The Association between Polymorphisms of
B7 Molecules (CD80 and CD86) and
Graves’ Ophthalmopathy in a Taiwanese
Population
Wen-Ling Liao, DrPH,
1,2Rong-Hsing Chen, MD,
3Hui-Ju Lin, MD, PhD,
4Yu-Huei Liu, PhD,
1,2Wen-Chi Chen, MD, PhD,
5Yuhsin Tsai, PhD,
2Lei Wan, PhD,
1,2,6Fuu-Jen Tsai, MD, PhD
1,2,6Objective:
This study evaluates whether B7 molecules (CD80 and CD86) could be used as genetic markers
for the development of Graves’ ophthalmopathy (GO).
Design:
Cross-sectional study.
Participants:
We included 471 patients with Graves’ disease (GD; 200 patients with GO and 271 patients
without GO) in a Chinese population in Taiwan.
Methods:
An endocrinologist with substantial experience in thyroid diseases identified GO. Blood samples
were taken for DNA extraction from GD subjects. The gene polymorphism of CD80 and CD86 was genotyped by
polymerase chain reaction in each patient.
Main Outcome Measures:
Genotypes of CD80 and CD86 polymorphism.
Results:
We found that the frequency of C allele at position rs_9831894 of the CD86 gene is different in
patients with GD (with and without GO; chi-square test, P
⫽ 0.0017). In addition, the multifactor
dimen-sionality reduction method was used to identify the best gene– gene interaction to predict the risk of GO. We
identified an interaction between CD80_rs9289131 and CD86_rs9872483 (sign test, P
⫽ 0.0010). Moreover,
the G-A haplotype was shown to have a protective effect in the development of ophthalmopathy among
patients with GD (odds ratio, 0.63; 95% confidence interval, 0.44 – 0.90). Moreover, among patients with GO,
the patients carrying the G-A haplotype had a lower level of free thyroxine T
4than those not carrying the G-A
haplotype (P
⫽ 0.0001).
Conclusions:
These results suggest that the polymorphisms of the CD86 gene may be used as genetic
markers for making the diagnosis and prognosis of GO. Therefore, GO could be a disease with complex genetic
factors, resulting from the existing gene– gene interaction found in the present study.
Financial Disclosure(s):
The authors have no proprietary or commercial interest in any of the materials
discussed in this article. Ophthalmology 2011;118:553–557 © 2011 by the American Academy of Ophthalmology.
Graves’ disease (GD) is a common autoimmune thyroid
disease, in which the body produces antibodies to the
re-ceptor of the thyroid-stimulating hormone. Graves’ disease
is characterized clinically by hyperthyroidism, diffuse
goi-ter, thyroid-specific autoantibodies, Graves’
ophthalmopa-thy (GO), and dermopaophthalmopa-thy.
1Recent studies support the
involvement of environmental triggers and predisposing
genes in the pathogenesis of GD.
2– 4Affecting 25% to 50% of patients with GD, GO is the
most common extrathyroidal manifestation.
4 –7It is
consid-ered to be an autoimmune inflammatory disorder affecting
the extraocular muscles and the orbital fatty and connective
tissues. Recent studies have shown that T-cell–mediated
immunity is likely to play an important role in the
autoim-mune inflammation of ophthalmopathy.
8Several genes,
such as cytotoxic T-lymphocyte antigen-4 (CTLA-4),
9 –11CD28,
12–14and protein tyrosine phosphatase nonreceptor 22
(PTPN22),
15related to T-cell activation have been
investi-gated, but, to date, only the CTLA-4 gene has been shown to
be associated with GD. From a meta-analysis
16that
in-cluded the results of 44 published and unpublished studies,
a significant association between the polymorphism of
CTLA-4 and GD was found in Asian and Caucasian
popu-lations. However, the number of susceptible genes related to
T-cell functions remains unknown.
In order to activate native T cells, a specific antigen that
can be presented by major histocompatibility complex
mol-ecules and antigen–major histocompatibility complex
inter-act with the T-cell receptor are necessary. Moreover, a
second signal that includes the interaction between CD28/
CTLA-4 and B7 molecules is also required for T-cell
acti-vation. B7 molecules are costimulatory molecules
ex-pressed on the surface of antigen-presenting cells. The
binding of B7 to CD28 on T cells initiates a costimulatory
signal for T-cell activation, proliferation, differentiation,
and the subsequent production of a number of cytokines. In
contrast, binding to CTLA-4, which has a higher affinity for
B7, downregulates T-cell activation and diminishes the
im-mune response by competing for the binding site of B7 to
CD28.
