中國醫藥大學機構典藏 China Medical University Repository, Taiwan:Item 310903500/41442
236
0
0
全文
(2)
(3) Acknowledgement 『轉眼之間,時光飛逝』也許不能夠形容我畢業所經歷的時間,我想用『長期抗 戰,革命成功』此類的話語比較能闡述我現在的心情。畢竟,中間經歷的酸甜苦辣可 能是無法言喻的。不過,現在我想說的是,我終於走過來啦! 在這一段漫長的日子裡,我應該感謝的人不勝其數。在這段學業和人生路途上, 我想最該感謝的應該是我的恩師楊寧蓀老師、郭盛助老師和吳金濱老師。首先。非常 感謝楊寧蓀老師,從我在中研院農生中心服國防役至今,謝謝您收留我這個國防役助 理到收留一個中國醫藥大學來的博士班學生的恩情,更感謝您從不間斷地用的熱情將 您的卓越研究經驗及看法和獨特人生經歷分享給我,由衷地感謝您的提拔和教誨,很 高興能在您的實驗室與您一同成長和學到與眾不同的科學辦法和思考方式,我會好好 地學習您的辯證法、多快好省和多問等科學方法, 『寧蓀,謝謝您!』 。此外,也非常 感謝郭盛助老師從我大學就上藥物化學課程時的精闢教導,那時就感覺到您是一位很 棒的老師;碩士班時共同指導碩士論文和給予指導,受益匪淺;後來,讀博士班就學 後,給予莫大的支持,對論文和投稿上總不吝嗇給予指導和幫忙,發自內心地很高興 有您這一位恩師, 『郭老師,謝謝您!』 。我還需要感謝我的一位恩師吳金濱老師,謝 謝您在大學時,就短暫收留我這個大學生在實驗室學習,後來碩士班也承蒙您的照 顧,見到許多不同的世面,學習到許多不同的科學方法和工業化的製程,最後得以順 利地畢業;也很感謝您在博士班的照顧和幫忙,『吳老師,謝謝您!』。 在這邊也非常感謝對此篇論文給予許多指導的老師,感謝莊聲宏所長、鍾景光老 師、王升陽老師、高銘欽老師和吳天賞院長,在口試時給予諸多寶貴的意見和著實受 用的指正,俾使這份論文能夠更加完整、豐富、有條理和有邏輯。這中間特別感謝莊 聲宏所長針對這份論文花了許多的心思給予更正,也非常感謝鍾景光老師對文章邏輯 寫作和投稿上的幫忙;更感謝 SARS 計畫時大力幫忙的王升陽老師,時常在人生的大 道上,給予許多的灌溉。另外,我也想感謝在中研院農生中心幫助我的老師們,謝謝 徐麗芬老師在 SARS 撰寫時,給予許多的指導和幫忙;謝謝蕭培文老師在許多實驗上 給予許多邏輯和設計上的指導;謝謝楊文欽不厭其煩地提供免疫學上的知識和在實驗 資源上的幫助;謝謝梁佑全老師、林崇熙老師和侯珈楨博士,實驗上和生活上的鼎力 相助,也感謝生醫所潘文涵老師和施嘉和老師在計畫合作時的指導和幫忙;此外,亦 非常感激中研院生化所饒淑娟博士在實驗上的大力幫忙和指導。還有其他機構也給予 我許多的指導,特別感謝陳穗生教授給我許多科學之事的灌輸和教導,也提供這篇論 文相當的幫忙和校正;感謝台北醫學大學李慶國老師和林天仁老師,中國醫藥大學黃 麗嬌老師、郭悅雄老師、連金城老師和邱紹智老師,高雄醫學大學張芳榮所長,中山.
(4) 大學廖志中老師在學習過程中的指正和生活上的打氣幫忙。我很想再感謝的就是我的 良師益友中興大學簡世昌教授,感謝他在婚禮的幫忙和博士班歷程中,不斷地提供精 神和生活上的灌溉,也提供了許多科學上的指導,非常感謝。 攻讀博士的過程中,太多的人需要感謝。首先,想要感謝在中研院楊寧蓀實驗室 裡曾經幫忙或一起工作過的同事們,謝謝我的難兄難弟魏紋祈魏傑克,很高興看到你 的卓越成長和感謝你給我的幫忙;謝謝繪名鞋姊在實驗和生活上的幫忙,謝謝現任同 事們碧雪、莉婷、永祥、薇婷、Arul、Tina、Blue、小倩、聖晏、詠宗、怡君、書翊、 佳奇、小昭和年輕有為的章杰,感謝楊寧蓀實驗室的前同事們曹小曹、徐師傅、琇慧、 文經、Terry、小美、瑩筑、雅玲、文心學姊、小米茶、淑貞、王正方博士、Kandan、、 Vani、Steve、顏伯任(Robert)、趙嶸、坤珀、育生、青晉、靜芳、淑敏;也感謝中研 院其他實驗室所有曾經幫過我的人,謝謝聿智、永志、阿伯、洪博、慶豐、俊維、佳 穎、逸人、恩豪、進賢、秋萍、育雯、瑀心、世川、怡均、清毅、郭博、秉忠、小佑、 恬岎、志翰、小賴、奕枚、包子、宗遠、政權、慧玲、啟彰、智揚、俊杰、皎君、書 甄、秉智、凱偉、子軒、均宇、偉君,謝謝你們的陪伴和幫忙。另外,我也想特別感 謝中國醫藥大學曾經幫助過我的貴人謝謝立琛常常不辭辛勞地幫忙,真心地非常感謝 你的熱心幫助,謝謝學長姐們俊仁美華夫婦、勝宏學長、健源學長,謝謝同學們 momo、華鑫、元照、亮董和怡倩的幫忙,謝謝學弟妹們泰霖、士鳴,也特別感謝廖 紋瑩蟲蟲時常的鼎力相助。此外,我想感謝我的死黨,恭喜也謝謝黎雁行博士,謝謝 大熊、芭樂。還有一直情義相挺的 Kenny 和老大,謝謝你們在我需要幫忙時,幫助 我! 最後,我想將這份論文獻給我最重要的親人們!首先,是我最親愛的父母親,爸 媽,您們是很偉大也是全世界最好的爸媽,很謝謝您們這麼地相信和栽培著我,謝謝 你們,沒有你們,就沒有這份論文,謝謝爸爸和媽媽!再來,我想要將這份論文獻給 我在天國的叔叔(乾爸),謝謝您一直鼓勵我,我真的很希望您可以看到我畢業的那 一刻,謝謝 K 爸,我也想謝謝我的乾媽,謝謝乾媽一路的支持和鼓勵;我也想獻給 我在天上的爺爺和奶奶,希望您們能夠感到高興。再者,我想要謝謝我的大哥致遠, 謝謝你一直都支持你的弟弟,我感受的到,哥,謝謝你!也謝謝我的兩個妹妹致儀和 致絹,相信你們也會為我高興,謝謝你們!還要謝謝我的另一爸媽我的岳父岳母,不 但不嫌棄我,更願意將他們最寶貴的掌上明珠許配給我,也僅以這份論文獻給您們! 在這最後的最後,壓軸中的壓軸,我想要以這份論文獻給剛成為我最親密的家人岳 穎,謝謝你當初在眾多競爭者中選擇了我一個窮學生,謝謝你在我學習歷程中的支 持,謝謝你走進我的生命中,更謝謝你給我機會在民國一百年完成了與妳的終身大事 和學業,謝謝你成為我生命中的一部份,沒有你就沒有這份論文,謝謝我最棒的老婆!.
(5) Table of Contents Page Table of Contents……………………………………………………………………............I Abbreviation…………………………………………………………………………….....VI Figures…………. …………………………………………………………………............XI Tables…………. ………………………………………………………………….......... XV Chinese Abstract………………………………………………………………..............XVII Abstract………. ……………………………………………………………….............XXII. Part I. Novel quinolone CMQ induces apoptosis and mitotic catastrophe in prostate cancer cells via reactive oxygen species- and mitochondria-dependent pathways………………...1 Chapter 1. Introduction ……………………………………………………………….…….1 1.1. Epidemiology of prostate cancer……………………………………...……….....1 1.2. The structure of microtubules and microtubule-binding agents (MBAs)………..5 1.3. Cell death including apoptosis and mitotic catastrophe…………………..……14 1.4. Relationship between p53 and microtubule-binding agents……………………21 1.5. CMQ as one of 2-phenyl-4-qunolone derivatives with potent anti-tumor activity………………………………………………………………………….24 Chapter 2. Rationale and Significance…….. …………………………………………….28. I .
