Cyclosporine-A enhances gingival ß-catenin stability via Wnt
signaling
Hsiao-Pei Tu*, †, Yen-Teen Chen*, ‡, Earl Fu*, E-Chin Shen§, Meng-Hsun Wu∥, Yen-Lin Chen¶, Cheng-Yang Chiang*, Hsien-Chung Chiu*
*: Department of Periodontology, School of Dentistry, National Defense Medical Center and Tri-Service General Hospital, Taipei, Taiwan.
†: Department of Dental Hygiene, China Medical University, Taichung, Taiwan, ROC
‡: Department of Education and Research, Cardinal Tien Hospital, New Taipei City, Taiwan, R.O.C.
§: Dental Department, Buddhist TZU CHI General Hospital, Sindian, Taipei, Taiwan, ROC
:
∥ Institute of Molecular and Genomic Medicine, National Health Research Institutes, Miaoli, Taiwan, ROC
¶: Department of Pathology, Cardinal Tien Hospital, School of Medicine, Fu-Jen Catholic University, New Taipei City, Taiwan
Correspondence to: Dr. Hsien-Chung Chiu School of Dentistry
National Defense Medical Center
PO Box 90048-507, Taipei, Taiwan, ROC Tel: +886-2-87927150
Fax: +886-2-87927145
Email: [email protected]
One sentence summary:
APC/Axin1 inhibition and Dvl-1 induction in rat gingiva indicate that cyclosporine-A enhances ß-catenin stability via Wnt signaling.
Figures: 4 Tables: 1
References: 35 Words: 2657
_________________________________________________________________ June 30, 2014 Journal of Periodontology
ABSTRACT
Background: We have found that cyclosporine-A Cyclosporine-A (CsA) increases ß-catenin mRNA and protein expression. The present study demonstrates that Wingless-Int (Wnt)/ß-catenin signaling inhibits ß-catenin degradation in the gingiva.
Methods: Forty 5-week-old male Sprague–Dawley rats were assigned to two study groups after
healing from following extraction of the right maxillary molars extractions. The rats in the experimental CsA group were fed 30 mg/kg CsA daily, whereas the control rats were fed mineral oil for 4 weeks. At the end of the study, all rats were sacrificed and the gingivae were obtained. The gingival morphology after CsA treatment was evaluated by histology, and the genes related to Wnt/ß-catenin signaling were initially screened by microarray. PCR, western blotting, and immunohistochemistry were used to examine the mRNA and protein expression of proliferating cell nuclear antigen (Pcna), cyclin D1, E-cadherin, ß-catenin, disheveled (Dvl)-1, glycogen synthase kinase 3β (GSK3β), Axin-1, and adenomatous polyposis coli (APC). Phosphoserine and ubiquitinylated ß-catenin were detected after immunoprecipitation.
Results: In rats treated with CsA, overgrowth overgrown of gingivae were observed and altered expression of genes related to Wnt/β-catenin signaling was detected by the microarray. The gingival mRNA and protein expression profiles for genes associated with Wnt/β-catenin signaling further confirmed the effect of CsA: β-catenin and Dvl-1 expression increased, but APC and Axin1 expression decreased. Western blotting and immunohistochemistry showed decreases in ß-catenin serine phosphorylation (33/37) and ubiquitinylation in the gingivae of CsA-treated rats.
Conclusion: Our results suggest that CsA-enhanced gingival β-catenin stability may be involved in
gene up-regulation or β-catenin degradation via the Wnt/ß-catenin pathway.
Key Words
INTRODUCTION
Gingival overgrowth, characterized by epithelial hyperplasia, interstitial fibrosis, blood vessel profile changes, and focal inflammatory cell infiltration, is a side effect of cyclosporine A (CsA) therapy . Various direct and/or indirect interactions between CsA and gingival cells have been investigated, including metabolic and synthetic effects3-6. CsA directly augments gingival epithelial cell proliferation, whereas over-expression of growth factors, including keratinocyte growth factor and epithelial growth factor, may also be indirectly associated with epithelial proliferation in overgrown gingiva 7-9. However, the mechanisms underlying these processes remain unknown.
