Oxidized Low-Density Lipoproteins Inhibit Endothelial Cell
Proliferation by Suppressing Basic Fibroblast Growth
Factor Expression
Chu-Huang Chen, MD, PhD; Wei Jiang, MD; David P. Via, PhD; Sherry Luo, BS; Tz-Rung Li, MS;
Yuan-Teh Lee, MD, PhD; Philip D. Henry, MD
Background—Hyperlipidemia inhibits proliferation of endothelial cells (ECs) in culture and angiogenesis in vivo and in arterial explants. Elucidation of the mechanisms may suggest novel therapies against atherosclerosis.
Methods and Results—Basic fibroblast growth factor (bFGF) expression and mitogenic effects were assessed in bovine aortic ECs incubated with oxidized LDL (ox-LDL). Compared with native LDL and lipoprotein-free controls, ox-LDL reduced bFGF mRNA levels in a time- and concentration-dependent manner, 100 mg/mL producing a maximum reduction of 40% to 50% within 24 to 48 hours. There were commensurate reductions in intracellular and extracellular bFGF concentrations, DNA and total RNA syntheses, and cell replication. FGF receptor 1 andb-actin mRNA levels were unchanged. Ox-LDL accelerated bFGF mRNA degradation in actinomycin D–treated cells. However, inhibition of bFGF expression by ox-LDL was attenuated by cyclohexamide, indicating a requirement for continuous new protein synthesis for posttranscriptional destabilization. Reduced syntheses of DNA and total RNA were completely restored by bFGF but not by vascular endothelial growth factor. Inhibition of total RNA synthesis achieved by exposing cells to a bFGF-neutralizing antibody was similar in magnitude to that induced by ox-LDL.
Conclusions—Cytotoxic effects of ox-LDL on ECs are attributable in part to suppression of bFGF expression.
(Circulation. 2000;101:171-177.)
Key Words: lipoproteinsn growth substances n endothelium n genes n angiogenesis
H
yperlipidemia impairs function of both large-vessel and microvascular endothelium. We have shown that com-pensatory macrovascular and microvascular growth is im-paired in rabbits with diet-induced hypercholesterolemia.1Impaired capillary-like microtube growth in arterial explants exposed to hypercholesterolemia in vivo or to oxidized LDL (ox-LDL) in vitro is associated with reduced basic fibroblast growth factor (bFGF) concentrations in the culture medium. The impairment can be partially reversed by exogenous bFGF.2,3Here, we determined whether impairment of
prolif-erative and angiogenic responses induced by ox-LDL is due to suppressed endothelial expression of bFGF or of its receptor, FGFR-1. The effects of bFGF on DNA and total RNA syntheses in the presence of ox-LDL were compared with those of vascular endothelial growth factor (VEGF), an endothelial cell (EC)–specific mitogen and potent angiogenic factor.4,5
Methods
CellsPrimary cultures of bovine aortic ECs (BAECs) were derived by a previously reported procedure.2Cell purity was assessed by uptake
of acetylated LDL labeled with 1,1 9-dioctadecyl-1,3,3,39,39-tetra-methylindocarbocyanine perchlorate (Biomedical Technologies), immunocytochemical staining for von Willebrand factor–related antigen with FITC-labeled monoclonal antibody (Incstar), and neg-ative staining fora-actin with HHF35 antibody (Enzo).2,6More than
98% of the cells exhibited responses typical of cells of endothelial origin. Cells at 8 to 12 passages, maintained in DMEM supplemented with 10% FBS and antibiotics (streptomycin 100mg/mL, penicillin 100 IU/mL, amphotericin B 0.25mg/mL), were used.
Preparation of Native LDL and Ox-LDL
Native LDLs (d51.019 to 1.063 g/mL) from pooled human plasma anticoagulated with EDTA 0.5 mg/mL were isolated by sequential ultracentrifugation.7 Ox-LDLs were prepared by exposing native
LDL for 24 hours at 37°C to 5mmol/L CuSO4in PBS.8Mildly
oxidized LDLs were prepared by exposing native LDLs to CuSO4for
Received February 23, 1999; revision received July 6, 1999; accepted July 21, 1999.
From the Department of Medicine, Baylor College of Medicine, Houston, Tex (C.-H.C., W.J., D.P.V., S.L., P.D.H.), and the Department of Internal Medicine, National Taiwan University Hospital, Taipei (T.-R.L., Y.-T.L.).
Guest Editor for this article was Jeffrey M. Isner, MD, St Elizabeth’s Medical Center, Boston, Mass.
Presented in abstract form at the Biomedicine 96 Meeting of the American Association of Physicians, Washington, DC, May 3– 6, 1996, and the 69th Scientific Sessions of the American Heart Association, New Orleans, La, November 10 –13, 1996, and published in abstract form (Circulation. 1996;94[suppl I]:I-277).