(6) Chapter 3. Results and Discussion…………………………………………………….…..31 3.1. Results. ………………………………………………………………….........…31 3.1.1. CMQ inhibits growth of both LNCaP and PC-3 cells………………...…31 3.1.2. CMQ induces G2/M cell cycle arrest that leads to apoptosis in LNCaP cells and endoreduplication in PC-3 cells………………………………..34 3.1.3. CMQ suppresses polymerization of microtubules in LNCaP and PC-3 cells………………………………………………………………………41 3.1.4. CMQ activates the intrinsic apoptotic pathway in both cell types, but activates the extrinsic pathway only in LNCaP cells. …………………...45 3.1.5. CMQ can confer caspase-dependent and ROS-mediated antitumor activity…...…………………………………………………………….....50 3.1.6. CMQ suppresses tumor growth in xenograft mouse models………….…52 3.2. Discussion…………………………………………………………………..…...56 Chapter 4. Conclusion…………………………………………………………………......64 Chapter 5. Materials and Methods…………………………………………….……...…...65. Part II. CMQ augments efficacy of dendritic cell-based cancer vaccines…………………71 Chapter 1. Introduction………………………………………………………………….…71 1.1. Cancer vaccines and dendritic cells……………………………………….....….71 II .
(7) 1.2. Damage-associated molecular patterns (DMAPs) and Immunogenic cell death (ICD)……………………………………………………………………………81 1.3. The relationship between microtubule-targeting/binding agents (MTAs/MBAs) and immunity……………………………………………………...……………87 Chapter 2. Rationale and Significance………………………………………………..…...88 Chapter 3. Results and Discussion…………………………………………………….......90 3.1. Results……………………………………………………………………..…….90 3.1.1. Effect of various microtubule-depolymerizing agents on growth of B16F10 melanoma cells…………………….....................................…....90 3.1.2. Effect of specific microtubule-depolymerizing agents on expression of immunogenic cell death-related proteins and tumor-associated antigens..93 3.1.3. Microtubule-depolymerizing agents can enhance the efficacy of therapeutic immunity provided by tumor cell lysate-pulsed dendritic cell vaccines……………………….………………………………………….96 3.1.4. Specific immune cell subsets involved in vaccine efficacy…………….100 3.1.5. Effect of DC culture conditions on vaccine efficacy…………….......…102 3.1.6. Effect of different administration routes on therapeutic immunity of DC-based vaccines pulsed with tumor cell lysates……………………..106 3.1.7. Microtubule-depolymerizing agents enhance maturation of dendritic cells III .
(8) and CD4+ T and CD8+ T cell proliferation……………………………..109 3.2. Discussion……………………………………………………………………...112 Chapter 4. Conclusion…………………………………………………………………....118 Chapter 5. Materials and Methods…………………………………………………..…...120. Part III. Specific plant terpenoids and lignoids possess potent antiviral activities against severe acute respiratory syndrome coronavirus……………………………………….....130 Chapter 1. Introduction…………………………………………………………………..130 1.1. Severe acute respiratory syndrome (SARS) and treatment of SARS……………………………………………………………………………...….130 1.2. Key steps in SARS-CoV replication as potential targets for anti-SARS-CoV drugs………………………………………………………………………………….133 1.3. SARS and anti-SARS drug development…………………………………...…135 Chapter 2. Rationale and Significance……………………………………………….......136 Chapter 3. Results and Discussion……………………………………………………….137 3.1. Results……………………………………………………………………..…...137 3.1.1. Anti-SARS-CoV activity of test phytocompounds and extracts as measured by cell-based cytopathogenic effect (CPE) assay……………137 3.1.2. Inhibition of SARS-CoV replication evaluated using ELISA………….144 IV .
(9) 3.1.3. Cytotoxic effects of test compounds on Vero E6 cells………...……….153 3.1.4. Inhibitory effects of test phytocompounds or extracts on SARS-CoV 3CL protease activity…………………………………………………………155 3.1.5. Structural modeling of compounds 13-15 with SARS-CoV 3CL protease…………………………………………………………………160 3.2. Discussion………………………………………………………………...……164 Chapter 4. Conclusion…………………………………………………………………....174 Chapter 5. Materials and Methods…………………………………………………....….175. References…………...………………………………………………………………..….184. V .
(10) Abbreviation 2P4Q. 2-phenyl-4-quinolone. ABC. ATP binding cassette. AIF. Apoptosis-inducing factor. ALL. acute lymphoblastic leukemias. APC. antigen-presenting cell. ATCC. American Type Culture Collection. BMDC. bone marrow-derived dendritic cells. BrdU. 5-bromo-2-deoxyuridine. CBE. 75% ethanol extract from Cibotium barometz. CBM. methanol extract from Cibotium barometz. CEA. carcinoembryogenic antigen. CICD. caspase-independent cell death. CC50. the cytotoxic concentration (CC50) of test compounds that. CCR7. CC chemokine receptor 7. CMQ. 2- (3-chlorophenyl)- 6, 7-methylenedioxyquinolin-4-one. concn. concentration. COX-2. cyclooxygenase-2. CPE. cytopathogenic effect. CRT. calreticulin. CTCL-CMQ. Dendritic cells were pulsed with CMQ-treated tumor cell lysate. CTL. cytotoxic T lymphocyte. CTH. n-hexane extract from Cassia tora. DAMPs. damage-Associated Molecular Patterns VI . .
(11) DAPI. 4',6-diamidino-2-phenylindole. DBM. methanol extract from Dioscorea batatas. DCs. dendritic cells. DMSO. dimethyl sulfoxide. DR. death receptor. DSB. double strand breaks. DTT. dithiothreitol. EC50. the effective concentration (EC50) for the inhibition of viral. ELISA. Enzyme-linked immunosorbent assay. END. endoreduplication. ER. endoplasmic reticulum. Eq. 1. Equation 1. Eq. 2. Equation 2. F-T. 4-6 cycles of freezing-thawing. FBS. fetal bovine serum. FITC. fluorescein isothiocyanate. FMKp. a bacterial membrane fragment of Klebsiella pneumonia. FMQ. 2-(3-fluorophenyl)-6,7-methylenedioxyquinolin-4-one. FRET. fluorescence energy transfer. GM-CSF. granulocyte macrophage colony-stimulating factor. GSH. n-hexane extract from Gentiana scabra. H2DCFDA. dichlorodihydrofluorescein diacetate. HMGB1. high-mobility group box 1. HSP70. heat shock protein 70 VII . .
(12) HSP90. heat shock protein 90. IAP. inhibitor of apoptosis. ICD. immunogenic cell death. I.D.. intradernally. IL-1β. interleukin-1β. IL-4. interleukin-4. IL-6. interleukin-6. IL-10. interleukin-10. IL-12. interleukin-12. iNOS. inducible nitric oxide synthase. I.N. intranodally. Irra. irradiation (60 Gray). I.T.. intratumorally. I.V.. intravenously. DX. doxorubicin. KLH. keyhole limpet hemocyanin. mAb. monoclonal antibodies. maDC. matured DC. MBAs. microtubule-binding agents. MC. mitotic catastrophe. MDAs. microtubule-depolymerizing agents. MDDC,. monocyte-derived dendritic cells. MDR. multidrug resistance. MFI. mean fluorescence intensity VIII . .
(13) MHC. major histocompatibility complex. MRP. multidrug resistance-associated protein. MTAs. microtubule-targeting agents. MTT. 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide. NAC. N-acetyl cysteine. N.C.. not calculable. NCCD. nomenclature committee on cell death. NF-κB. nuclear factor kappa B. NIC. niclosamide. NMDA. N-nethyl D-aspartate. NSCLC. non-small-cell lung carcinomas. N.T.. not tested. OD. Optical density. PBMC. peripheral blood mononuclear cells. PBST. PBS containing 0.1% Tween 20. PCD. Programmed cell death. PDB. Protein Data Bank. PE. phycoerythrin. PI. propidium iodide. PMSF. phenylmethylsulfonnylfluoride. PSMA. prostate-specific membrane antigen. PTX. paclitaxel. S.C.. subcutaneously. SCID. severe combined immunodeficient IX . .