17Therefore, B7 molecules—B7.1 (CD80) and B7.2
(CD86)—also play important roles in T-cell activation.
From in vitro and in vivo studies, a role of the
costimu-latory B7 molecules in the regulation of T-cell proliferation
and immune response has been suggested.
18 –20Some
stud-ies have investigated the association of polymorphisms of
B7 molecules with several systemic autoimmune diseases,
including multiple sclerosis,
21rheumatoid arthritis, and
sys-temic lupus erythematosus
22; however, the role of B7
mol-ecules in organ-specific autoimmune diseases remains
un-clear. Therefore, in the present study, we investigated the
association between single nucleotide polymorphisms
(SNPs) in the genes of B7 molecules (CD86 and CD80) and
the susceptibility of developing GO in Taiwanese patients
with GD.
Methods
Patients and Data Collection
A total of 471 patients with GD visiting at China Medical Uni-versity Hospital, Taiwan, were enrolled in this study. All partici-pants were interviewed and examined by an endocrinologist with substantial experience in thyroid diseases. The diagnosis of GD was based on clinical symptoms and biochemical confirmation of hyperthyroidism, diffused goiter, and the presence of ⱖ1 of the following observations: positive results for thyroid-stimulating hormone receptor antibody tests, diffusely increased iodine-131 uptake in the thyroid gland, and exophthalmos. Patients with GD were categorized according to the NOSPECS system recom-mended by the American Thyroid Association.23The GD patients
who had proptosis with or without more severe form (classes 3– 6) were defined as having GO. The degree of proptosis was measured using an exophthalmometer and was defined as a distance of the apex of the cornea from the lateral orbital rim⬎18 mm in either eye or a 2-mm difference in the degree of protrusion between the 2 eyes. The data regarding age, gender, history of tobacco use, thyroid gland pathology, and the affected anatomic site were extracted from full medical records. Blood samples were collected by venipuncture for genomic DNA isolation and serologic tests at the time of enrollment in the study. Informed consent was obtained from each participant before his or her inclusion in this study. The study was approved by the ethics committee of China Medical University hospital.
Genomic DNA Extraction and Genotyping
The genomic DNA was extracted from peripheral blood leuko-cytes using Genomic DNA kit (Qiagen, Valencia, CA) in accor-dance with the manufacturer’s instructions. To select the most representative SNPs by capturing the majority genetic variation, SNP genotype information was downloaded in December 2008 from the HapMap Han Chinese in Beijing ⫹ JPT population. HapMap genotypes were analyzed within Haploview and Tag SNPs were selected using the Tagger function. Eight SNPs for each gene (CD80 and CD86) met the criteria were selected. The following are SNPs we selected for analyses. SNPs belong to CD86 gene: rs129055 (A/G), rs11717893 (A/G), rs2715267 (A/C),
rs2715272 (A/G), rs4678186 (A/G), rs765945 (A/C), rs9831894 (A/C), and rs9872483 (A/G). SNPs belong to CD80 gene: rs9289131 (A/G), rs7642502 (A/G), rs1523311 (A/G), rs2049502 (A/G), rs2228017 (A/G), rs16829957 (A/G), rs3915165 (A/C), and rs16829988 (A/G). Genotyping was performed using an allele-specific extension and ligation assay according to the manufactur-er’s instructions (Illumina, San Diego, CA).
Haplotype Analysis
Haplotype frequencies and effects were examined using the sta-tistical package Haplo.stats24 in software language R (R 2.8.1).
The function Haplo.score was used to assess differences in hap-lotype frequencies between cases and controls and to calculate a global score test that was used to evaluate the overall significance. Effects of individual haplotypes were also examined by comparing the ophthalmopathy risk associated with each inferred haplotype with the risk associated with the highest estimated frequency haplotype. The estimated odds ratio (OR) and 95% confidence intervals (CIs) were obtained using the function Haplo.glm.