The Wingless-Int (Wnt)/β-catenin signaling pathway controls cell-to-cell communication in the embryo and adult (i.e., cell proliferation and differentiation during development and healing) 10. Evidence suggests that in the absence of Wnt, β-catenin is phosphorylated by glycogen synthase kinase 3β (GSK3β) in a cytoplasmic destruction complex that contains adenomatous polyposis coli (APC) and Axin-1, targeting β-catenin for ubiquitination and degradation. When Wnt binds to the frizzled receptor, disheveled (Dvl) is hyperphosphorylated and activated. Activated Dvl-1 inhibits GSK3β activity and phosphorylation of Axin-1, APC, and β-catenin, and it stabilizes β-catenin, which is then translocated to the nucleus where it stimulates the expression of Wnt-responsive genes.
Our previous study showed that CsA inhibits E-cadherin expression in rat gingiva and may thus induce epithelial cell proliferation through free β-catenin 11. In this study, a hypothesis that CsA enhances gingival ß-catenin stability via Wnt signaling is proposed. In this study, We therefore assessed the expression of proteins associated with the Wnt/β-catenin signaling pathway, including Dvl-1, GSK-3ß,
Axin, and APC, in the gingivae of CsA-treated rats. We also determined the levels of serine-phosphorylated (33/37) and ubiquitinylated β-catenin.
MATERIALS AND METHODS Animal experiment
Forty of five-week-old male Sprague–Dawley rats, weighing 120–150 g, were randomly assigned and divided to experimental CsA and control groups after a 3-week healing period following extraction of all right maxillary molars as described in our previous study 12. Animals in the experimental CsA group received CsA# (30 mg/kg body weight in mineral oil) daily by gastric feeding for 4 weeks, whereas the control group received mineral oil only. At the end of the experiment, all animals were sacrificed using carbon dioxide inhalation. Five gingival specimens from each group were collected and pooled for the initial microarray screen for genes related to the Wnt/β-catenin pathway. The other specimens, 15 per group, were divided and either stored and prepared for mRNA and protein assays or used for histology and immunohistochemistry (IHC). Proliferating cell nuclear antigen (Pcna), cyclin D1, E-cadherin, ß-catenin, Dvl-1, GSK3β, Axin-1, and APC, as well as phosphorylated and ubiquitinylated ß-catenin, were selected for investigation. All animals were kept in a specific pathogen-free facility and handled according to protocols approved by Institutional Animal Care and Use Committee, National Defense Medical Center, Taipei, Taiwan (IACUC-05-194).
RNA extraction and microarray analysis
Gingival samples were homogenized in TRIzol reagent** with a glass homogenizer, and total RNA was quantified by spectrophotometry at 260 nm. cDNA was generated with reverse transcriptase enzyme SuperScript III** and the GeneAmp 9700®-PCR System polymerase chain reaction (PCR) machine††. The cDNA was
labeled with CyDye by using the MessageAmp aRNA Kit‡‡. Reference cRNA from the control group was labeled with Cy3; cRNA from the CsA-treated group was labeled with Cy5. Labeled aRNA was purified to remove uncoupled CyDyes before being applied to the microarray. Hybridization was performed on the microarray Rat Oligo 1A Microarray (G4130A)§§ containing approximately 22,000 oligonucleotides that represent more than 17,000 genes. Hybridization and wash conditions were designed according to the standard Agilent 60-mer oligo microarray processing protocol§§.
Semi-quantitative and real-time quantitative RT-PCR
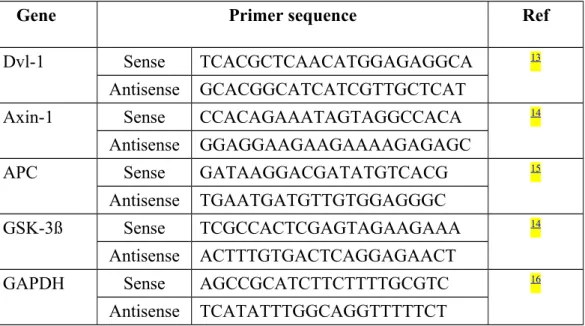
The cDNAs were compared by quantitative PCR under the following cycling conditions: denaturation at 94°C, annealing at 58–64°C, and extension at 72°C. The PCR primers for rat Dvl-1, Axin-1, APC, GSK-3ß and GAPDH were shown in (Table 1). follows: rat Dvl 1 primers 13, and; rat Axin-1 primers 14, 5′-CCACAGAAATAGTAGGCCACA-3′ and 5′-GGAGGAAGAAGAAAAGAGAGC-3′; rat APC primers 15, GATAAGGACGATATGTCACG-3′ and 5′-TGAATGATGTTGTGGAGGGC-3′; rat GSK-3ß primers 14, 5′-TCGCCACTCGAGTAGAAGAAA-3′ and 5′-ACTTTGTGACTCAGGAGAACT-3′; and rat glyceraldehyde-3-phosphate dehydrogenase (GAPDH) primers, 5′-AGCCGCATCTTCTTTTGCGTC-3′ and 5′-TCATATTTGGCAGGTTTTTCT-3′ 16. Amplified products were analyzed on agarose gels stained with ethidium bromide (EtBr). The RT-PCR product bands were scanned and the expression levels were normalized to those of glyceraldehyde-3-phosphate dehydrogenase (GAPDH).