Correspondence to Chu-Huang Chen, MD, PhD, Baylor College of Medicine, 6565 Fannin, MS A-601, Houston, TX 77030. E-mail [email protected]
© 2000 American Heart Association, Inc.
Circulation is available at http://www.circulationaha.org
Ox-LDL contained thiobarbituric acid–reactive substance concentra-tions of 18 to 22 nmol/mg LDL protein; mildly oxidized LDL contained 6 to 8 nmol/mg LDL protein.
Protocol
Cell cultures grown to subconfluence were washed 3 times with serum-free medium and maintained under serum-free conditions for another 24 hours before experiments. To demonstrate a concentration-dependent effect of ox-LDL on bFGF and on FGFR-1 expression, cells were incubated with 25 to 200mg/mL ox-LDL for 24 hours. In other experiments, the effect of mildly oxidized LDL was compared with that of ox-LDL. To determine reaction time course, cells were treated with 100mg/mL ox-LDL for 6, 12, 24, and 48 hours. To ascertain whether the effects of ox-LDL on DNA and RNA syntheses could be reversed by recombinant human bFGF or VEGF-165 (R&D Systems), cells were treated with each in the presence or absence of ox-LDL. In some experiments, bFGF-neutralizing antibody (R&D Systems) was added.
For the study of DNA and RNA syntheses, cell growth, and intracellular bFGF protein, 1003104cells were seeded in each well
of 12-well Corning cell culture plates.
Reverse Transcriptase–Polymerase Chain Reaction
To determine bFGF mRNA levels, total RNA was extracted from control or ox-LDL–treated cells.9One microgram of the extracted
total RNA was reverse transcribed in 10 mL reaction mixture containing 2.5 U Moloney murine leukemia virus reverse transcrip-tase (RT) for 45 minutes at 42°C. A fraction of the synthesized cDNA was subjected to polymerase chain reaction (PCR) amplifi-cation (GeneAmp, Perkin-Elmer Cetus Co). The bFGF primers were 59-GGAGTGTGTGCTAACCGTTACCTGGCTATG-39 (upstream) and 59-TCAGCTCTTAGCAGACATTGGAAGAAAAAG-39 (downstream). The FGFR-1 primers were 5 9-AAGGACAAACCC-AACCGTGTGACC-39 (upstream) and 59-CCCAAAGTCTGCTA-TCTTCATCAC-39 (downstream).10 The cDNA mixture
supple-mented with the primers, Taq DNA polymerase (AmpliTaq), PCR buffer, dNTP, and MgCl2was then incubated in a PTC-100
program-mable thermal cycler (step-cycle file for 30 cycles at 94°C for 1 minute, 54°C for 1 minute, and 72°C for 1 minute, followed by a time delay file at 72°C for 10 minutes). As an internal control, levels ofb-actin mRNA were estimated under the same cycling conditions as used in bFGF amplification with the following primers: 5 9-AACCGCGAGAAGATGACCCAGATCATGTTT-39 (upstream) and 59-AGCAGCCGTGGCCATCTCTTGCTCGAAGTC-39 (down-stream).11 The PCR profile was optimized in preliminary
experi-ments to ascertain linearity of amplification for bFGF, FGFR-1, and
b-actin genes. A fraction of each PCR product (10 mL) was
electrophoresed in 2% agarose gel, and DNA bands stained with ethidium bromide were visualized by ultraviolet transillumination.
RNase Protection Assay
RNase protection assays were performed with a Guardian kit (Ambion)12with the bFGF RNA probe prepared by in vitro
tran-scription from a bFGF cDNA insert previously cloned in the TA vector (Invitrogen). An RT-PCR product from BAECs using bFGF primers as above was inserted into the TA cloning vector and completely sequenced. This plasmid was linearized with EcoRV, and the antisense RNA was synthesized with SP6 RNA polymer-ase and [a-32P]uridine triphosphate (DuPont NEN), followed by
purification through a G-50 column (Boehringer). Labeled probe (150 000 cpm) was then mixed with 20mg total RNA extracted
from cells subjected to selected treatments and incubated at 42°C overnight. After hybridization, the mixture was treated with ribonuclease to degrade single-strand, unhybridized probes. Sam-ples of the mixture were separated by electrophoresis in urea polyacrylamide gel and exposed to x-ray films at270°C with an intensifying screen. Relative mRNA levels were quantified by densitometry with an Ultro Scan densitometer (LKB Produkter).