Statistical Analyses
The genotype and allele frequency distributions in the polymor-phisms in Graves’ patients with or without ophthalmopathy were analyzed by the chi-square test or Fisher exact test for differences in proportions. The OR was calculated from genotype frequencies and allelic frequencies with 95% CI by using unconditional logis-tical regression adjusting for age of diagnosis, gender and smoking history. The multifactor dimensionality reduction (MDR) method with version 1.1.0 of the open source software package (Dart-mouth Medical School, Hanover, NH) was used to detect the best locus–locus interaction models. The interaction dendrogram was built according to hierarchical clustering algorithm. The effect of carrying high-risk haplotypes on serology test among the GO group was evaluated by the 2 sample independent t-test. All statistical analyses were conducted using SAS statistical software, version 9.1 (SAS Inc., Cary, NC) and P⬍0.05 (2 sided) was used as the level of significance.
Results
We studied 200 GO cases and 271 GO controls, and a total of 16 SNPs were selected from the genes of CD80 (n⫽ 8) and CD86 (n ⫽ 8) to perform genotyping and to investigate the effect of B7 (CD80 and CD86) polymorphisms on GO.
Allele and Genotype Frequencies of CD80 and
CD86 Polymorphisms
We did not find a significant deviation from the Hardy-Weinberg equilibrium for any SNP (P⬎0.05). This finding indicated that the Table 3. Summarizing Multifactor Dimensionality Reduction Models for Gene–Gene Interaction in Graves’ Ophthalmopathy
Risk Number of Factors Best Candidate Models Testing Accuracy (%) P-Value Cross-Validation Consistency 1 CD86_rs9831894 55.6 0.0107 10/10 2 CD80_rs9289131 CD86_rs9872483 57.2 0.0010 8/10 3 CD80_rs16829957 CD86_rs9831894 CD86_rs6765945 49.0 0.3770 4/10
possibility of a bias resulting from population stratification or a genotyping error was unlikely. In genotype association tests, the polymorphism at position rs9831894 (A¡C) in the CD86 gene was statistically associated with GO (P⫽ 0.0071). Furthermore, in allele frequency analyses, the frequency of the C allele at position rs9831894 was significantly higher in patients without GO than in those with GO, with an OR of 0.73 (95% CI, 0.55– 0.97; Table 1, available online athttp://aaojournal.org). However, the distribution of the genotype or the allele frequency of the SNPs within the CD80 gene was not statistically different between the 2 groups (Table 2, available online athttp://aaojournal.org).
Multifactor Dimensionality Reduction Analysis
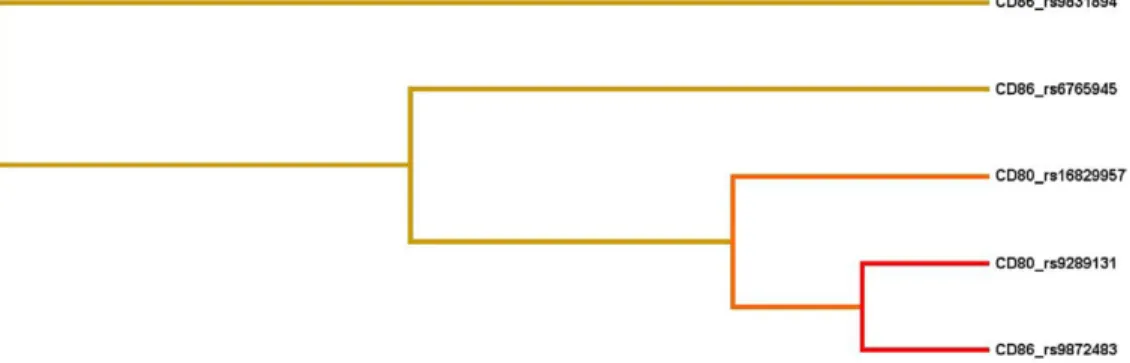
We used MDR analysis to identify the best interaction models among a total of 16 SNPs in the genes of CD80 and CD86.Table 3 summarizes the best interaction models obtained from MDR analysis. Consistent with the individual SNP analysis, in the 1-lo-cus model, the SNP CD86 gene_rs9831894 was the best candidate for predicting a high risk for GO (testing accuracy, 55.6%; cross-validation consistency, 10; sign test, P⫽ 0.0107). The best inter-action model was a 2-locus model composed of the SNPs CD86 gene _rs9872483 and CD80 gene _rs9289131 (testing accuracy, 57.2%; cross-validation consistency, 8; sign test, P ⫽ 0.0010). From the interaction dendrogram (Fig 1), a strong synergistic effect of these 2 loci was found in modeling the risk of GO.