Gene expression of Pcna, cyclin D1, and E-cadherin was confirmed by real-time PCR machine∥∥13. In brief, the probes and primers for rat Pcna, cyclin D1, E-cadherin,
and β-catenin were selected from primers available with the TaqMan Assay-on-Demand gene expression commercial kit¶¶. Reactions were performed in triplicate in 50-µL reaction volumes of 1× PCR buffer A, 2.5 mM MgCl2, each primer at 0.4 µM, each dNTP at 200 µM, 100 nM probe, and 0.025 U/µL Taq Gold PCR enzyme. Each primer/probe set (5–10 µL) was added, and the reactions were performed under the following conditions: 95°C for 12 min followed by 40 cycles of 95°C for 20 s and 60°C for 1 min. The data were analyzed using sequence detection software that calculated the threshold cycle for each reaction 14.
Immunohistochemistry
After deparaffinization and hydration, tissue sections were boiled in antigen retrieval DAKO buffer ## for 15 min for antigen retrieval. Endogenous peroxidase activity was quenched by incubation for 5 min with 0.1% hydrogen peroxide in distilled water. Tissue sections were washed twice for 5 min in phosphate-buffered saline (PBS). The sections were incubated for 2 h with unconjugated primary monoclonal and polyclonal antibodies for Pcna***, cyclin D1 ***, E-cadherin †††, Dvl-1 ***, GSK-3ß ‡‡‡, Axin 1§§§, APC***, and β-catenin phosphoserine (33/37) †††17, followed by incubation with the biotinylated secondary antibody, streptavidin-conjugated horseradish peroxidase (HRP) complexes, and 3-amino-9-ethyl carbazole (AEC) solution for an additional 30 min, 30 min, and 10 min, respectively. Between incubation periods, the cells and sections were washed with sterile PBS. The specimens were then counterstained with hematoxylin, dehydrated, and mounted. Cells stained for Dvl-1, GSK-3ß, Axin-1, APC, and β-catenin phosphoserine (33/37) were examined by fluorescence microscopy.
We used a motor-driven Teflon-glass homogenizer to homogenize gingival tissues at 4°C. The homogenates were centrifugedat 13,000 g for 10 min at 5°C, total protein supernatants were collected, and protein concentrations were determined by the Bradford method 18. Western blotting was performed as previously described 19. The protein samples were resolved by 10% SDS-PAGE and transferred to Millipore Immobilon-PSQ Transfer PVDF membranes¶¶¶ with an electrical protein transfer machine###. a Hoefer Semi-Dry Transfer system. The membranes were blocked with a solution of 5% nonfat milk and 1% Tween 20 in Tris-buffered saline; incubated with primary antibodies to E-cadherin†††, β-catenin†††, phosphorylated serine (33/37) of β-catenin†††, and α-actin∥∥∥ at 4C overnight; and washed with PBS. HRP-conjugated secondary antibodies and ImmobilonTM Western HRP Substrate Luminol Reagent**** were used for antibody detection. Images were visualized by autoradiography.
For immunoprecipitation, 500 µg of protein from eachpreparation was used after the additionof 4 µg of β-catenin antibody‡‡‡ (Cell Signaling Technology)for 24 hat 4°C. Protein A-Sepharose 4 Fast Flow Beads(30 µL)30ul of beads†††† were added to each sample; after incubation by rotation at 4°C for 1 h, the immunoprecipitated complexes were collected by centrifugation at 3,000 rpm1,600g for 5 min. The complexes were separated by 12% SDS-PAGE and blotted with an anti-ubiquitin antibody‡‡‡, and β-catenin degradation in the gingiva after CsA treatment was examined.