Northern Blot Analysis
Total RNA was isolated from cultured cells with RNAzol (Biotecx Laboratories, Inc). Twenty micrograms of total RNA was subjected to electrophoresis in each lane of 0.7% agarose/2.2 mol/L formalde-hyde gel, then transferred to Nytran membranes (Schleicher & Schuell).13 The bFGF cDNA insert was excised from plasmid
constructs with EcoRI and gel-purified (Geneclean Kit-Bio 101, Inc) to be used as probes. After labeling with [a-32P]dCTP to a high
specific activity ('109 cpm/mg) by a random priming method,
probes were hybridized to the blots (106cpm/mL) overnight at 42°C
in a solution containing 40% formamide, 53SSC, 53Denhardt’s solution, 0.5% SDS, 250 mg/mL salmon sperm DNA, and 10% dextran sulfate. The blots were then washed under high stringency at 65°C in 0.23SSC plus 0.1% SDS. Films were exposed at 270°C for 3 days with 2 intensifying screens for autoradiography.b-Actin was used in each experiment as internal control (not shown).
Actinomycin D and Cyclohexamide
To inhibit RNA transcription, cells were treated with 50 ng/mL actinomycin D, which decreased [3H]uridine uptake by.95%. To
Figure 1. Effects of LDL preparations on bFGF andb-actin expression evaluated by RT-PCR. A, Relative bFGF mRNA lev-els were expressed in BAECs incubated for 24 hours with PBS (C, control), 100mg/mL native LDL (N), or 50 or 100 mg/mL ox-LDL (Ox). M denotes marker. B, Cells were incubated for 24 hours with PBS (control), 100mg/mL mildly oxidized LDL (Cu21 for 4 hours), and 100mg/mL ox-LDL (Cu21for 24 hours). F and b denote bFGF and b-actin, respectively.
Figure 2. Effects of ox-LDL on bFGF, FGFR-1, andb-actin expression evaluated by RT-PCR. BAECs were incubated for 24 hours with PBS (C, control) or 50 or 100mg/mL ox-LDL. M denotes marker.
determine whether ox-LDL accelerated posttranscriptional degrada-tion of bFGF mRNA, ox-LDL was included in the medium of actinomycin D–treated cells.14To determine whether bFGF mRNA
degradation required new protein synthesis, cyclohexamide 1.5
mg/mL was used.14
Enzyme-Linked Immunosorbent Assay
Extracellular bFGF concentrations in culture medium collected during the serum-free period were measured by ELISA with a Quantikine Kit (R&D Systems). Medium samples and bFGF stan-dards were incubated at room temperature for 2 hours in wells of the microtiter plate coated with a murine bFGF monoclonal antibody. After a washing, the cells were incubated for 2 hours with a rabbit polyclonal antibody against bFGF conjugated to horseradish perox-idase. The bFGF concentration in each well was estimated spectro-photometrically at 450 nm by use of standard curves.3The bFGF
concentrations in cell lysates prepared with Nonidet P-40 (Sigma) were assayed similarly.
DNA and Total RNA Syntheses
To evaluate treatment effects on DNA synthesis, 3mCi/mL [3
H]thy-midine (Moravek Biomedicals) was included in the medium during the final 4 hours of incubation. Incubation was terminated by decanting the medium and fixing the cells with 1 mL of 10% (wt/vol) cold trichloroacetic acid for 15 minutes at 4°C. [3H]Thymidine
incorporated in extracted DNA was assayed by scintillation spec-trometry.2,3To evaluate total RNA synthesis, 5mCi/mL [3H]uridine
was included during the final 2 hours of incubation. Cells were fixed
with trichloroacetic acid, and RNA was extracted for detection of incorporated [3H]uridine.15
Statistical Analysis
The significance of the differences between group means was assessed by a 2-sided Student’s t test for single comparisons and Bonferroni’s test for multiple comparisons. Probability values,0.05 were considered significant. Results are expressed as mean6SD. A GB-STAT program (Dynamic Microsystems, Inc) was used.
Results
Effects of Ox-LDL on bFGF and on FGFR-1 ExpressionIncubation of the cells with 50 and 100 mg/mL ox-LDL suppressed, in a concentration-dependent manner, the expres-sion of bFGF mRNA as assessed by RT-PCR but had no effect on “housekeeping” b-actin expression (Figure 1A). Native LDL and mildly oxidized LDL had no effect on these expressions (Figure 1A and 1B). Expression of FGFR-1 was not inhibited by ox-LDL at concentrations up to 100mg/mL (Figure 2). At ox-LDL concentrations from 25 to 200mg/mL, RNase protection assays demonstrated a concentration-dependent decrease in bFGF mRNA (Figure 3). The reduc-tions were clearly demonstrable at 50mg/mL and were '50% at 100mg/mL ox-LDL, a concentration that did not increase
Figure 3. Effects of increasing concentrations of ox-LDL on bFGF andb-actin mRNA levels assessed by RNase protection assay and den-sitometry in BAECs incubated for 24 hours with 25, 50, 100, or 200mg/mL ox-LDL, relative to PBS control (C).