Frequencies of the CD80 and CD86 Haplotype
Analysis of the rs9289131 (CD80 gene) and rs9872483 (CD86 gene) haplotype was used to investigate a potential gene– gene interaction. The overall global test showed a difference in the frequency of haplotypes between the cases and controls forrs9289131-rs9872483 (P ⫽ 0.0246). The haplotype G-A was significantly inversely associated with a high risk for GO (OR, 0.63; 95% CI, 0.44 – 0.90;Table 4). Furthermore, we investigated the effect of G-A haplotypes on clinical serology tests among GO patients. The patients carrying the G-A haplotype had a lower level of free thyroxine (free T4) compared with those not carrying the G-A haplotype (P⫽ 0.0001;Table 5).
Discussion
From previous studies, the importance of immunity-related
genes in autoimmune diseases has been discussed, and the
crucial role of the costimulatory molecules—CD80 and
CD86 —for T-cell activation and inhibition has been
estab-lished. However, the relationship between gene
polymor-phisms and the susceptibility of developing GO remains
unclear. In the present study, we investigated the association
between B7 molecules (CD80 and CD86) polymorphisms
and GO in a Chinese population in Taiwan.
Both CD80 and CD86 genes are located on chromosome
3q21 and have similar costimulatory ligands to activate T
cells. CD86 could be expressed on resting T, B, and natural
killer cells, or monocytes.
25CD86 can be quickly induced
on activated macrophages, B cells, and natural killer cells,
26but not CD80. In addition, the role of costimulation via
CD80 or CD86 in the polarization of the T-helper response
is different. CD80 preferentially acts as a costimulator for
the generation of T-helper 1 cells, whereas CD86
costimu-lation induces the differentiation toward the T-helper 2
functional phenotype.
27Table 4. Haplotype Frequency of CD80_rs9289131 and CD86_rs9872483 among Graves’ Disease Patients
Haplotype Frequency (%) Haplotype-Specific Test P-Value OR (95% CI) Without GO (n⫽ 542) With GO (n⫽ 400) G-A 39 29 0.00747 0.63 (0.44,0.90) A-G 17 16 0.49784 0.80 (0.50, 1.29) G-G 34 39 0.29026 1 A-A 11 17 0.09272 1.32 (0.85, 2.05)
PGlobal Score Testⴝ 0.0246
CI⫽ confidence interval; GO ⫽ Graves’ ophthalmopathy; OR ⫽ odds ratio.
Figure 1. Interaction dendrogram. The interaction dendrogram reveals a strong interaction effect of CD80 and CD86 these 2 genes in modulating the risk of Graves’ ophthalmopathy. The location of the longitudinal connecting bars indicates the strength of the dependence: left is weaker and right is stronger. The hierarchical cluster analysis with average linkage place CD80 rs9289131 and CD86 rs9872483 on the same branch.
To the best of our knowledge, this is the first
population-based study to show that the organ-specific autoimmune
disease GD was associated with SNPs in the CD86 gene.
Moreover, the interaction between CD80 and CD86 genes
was found in the present study. However, an association
between SNPs in the CD80 gene and GO was not found in
the present study, probably because the number of patients
diagnosed with GO might have been too small to detect the
effect of CD80. A larger sample size should be considered
in future studies. From previous studies, CD86
polymor-phisms were found to be associated with asthma and related
allergic disorders.
28,29However, Matsushita et al
22found
that polymorphisms in the genes of CD80, CD86, and
CTLA-4 were not associated with systemic autoimmune
diseases such as rheumatoid arthritis and systemic lupus
erythematosus in a Japanese population.
By using genotype and allele frequencies analyses, we
demonstrated that the C genotype at position rs9831894 of
the CD86 gene may play a protective role in the
develop-ment of ophthalmopathy among GD patients. Furthermore,
we tested the genotype frequency of rs9831894 between GD
patients and a normal population of Han Chinese in Beijing
(data from HapMap database) to investigate its protective
role in the development of GD. The association with GD
was borderline significant (P
⫽ 0.0505). This could be
owing to a lower statistical power in the small Han Chinese
in Beijing population (n
⫽ 83). These results provide
evi-dence of a protective role of CD86 in the development of
ophthalmopathy among GD patients. Furthermore, we
in-vestigated the potential interaction between CD80 and
CD86 by using MDR analysis. Interestingly, in a 2-locus
interaction
model,
CD80_rs9289131-CD86_rs9872483,
was identified as the best model to predict the risk of GO,
although none of these SNPs were identified to be
associ-ated with GO independently. The protective effects of the
G-A haplotype of CD80 –CD86 on symptoms of
ophthal-mopathy was found. Moreover, the higher percentage of GO
patients carrying the G-A haplotype had normal levels of
free T
4compared with those not carrying the G-A haplotype
(54.46% vs 39.47%; P
⫽ 0.0483). This finding suggests that
the B7 genes play a protective role.