Statistical Analysis
Student’s t-test was used to evaluate differences in mRNA, protein expression, and histological tissue areas in the control and CsA groups. P < 0.05 represented a significant difference.
# : Sandimmune, Sandoz, Basel, Switzerland **: Invitrogen Corporation, Carlsbad, CA, USA ††: Applied Biosystems, Foster City, CA, USA ‡‡: Ambion, Houston, TX, USA
§§: Agilent Technologies, Santa Clara, CA, USA :
∥∥ Applied Biosystems, Foster City, CA, USA
¶¶: TaqMan Assay-on-Demand gene expression, Applied Biosystems, Foster City, CA, USA Life Technologies, San Diego, CA, USA
##: DakoCytomation, San Diego, CA, USA
***: Santa Cruz Biotechnology, Inc. Santa Cruz, CA, USA †††: BD Biosciences, San Jose, CA, USA
‡‡‡: Cell Signaling Technology, Inc., Beverly, MA §§§: Biorbyt, Cambridge, UK
:
∥∥∥ Abcam, Cambridge, MA, USA
¶¶¶: Immobilon-PSQ Transfer PVDF membranes, Millipore, Billerica, MA ###: Hoefer Semi-Dry Transfer system, Hoefer, Inc., Holliston, MA, USA. ****: ImmobilonTM Western HRP Substrate Luminol Reagent, Millipore, Billerica, MA
RESULTS
Wnt/β-catenin-associated gene expression in CsA-induced overgrown gingiva
Compared to the control rats, CsA-treated rats showed increased gingival tissue area, whereas epithelial and connective tissue area remained unchanged (Figure 1A) areas, including the epithelial, connective tissue, and total soft tissue areas, whereas the ratio of the epithelial to connective tissue area remained unchanged (Figure 1A). In the gingiva, the expression of genes related to the Wnt/β-catenin pathway was initially screened and compared between the control and CsA-treated rats by microarray (Figure 1B). Expression of Dvl-1 (1.27) and GSK-3ß (1.35) increased in the gingivae of CsA-treated rats compared to that in control rats, whereas expression of E-cadherin (0.95), APC (0.79), and Axin-1 (0.81) was reduced. Expression of Pcna (1.11) and cyclin D1 (1.20), downstream genes in the Wnt/β-catenin pathway, was also increased. The increased expression of Pcna and cyclin D1 in the gingiva of CsA-treated rats was confirmed by RT-PCR and IHC staining (Figure 1C).
CsA and expression of proteins associated with Wnt/ß-catenin signaling
Gingival β-catenin mRNA and protein expression was increased in CsA-fed rats (Figure 2A-C). Genes associated with Wnt/β-catenin signaling were affected by CsA; Dvl-1 expression was increased (Figure 3A), but that of APC and Axin-1 was decreased in CsA-treated rats (Figure 3B). Western blotting showed a reduction in serine phosphorylation (33/37) in rats treated with CsA relative to control rats (Figure 4A). Moreover, the ubiquitinylation of β-catenin was greatly reduced in the gingivae of treated rats, whereas the β-catenin level did not differ between the CsA-treated rats and the control rats (Figure 4B).
DISCUSSION
We studied the network of proteins associated with the Wnt/β-catenin signaling pathway in rat edentulous gingivae to understand its role in CsA-induced gingival changes changes. in the previous studies . Although the gingival enlargement is usually pronounced at sites with teeth in human, the severe mucosa hyperplasia in the edentulous maxilla associated with CsA therapy was also reported21. In our previous study, we have demonstrated the CsA induced overgrowth could be observed on either the interdental papilla or the edentulous gingiva in this rat model. The Wnt/β-catenin signaling Signaling is initiated by binding of Wnt ligand to its receptors, the frizzled proteins and lipoprotein receptor-related proteins 22, which trigger a cascade that results in the displacement of GSK-3β from the APC/Axin/GSK-3ß complex (Figure 4C) . In the absence of Wnt signaling (off-state), β-catenin is targeted for degradation by the complex. Appropriate phosphorylation of β-catenin by coordinated action of GSK-3ß and casein kinase 1 leads to its ubiquitination and proteasomal degradation. In the presence of Wnt binding (on-state), Dvl is activated by phosphorylation and recruits GSK-3ß from the degradation complex, which allows β-catenin to be stabilized and translocated to the nucleus, where it stimulates expression of Wnt-responsive genes 25.