Effects of LDL Preparations on DNA and Total RNA Syntheses and on Cell Proliferation at 24 Hours
Assessment PBS (Control) LDL 100mg/mL Ox-LDL 25mg/mL Ox-LDL 50mg/mL Ox-LDL 100mg/mL Ox-LDL 200mg/mL [3H]thymidine incorporation (DNA), cpm31024/well 8668 8469 75610 65610* 60615* 29615†
[3H]uridine incorporation (RNA), cpm31024/well 5363 5264 4865 4663* 4262* 25610†
Cell count, cells31024/well 210613 202611 194614 186616 152614* 102622†
Dead cells, %/well 14610 13612 15610 17612 17613 56613†
n56 in all treatments; 1003104cells/well at inoculation.
the percentage of dead cells (Table). At higher concentra-tions, up to 200mg/mL, there was an increase in cell death but no further reduction in bFGF mRNA (Figure 3).
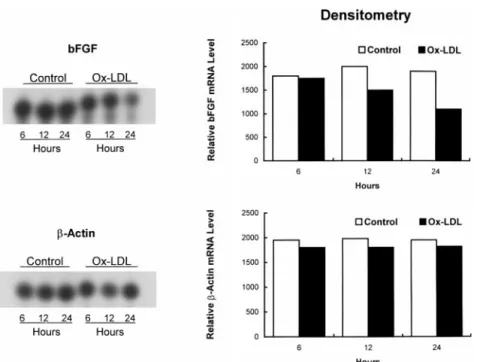
In time-course experiments performed with an ox-LDL concentration of 100 mg/mL, reduction of bFGF mRNA became apparent at 12 hours and reached the maximum of 40% to 50% at 24 to 48 hours (Figures 4, 5, and 6). Posttranscriptional Destabilization
Actinomycin D 50 ng/mL decreased [3H]uridine uptake by
.95% (from 33.562.5 to 1.060.5 cpm, n56, P,0.001). In
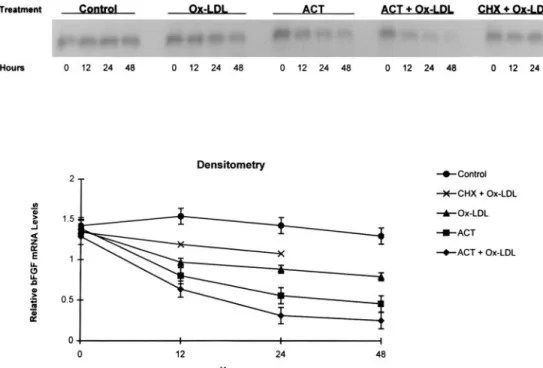
the presence of actinomycin D, bFGF mRNA decreased by 60% and cell viability was detectably impaired after 24 hours (Figure 6). Combined treatment with actinomycin D and ox-LDL accelerated bFGF mRNA degradation; message reductions reached 50% by 12 hours (n54). Reductions with combined treatment exceeded those seen with ox-LDL or actinomycin D alone and resulted in barely visible messages
mRNA that required sustained synthesis of new protein(s). Intracellular and Extracellular
bFGF Concentrations
Ox-LDL treatment produced concentration-dependent de-creases in intracellular bFGF peptide detectable by ELISA. In cells exposed to 0, 25, 50, or 100 mg/mL ox-LDL for 24 hours, the intracellular bFGF concentration (n54) averaged
316624, 177660, 153653, and 115632 pg/mg protein,
respectively (Figure 7). These peptide reductions paralleled those of bFGF mRNA, although bFGF mRNA reductions at the lowest ox-LDL concentration were not detectable. The average extracellular bFGF concentration was 7.660.8 pg/mL (n54) for PBS controls. After 24-hour incubation with 50mg/mL ox-LDL, assayable bFGF was reduced to 5.260.4 pg/mL (P,0.05); with 100mg/mL ox-LDL, bFGF was not detectable (,5 pg/mL). At an ox-LDL concentration of 200
mg/mL, most cells ruptured, and the bFGF concentration in
the medium surged to 32614 pg/mL (P,0.01). Effects of Ox-LDL on DNA and Total RNA Syntheses and Cell Proliferation
Ox-LDL decreased DNA and total RNA syntheses in a concentration-dependent manner (Table). At 50mg/mL, syn-thesis reductions were moderate but statistically significant. At 100 and 200mg/mL, synthesis decreases were accompa-nied by reductions in total cell count. At 100mg/mL, few trypan blue–positive cells were detectable, but at 200mg/mL, many cells exhibited disrupted membranes and .50% showed positive staining.
