake in the control and formalin groups are shown inFig. 3. Repeated-measures ANOVAs with Fisher’s post hoc tests indicated the presence of significant activations in the bilateral Cg, M, S1, S2, IC, VC, CPu, HIP, PAG, amygdala (Amyg), Th, and HT, whereas no changes were evident in muscle tissue (Mu). In addition, clear lateralization was only ob-served in S1 and HT (Fig. 3).
Antinociceptive effects of lidocaine and morphine
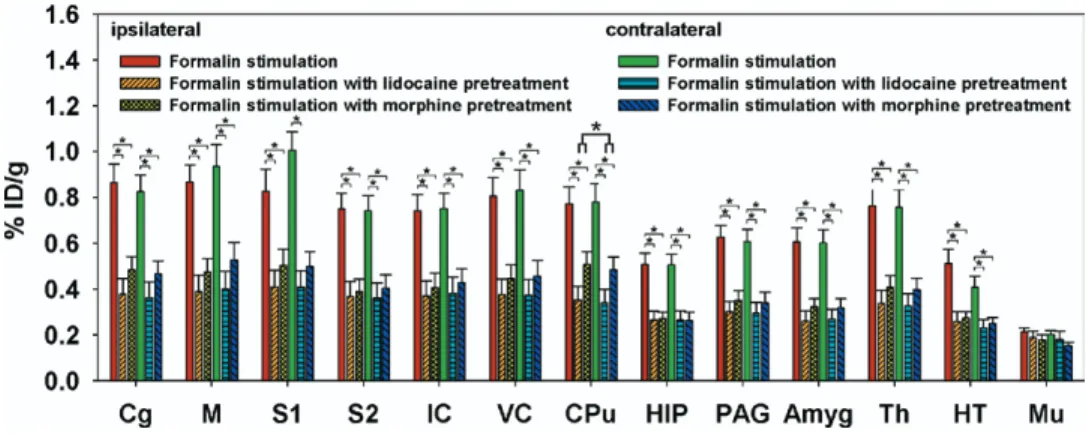
The effects of antinociceptive drugs are shown inFig. 2B, C, andFig. 4. These reduced the brain18
F-FDG uptake, with no clear lateralized differences evident in the sensory cortices. Factorial ANOVAs with Fisher’s post hoc tests have shown that18
F-FDG uptake in the bilateral Cg, M, S1, S2, IC, VC, CPu, HIP, PAG, Th, Amyg, and HT was lower for pretreatment with lidocaine and morphine than for
formalin stimulation alone. Among the measured areas,
18
F-FDG uptake in the CPu was higher for morphine treat-ment than for lidocaine treattreat-ment.
DISCUSSION
The present study clearly revealed formalin-induced noci-ceptive maps, with strong activations evident in the Cg, M, S1, S2, IC, VC, CPu, HIP, PAG, Amyg, Th, and HT. Although functional imaging techniques have been used previously to examine pain-related responses in rodent brains, such as a blood-flow-based autoradiographic method (Morrow et al., 1998) and fMRI (Tuor et al., 2000; Shah et al., 2005; Shih et al., in press, 2008b), the method employed in the current study provides new information and methodological improvements for preclinical pain re-search. First, our findings complement the previous pain studies in animals by revealing the relative metabolic changes instead of the responses related to blood flow. Second, the 18
F-FDG microPET technique provides an opportunity for in vivo, longitudinal follow-up of the re-sponse in the same animal, whereas the blood-flow-based autoradiographic method can only be applied once in each animal. Third, fMRI is usually performed under anesthesia, which would alter the representation of behavior-related brain functions and has been reported to strongly influence
Fig. 2. Incidence18F-FDG microPET maps overlaid on the MRI images showing the cerebral metabolic changes in three groups of rats, quantified
as the averaged %ID/g. (A) Averaged response of seven rats subjected to left hind-paw formalin stimulation alone. (B) Averaged response of six rats subjected to left hind-paw formalin stimulation with lidocaine pretreatment. (C) Averaged response of six rats subjected to left hind-paw formalin stimulation with morphine pretreatment. Clear lateralization is only evident in (A), in which the18F-FDG uptake is highest in the contralateral S1 and
the responsive region match with that of the hind-limb region defined by the rat brain atlas. The image position was 1.8 mm posterior to the bregma.
Fig. 3. Formalin-induced metabolic increases and cerebral laterality. Repeated-measures ANOVAs with Fisher’s post hoc tests indicated activations
in the bilateral Cg, M, S1, S2, IC, VC, CPu, HIP, PAG, Amyg, Th, and HT. Among the measured areas, clear lateralization was evident in S1 and HT, with18
F-FDG uptake being higher in the contralateral S1 than on the ipsilateral side, and in the ipsilateral HT than on the contralateral side. * Denotes P⬍0.05. Error bars represent S.E.M. values.
neuronal activity (Lindauer et al., 1993; Matsumura et al., 2003). Fourth, most fMRI studies in rodents have utilized invasive surgical procedures such as cannulation to con-trol anesthesia, deliver pharmacological compounds, or monitor physiological parameters. Neuronal activation might be affected by the associated surgical pain that would activate the responsive brain region prior to stimu-lation. In other words, neurovascular coupling around re-sponsive nuclei might already be in a responding state rather than a baseline state due to the surgical wound. These invasive procedures therefore inevitably generate confounding pain reactions. Moreover, if the invasive pro-cedures induce bleeding, the measured hemodynamic re-sponses are very likely to be affected by blood flow changes resulting from the alteration of blood pressure. It is therefore difficult to determine whether the observed activation responses are purely caused by the nociceptive stimulus. The current study avoided these invasive proce-dures, with the resulting activation responses appearing to be more distinguishable than in previous fMRI studies (Tuor et al., 2000; Shah et al., 2005; Shih et al., in press, 2008b). Lastly, fMRI is often criticized for the difficulty of detecting subcortical responses due to the position of the receiver coil, which usually results in stronger signals from cortical regions than from subcortical regions of the brain. Furthermore, the brain areas influenced by strong suscep-tibility artifacts such as the Amyg cannot be uniformly measured by fMRI. These disadvantages can be over-come by using microPET, which also shows better homo-geneity and fewer artifacts near the air-tissue interface.
Among the measured areas, bilateral activations of the IC and VC as well as S1, S2, Th, and Cg in the spinotha-lamic tract are consistent with fMRI data (Tuor et al., 2000; Malisza et al., 2003; Shah et al., 2005; Shih et al., in press, 2008b). Although fMRI can consistently detect these acti-vations, clear lateralization is difficult to be observed. Our results indicate that18
F-FDG microPET shows better de-lineation of the sensory-motor laterality of formalin-induced nociception than fMRI, since the 18
F-FDG uptake was significantly higher in the contralateral S1 than on the ipsilateral side (Figs. 2and3). A possible explanation of
this feature is the lack of anesthesia, and the use of a glucose analog rather than hemodynamic compensation, which is less sensitive to the blood-pressure-induced non-specific activation in fMRI (Tuor et al., 2002). It has also been suggested that the CPu and M are activated by painful stimuli (Chudler et al., 1993; Lorenz et al., 2003; Malisza et al., 2003), with bilateral activation in these areas possibly associated with the representation of mirror pain behavior in animals, where licking of the contralateral un-treated paw was observed following unilateral formalin stimulation (Aloisi et al., 1993).
In the subcortical regions, the PAG and HT were also activated, both of which are known to be intimately in-volved in pain modulation. The PAG is considered a major integration site for nociception signals and a center of analgesic action for opiates that influences the activity of the spinal cord dorsal horn through projections from the ventromedial medulla (Mason, 2005). Previous studies have shown that a nociception signal in the HT can simul-taneously excite the central autonomic nervous system, and that the HT might play a complex role in antinocicep-tive reactions (Pinto-Ribeiro et al., 2008). Our findings suggest that the18
F-FDG uptake was higher in the ipsilat-eral HT than on the contralatipsilat-eral side; however, the un-derlying mechanism remains obscure. Increased glucose metabolism in the Amyg during activation has been ob-served in a rat model of neuropathic pain (Mao et al., 1993). The findings of several studies indicate that the Amyg is associated with unconscious memorization of no-ciceptive stimuli and involved in emotional processes and automatic functions (Bingel et al., 2002; Alkire and Nathan, 2005; Neugebauer, 2007). As far as the activity in HIP is concerned, an immunohistochemical study has shown that a unilateral injection of formalin induced bilateral c-fos expression in the HIP, suggesting that the HIP is linked by numerous reciprocal neural connections and further impli-cating that this area is involved in the perception of chronic pain (Aloisi et al., 1997).
In addition to formalin-induced nociception, the present study also examined the antinociceptive effects of lido-caine and morphine.Fig. 4shows that pretreatment with
Fig. 4. Effects of lidocaine and morphine pretreatments on formalin-induced metabolic changes. Pretreatment with lidocaine and morphine reduced 18F-FDG uptake by different amounts. Factorial ANOVAs with Fisher’s post hoc tests indicated significant decreases in the bilateral Cg, M, S1, S2,
IC, VC, CPu, HIP, PAG, Amyg, Th, and HT compared with the groups subjected to formalin stimulation alone. * Denotes P⬍0.05. Error bars represent S.E.M. values.
lidocaine and morphine resulted in different reductions in
18
F-FDG uptake in the measured brain areas, indicating that formalin-induced metabolic changes can be inhibited by these two drugs. Moreover, the inhibition by lidocaine might also result from the blocking effect of voltage-gated sodium channels, thus reducing glucose metabolism glo-bally (Abdi et al., 1998; Gold and Thut, 2001; Rykacze-wska-Czerwinska, 2006), whereas morphine has been widely used for pain alleviation (Yaksh et al., 1988; Frank-lin et al., 1990; Lamas et al., 1994; Tuor et al., 2000; Shah et al., 2005) and also reportedly induces additional anes-thesia, thus attenuating glucose utilization in widespread brain regions (Cohen et al., 1991; Chudler and Dong, 1995). Although the mechanisms underlying widespread in-hibition in the brain are not fully understood, it cannot be ruled out that the down-regulation of global metabolic states is initiated by the brain processing of antinociceptive signals aimed at reducing sensory-motor reactions, and also emo-tional responses. Furthermore, the differences in the effects of lidocaine and morphine at the used dosages were largest in the CPu, which suggests that this region exhibits higher neuronal activities under morphine regulation.
CONCLUSION
The present study established an18
F-FDG microPET pro-tocol that allowed both serial and longitudinal measure-ments of nociception-induced metabolic changes in the brains of conscious rats. The formalin-induced nociception maps revealed several brain regions that are possibly involved in various aspects of pain processing. The whole-brain formalin-induced nociceptive responses following li-docaine and morphine pretreatment were also examined. This microPET imaging protocol should be useful in further evaluations of the effects of new pain-relief drugs and in developing preclinical therapeutic strategies for pain.
Acknowledgments—The authors acknowledge technical support from the Functional and Micro-Magnetic Resonance Imaging Cen-ter and Molecular-Genetic Imaging Core supported by the Na-tional Research Program for Genomic Medicine, NaNa-tional Science Council, Taiwan, PR China (3112-B-001-009 and NSC96-3112-B-001-004).
REFERENCES
Abdi S, Lee DH, Chung JM (1998) The anti-allodynic effects of ami-triptyline, gabapentin, and lidocaine in a rat model of neuropathic pain. Anesth Analg 87:1360 –1366.
Alkire MT, Nathan SV (2005) Does the amygdala mediate anesthetic-induced amnesia? Basolateral amygdala lesions block sevoflu-rane-induced amnesia. Anesthesiology 102:754 –760.
Aloisi AM, Porro CA, Cavazzuti M, Baraldi P, Carli G (1993) ‘Mirror pain’ in the formalin test: behavioral and 2-deoxyglucose studies. Pain 55:267–273.
Aloisi AM, Zimmermann M, Herdegen T (1997) Sex-dependent effects of formalin and restraint on c-Fos expression in the septum and hippocampus of the rat. Neuroscience 81:951–958.
Bingel U, Quante M, Knab R, Bromm B, Weiller C, Buchel C (2002) Subcortical structures involved in pain processing: evidence from single-trial fMRI. Pain 99:313–321.
Chudler EH, Dong WK (1995) The role of the basal ganglia in noci-ception and pain. Pain 60:3–38.
Chudler EH, Sugiyama K, Dong WK (1993) Nociceptive responses in the neostriatum and globus pallidus of the anesthetized rat. J Neu-rophysiol 69:1890 –1903.
Cohen SR, Kimes AS, London ED (1991) Morphine decreases cere-bral glucose utilization in limbic and forebrain regions while pain has no effect. Neuropharmacology 30:125–134.
Franklin KB, Abbott FV, English MJ, Jeans ME, Tasker RA, Young SN (1990) Tryptophan-morphine interactions and postoperative pain. Pharmacol Biochem Behav 35:157–163.
Gold MS, Thut PD (2001) Lithium increases potency of lidocaine-induced block of voltage-gated Na⫹currents in rat sensory neu-rons in vitro. Pharmacol Exp Ther 299:705–711.
Lamas X, Farre M, Moreno V, Cami J (1994) Effects of morphine in postaddict humans: a meta-analysis. Drug Alcohol Depend 36:147–152.
Lindauer U, Villringer A, Dirnagl U (1993) Characterization of CBF response to somatosensory stimulation: model and influence of anesthetics. Am J Physiol 264:H1223–H1228.
Lorenz J, Minoshima S, Casey KL (2003) Keeping pain out of mind: the role of the dorsolateral prefrontal cortex in pain modulation. Brain 126:1079 –1091.
Malisza KL, Gregorash L, Turner A, Foniok T, Stroman PW, Allman AA, Summers R, Wright A (2003) Functional MRI involving painful stimulation of the ankle and the effect of physiotherapy joint mo-bilization. Magn Reson Imaging 21:489 – 496.
Mao J, Mayer DJ, Price DD (1993) Patterns of increased brain activity indicative of pain in a rat model of peripheral mononeuropathy. J Neurosci 13:2689 –2702.
Mason P (2005) Deconstructing endogenous pain modulations. J Neu-rophysiol 94:1659 –1663.
Matsumura A, Mizokawa S, Tanaka M, Wada Y, Nozaki S, Nakamura F, Shiomi S, Ochi H, Watanabe Y (2003) Assessment of microPET performance in analyzing the rat brain under different types of anes-thesia: comparison between quantitative data obtained with micro-PET and ex vivo autoradiography. Neuroimage 20:2040 –2050. McMahon SB, Koltzenburg M (2005) Wall and Melzack’s textbook of
pain (McMahon SB, Koltzenburg M, eds), Oxford, UK: Elsevier Churchill Livingstone.
Morrow TJ, Paulson PE, Danneman PJ, Casey KL (1998) Regional changes in forebrain activation during the early and late phase of formalin nociception: analysis using cerebral blood flow in the rat. Pain 75:355–365.
Neugebauer V (2007) The amygdala: different pains, different mech-anisms. Pain 127:1–2.
Ogawa S, Lee TM, Kay AR, Tank DW (1990a) Brain magnetic reso-nance imaging with contrast dependent on blood oxygenation. Proc Natl Acad Sci U S A 87:9868 –9872.
Ogawa S, Lee TM, Nayak AS, Glynn P (1990b) Oxygenation-sensitive contrast in magnetic resonance image of rodent brain at high magnetic fields. Magn Reson Med 14:68 –78.
Ohashi K, Ichikawa K, Chen L, Callahan M, Zasadny K, Kurebayashi Y (2008) MicroPET detection of regional brain activation induced by colonic distention in a rat model of visceral hypersensitivity. J Vet Med Sci 70:43– 49.
Paxinos G, Watson C (1998) The rat brain in stereotaxic coordinates: San Diego: Academic Press.
Phelps ME (2000) PET: the merging of biology and imaging into molecular imaging. J Nucl Med 41:661– 681.
Phelps ME, Huang SC, Hoffman EJ, Selin C, Sokoloff L, Kuhl DE (1979) Tomographic measurement of local cerebral glucose met-abolic rate in humans with (F-18)2-fluoro-2-deoxy-D-glucose: val-idation of method. Ann Neurol 6:371–388.
Pinto-Ribeiro F, Ansah OB, Almeida A, Pertovaara A (2008) Influence of arthritis on descending modulation of nociception from the para-ventricular nucleus of the hypothalamus. Brain Res 1197:63–75. Qi J, Leahy RM, Cherry SR, Chatziioannou A, Farquhar TH (1998)
High-resolution 3D bayesian image reconstruction using the mi-croPET small-animal scanner. Phys Med Biol 43:1001–1013.
Rykaczewska-Czerwinska M (2006) Antinociceptive effect of lidocaine in rats. Pharmacol Rep 58:961–965.
Schiffer WK, Mirrione MM, Dewey SL (2007) Optimizing experimental protocols for quantitative behavioral imaging with 18F-FDG in ro-dents. J Nucl Med 48:277–287.
Shah YB, Haynes L, Prior MJ, Marsden CA, Morris PG, Chapman V (2005) Functional magnetic resonance imaging studies of opioid receptor-mediated modulation of noxious-evoked BOLD contrast in rats. Psychopharmacology (Berl) 180:761–773.
Shih YY, Chang C, Chen JC, Jaw FS (2008a) BOLD fMRI mapping of brain responses to nociceptive stimuli in rats under ketamine anesthesia. Med Eng Phys, in press.
Shih YY, Chen YY, Chen CC, Chen JC, Chang C, Jaw FS (2008b) Whole-brain functional magnetic resonance imaging mapping of acute nocicep-tive responses induced by formalin in rats using atlas registration-based event-related analysis. J Neurosci Res 86:1801–1811.
Shih YY, Chen YY, Chen JC, Chang C, Jaw FS (2007) ISPMER: Integrated system for combined PET, MRI, and electrophysiologi-cal recording in somatosensory studies in rats. Nucl Instrum Meth-ods A 580:938 –943.
Tjolsen A, Berge OG, Hunskaar S, Rosland JH, Hole K (1992) The formalin test: an evaluation of the method. Pain 51:5–17. Tuor UI, Malisza K, Foniok T, Papadimitropoulos R, Jarmasz M,
Somorjai R, Kozlowski P (2000) Functional magnetic reso-nance imaging in rats subjected to intense electrical and nox-ious chemical stimulation of the forepaw. Pain 87:315– 324.
Tuor UI, McKenzie E, Tomanek B (2002) Functional magnetic reso-nance imaging of tonic pain and vasopressor effects in rats. Magn Reson Imaging 20:707–712.
Yaksh TL, Al-Rodhan NR, Jensen TS (1988) Sites of action of opiates in production of analgesia. Prog Brain Res 77:371–394.
(Accepted 7 July 2008) (Available online 11 July 2008)