Significantly increased higher gingival expression of Dvl-1 was observed in CsA-treated rats than in the controls (Figure 3A). This finding along with and the significant decrease in APC and Axin expression (Figure 3B) suggest that CsA inhibits formation of the β-catenin destruction complex, thereby increasing cytoplasmic β-catenin levels. However, the analysis of GSK-3β yielded conflicting results. Although microarray analysis showed increased GSK-3β expression in CsA
rats (Figure 1B), RT-PCR revealed a non-significant decrease, and IHC showed a meaningful decrease (Figure 3B). The protein half-life of GSK-3β may account for these discrepancies, but the mechanism remains unclear at this time. Nevertheless, the reduction of catenin phosphorylation (Figure 4) and hypophosphorylation of β-catenin in the gingiva of CsA-treated rats suggests stability against ubiquitin-mediated degradation.
A large complex that includes APC and Axin binds to GSK-3ß and CK1 to phosphorylate ß-catenin, thereby reducing its ß-catenin. The APC protein normally forms a complex with GSK-3ß and Axin via interactions with a 20-amino acid segment and SAMP repeats. This complex is then able to bind ß-catenin in the cytoplasm. With the help of casein CK1 CK1, which performs an initial phosphorylation of ß-catenin, GSK-3ß is able to phosphorylate ß-catenin a second time, which targets ß-catenin for ubiquitination and degradation by cellular proteosomes and thus prevents ß-catenin from translocating into the nucleus, where it acts as a transcription factor for proliferation-associated genes. Mutations in the APC gene typically lead to a truncated APC protein that lacks the ß-catenin binding site and therefore cannot degrade ß-catenin. 26
Axin is a negative regulator of the Wnt signaling pathway. Extensive evidence has shown that rat Axin (rAxin) and its homolog, Axil (for Axin-like), function as GSK-3ß-interacting proteins. Both Axin and Axil bind not only to GSK-3ß but also to beta-catenin and they promote GSK-3ß-dependent phosphorylation of beta-catenin. In addition, the regulators of G protein signaling (RGS) domain of rAxin directly interacts with APC, and expression of rAxin in COS and SW480 cells stimulates the degradation of ß-catenin.32-35 In this manner, Axin family members downregulate ß-catenin. In the present study, RT-PCR and IHC indicated a significant decrease in the
expression of APC and Axin-1 (Figure 3B). These results imply that CsA may decrease the formation of the complex that degrades ß-catenin, thereby increasing the amount of the free form of cytoplasmic ß-catenin.
Taken together, our results demonstrated that CsA increased the ß-catenin expression(Figure 2A-C), and decreased the ß-catenin degradation (Figure 4A-B). In combined with the downregulation of E-cadherin, which was also induced by CsA11, CsA lead to increased free form of ß-catenin, which translocated to cell nuclear and modulate the cell proliferation, as well as the gingival morphological changes.
In summary, we examined the network of proteins associated with the Wnt/β-catenin signaling pathway in the gingivae of CsA-treated rats. CsA increased the gingival expression of Dvl-1, whereas the expression of Axin and APC was reduced. This finding and the decrease in levels of phosphorylated and ubiquitinylated ß-catenin lead us to suggest that CsA enhances β-ß-catenin stability via the Wnt/β-ß-catenin pathway and increases β-catenin gene expression. These findings shed new light on the development of CsA-induced gingival overgrowth.
ACKNOWLEDGMENT
This study was supported in part by the National Science Council, Republic of China (NSC-94-2314-B-016-033), and the C.Y. Foundation for Advancement of Education, Sciences, and Medicine Taipei, Taiwan, Republic of China. The authors report no conflicts of interest related to this study.