(14) S.D.. standard deviation. SI. Selective index. ROS. reactive oxygen species. SARS. severe acute respiratory syndrome. SARS -CoV. SARS coronavirus. TAAs. tumor-associated antigens. TBAs. tubulin-binding asgents. TCH. n-hexane extract from Taxillus chinensis. TCLs. tumor cell lysates. TCL-C. DCs pulsed with colchicne-treated tumor cell lysate for 12h,. TCL-CMQ. DCs pulsed with CMQ-treated tumor cell lysate for 12h,. TCL-DMSO. DCs pulsed with DMSO-treated tumor cell lysate for 12h,. TCL-DX. DCs pulsed with DX-treated tumor cell lysate for 12h,. TCL-FMQ. DCs pulsed with FMQ-treated tumor cell lysate for 12h,. TCM. traditional Chinese medicines. TNF-α. tumor necrosis factor-α. TReg. regulatory T cells. V. vincristine. VAL. valinomycin. Z-IETD-FMK. caspase-8 inhibitor. Z-VAD-FMK. caspase family inhibitor. X .
(15) Figures Page Figure 1. Ten leading cancer types for the estimated new cancer cases and deaths, by sex, United States, 2010………………………………………………………………….……....3 Figure 2. Various chemical structures of microtubule-binding agents, grouped according to their binding domains……………………………………………………….…….………...7 Figure 3. The formation of microtubules and the binding sites for microtubule inhibitors. …………………………………….……………………………………..............8 Figure 4. The role of the microtubules in cell cycle………………………………..….......10 Figure 5. Schematic representation of detailed mechanisms that govern the extrinsic and intrinsic apoptotic pathways and caspase-independent cell death…………………..…......17 Figure 6. Schematic representation of current concepts of mitotic catastrophe…………...18 Figure 7. Pathways leading to mitotic catastrophe and cell death after DNA damage……20 Figure 8. Schematic representation of putative mechanisms of a cell after exposure to low and high concentrations of the microtubule-depolymerizing agents, nocodazole……...…22 Figure 9. The relationship between mitotic defects with numeral abnormalities of spindles and p53 …………………………………….……………………………………...............23 Figure 10. Chemiceal structure and cytotoxicty of CMQ in human prostate carcinoma and other cells…..……………………………....……………………………………...............33. XI .
(16) Figure 11. CMQ induces apoptosis and mitotic catastrophe via G2/M cell cycle arrest.....37 Figure 12. CMQ induces DNA damage and fragmentation……………………….............39 Figure 13. CMQ suppresses polymerization of microtubules in LNCaP and PC-3 cells....43 Figure 14. CMQ triggers the ROS-mediated intrinsic apoptotic pathway in both LNCaP and PC-3 cells, whereas it activates the extrinsic apoptotic pathway only in LNCaP cells………………………………………………………………………………………..47 Figure 15. ROS plays a role in antitumor activity of CMQ that is mediated by ROS…....51 Figure 16. CMQ suppressed PC-3 and LNCaP tumor growth in subcutaneous xenograft SCID mice………………………………………………………………………………...54 Figure 17. Hypothetical mechanisms for the action mode of CMQ-induced cell cycle arrest and cytotoxicity in LNCaP cells and PC-3 cells…………………………………………..63 Figure 18. The different functions of immature and mature dendritic cells………………75 Figure 19. The approach of ex vivo differentiation and activation of DCs for cancer immunotherapy……………………………………………………………………………77 Figure 20. Various factors to be considered in DC-based immunotherapy……………….77 Figure 21. Critical events required to trigger dendritic cell activation by dying tumor cells………………………………………………………………………………………..83 Figure 22. The key steps for the induction of an antitumor immune response……………85 Figure 23. Schematic representation of the main immunogenic determinants of dying XII .
(17) tumor cells…………………………………………………………………………………86 Figure 24. Chemical structures of microtubule-depolymerizing agents and doxorubicin tested in this study…………………………………………………………………………91 Figure 25. Effect of MDAs and doxorubicin on cell viability of B16F10 melanoma cells.92 Figure 26. Expression of damage-associated molecular patterns (DAMPs) and tumor-associated antigens in tumor cell lysates of B16F10 melanoma treated with different MDAs…………………………………………………………………………………….94 Figure 27. Therapeutic immunity of DC vaccines pulsed with various tumor cell lysates, against B16 melanoma……………………………………………………………………98 Figure 28. Determination of immune cell subsets responsible for protective immunity induced by test DC vaccines……………………………………………………………..101 Figure 29. Effect of GM-CSF and IL-4 on IL-12p70 expression in DCs treated with tumor cell lysates………………………………………………………………………….…….104 Figure 30. Effect of different administrative routes on DC-based vaccines pulsed with specific tumor cell lysates………………………………………………………………..108 Figure 31. Effect of treatment with different MDAs on expression of cell-surface markers in mouse bone marrow-derived dendritic cells and CD4+ and CD8+ T-cell proliferation.110 Figure 32. Schematic diagram of the SARS coronavirus structure………………………131 Figure 33. A summary of the agents used clinically and studied experimentally regarding XIII .
(18) the pharmacological treatment and prevention of SARS………………………………...132 Figure 34. Different drug targets in the life cycle of SARS-CoV………………………..134 Figure 35. Characterization of compound inhibition of cytopathogenic effect (CPE) of SARS-CoV on Vero E6 cells using a cell-based assay…………………………………...139 Figure 36. Chemical structures of the 22 compounds that exhibit significant inhibitory activity against cytopathogenic effect of SARS-CoV on Vero E6 cells………………….140 Figure 37. The inhibitory effects of test compounds on replication of SARS-CoV and on proliferation of Vero E6 cells…………………………………………………………….146 Figure 38. Inhibitory effect of test extracts on replication of SARS-CoV and on proliferation of Vero E6 cells…………………………………………………………….148 Figure 39. The principle of the SARS-CoV 3CL protease inhibition assay using fluorogenic substrate……………………………………………………………………..156 Figure 40. The inhibitory properties of betulinic acid (13) and savinin (16) on the enzymatic activity of SARS-CoV 3CL protease…………………………………………158 Figure 41. Inhibition of the enzymatic activity of SARS-CoV 3CL protease by CBM and DBM……………………………………………………………………………………..159 Figure 42. Structural modeling of the binding of compounds 13-16 to SARS-CoV 3CL protease…………………………………………………………………………………..162 Figure 43. Schematic representation of different preparations of test herbal extracts…..177 XIV .
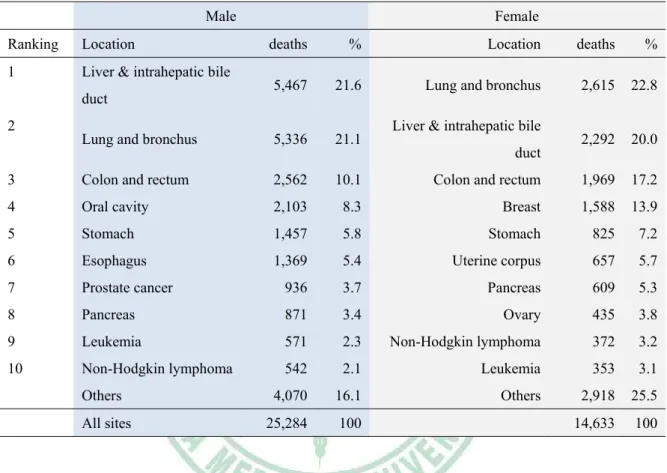
(19) Tables Page Table 1. Ten leading cancer types for the estimated cancer–caused deaths, by sex, Taiwan, 2009…………………….…………………………………………………………………...4 Table 2. Anti-mitotic drugs including microtubule-polymerizing agents and microtubule-depolymerizing agents in clinical use……...……………………...…………12 Table 3. Characteristics of microtubule-binding agents. …………………….…................13 Table 4. Comparison of major cell death pathways observed in drug-treated cells……….18 Table 5. Specific inducers of mitotic catastrophe (MC) and associated events…………...19 Table 6. A summarized list of bioactivities of quinolone derivatives and 2-phenyl-4quinolone derivatives……………………………………………………………………...25 Table 7. Overview of clinical trials using dendritic cell-based cancer immunotherapy…..78 Table 8. Major determinants of immunogenic cancer cell death and their effects on DCs.84 Table 9. Phytocompounds tested as effective against CPE of SARS-CoV on Vero E6 cells.....................................................................................................................................142 Table 10. Effect of Traditional Chinese Medicine extracts on cytopathogenic effect (CPE) of SARS-CoV on Vero E6 cells…………………………………………………………..143 Table 11. Inhibition of Vero E6 cell proliferation and SARS-CoV replication by test phytocompounds…………………………………………………………………………150 XV .