Figure 4. Time-course effects of ox-LDL on bFGF andb-actin expression evaluated by RT-PCR. BAECs were incubated with PBS (control) or 100mg/mL ox-LDL for 6, 12, or 24 hours.
Figure 5. Time-course effects of ox-LDL on bFGF andb-actin mRNA levels assessed by RNase protection assay and densitometry in BAECs incubated with 100 mg/mL ox-LDL for 6, 12, or 24 hours rela-tive to PBS control (C).
bFGF and VEGF Efficacies in Counteracting Ox-LDL Inhibition of DNA and Total
RNA Syntheses
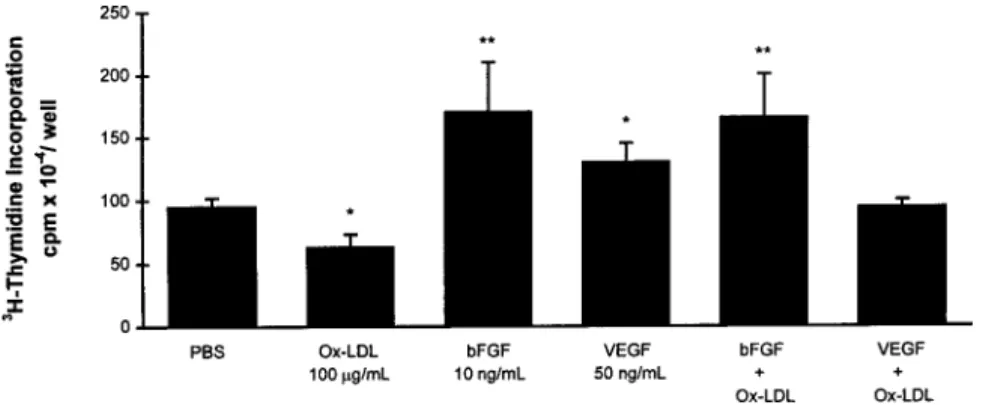
Applied alone, bFGF 10 ng/mL increased DNA synthesis 2-fold, and VEGF-165 50 ng/mL evoked an increase of lesser magnitude (n56; Figure 8). VEGF-165 and VEGF-121 had similar effects; maximal stimulation was at 50 ng/mL (data not shown). DNA synthesis was increased to a similar extent by bFGF whether ox-LDLs were present or absent, which suggests that bFGF prevented inhibition by ox-LDL 100
mg/mL. In contrast, VEGF-stimulated DNA synthesis was
sensitive to ox-LDL inhibition.
bFGF increased total RNA synthesis to the same extent in the presence or absence of ox-LDL (n56; Figure 9). VEGF-165 applied alone failed to increase total RNA synthesis or to counteract its inhibition by ox-LDL. bFGF-neutralizing anti-body 10mg/mL produced a maximal decrease in total RNA synthesis, very close to that obtained with 100 mg/mL ox-LDL.
Discussion
We demonstrated that ox-LDLs reduce steady-state bFGF mRNA levels in cultured vascular ECs. The time- and concentration-dependent suppression of bFGF expression was indicated by decreases in both intracellular and extracel-lular bFGF concentrations. Expression of 2 important refer-ence genes, FGFR-1 and b-actin, was preserved, consistent with selective bFGF gene inhibition. This would explain why exogenous bFGF was capable of completely restoring DNA and total RNA syntheses, even in the presence of ox-LDL. The restorative effects of bFGF were not matched by those of VEGF, an EC-specific mitogen.4,5 These results support
findings that hyperlipidemic impairment of angiogenesis is associated with reduced availability of bFGF and can be corrected by exogenous bFGF.2,3
The progressive reduction in bFGF expression reflected, in part, accelerated posttranscriptional mRNA degradation. Ox-LDL shortened the bFGF half-life from 24 to'12 hours after transcription was inhibited by actinomycin D. Sensitive to
Figure 7. Effects of LDL preparations on intracellular bFGF concentration (24-hour incubations). *P,0.05, **P,0.01 vs PBS control.
Figure 6. Effects of actinomycin D (ACT, 50 ng/mL, 0 through 48 hours) and cyclohexamide (CHX, 1.5mg/mL, 0 through 24 hours) in presence or absence of 100mg/mL ox-LDL on bFGF expression, compared with PBS (control), assessed by Northern blot analysis and densitometry.
cyclohexamide, enhanced degradation of bFGF mRNA de-pended on newly synthesized protein(s). The effects resemble endothelial nitric oxide synthase expression by ox-LDL, attributed to combined posttranscriptional mRNA degrada-tion and early transcripdegrada-tional inhibidegrada-tion.14
Maximal inhibition of bFGF expression required 24 to 48 hours of incubation with ox-LDL, but significant inhibition was evident by 12 hours. Similar early and progressive reductions induced by ox-LDL were reported for the nitric oxide synthase gene.14bFGF message level reductions
cor-related with reductions in immunoassayable bFGF concen-tration in both cytoplasm and culture medium. This may have functional implications, because bFGF exerts its effects through both intracellular and extracellular mechanisms. Ox-LDL decreased DNA synthesis, and to a lesser extent total RNA synthesis, in a concentration-dependent fashion. With concentrations up to 100 mg/mL, only reductions in total cell number were noted, but with higher concentrations, increases in dead cells were evident. Also, concentration-dependent effects of ox-LDL on assayable extracellular bFGF were observed only up to 100 mg/mL. At 200 mg/mL, extracellular bFGF surged, reflecting release of peptide from irreversibly damaged cells.
In many systems, ox-LDLs exert biphasic effects: low concentrations or brief incubations are stimulatory, and high concentrations or prolonged incubations are inhibitory. Com-plex, concentration-dependent effects of ox-LDL may reflect actions of distinct stimulatory or inhibitory mediators such as platelet-activating factor (PAF)16,17or oxysterols.18However,
single mediators, such as lysophosphatidylcholines, may by themselves exert biphasic effects.19The present experiments
further illustrate the complex actions of ox-LDL. As noted, ox-LDL suppressed bFGF expression without apparent effect
on FGFR-1 and b-actin, although suppressant effects of ox-LDL have been reported for other genes such as nitric oxide synthase14and thrombomodulin.20Yet ox-LDLs tend to
activate vasoconstrictor, proinflammatory, and procoagula-tory genes.21–24Suppressant effects on DNA and total RNA
syntheses as observed here may reflect inflammatory re-sponses leading to apoptosis.25
In contrast to bFGF, VEGF failed to reverse the inhibitory effects of ox-LDL on DNA and total RNA syntheses. In the absence of ox-LDL, high-dose VEGF increased DNA but not RNA synthesis. VEGF has been shown to increase DNA synthesis in bovine ECs26and to improve collateral development
indices in the ischemic limbs of Watanabe heritable hyperlipid-emic rabbits.27The role of VEGF in RNA synthesis, however,
has not been investigated. In contrast, bFGF acts as an autocrine stimulator and can stabilize RNA in the presence of actinomycin D. Also, it increases total cellular RNA synthesis in neuropeptide-producing 44-2C cells treated withaA, an inhib-itor of RNA polymerase II.15Although [3H]uridine uptake has
limitations as an index, its increase or decrease yields useful qualitative information on RNA synthesis.
Although bFGF has been used to induce angiogenesis,28,29its
role in regulating angiogenesis remains incompletely defined. Other factors, including VEGF and its receptors, angiopoietin and the Tie2 receptor, transforming growth factor-a, and plate-let-derived growth factor B, have also been recognized as important.30,31The present findings and those of our previous
reports with the explant model do not provide mechanistic information. The data clearly indicate, however, that vascular ECs of various species (human, rabbit, bovine) are unable to replicate or form microvessels when the cells are deprived of bFGF.2,3We and others have emphasized as a potential
mech-anism the importance of phospholipid mediators contained in
Figure 9. Effects of ox-LDL, bFGF, and VEGF-165, alone or in combination, and of bFGF-neutralizing antibody (bFGF-Ab) on total RNA synthesis (24-hour incuba-tions). *P,0.05 vs PBS control.
modified LDL.8,16,17,23,32–34There is evidence that these
media-tors, including PAF and lysophosphatidylcholines and their phospholipid analogues, act by means of PAF receptors.35
Recently, ox-LDLs have been shown to downregulate PAF receptor expression, consistent with the action of ox-LDL phospholipids through the PAF receptor pathway. It is clear, however, that other lipids in ox-LDLs, in particular oxysterols,18
are likely to play roles in mediating its effects.
Acknowledgments
This study was supported in part by Grant-in-Aid 95G-239 from the American Heart Association, Texas Affiliate, Austin, Tex, and Scientist Development Grant 9630095N from the American Heart Association National Center, Dallas, Tex. Li and Lee were supported by grant NSC85-2331-B-002-123 from the National Science Council, Taipei, Taiwan. The authors are grateful to Suzanne Simpson for editorial assistance and to Bo Dakus for computer graphics.
References
1. Bucay M, Nguy J, Barrios R, Chen CH, Henry PD. Impaired adaptive vascular growth in hypercholesterolemic rabbit. Atherosclerosis. 1998;139: 243–251.