Legends
Figure 1. CsA treatment and expression of genes associated with cell proliferation in rat gingiva. (A) Area of gingival tissue in control and CsA-treated rats (H & E staining, magnification at ×10) (Means and standard deviations, *: significant difference at p < 0.05). (B) Gingival expression profile of genes associated with cell proliferation in control and CsA-treated rats determined using micro-array analysis. (C) mRNA and protein expression of Pcna and cyclin D1 in gingivae of rats (means and standard deviations, *: significant difference at p < 0.001, and IHC images: original magnification ×25).
Figure 2. CsA treatment and expression of ß-catenin in rat gingiva. (A-B) mRNA and protein expression for β-catenin in gingivae of control and CsA-treated rats (means and standard deviations, *: significant difference at p < 0.01). (C) Expression and distribution of ß-catenin in the rat gingiva, examined by IHC (original magnification ×25).
Figure 3. CsA treatment and expression of genes associated with β-catenin degradation in the rat gingiva. (A) mRNA and protein expression of Dvl-1 in gingivae of control and CsA-treated rats, determined by RT-PCR and IHC (original magnification ×25). (B) mRNA and protein expression of GSK-3β, APC, and Axin-1 in rat gingiva (means and standard deviations, *significant difference at P < 0.001). Figure 4. Expression of serine-phosphorylated (33/37) and ubiquitinylated β-catenin in rat gingiva. (A) Expression of serine-phosphorylated (33/37) and ubiquitinylated β-catenin in gingivae of control and CsA-treated rats, examined by Western blot after IP (means and standard deviations, * and **: significant difference at P < 0.05 and P < 0.001, respectively). (B) Expression of serine-phosphorylated (33/37) ß-catenin in gingivae from a control rat and a CsA-treated rat (IHC staining, original magnification ×25). (C) A model of CsA-enhanced β-catenin expression in a gingival cell:inhibitionof β-catenindegradation via the Wnt/β-catenin pathway.
Table 1: The primers used in RT-PCR for rat Dvl 1, Axin-1, APC, GSK-3ß and GAPDH.
Gene Primer sequence Ref
Dvl-1 Sense TCACGCTCAACATGGAGAGGCA 13
Antisense GCACGGCATCATCGTTGCTCAT
Axin-1 Sense CCACAGAAATAGTAGGCCACA 14
Antisense GGAGGAAGAAGAAAAGAGAGC
APC Sense GATAAGGACGATATGTCACG 15
Antisense TGAATGATGTTGTGGAGGGC
GSK-3ß Sense TCGCCACTCGAGTAGAAGAAA 14
Antisense ACTTTGTGACTCAGGAGAACT
GAPDH Sense AGCCGCATCTTCTTTTGCGTC 16
References:
1. Rateitschak-Pluss EM, Hefti A, Lortscher R, Thiel G. Initial observation that cyclosporin-A induces gingival enlargement in man. J Clin Periodontol 1983;10:237-246.
2. Deliliers GL, Santoro F, Polli N, Bruno E, Fumagalli L, Risciotti E. Light and electron microscopic study of cyclosporin A-induced gingival hyperplasia. J
Periodontol 1986;57:771-775.
3. Chin YT, Tu HP, Chen YT, et al. Expression and bioactivities of endothelin-1 in gingiva during cyclosporine A treatment. J Periodontal Res 2009;44:35-42. 4. Chiang CY, Tu HP, Chen YT, et al. Up-regulation of retinoblastoma protein
phosphorylation in gingiva after cyclosporine A treatment: an in vivo and in vitro study. J Periodontal Res 2011;46:158-163.
5. Chiu HC, Lu YT, Chin YT, et al. Cyclosporine A inhibits the expression of membrane type-I matrix metalloproteinase in gingiva. J Periodontal Res 2009;44:338-347.
6. Bartold PM. Regulation of human gingival fibroblast growth and synthetic activity by cyclosporine-A in vitro. J Periodontal Res 1989;24:314-321. 7. Tu HP, Fu E, Chen YT, Wu MH, Cheng LC, Yang SF. Expression of p21 and
p53 in rat gingival and human oral epithelial cells after cyclosporine A treatment. J Periodontal Res 2008;43:32-39.
8. Hyland PL, McKeown ST, Mackenzie IC, Irwin CR. Regulation of keratinocyte growth factor and scatter factor in cyclosporin-induced gingival overgrowth. J Oral Pathol Med : official publication of the International
Association of Oral Pathologists and the American Academy of Oral Pathology 2004;33:391-397.