(20) Table 12. Effect of test extracts on Vero E6 cell proliferation and SARS-CoV replication………………………………………………………………………………...152 Table 13. Kinetic properties of specific compounds that inhibit the enzymatic activity of SARS-CoV 3CL protease………………………………………………………………...157 Table 14. IC50 values of test extracts on the enzymatic activities of SARS-CoV 3CL protease…………………………………………………………………………………..159. XVI .
(21) 中文摘要 第一部分: 在第一個研究部分,我們利用 CMQ 作為候選藥物探討在 p53 基因表 達的前列腺癌 LNCaP 細胞和 p53 基因缺乏的前列腺癌 PC-3 細胞的抗癌 活性。體外結果顯示 CMQ 可抑制這兩種癌細胞的增生,這種抑制效果與 微管解聚效果和 G2/M 細胞週期中止有關。CMQ 在 LNCaP 細胞引起嚴 重 的 凋 亡 (apoptosis) 效 果 , 但 是 , 在 PC-3 細 胞 則 引 起 核 內 複 製 (endoreduplication)和延後性的細胞死亡。細胞週期的封鎖被發現與升高 的活性氧(reactive oxygen species, ROS)的升高量有關,進而啟動了粒腺體 凋亡路徑及後續的 caspase-3 和 caspase-9 的活化。此外,CMQ 可以活化 LNCaP 細胞中的 Caspase-8,在 PC-3 細胞則不然。腹腔內注射 CMQ 有 效性抑制在帶有 LNCaP 和 PC-3 之免疫不全小鼠之癌細胞生長。這些實 驗結果顯示 CMQ 在這兩種前列腺癌細胞具有可區分的抗癌效果。. 關鍵詞:CMQ;2-phenyl-4-quinolone;細胞凋亡;有絲分裂災難;抗癌 活性 XVII .
(22) 第二部分: 損害相關分子模式(damage-Associated Molecular Patterns, DAMPs)和 免疫性的細胞死亡(immunogenic cell death, ICD)相關,而且能促進樹突狀 細 胞 (dendritic cells, DCs) 的 成 熟 和 抗 原 呈 現 。 特 定 的 微 管 解 聚 劑 (microtubule-depolymerizing agents, MDAs)例如秋水仙素已經被發現能抗 癌和活化樹突狀細胞。在這個部分的研究,我們評估三種了 MDAs,包 括秋水仙素和兩種 2-phenyl-4-quinolone (2P4Q)衍生物,誘發在癌細胞免 疫性細胞死亡的能力及活化樹突狀細胞和增加 T 細胞的能力。這三個測 試的 MDAs 可以使 HSP70 (heat shock protein 70)、HSP90 (heat shock protein 90)和 HMGB1 (high-mobility group box 1)的 DAMPs 增加,但卻不 能增加 calreticulin(CRT)的表現量。樹突狀細胞加上經由 MDAs 處理過 內的腫瘤細胞裂解液 (tumor cell lysates, TCLs)為基礎的樹突狀細胞疫苗 有效地抑制癌細胞生長和增加小鼠存活率,並且對癌細胞具有細胞毒性 T 淋巴球(cytotoxic T lymphocyte, CTL)的活性。在體內利用抗體刪除的實驗 中,顯示 CD8+ T 細胞和自然殺手細胞才是主要影響此項抗癌活性的主要 細胞,而非 CD4+ T 細胞。此外,在處理和刺激樹突狀細胞的過程中,加 入 白 介 素 -4 (interleukin-4, IL-4) 和 顆 粒 細 胞 巨 噬 細 胞 群 落 刺 激 因 子 (granulocyte macrophage colony-stimulating factor, GM-CSF)可以有效地增 加白介素-12 (interleukin-12, IL-12)和降低白介素-10 (interleukin-10, IL-10) XVIII .
(23) 的分泌量。在另一方面,MDAs 也可以引起樹突狀細胞表面標記成熟表 現,和增加 CD4+ T 細胞和 CD8+ T 細胞與樹突狀細胞(DCs)之增生。綜觀 來說,MDAs 包括臨床使用的秋水仙素可以引起免疫性細胞死亡,MDA 處理過內的腫瘤裂解液(tumor cell lysates, TCLs)為基礎的樹突狀細胞疫 苗可以產生強效的抗癌免疫活性。這個研究方法也許可以在未來臨床上 癌症疫苗發展的指標。. 關鍵詞:免疫性的細胞死亡;2-phenyl-4-quinolone;樹突細胞;秋水仙素; 癌症疫苗 XIX .
(24) 第三部分: 為避免抗嚴重急性呼吸道症候群(severe acute respiratory syndrome, SARS)這種致命疾病的捲土重來,發展抗嚴重急性呼吸道症候群冠狀病毒 (SARS coronavirus,. SARS-CoV)藥物是重要的。在這個研究中,利用. SARS-CoV 感染 Vero E6 細胞所產生的細胞病變效應(cytopathogenic effect,CPE)為測定法來評估超過兩百個植化物和超過兩百個中草藥萃取 物之抗 SARS-CoV 活性。植化物中,十個雙萜類化合物、兩個倍半萜類 化合物、兩個三萜類化合物、五個木酚素類化合物、薑黃素和兩個參考 有效控制組 niclosamide 和 valinomycin 在濃度 3.3-10 μM 皆為 CPE 抑制 劑. 六種植化萃取物在濃度 25-200 μg/ml 也為強的抑制劑,分別為從龍膽 (Gentiana scabra)萃取的 GSH、從淮山藥(Dioscorea batatas)萃取的 DBM、從決明子(Cassia tora)萃取的 CTH、從桑寄生(Taxillus chinensis) 萃取的 TCH 和從狗脊(Cibotium barometz)萃取的 CBE 和 CBM。並針 對二十二個化合物和六個植物萃取物進行抑制 50 %Vero E6 細胞增殖的 樣品濃度(CC50)和抑制 50 %病毒繁殖的樣品濃度(EC50)之測定。最有效的 植化物 1、5、6、8、14 和 16 其選擇性指數值(Selective index values, SI, SI= CC50/ EC50)分別為 58、>510、111、193、180 和>667。CBE、 GSH、 DBM、 CTH 和 TCH 這五種最有效萃取物其 SI 分別為> 59.4、> 57.5、> 62.1、> 59.4 和 > 92.9。 Betulinic acid (13) 和 savinin (16) 是 SARS-CoV 3CL 蛋 XX .
(25) 白質酶競爭性抑制劑,其 Ki 值 分別為 8.2 ± 0.7 M 和 9.1 ± 2.4 M。 CBM 和 DBM 也顯示能制 SARS-CoV 3CL 蛋白質酶活性,其 IC50 分別為 39 μg/ml 和 44 μg/ml。我們的研究顯示特定的雙萜類化合物、木酚素類化 合物和這六種化合物具有強效的抗 SARS 效果。. 關鍵詞:嚴重急性呼吸道症候群;萜類;木酚素類;植物萃取物;3CL 蛋白質酶. XXI .
(26) Abstract Part I. In. this. study,. we. investigated. the. effect. of. 2-. (3-chlorophenyl)-. 6,. 7-methylenedioxyquinolin-4-one (CMQ) as a drug candidate for anti-tumor activities in p53-expressing LNCaP cells and p53-null PC-3 cells of prostate cancers. In vitro results showed that CMQ-1 inhibited tumor cell growth in both cell types, and the inhibition was associated with microtubule-depolymerizing activity and G2/M cell cycle arrest in both cell lines. Intriguingly, CMQ triggered a strong apoptotic activity in LNCaP cells but induced an endoreduplication activity and delayed cell death in the tested PC-3 cells. The cell cycle blockade was found to be associated with an elevated level of reactive oxygen species (ROS), followed by activation of the mitochondrial apoptotic pathway, with subsequent consecutive activation of caspases-9 and -3. In addition, CMQ significantly activated caspase-8 in LNCaP cells but not in PC-3 cells. Intraperitoneal injection of CMQ significantly suppressed tumor growth in SCID mice bearing LNCaP or PC-3 xenografts. Our findings suggest that CMQ may display differential antitumor activities in different prostate cancers, and this may have clinical applications.. Keywords: CMQ, 2-phenyl-4-quinolone, apoptosis, mitotic catastrophe, anti-tumor activity XXII .