2. Chen CH, Nguyen HH, Weilbaecher D, Luo S, Gotto AM Jr, Henry PD. Basic fibroblast growth factor reverses atherosclerotic impairment of human coronary angiogenesis-like responses in vitro. Atherosclerosis. 1995;116: 261–268.
3. Chen CH, Cartwright J Jr, Li Z, Luo S, Nguyen HH, Gotto AM Jr, Henry PD. Inhibitory effects of hypercholesterolemia and oxidized LDL on angiogenesis-like endothelial growth in rabbit aortic explants: essential role of basic fibroblast growth factor. Arterioscler Thromb Vasc Biol. 1997;17: 1303–1312.
4. Leung DW, Cachianes G, Kuang WJ, Goeddel DV, Ferrara N. Vascular endothelial growth factor is a secreted angiogenic mitogen. Science. 1989; 246:1306 –1309.
5. Takeshita S, Kearney M, Loushin C, Brogi E, Zheng LP, Horowitz J, Ferrara N, Symes JF, Isner JM. In vivo evidence that vascular endothelial growth factor stimulates collateral formation by inducing arterial cell proliferation in a rabbit ischemic hindlimb. J Clin Invest. 1994;93:662– 670.
6. Voyta JC, Via DP, Butterfield CE, Zetter BR. Identification and isolation of endothelial cells based on their increased uptake of acetylated low density lipoprotein. J Cell Biol. 1984;99:2034 –2040.
7. Steinbrecher UP, Parthasarathy S, Leake DS, Witztum JL, Steinberg D. Modification of low density lipoprotein by endothelial cells involves lipid peroxidation and degradation of low density lipoprotein phospholipids. Proc
Natl Acad Sci U S A. 1984;81:3883–3887.
8. Mangin E Jr, Kugiyama K, Nguy J, Kerns S, Henry PD. Effects of lysolipids and oxidatively modified low density lipoprotein on endothelium-dependent relaxation of rabbit aorta. Circ Res. 1993;72:161–166.
9. Chomczynski P, Sacchi N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal Biochem. 1987; 162:156 –159.
10. Brogi E, Winkles JA, Underwood R, Clinton SK, Alberts GF, Libby P. Distinct patterns of expression of fibroblast growth factors and their receptors in human atheroma and nonatherosclerotic arteries. J Clin Invest. 1993;92: 2408 –2418.
11. Bonilla-Felix M, Jiang W. Expression and localization of prostaglandin EP3 receptor mRNA in the immature rabbit kidney. Am J Physiol. 1996;271(1 pt 2):F30 –F36.
12. Becks GP, Logan A, Phillips ID, Wang JF, Smith C, DeSousa D, Hill DJ. Increase of basic fibroblast growth factor (FGF) and FGF receptor messenger RNA during rat thyroid hyperplasia: temporal changes and cellular distribution. J Endocrinol. 1994;142:325–338.
13. Tomanek RJ, Lotun K, Clark EB, Suvarna PR, Hu N. VEGF and bFGF stimulate myocardial vascularization in embryonic chick. Am J Physiol. 1998;274(5 pt 2):H1620 –H1626.
14. Liao JK, Shin WS, Lee WY, Clark SL. Oxidized low-density lipoprotein decreases the expression of endothelial nitric oxide synthase. J Biol Chem. 1995;270:319 –324.
15. Zeytin FN, Rusk SF, Raymond V, Mandell AJ. Fibroblast growth factor stabilizes ribonucleic acid and regulates differentiated functions in a multipeptide-secreting neuroendocrine cell line. Endocrinology. 1988;122:1121–1128.
16. Lehr HA, Seemu¨ller J, Hu¨bner C, Menger MD, Messmer K. Oxidized LDL-induced leukocyte/endothelium interaction in vivo involves the receptor for platelet-activating factor. Arterioscler Thromb. 1993;13:1013–1018. 17. Watson AD, Navab M, Hama SY, Sevanian A, Prescott SM, Stafforini DM,
McIntyre TM, Du BN, Fogelman AM, Berliner JA. Effect of platelet acti-vating factor-acetylhydrolase on the formation of minimally oxidized low density lipoprotein. J Clin Invest. 1995;95:774 –782.
18. Hughes H, Mathews B, Lenz ML, Guyton JR. Cytotoxicity of oxidized LDL to porcine aortic smooth muscle cells is associated with the oxysterols 7-ketocho-lesterol and 7-hydroxycho7-ketocho-lesterol. Arterioscler Thromb. 1994;14:1177–1185. 19. Stiko A, Regnstro¨m J, Shah PK, Cercek B, Nilsson J. Active oxygen species
and lysophosphatidylcholine are involved in oxidized low density lipoprotein activation of smooth muscle cell DNA synthesis. Arterioscler Thromb Vasc
Biol. 1996;16:194 –200.