9. Chin YT, Chen YT, Tu HP, et al. Upregulation of the expression of epidermal growth factor and its receptor in gingiva upon cyclosporin A treatment. J
Periodontol 2006;77:647-656.
10. Logan CY, Nusse R. The Wnt signaling pathway in development and disease.
Annu Rev Cell Dev Biol 2004;20:781-810.
11. Tu HP, Chen YT, Shieh YS, et al. Cyclosporin-induced downregulation of the expression of E-cadherin during proliferation of edentulous gingival epithelium in rats. J Periodontol 2006;77:832-839.
12. Fu E, Hsieh YD, Shen EC, Nieh S, Mao TK, Chiang CY. Cyclosporin-induced gingival overgrowth at the newly formed edentulous ridge in rats: a morphological and histometric evaluation. J Periodontol 2001;72:889-894. 13. Zhou X, Hu X, Xie J, Xu C, Xu W, Jiang H. Exogenous high-mobility group
box 1 protein injection improves cardiac function after myocardial infarction: involvement of Wnt signaling activation. J Biomed Biotechnol 2012;2012:743879.
14. Sklepkiewicz P, Schermuly RT, Tian X, et al. Glycogen synthase kinase 3beta contributes to proliferation of arterial smooth muscle cells in pulmonary hypertension. PloS one 2011;6:e18883.
15. Kishimoto Y, Takata N, Jinnai T, et al. Sulindac and a cyclooxygenase-2 inhibitor, etodolac, increase APC mRNA in the colon of rats treated with azoxymethane. Gut 2000;47:812-819.
16. Drissi H, Lomri A, Lasmoles F, Holy X, Zerath E, Marie PJ. Skeletal unloading induces biphasic changes in insulin-like growth factor-I mRNA levels and osteoblast activity. Exp Cell Res 1999;251:275-284.
17. Gu Y, Turck CW, Morgan DO. Inhibition of CDK2 activity in vivo by an associated 20K regulatory subunit. Nature 1993;366:707-710.
18. Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal
Biochem 1976;72:248-254.
19. Chiu HC, Lan GL, Chiang CY, et al. Upregulation of heme oxygenase-1 expression in gingiva after cyclosporin A treatment. J Periodontol 2008;79:2200-2206.
20. Chen YT, Tu HP, Chin YT, et al. Upregulation of transforming growth factor-beta1 and vascular endothelial growth factor gene and protein expression in cyclosporin-induced overgrown edentulous gingiva in rats. J Periodontol 2005;76:2267-2275.
21. Gelbard S, Aoskar Y, Zalkind M, Stern N. Effect of impression materials and techniques on the marginal fit of metal castings. J Prosthet Dent 1994;71:1-6. 22. Nelson WJ, Nusse R. Convergence of Wnt, beta-catenin, and cadherin
pathways. Science 2004;303:1483-1487.
23. Mosimann C, Hausmann G, Basler K. Beta-catenin hits chromatin: regulation of Wnt target gene activation. Nat Rev Mol Cell Biol 2009;10:276-286.
24. Angers S, Moon RT. Proximal events in Wnt signal transduction. Nat Rev Mol
Cell Biol 2009;10:468-477.
25. Willert K, Jones KA. Wnt signaling: is the party in the nucleus? Genes Dev 2006;20:1394-1404.
26. Polakis P. Wnt signaling and cancer. Genes Dev 2000;14:1837-1851.
27. Cadigan KM, Nusse R. Wnt signaling: a common theme in animal development. Genes Dev 1997;11:3286-3305.
polyposis coli interaction. EMBO J 2000;19:2270-2279.
29. Roberts DM, Pronobis MI, Poulton JS, et al. Deconstructing the sscatenin destruction complex: mechanistic roles for the tumor suppressor APC in regulating Wnt signaling. Mol Biol Cell 2011;22:1845-1863.
30. Yamamoto H, Kishida S, Uochi T, et al. Axil, a member of the Axin family, interacts with both glycogen synthase kinase 3beta and beta-catenin and inhibits axis formation of Xenopus embryos. Mol Cell Biol 1998;18:2867-2875.