(27) Part II. Damage-associated molecular patterns (DAMPs) are associated with immunogenic cell death and have the ability to enhance maturation and antigen presentation of dendritic cells (DCs). Specific microtubule-depolymerizing agents (MDAs) such as colchicine have been shown to confer anti-cancer activity and also trigger activation of DCs. In the second section, we evaluated the ability of three MDAs (colchicine and two 2-phenl-4-quinolone analogues) to induce immunogenic cell death in test tumor cells, activate DCs, and augment T-cell proliferation activity. Three test phytochemicals considerably increased the expression of DAMPs including HSP70, HSP90 and HMGB1, but had no effect on expression of calreticulin (CRT). DC vaccines pulsed with MDA-treated tumor cell lysate (TCLs) had a significantly inhibitory effect on tumor growth, showed cytotoxic T-lymphocyte activity against tumors, and increased the survival rate of test mice. In vivo antibody depletion experiments suggested that CD8+ T cells and NK cells, but not CD4+ T cells, were the main effector cells responsible for the observed anti-tumor activity. In addition, culture of DCs with GM-CSF and IL-4 during the pulsing and stimulation period significantly increased the production of IL-12 and decreased production of IL-10. On the other hand, MDAs also induced phenotypic maturation of DCs and augmented CD4+ and CD8+ T-cell proliferation when co-cultured with DCs. Taken together, specific MDAs including the clinical drug, colchicine, can induce immunogenic cell death in tumor cells, XXIII .
(28) and DCs pulsed with MDA-treated tumor cell lysates (TCLs) can generate potent anti-tumor immunity in mice. This approach may warrant future clinical evaluation as a cancer vaccine.. Keywords: immunogenic cell death; colchicine; 2-phenyl-4-quinolone; dendritic cells; cancer vaccine XXIV .
(29) Part III. Development of anti-severe acute respiratory syndrome associated coronavirus (SARS-CoV) agents is pivotal to prevent the reemergence of the life-threatening disease, SARS. In this study, more than 200 phytocompounds and more than 200 extracts from Chinese medicinal herbs were evaluated for activity against anti-severe acute respiratory syndrome associated coronavirus (SARS-CoV) activities using a cell-based assay measuring SARS-CoVinduced cytopathogenic effect on Vero E6 cells. Ten diterpenoids, two sesquiterpenoids, two triterpenoids, five lignoids, curcumin and reference controls, niclosamide and valinomycin were potent inhibitors at concentrations between 3.3 and 10 μM. Six herbal extracts, one each from Gentiana scabra, Dioscorea batatas, Cassia tora and Taxillus chinensis (designated as GSH, DBM, CTH and TCH, respectively), and two from Cibotium barometz (designated as CBE and CBM), were found to be potent inhibitors of SARS-CoV at concentrations between 25 and 200 μg/ml. The concentrations of the 22 compounds and six extracts to inhibit 50% of Vero E6 cell proliferation (CC50) and viral replication (EC50) were measured. The selective index values (SI = CC50/EC50) of the most potent compound 5 and 16 were >510 and >667, respectively. In addition, the resulting selective index values of the most effective extracts CBE, GSH, DBM, CTH and TCH were > 59.4, > 57.5, > 62.1, > 59.4, and > 92.9, respectively. Betulinic acid (13) and savinin (16) were competitive inhibitors of SARS-CoV 3CL protease with Ki values = 8.2 XXV .
(30) ± 0.7 and 9.1 ± 2.4 M, respectively. Among these extracts, CBM and DBM also showed significant inhibition of SARS-CoV 3CL protease activity with IC50 values of 39 μg/ml and 44 μg/ml, respectively. Our findings suggest that specific abietane-type diterpenoids and lignoids as well as these six herbal extracts exhibit strong anti-SARS-CoV effects.. Keywords: SARS; terpenoids; lignoids; phytoextracts; 3CL protease . XXVI .
(31) Part I. Novel quinolone CMQ induces apoptosis and mitotic catastrophe in prostate cancer cells via reactive oxygen species (ROS)- and mitochondriadependent pathways Chapter 1. Introduction. 1.1. Epidemiology of prostate cancer Cancer is probably the most extensive public health problem in the world including Taiwan. Prostate cancer is the most frequently diagnosed malignancy and the second leading cause of cancer-related deaths in men in the United States (1-3). In 2010, prostate cancer alone accounts for approximately 28 % of incidental cases and for 11 % of total cancer deaths in men (Figure 1) (3). Although the incidence of prostate cancer is lower in Asian countries, however, this type of cancer is one of the top 10 fatal cancers in Taiwan (Table 1) (4). Regional and early diagnosed tumors can be treated by radiation therapy and prostatectomy. However, it has been reported that numerous patients treated with prostatectomy suffer relapse (5, 6). The androgen-ablation therapy may be effective for the patients with relapsed disease at the initial stage, but the cancers eventually progress to hormone-refractory prostate cancer and metastasize to other organs, , leaving a majority of. 1 .
(32) the sufferers dead within a few years (2, 7, 8). To date, there is no effective treatment for locally advanced or metastatic prostate cancer. Therefore, it is particularly important to discover effective therapeutic agents that can treat androgen-dependent or/and androgen-independent prostate cancers without further progression and high mortality. In this context, many novel approaches for the treatment of prostate cancer are currently under development, including new androgen receptor antagonists, cytotoxic agents, antimitotic drugs, antiangiogenic agents, and immune-based therapies (2). Based on this rationale and importance, in this study, we investigate a novel quinolone derivative, CMQ, as a candidate anti-tumor agent in prostate cancer cells.. 2 .
(33) Figure 1. Ten leading cancer types for the estimated new cancer cases and deaths, by sex, United States, 2010. [Adopted from Jemal et al. (3) ]. 3 .
(34) Table 1. Ten leading cancer types for the estimated cancer–caused deaths, by sex, Taiwan, 2009.. Estimated deaths caused by cancer (2009 in Taiwan) Male Ranking. Location. 1. Liver & intrahepatic bile. Female deaths. %. Location. deaths. %. 5,467. 21.6. Lung and bronchus. 2,615. 22.8. Lung and bronchus. 5,336. 21.1. 2,292. 20.0. 3. Colon and rectum. 2,562. 10.1. Colon and rectum. 1,969. 17.2. 4. Oral cavity. 2,103. 8.3. Breast. 1,588. 13.9. 5. Stomach. 1,457. 5.8. Stomach. 825. 7.2. 6. Esophagus. 1,369. 5.4. Uterine corpus. 657. 5.7. 7. Prostate cancer. 936. 3.7. Pancreas. 609. 5.3. 8. Pancreas. 871. 3.4. Ovary. 435. 3.8. 9. Leukemia. 571. 2.3. Non-Hodgkin lymphoma. 372. 3.2. 10. Non-Hodgkin lymphoma. 542. 2.1. Leukemia. 353. 3.1. 4,070. 16.1. Others. 2,918. 25.5. 25,284. 100. 14,633. 100. duct 2. Others All sites. Liver & intrahepatic bile duct. The table was modified from Department of Health, Executive Yuan, Taiwan. (http://www.doh.gov.tw/CHT2006/DM/DM2_p01.aspx?class_no=25&level_no=1&doc_no =76013 ).. 4 .
(35) 1.2. The structure of microtubules and microtubule-binding agents (MBAs) In general, antimitotic drugs constitute a class of anticancer drugs that target the microtubule system (9, 10). A particularity of microtubule-binding agents compared to other categories of anticancer drugs is their various structural diversity and in many cases, their structural complexity (Figure 2). As shown in Figure 3a, microtubules, the key components of cytoskeletal filaments in all eukaryotic cells, are composed of a backbone of α- and β- tubulin heterodimers (9-11). To the best of our knowledge, in terms of drugs which interfere the tubulin polymerization, there are typically three binding-domains within microtubule structures (Figure 3b). Microtubule dynamics and stability, strongly associated with the rearrangement of depolymerization and polymerization, as shown in Figure 3c, are critical for cells undergoing replication and division where microtubule architecture vividly changes. Microtubules are also associated with various intracellular components, including mitochondria, the endoplasmic reticulum, the Golgi apparatus and lysosomes. As seen in Figure 4a and 4b, the mitotic spindle microtubules allow the appropriate alignment of chromosomes during metaphase, followed by the equal distribution of chromatids between the two daughter cells during anaphase (12). An increasing number of chemically diverse substances, originating from natural sources, have been discovered bound to tubulin and/or microtubules, interacting with microtubule 5 .