20. Ishii H, Kizaki K, Horie S, Kazama M. Oxidized low density lipoprotein reduces thrombomodulin transcription in cultured human endothelial cells through degradation of the lipoprotein in lysosomes. J Biol Chem. 1996;271: 8458 – 8465.
21. Boulanger C, Tanner FC, Be´a ML, Hahn AWA, Werner A, Lu¨scher TF. Oxidized low density lipoproteins induce mRNA expression and release of endothelin from human and porcine endothelium. Circ Res. 1992;70: 1191–1197.
22. Terkeltaub R, Banka CL, Solan J, Santoro D, Brand K, Curtiss LK. Oxidized LDL induces monocytic cell expression of interleukin-8, a chemokine with T-lymphocyte chemotactic activity. Arterioscler Thromb Vasc Biol. 1994;14: 47–53.
23. Khan BV, Parthasarathy SS, Alexander RW, Medford RM. Modified low density lipoprotein and its constituents augment cytokine-activated vascular cell adhesion molecule-1 gene expression in human vascular endothelial cells. J Clin Invest. 1995;95:1262–1270.
24. Fei H, Berliner JA, Parhami F, Drake TA. Regulation of endothelial cell tissue factor expression by minimally oxidized LDL and lipopolysaccharide.
Arterioscler Thromb. 1993;13:1711–1717.
25. Escargueil-Blanc I, Meilhac O, Pieraggi MT, Arnal JF, Salvayre R, Ne`gre-Salvayre A. Oxidized LDLs induce massive apoptosis of cultured human endothelial cells through a calcium-dependent pathway: prevention by aurin-tricarboxylic acid. Arterioscler Thromb Vasc Biol. 1997;17:331–339. 26. Thieme H, Aiello LP, Takagi H, Ferrara N, King GL. Comparative analysis
of vascular endothelial growth factor receptors on retinal and aortic vascular endothelial cells. Diabetes. 1995;44:98 –103.
27. Van Belle E, Rivard A, Chen D, Silver M, Bunting S, Ferrara N, Symes JF, Bauters C, Isner JM. Hypercholesterolemia attenuates angiogenesis but does not preclude augmentation by angiogenic cytokines. Circulation. 1997;96: 2667–2674.
28. Baffour R, Berman J, Garb JL, Rhee SW, Kaufman J, Friedman P. Enhanced angiogenesis and growth of collaterals by in vivo administration of recom-binant basic fibroblast growth factor in a rabbit model of acute lower limb ischemia: dose-response effect of basic fibroblast growth factor. J Vasc Surg. 1992;16:181–191.
29. Lazarous DF, Scheinowitz M, Shou M, Hodge E, Rajanayagam S, Hunsburger S, Robison WG Jr, Stiber JA, Correa R, Epstein SE, Unger EF. Effects of chronic systemic administration of basic fibroblast growth factor on collateral development in the canine heart. Circulation. 1995;91:145–153. 30. Folkman J, D’Amore PA. Blood vessel formation: what is its molecular
basis? Cell. 1996;87:1153–1155.
31. Maisonpierre PC, Suri C, Jones PF, Bartunkova S, Wiegand SJ, Radziejewski C, Compton D, McClain J, Aldrich TH, Papadopoulos N, Daly TJ, Davis S, Sato TN, Yancopoulos GD. Angiopoietin-2, a natural antagonist for Tie2 that disrupts in vivo angiogenesis. Science. 1997;277:55– 60.
32. Heery JM, Kozak M, Stafforini DM, Jones DA, Zimmerman GA, McIntyre TM, Prescott SM. Oxidatively modified LDL contains phospholipids with platelet-activating factor-like activity and stimulates the growth of smooth muscle cells. J Clin Invest. 1995;96:2322–2330.
33. Kugiyama K, Kerns S, Morrisett JD, Roberts R, Henry PD. Impairment of endothelium-dependent arterial relaxation by lysolecithin in modified low-density lipoproteins. Nature. 1990;344:160 –162.
34. Ohlsson BG, Englund MC, Karlsson AL, Knutsen E, Erixon C, Skribeck H, Liu Y, Bondjers G, Wiklund O. Oxidized low density lipoprotein inhibits lipopolysaccharide-induced binding of nuclear factor-kB to DNA and the subsequent expression of tumor necrosis factor-a and interleukin-1b in mac-rophages. J Clin Invest. 1996;98:78 – 89.
35. Ogita T, Tanaka Y, Nakaoka T, Matsuoka R, Kira Y, Nakamura M, Shimizu T, Fujita T. Lysophosphatidylcholine transduces Ca21signaling via the plate-let-activating factor receptor in macrophages. Am J Physiol. 1997;272(1 pt 2):H17–H24.