In conclusion, this study provides evidence that the
CD86 gene polymorphism is related to GO in GD patients.
Furthermore, a protective role of a gene– gene interaction
between CD80 and CD86 in the development of GO and in
the clinical response was identified.
References
1. Mishra A, Mishra SK. Multicentre study of thyroid nodules in patients with Graves’ disease (Br J Surg 2000;87:1111-13) [letter]. Br J Surg 2001;88:313.
2. Gianoukakis AG, Smith TJ. Recent insights into the patho-genesis and management of thyroid-associated ophthalmopa-thy. Curr Opin Endocrinol Diabetes Obes 2008;15:446 –52. 3. Anvari M, Khalilzadeh O, Esteghamati A, et al. Genetic
susceptibility to Graves’ ophthalmopathy: the role of polymor-phisms in proinflammatory cytokine genes. Eye (Lond) 2010; 24:1058 – 63.
4. Gianoukakis AG, Khadavi N, Smith TJ. Cytokines, Graves’ disease, and thyroid-associated ophthalmopathy. Thyroid 2008;18:953– 8.
5. Perros P, Neoh C, Dickinson J. Thyroid eye disease. BMJ 2009;338:b560.
6. Kuriyan AE, Phipps RP, Feldon SE. The eye and thyroid disease. Curr Opin Ophthalmol 2008;19:499 –506.
7. Khoo TK, Bahn RS. Pathogenesis of Graves’ ophthalmo-pathy: the role of autoantibodies. Thyroid 2007;17:1013– 8. 8. Bednarczuk T, Hiromatsu Y, Inoue Y, et al. T-cell-mediated
immunity in thyroid-associated ophthalmopathy. Thyroid 2002;12:209 –15.
9. Esteghamati A, Khalilzadeh O, Mobarra Z, et al. Association of CTLA-4 gene polymorphism with Graves’ disease and ophthalmopathy in Iranian patients. Eur J Intern Med 2009; 20:424 – 8.
10. Kouki T, Sawai Y, Gardine CA, et al. CTLA-4 gene polymor-phism at position 49 in exon 1 reduces the inhibitory function of CTLA-4 and contributes to the pathogenesis of Graves’ disease. J Immunol 2000;165:6606 –11.
11. Yanagawa T, Hidaka Y, Guimaraes V, et al. CTLA-4 gene polymorphism associated with Graves’ disease in a Caucasian population. J Clin Endocrinol Metab 1995;80:41–5.
12. Bossowski A, Stasiak-Barmuta A, Urban M. Relationship between CTLA-4 and CD28 molecule expression on T lym-phocytes and stimulating and blocking autoantibodies to the TSH-receptor in children with Graves’ disease. Horm Res 2005;64:189 –97.
13. Bossowski A, Stasiak-Barmuta A, Urban M, Rinderle C. Analysis of costimulatory molecules (CD28-CTLA-4/B7) ex-pression on chosen mononuclear cells in adolescents with Graves’ disease during methimazole therapy [in Polish]. En-dokrynol Diabetol Chor Przemiany Materii Wieku Rozw 2004;10:93–101.
14. Tomer Y, Greenberg DA, Barbesino G, et al. CTLA-4 and not CD28 is a susceptibility gene for thyroid autoantibody pro-duction. J Clin Endocrinol Metab 2001;86:1687–93. 15. Ichimura M, Kaku H, Fukutani T, et al. Associations of
protein tyrosine phosphatase nonreceptor 22 (PTPN22) gene polymorphisms with susceptibility to Graves’ disease in a Japanese population. Thyroid 2008;18:625–30.
16. Kavvoura FK, Akamizu T, Awata T, et al. Cytotoxic T-lym-phocyte associated antigen 4 gene polymorphisms and auto-immune thyroid disease: a meta-analysis. J Clin Endocrinol Metab 2007;92:3162–70.
17. Janeway CA, Travers P, Walport M, Schlomchik M. Immunobiology: The Immune System in Health and Disease. 6th ed. New York: Garland Sci.; 2005:328 –9.