(36) polymerization, leading to mitosis arrest, resulting in apoptosis of cancer cells or other types of cell deaths (9, 11, 13, 14). According to their interference with microtubule dynamics and stability, antimitotic drugs are classified into two distinct groups, as summarized in Table 2. Microtubule-stablizing agents or microtubule-polymerizing agents, which bind to fully formed microtubules and promote polymerization of tunulin subunits, include the taxanes, such as paclitaxel and doxetaxel, and the epothilones. The second group, microtubule-destabilizing agents or microtubule-depolymerizing agents (MDAs), which bind mainly to the tubulin dimer and inhibit their polymerization into microtubules, include the vinca alkaloids, such as vincristine, vinblastine, and colchicine. Both the taxanes and the vinca alkaloids are currently used as chemotherapeutic drugs for various human cancers, whereas agents binding to the colchicine-binding site have been recognized as potential targets (14, 15). Although there are numerous microtubule-binding agents in clinical use (Table 2), the long-term use of many MBAs may result in drug resistance (Table 3). Therefore, it is vitally important to discover and develop new drug candidates to improve this difficult situation.. 6 .
(37) Figure 2. Various chemical structures of microtubule-binding agents, grouped according to their binding domains. a: vinca-domain binders; b: colchicine-domain binders; c: taxol-domain binders; d: others. This figure shows the extreme chemical diversity and the complexity of these agents. This figure is adopted from Dumontet et al. (10). 7 .
(38) Figure 3. The formation of microtubules and the binding sites for microtubule inhibitors. (a) The formation of microtubules. The soluble tubulin dimers, each containing one α-tubulin peptide and one β-tubulin peptide, polymerize to form a microtubule nucleus. Additional dimers are added head-to-tail; resulting microtubules are highly dynamic structures with a (+) end, characterized by an exposed β-tubulin peptide and a (–) end, characterized by an exposed α-tubulin peptide. (b) The binding sites of different microtubule inhibitors. Three binding sites of microtubule inhibitors are shown (orange triangles, purple squares and yellow diamonds). While vinca alkaloids bind to microtubule. 8 .
(39) ends, colchicine binds to soluble dimers, which are incorporated incorporated into the microtubules. Taxanes, such as paclitaxel, bind along the interior surface of the microtubules. The figures (a) and (b) are adopted from Dumontet et al. (10). (c) The dynamics and stability of microtubules. Microtubules are dynamic polymers assembled from tubulin heterodimers, which are organized so that the microtubules have an intrinsic polarity. Microtubules undergo periods of polymerization and depolymerization and interconvert randomly between these states, a property known as dynamic instability. This figure (c) was adopted from Walczak et al. (12).. 9 .
(40) Figure 4. The role of the microtubules in cell cycle. (a) The structure of the mitotic spindle during mitosis. Mitosis is staged into individual phases. During prometaphase, the spindle microtubules are connected with kinetochore. 10 .
(41) (k)-fibres (bundles of stabilized microtubules) and the kinetochores on the chromosomes, allowing the chromosome to align at the spindle equator,; this defines metaphase. The microtubules are uniformly oriented with their minus (-) ends at the centrosome and their plus (+) ends extending towards the spindle equator, where they often overlap. The astral microtubules emanate from the centrosomes and extend their plus (+) ends towards the cell’s cortex. The movement of the chromosomes towards the poles occurs during anaphase A, and the two spindle poles separate during anaphase B. The nuclear envelope begins to reform and the DNA begins to decondense during telophase. An organized central spindle bundle of microtubules is also present. This figure is adopted from Walczak et al. (12). (b) The microtubule changes during the cell cycle. The structures of microtubule (shown in green) undergo marked morphological changes to regulate specific functions throughout the cell cycle. Microtubule dynamics vary during the cell cycle, being most dynamic during mitosis and least dynamic in interphase cells. Most of tubulin-binding agents (TBAs) act upon the dynamics of the spindle microtubules, which are important for normal spindle function. Disruption of spindle microtubules causes mitotic arrest, leading to cell death through various mechanisms. DNA is shown in blue. This figure is adopted from Kavallaris et al.. (11). 11 .
(42) Table 2. Anti-mitotic drugs including microtubule-polymerizing agents and microtubule-depolymerizing agents currently in clinical use. Microtubule-polymerizing/-stabilizing agents Compound class. Compound. Clinical status. Taxanes. Paclitaxel. Approved for ovarian cancer, breast cancer and NSCLC; in. (taxol-domain binder). clinical trials for various solid tumour types Docetaxel. Approved for NSCLC, breast, prostate, stomach, head and neck cancer; in clinical trials for various solid tumour types. Epothilones. Cabazitaxel. Approved for metastatic hormone-resistant prostate cancer. Nab-paclitax. Approved for breast cancer; in clinical trials for various solid. el. tumours. Clinical trials for prostate cancer. Larotaxel. Phase III trials for pancreatic cancer. Ixabepilone. Approved for breast cancer; in clinical trials for solid tumours. (taxol-domain binder) Mcrotubule-depolymerizing/-destabilizing agents Compound class. Compound. Clinical status. Vinca alkaloids. Vincristine. Approved for ALL, lymphomas, various solid tumours; in. (vinca-domain binder). clinical trials for various tumour types Vinblastine. Approved for lymphomas and various solid tumours; in clinical trials for various tumour types. Vinorelbine. Approved for breast cancer and NSCLC; in clinical trials for various tumour types. Vindesine. Approved for ALL, lymphomas and lung cancer; in clinical trials for various tumour types. Vinflunine. Approved for bladder cancer; in clinical trials for breast cancer in combination with trastuzumab. Dolastatins. Romidepsin. (vinca-domain binder) Halichondrin. Approved for cutaneous T cell lymphoma; in clinical trials for myeloma, lymphoma and solid tumours. Eribulin. Phase III trials for advanced breast cancer. Ombrabulin. Phase III trials for sarcoma. (vinca-domain binder) Combretastatins (colchicine-domain binder). This table was modified from Dumontet et al. (10) 12 .
(43) Table 3. Characteristics of microtubule-binding agents.. This table is adopted from Dumontet et al.(10). 13 .
(44) 1.3. Cell death including apoptosis and mitotic catastrophe Various types of cell death have recently been defined and classified by the Nomenclature Committee on Cell Death (NCCD); these include apoptosis, autophagy, necrosis, cornification, and atypical cell death such as mitotic catastrophe (16). Programmed cell death (PCD) is a natural process for removing unwanted cells such as those with aberrant substratum attachment, potentially harmful mutations, or abnormal alterations in cell-cycle control (17). Apoptosis, a major type of PCD, is essentially a regulating mechanism by which cells undergo programmed cell death (18). In many malignant cells, the normal process for removing unwanted cells is deregulated (17). Anti-apoptotic mechanisms contribute to the development of cancer and to the resistance of cancer cells to anticancer chemotherapies. An increasing number of studies have demonstrated that promoting apoptosis is a good strategy for cancer drug discovery (17, 19-22). To date, at least two major apoptotic pathways have been described previously: the intrinsic and the extrinsic pathway (see Figure 5) (18, 19, 23). The intrinsic pathway is a mitochondrion-mediated process, influencing mitochondrial permeability and resulting in cytochrome c release and activation of caspase-9. The extrinsic apoptotic pathway, on the other hand, is triggered by the binding of specific ligands such as Fas ligands and TRAIL (tumor-necrosis factor-related apoptosis-inducing ligand) to membrane death receptors (DRs) such as Fas (CD95/APO-1), DR4 (TRAIL-R1), and DR5 (TRAIL-R2), resulting in 14 .
(45) recruitment of adaptor molecules and initiating activation of caspase-8 as well as distal executioner caspases including caspase-3. Additionally, the endoplasmic reticulum stress pathway is recognized as the third pathway to induce apoptosis (24). On the other hand, the p53 tumor suppressor functions as a key regulator of both intrinsic and extrinsic pathways (18). However, although current chemotherapeutic drugs can induce apoptosis, there is still an urgent need to find candidate drugs that will target an apoptotic pathway with a less adverse effect for practical, more productive cancer chemotherapy. Mitotic catastrophe is a type of cell death characterized by the occurrence of aberrant mitosis with the formation of large multi-nucleated cells, which may be morphologically distinguishable from apoptotic cells (25-27). However, mitotic catastrophe is a term sometimes used restrictively to illustrate a type of cell death that occurs during or after a defective mitosis, which takes place through apoptosis or necrosis, rather than a cell death itself (25, 26, 28). Moreover, it may be associated with complicated mechanisms of induction, as shown in Figure 6. Several antitumor drugs and ionizing radiation have been shown to induce mitotic catastrophe (26), but precisely how the ensuing lethality is controlled or what mechanisms are involved is less characterized. The type of cell death caused by antitumor therapy may be evaluated by the mechanisms of action of the antitumor agents, the dosing regimen, and perhaps the genetic background of the cells under treatment with anti-cancer agents. For example, oxaliplatin can trigger different 15 .
(46) types of cell death such as mitotic catastrophe and apoptosis in esophageal cancer cells (29). The proteasome inhibitor Bortezomib induces cell death via mitotic catastrophe and apoptosis in B-cell lymphoma cell lines (30). Doxorubicin may induce one of two distinct modes of cell death: apoptosis or cell death through mitotic catastrophe accompanied by senescence-like phenotype (31). The wild-type of p53 promotes apoptosis or senescence, whereas mitotic catastrophe is independent of p53 (25, 26). Mitotic catastrophe may be regarded as a delayed response of p53-mutant or p53-null tumors that are resistant to some damage especially for DNA damage (see Figure 7). In this context, characterization of the mechanisms of treatment-induced mitotic catastrophe should contribute to further improvement of the anti-tumor chemotherapy, due to the fact that the majority of solid tumors bear an inactive p53 protein (25).. 16 .
(47) Figure 5. Schematic representation of detailed mechanisms that govern the extrinsic and intrinsic apoptotic pathways and caspase-independent cell death. Essentially, the common denominator in apoptotic cell death processes is permeabilization of the outer mitochondrial membrane and the release into the cytosol of proteins that trigger death programs. The apoptotis-related caspases appear in yellow and red circles, respectively. Inhibitors of apoptosis are shown in green boxes. [Adopted from Blank et al.(18)]. 17 .
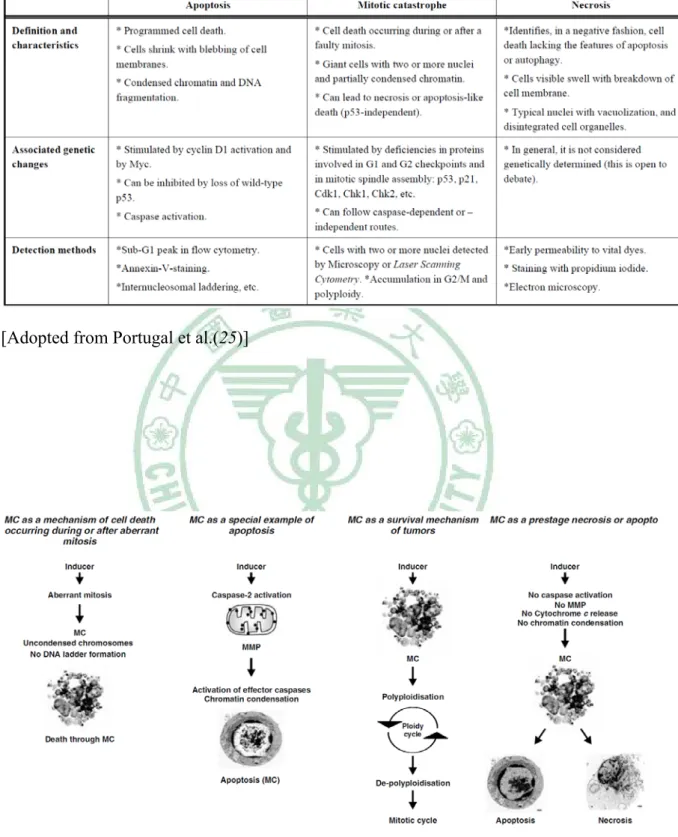
(48) Table 4. Comparison of major cell death pathways observed in drug-treated cells. [Adopted from Portugal et al.(25)]. Figure 6. Schematic representation of current concepts of mitotic catastrophe. [Adopted from Vakifahmetoglu et al. (26)] 18 .
(49) Table 5. Specific inducers of mitotic catastrophe (MC) and associated events. [Adopted from Vakifahmetoglu et al. (26)]. 19 .
(50) Figure 7. Pathways leading to mitotic catastrophe and cell death after DNA damage. Several key events occurring after a faulty mitosis are represented. DNA damage in p53-deficient cells can result in an exit from mitosis without cytokinesis, or in cell death occurring during mitosis. The relationship of mitotic catastrophe with apoptosis and/or necrosis is indicated. [Adopted from Portugal et al.(25)]. 20 .
(51) 1.4. Relationship between p53 and microtubule-binding agents Microtubule inhibitors can block the cell cycle at G2/M-phase by damaging the mitotic spindle. However, under certain conditions, specific cells escape these effects and become aneuploid, polyploid and/or micronucleated, or multi-nucleated (18, 32). For example, as shown in Figure 8, exposure to high doses of the microtubule-depolymerzing agent nocodazole may result in polyploidy due to mitotic slippage in the absence of a functional spindle. Furthermore, it is shown that p53 plays an important role in inducing cell death or ensuring cell survival. Also, different concentrations may cause different types of cell death. The mitotic defect with characteristics of monopolar, pseudo-bipolar, tripolar or multipolar spindles is possibly caused by different microtubule inhibitors including MDAs. As seen in Figure 9, these abnormal spindles can result in aneuploidy, polyploidy or other types of aberrant chromosomes. However, depending on whether cells express the p53 gene, cells are likely to die or to survive abnormally (33). We therefore hypothesize that cells expressed with or without expression of the p53 gene may be strongly related to different types of cell death or cell survival induced by microtubule inhibitors.. 21 .
(52) Figure 8. Schematic representation of putative mechanisms of a cell after exposure to low and high concentrations of the microtubule-depolymerizing agents, nocodazole. [Adopted from Decordier, I. et.al. (32).]. 22 .
(53) Figure 9. The relationship between mitotic defects with numeral abnormalities of spindles and p53. [Adopted from Fukasawa, K.et.al. (33).]. 23 .
(54) 1.5. CMQ as one of 2-phenyl-4-qunolone derivatives with potent anti-tumor activity As shown in Table 6, quinolone, a general structure occurring in natural alkaloids and its derivatives, exhibits several pharmacological activities including antibacterial activity, inhibition of leukotriene biosynthesis and antitumor activity (34-36). Furthermore, it has been reported that the 2-phenyl-4-quinolone derivatives exhibited various pharmacological activities including suppression of hind-paw edema and cutaneous vascular plasma extravasation (37), regulation of serotonin-mediated changes and permeability of endothelial monolayers (38), prevention of serotonin-induced increases in endothelial permeability to albumin (39), involvement in cyclic AMP generation in the inhibition of respiratory burst in rat neutrophils (40), antiplatelet activity (41), potent antagonists for glycine-binding site of the N-Methyl D-aspartate (NMDA) receptor (42),. potent. 5-hydroxytryptamine [5HT(1B)] antagonists (43), and especially anti-tumor activities in cancer cell lines by inhibiting polymerization of tubulins (Table 6) (44-52). Based on the rationale designs and efficacy results for 2-phenyl-4-quinolone derivatives, we found that 2-(3-chlorophenyl)-6, 7-methylenedioxyquinolin-4-one (CMQ) is a good candidate for further investigation of antitumor activities (45). Therefore, we investigated the effect of CMQ as a drug candidate for anti-tumor activities in p53-expressing LNCaP cells and p53-null PC-3 cells of prostate cancers. 24 .
(55) Table 6. A summarized list of bioactivities of quinolone and 2-phenyl-4- quinolone derivatives. Compounds. Bioactivities. Reference. quinolones. Antitumor quinolones with mammalian topoisomerase II mediated DNA cleavage activity. Yamashita et al. 1992. (36). 2-phenyl-4-quinolone derivatives. Positive inotropic effect in rat cardiac tissues. Su et al. 1993.(53). 1,6,7,8-substituted 2-(4'-substituted phenyl)-4-quinolones. Antimitotic agents interacting with tubulin. Kuo et al. 1993.(44). 2',3',4',5',5,6,7-substitute d 2-phenyl-4-quinolones. Cytotoxicity, and inhibition of tubulin polymerization. Li et al. 1994.(45). 2-phenyl-4-quinolone derivatives. Suppress hind-paw edema and cutaneous vascular plasma extravasation in mice. Wang et al. 1994.(37). quinolone derivatives. Antibacterial activity. Kamenska et al. 1996.(54) Renau et al. 1996.(55) Renau et al. 1996.(56) Yoshida et al. 1996.(57). 2-phenyl-4-quinolone. Regulate serotonin-mediated changes in the morphology and permeability of endothelial monolayers. Lee et al. 1997.(38). 2-phenyl-4-quinolone. Prevents serotonin-induced increases in endothelial permeability to albumin. Lee et al. 1998.(39). 2-phenyl-4-quinolone. Involve in cyclic AMP generation in the inhibition of respiratory burst in rat neutrophils. Wang et al. 1998.(40). 6,7,2',3',4'-substituted-1, 2,3,4-tetrahydro-2-pheny l-4-quinolones. Antimitotic antitumor agents. Xia et al. 1998.(46). 2-phenyl-4-quinolone. Antiplatelet activity. Huang et al. 1998.(41). 25 .
(56) fluoroquinolones. Inhibitiobn on DNA gyrase and topoisomerase II & IV. Hooper et al. 1999.(34). Fluorinated 2-phenyl-4-quinolone. Antimitotic antitumor agents. Xia et al. 2001.(47). Alkylation of 2-phenyl-4-quinolones. Synthesis. Hadjeri et al. 2001.(58). 2-phenyl-4-quinolone acetic acids and their esters. Antimitotic antitumor agents. Xia et al. 2003.(59). quinolone alkaloids. Inhibition of leukotriene biosynthesis by quinolone alkaloids from the fruits of Evodia rutaecarpa. Adams et al. 2004.(35). 5-hydroxy-7-methoxy-2phenyl-4-quinolones. Antimitotic antitumor agents. Hadjeri et al. 2004. (60). 2-phenyl-4-quinolone Antitumor-promoter and 9-oxo-9,10-dihydroacridi ne derivatives. Nakamura et al. 2005.(61). 3',6-substituted 2-phenyl-4-quinolone-3carboxylic acid derivatives. Antimitotic antitumor agents. Lai et al. 2005.(62). 2-phenyl-4-quinolone. Antimitotic antitumor agents. Chen et al. 2007.(48). quinoline-2-carboxylic acid (4-morpholin-4-yl-pheny l)amides. Potent 5HT(1B) antagonists. Horchler et al. 2007. (43). 7-choloro-4-hydroxy-3-[ Antagonist for the glycine-binding site of the 3-(4-methoxybenzyl) N-Methyl D-aspartate (NMDA) receptor phenyl]-2(1H)-quinolone. Matsumoto et al. 2007.(63). 2-(3-Fluorophenyl)-6-me Induces mitotic phase arrest in A549 cells thoxyl-4-oxo-1,4-dihydr oquinoline-3-carboxylic acid (YJC-1). Hsu et al. 2007.(64). 2-(2-fluorophenyl)-6,7Promoted G2/M arrest through inhibition of methylenedioxyquinolin- CDK1 and induced apoptosis through the 4-one (CHM-1) mitochondrial-dependent pathway in CT-26. Wang et al. 2007.(65). 26 .
(57) murine colorectal adenocarcinoma cells Potent and selective antimitotic antitumor activity against human hepatocellular carcinoma in vitro and in vivo. Wang et al. 2008.(49). Promoted G2/M arrest through inhibition of CDK1 and induced apoptosis through the mitochondrial-dependent pathway in CT-26 murine colorectal adenocarcinoma cells. Chou et al. 2009.(50). Induces apoptosis and inhibits metastasis in a human osterogenic sarcoma cell line. Hsu et al. 2009.(52). Induces DNA damage and inhibits DNA repair gene expressions in a human osterogenic sarcoma cell line. Chen et al. 2010.(66). Induces apoptosis of human umbilical vein endothelial cells via p53-mediated death receptor 5 up-regulation. Tsai et al. 2010.(51). Inhibits murine WEHI-3 leukemia in BALB/c mice in vivo. Lai et al. 2010.(67). 2-(2-fluorophenyl)-6,7Antimitotic antitumor agents methylenedioxyquinolin4-one monosodium phosphate (CHM-1-P-Na). Chou et al. 2010.(68). 2(3chlorophenyl)6,7 methylenedioxyquinoli n4one (CMQ). Wen et al. 2011.(69). Augment efficacy of dendritic cell-based cancer vaccines. 27 .
(58) Chapter 2. Rationale and Significance The rationale of this study used CMQ as a lead compound for investigating the anti-tumor activity in LNCaP and PC-3 cells are as follows. It has been reported that derivatives of 2-phenyl-4-quinolone induce cytoxicity and apoptosis in human cancer cell lines by inhibiting the polymerization of tubulins and interfering with microtubule organization (44-52). In a previous study, we reported that 2-(3-chlorophenyl)-6,7-methylenedioxyquinolin-4-one (CMQ) holds potential for further research and development into a specific antitumor agent (Fig. 1A) (45). To date, there is no effective treatment for either locally advanced or metastatic prostate cancer. Usually, these cancers are androgen-independent and p53 -null or -mutated. Therefore, various new therapeutic approaches, including new androgen receptor antagonists, antimiotic drugs, antiangiogenic agents, and immune-based therapies are being actively evaluated (2). On the other hand, notably, we understand two key pieces of information as follows: prostrate cancers are androgen dependent [6-7], and TRAIL and p53 play key roles in apoptosis (18, 23). However, there is no available research which compares the effects of 2-phenyl-4-quinolone derivatives on cancer cells concurrently with or without the characteristics described above. Therefore, in this study, we evaluated the extent to which CMQ exerts antitumor activities in prostate cancer cells including LNCaP cells 28 .
數據



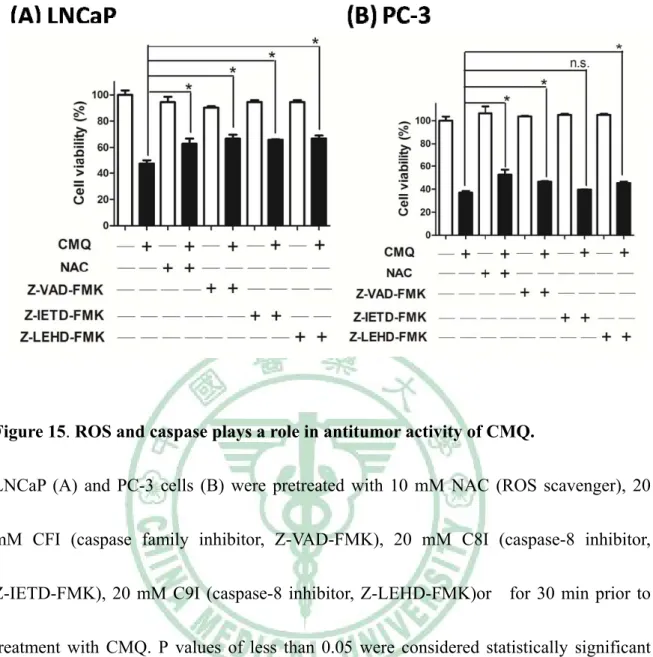
+7




相關文件
臺大機構典藏NTUR (National Taiwan University 二 Repository, http://ntur.lib.ntu.edu.tw) 經驗與協助推 動臺灣學術機構典藏TAIR (Taiwan Academic Institutional Repository,
The students were either copying the notes on the board //or correcting the mistakes on the test(quiz), //but Peter and Jack weren't. The cat which/ that almost drowned
The accuracy of a linear relationship is also explored, and the results in this article examine the effect of test characteristics (e.g., item locations and discrimination) and
By correcting for the speed of individual test takers, it is possible to reveal systematic differences between the items in a test, which were modeled by item discrimination and
N248-365 of SARS CoV Telomere binding protein.. Gallery of
◦ Lack of fit of the data regarding the posterior predictive distribution can be measured by the tail-area probability, or p-value of the test quantity. ◦ It is commonly computed
D., (2004) 'Mindfulness-Based Cognitive Therapy for Depression: Replication and Exploration of Differential Relapse Prevention Effects', Journal of Consulting and Clinical
After the SARS crisis in 2003, there is a huge and on-going reform in the post-graduate medical training program to improve the medical service in Taiwan. Our institute has