31. Ikeda S, Kishida S, Yamamoto H, Murai H, Koyama S, Kikuchi A. Axin, a negative regulator of the Wnt signaling pathway, forms a complex with GSK-3beta and beta-catenin and promotes GSK-GSK-3beta-dependent phosphorylation of beta-catenin. EMBO J 1998;17:1371-1384.
32. Behrens J, Jerchow BA, Wurtele M, et al. Functional interaction of an axin homolog, conductin, with beta-catenin, APC, and GSK3beta. Science 1998;280:596-599.
33. Hart MJ, de los Santos R, Albert IN, Rubinfeld B, Polakis P. Downregulation of beta-catenin by human Axin and its association with the APC tumor suppressor, beta-catenin and GSK3 beta. Curr Biol 1998;8:573-581.
34. Kishida M, Koyama S, Kishida S, et al. Axin prevents Wnt-3a-induced accumulation of beta-catenin. Oncogene 1999;18:979-985.
35. Kishida S, Yamamoto H, Ikeda S, et al. Axin, a negative regulator of the wnt signaling pathway, directly interacts with adenomatous polyposis coli and regulates the stabilization of beta-catenin. J Biol Chem 1998;273:10823-10826.
Figure 1.
A. Histomorphology in rat gingiva (means ± SDs) (*: significant difference vs the control group, p < 0.05). E p it h el iu m /C T r at io , ( % ) 0.0 0.5 1.0 1.5 2.0
Tissue area of gingival
T is su e ar ea , ( m m 2 ) 0.0 1.5 3.0 4.5 6.0 Control CsA Total
Sofe tissue Epithelium Connective tissue
Epi/CT ratio
*
*
*
B. Ratio of mRNA fluorescence intensity for genes associated with Wnt/β-catenin pathway by microarray.
Gene name (Unigene) Fold change
Cy5/Cy3 GenBank Accession No.
Rat β-catenin 1.01 NM_053357
Rat E-cadherin 0.95 NM_031334
Rat Dvl-1 1.27 NM_031820
Rat GSK-3β 1.35 NM_032080
Rat APC protein 0.79 AA956063
Rattus Axin 1 0.81 NM_024405
Rat Pcna 1.11 NM_022381
Rat cyclin D1 1.20 NM_171992
Control
C. Transcript and protein expression of genes associated with gingival cell proliferation
mRNA expression, Real time-PCR Immunohistochemistry for Pcna & Cyclin D1
Pcna Cyclin D1 R el at iv e R N A E xp re ss io n (¡Ñ 10 4 ) 0 3 6 9 12 Control CsA
*
*
Control CsA Pcna Cyclin D1Figure 2.
A. mRNA expression, by Real time-PCR B. protein expression, Western Blot
Control CsA
Anti-β-catenin IB
Anti-α-actin IB
C. Immunohistochemistry for β-catenin protein expression
Control
CsA
Figure 3.
A. Transcript and protein expressions of DVL-1
Control CSA Control CsA
DVL-1 GAPDH
B. Transcript and protein expressions in the β-catenin degradation complex
Control CSA Control CsA
GSK-3β GSK-3β APC Axin 1 APC Axin 1 GAPDH
Figure 4.
A. Western Blot of phosphoserine (33/37) & ubiquitination in β-catenin
Anti-phosphoserine (33/37) of β-catenin, Western Blot
Immunohistochemistry for phosphoserine (33/37) of β-catenin
Control CSA Control CSA
Phosphoserine (33/37)
α-actin
B. Anti-β-catenin IP & Anti-ubiquitin IB
Control CSA
Ubi-β-catenin
C. A model of CsA-enhanced β-catenin expression in a gingival cell:
the
inhibition
of β-catenin
degradation via the Wnt/β-catenin pathway
: E-Cadherin : Wnt : Frizzled : GSK-3b : Dvl-1 : APC : Polyubiquitination : TCF : Degradation of b-catenin : phosphoserine 33 : phosphoserine 37 : Increase : decrease
Wnt: Wingless-Int, GSK-3b: Glycogen synthase kinase 3β, APC: Adenomatous polyposis coli, Dvl-1: Disheveled-1, TCF: T-cell factor, CsA: cyclosporine A
Cytosol Axin-1 Nucleus B-catenin B -c ate nin B-catenin B-catenin B-catenin B-catenin Cyclin D1, c-Myc B-catenin T C F B-catenin Cs A Cs A Cs A Cs A P P P P P P