18. Paust S, Lu L, McCarty N, Cantor H. Engagement of B7 on effector T cells by regulatory T cells prevents autoimmune disease. Proc Natl Acad Sci U S A 2004;101:10398 – 403. 19. Chang TT, Kuchroo VK, Sharpe AH. Role of the B7-CD28/ Table 5. Effect of Carrying the G-A Haplotype on Clinical
Serology Test in Graves Disease Ophthalmopathy Patients
G-A Haplotype P-Value (t-test) With (n⫽ 101), Mean (SD) Without (n⫽ 76), Mean (SD) Free T 4 1.49 (1.09) 2.21 (1.61) 0.0001 TSH 2.46 (8.38) 1.17 (3.12) ⬎0.05 TRAb* 49.47 (23.47) 54.14 (27.00) ⬎0.05
CTLA-4 pathway in autoimmune disease. Curr Dir Autoimmun 2002;5:113–30.
20. Ziller C, Stoeckel F, Boon L, Haegel-Kronenberger H. Tran-sient blocking of both B7.1 (CD80) and B7.2 (CD86) in addition to CD40-CD40L interaction fully abrogates the im-mune response following systemic injection of adenovirus vector. Gene Ther 2002;9:537– 46.
21. Teutsch SM, Booth DR, Bennetts BH, et al. Association of common T cell activation gene polymorphisms with multiple sclerosis in Australian patients. J Neuroimmunol 2004;148: 218 –30.
22. Matsushita M, Tsuchiya N, Oka T, et al. New polymorphisms of human CD80 and CD86: lack of association with rheuma-toid arthritis and systemic lupus erythematosus. Genes Immun 2000;1:428 –34.
23. Werner SC. Modification of the classification of the eye changes of Graves’ disease: recommendations of the Ad Hoc Committee of the American Thyroid Association. J Clin En-docrinol Metab 1977;44:203– 4.
24. Schaid DJ, Rowland CM, Tines DE, et al. Score tests for association between traits and haplotypes when linkage phase is ambiguous. Am J Hum Genet 2002;70:425–34.
25. Azuma M, Ito D, Yagita H, et al. B70 antigen is a second ligand for CTLA-4 and CD28. Nature 1993;366:76 –9. 26. Hathcock KS, Laszlo G, Pucillo C, et al. Comparative analysis
of B7-1 and B7-2 costimulatory ligands: expression and func-tion. J Exp Med 1994;180:631– 40.
27. Battifora M, Pesce G, Paolieri F, et al. B7.1 costimulatory mol-ecule is expressed on thyroid follicular cells in Hashimoto’s thyroiditis, but not in Graves’ disease. J Clin Endocrinol Metab 1998;83:4130 –9.
28. Chen YQ, Shi HZ. CD28/CTLA-4 —CD80/CD86 and ICOS—B7RP-1 costimulatory pathway in bronchial asthma. Allergy 2006;61:15–26.
29. Corydon TJ, Haagerup A, Jensen TG, et al. A functional CD86 polymorphism associated with asthma and related allergic disorders. J Med Genet 2007;44:509 –15.
Footnotes and Financial Disclosures
Originally received: February 9, 2010. Final revision: July 22, 2010. Accepted: July 23, 2010.
Available online: September 29, 2010. Manuscript no. 2010-214.
1Genetic Center, China Medical University Hospital, Taichung, Taiwan. 2School of Chinese Medicine, China Medical University, Taichung,
Taiwan.
3School of Post Baccalaureate Chinese Medicine; China Medical
Univer-sity, Taichung, Taiwan.
4Department of ophthalmology, China Medical University Hospital,
Tai-chung, Taiwan.
5Graduate Institute of Integrated Medicine; China Medical University,
Taichung, Taiwan.
6Department of Health and Nutrition Biotechnology, Asia University,
Taichung, Taiwan. Financial Disclosure(s):
The authors have no proprietary or commercial interest in any of the materials discussed in this article.
Supported by a grant from the National Science Council (98-2320-B-039-008-MY3), Taipei, Taiwan, and a grant from the China Medical University Hospital (DMR-93-45), Taichung, Taiwan.
L.W. and F-J.T. contributed equally to this work.
Correspondence: Fuu-Jen Tsai, MD, PhD, Genetic Center, China Medical University Hospital, No.2 Yuh-Der Road, 404 Taichung, Taiwan. E-mail: