國 立 交 通 大 學
生物科技研究所
碩士論文
利用定點突變進行氧化鯊烯環化酵素家族
假設活性區中胺基酸之特性探討
Investigating the Characteristics of Putative
Active-Site Amino Acids in Oxidosqualene
Cyclases by Site-Directed Mutagenesis
研究生:陳奕齊
指導教授:吳東昆 博士
中華民國九十九年
Investigating the Characteristics of Putative Active-Site
Amino Acids in Oxidosqualene Cyclases by
Site-Directed Mutagenesis
研究生:陳奕齊
Student: Yi-Chi Chen
指導教授:吳東昆 博士
Advisor: Dr. Tung-Kung Wu
國立交通大學
生物科技研究所
碩士論文
A Thesis
Submitted to Department of Biological Science and Technology
College of Science
National Chiao Tung University
in partial Fulfillment of the Requirements
for the Degree of
Master
in
Biological Science and Technology
July, 2010
Hsinchu, Taiwan, Republic of China
I
利用定點突變進行氧化鯊烯環化酵素家族
假設活性區中胺基酸之特性探討
學生:陳奕齊
指導教授:吳東昆 博士
國立交通大學 生物科技研究所 碩士班
摘要
氧化鯊烯環化酵素存在於大多數生物體之中,他們會將直鏈狀的氧化鯊烯,反應 成特定的多環結構。在不同的生物物種,例如植物、動物與真菌間,會經由不同的反 應機制而形成不同的產物。催化過程包含氧化鯊烯上環氧基開環的起始反應、中途複 雜的環化和重組的步驟、以及最後的去質子反應,形成高度專一性的產物。為了比較 不同類型的氧化鯊烯環化酵素,我們選用了酵母菌中的氧化鯊烯-羊毛硬脂醇環化酵素、 阿拉伯芥中的環阿屯醇環化酵素及豌豆中的β-麥胚固醇環化酵素這三種來利用定點 突變的方式,分析存在於酵素活性區中的相對應胺基酸,希望能從實驗分析找出它們 的功能以及重要性。 在酵母菌的氧化鯊烯-羊毛硬脂醇環化酵素實驗中,我們利用定點飽和突變的方法, 針對 Cys703 來做分析。分析結果顯示經由不同的胺基酸突變,產生了八種不同的產 物,除了原先就會產生的羊毛硬脂醇之外,產生六種已知的環化中間物和一個先前未 曾被發現過的未知物。而其中之一的四環產物較為被關注,它在十七號碳的型態擁有 向上的氫和向下的長碳側鏈,與正常的結構相反,所以可以得知 Cys703 在決定十七 號碳上的側鏈為向上或向下型態佔有一定的重要性。此外,除了此產物以外,其他環 化中間物皆和 Phe699 突變點的分析十分類似,加上與離受質較遠的 Cys703 相比,II Phe699 為活性區第一層胺基酸且影響力大,可以推測 Cys703 和第一層胺基酸的穩 定性息息相關。 另一方面為阿拉伯芥中的環阿屯醇環化酵素及豌豆中的β-麥胚固醇環化酵素的 分析。前者與氧化鯊烯-羊毛硬脂醇環化酵素相同,在受質的摺疊會經由椅形-船形-椅 形形成原脂醇碳陽離子中間物;而後者則會經由椅形-椅形-椅形形成達瑪烯碳陽離子 中間物。此兩種酵素經由不同的結構和機制,分別形成環阿屯醇及β-麥胚固醇。實驗 分析方法是利用丙胺酸掃描法,選定十五個不同位置的胺基酸做突變,探討產物的差 異性。在環阿屯醇環化酵素的定點突變中,產生了一些和氧化鯊烯-羊毛硬脂醇環化酵 素突變中出現過的產物,推測是因為結構和機制非常的類似,因此儘管是不同物種, 但還是可以經由突變而產生相同的產物。而在β-麥胚固醇環化酵素的定點突變中,只 分析出兩種產物,推測可能是因為在β-麥胚固醇環化酵素這種椅形-椅形-椅形的機制 中需要比較精確的結構才可以使反應穩定,而詳細的反應機制則需要更進一步地針對 活性區周圍的胺基酸做深入的研究。
III
Investigating the Characteristics of Putative Active-Site Amino
Acids in Oxidosqualene Cyclases by Site-Directed Mutagenesis
Student: Yi-Chi Chen Advisor: Dr. Tung-Kung Wu
Institute of Biological Science and Technology National Chiao Tung University
Abstract
Oxidosqualene cyclases catalyze the biotransformation of the linear form substrate, (3S)-2,3-oxidosqualene, into tetracyclic or pentacyclic triterpene product. Different species of organisms including S. cerevisiae ERG7, A. thaliana CAS1 and P. sativum PSY operate this complex reaction through different conformation intermediates within the oxidosqualene cyclization process. According to previous reports, by utilizing the diverse structural and stereochemical control in various catalytically important amino acid residue mutants, oxidosqualene cyclases produced diverse product profiles ranging from monocyclic to polycyclic triterpene alcohols. These data implied that the plastic enzyme could thus be redesigned to obtain a novel reactivity from this complex enzyme, but with the characteristics of well-known high product specificity. Moreover, in order to further illustrate other critical amino acids involved in the catalytic significance and enzymatic plasticity of ERG7, CAS1 and PSY, we described herein a series of site-saturated mutations on
IV
the Cys703 residue of ERG7 and fifteen alanine-scanning mutations of CAS1 and PSY, respectively.
In the mutations of Cys703, a diverse products profile, including four known truncated cyclization tricyclic structures, four known tetracyclic structures and a novel product were identified from various ERG7C703X mutants. The product characterization and homology modeling results suggested that the Cys703 may indirectly affect the C-17 cation stabilization and the final deprotonation step, via the intermediary of the Phe699 residue, resulting in the production of either 17α or 17β side chain product derivatives. Structure-function-mechanism relationships of Cys703 and Phe699 on the catalytic activity of OSC could thus be discussed in a series of ERG7Phe699X/Cys703I double mutants.
In parallel, alanine-scanning mutagenesis was carried out on putative active site residues of CAS1 and PSY to further analyze the mutagenic effect on oxidosqualene cyclization in different sources. For mutation on CAS1, the product profiles of different mutants showed that there are highly relevant comparisons between CAS1 and ERG7. For example, the same bicyclic compound (9R,10S)-polypoda-8(26),13E,17E,21-tetraen-3β-ol was found in the CAS1Tyr734
mutant and its corresponding residue in the ERG7Tyr707 mutant. The results show that although they exist in totally different species, the relationships between them are still relevent. For the PSY mutation, there were only β-amyrin and lupeol synthesized from these mutants. This phenomenon allows us to predict that the producing of β-amyrin in PSY needs more precise structural control of substrate in the enzymatic active site. Thus, the detailed understanding of critical residues should be investigated after the functional roles of the neighboring amino acids in the active site are confirmed.
V
謝誌(Acknowledgement)
碩士班的生活終於到了尾聲了,當謝誌寫完,也就是我畢業的時候了吧!回過頭, 看著這兩年來的點點滴滴,從剛進實驗室什麼都不懂的小碩一開始,每件事都需要詢 問學長姊、常常做很多很天兵的舉動,漸漸的開始比較熟練、比較有自己的想法,到 現在看著學弟妹們努力的為實驗奮鬥。在這裡除了學習到很多的實驗技巧和學識外, 更多的東西是無法用文字表達的。因此,我在這要深深感謝每一位在身邊陪伴與教導 我的人,沒有你們,我絕對無法只靠自己走到這一步。這本論文除了呈現兩年來的成 果外,還有滿載的感謝與回憶。 最首先當然是要感謝我的指導教授吳東昆老師,感謝老師讓我加入這個實驗室, 提供完善的設備與環境,讓我能專心的做研究,除此之外,老師無論在做人處事的態 度、研究生應有的精神與能力及論文寫作上,都給我很好的建議與典範,雖然即將離 開實驗室,但我會時時記得老師說過的話,相信在接下來要開始的博士班生涯會有很 大的幫助。 接著,感謝李耀坤老師和廖奕翰老師,謝謝兩位老師在忙碌之餘尚能抽空來幫我 做口詴審核,給予我的論文許多良好建議,讓我注意到不足的地方,並能修改、補強 使論文內容更趨完善,老師精闢的見解也使我想到以往不曾思考過的問題。也感謝清 華大學貴儀中心的彭菊蘭女士,謝謝您在 NMR 光譜的分析上提供許多協助。 接下來是要感謝實驗室的成員們。首先要感謝的是 OSC 組的學長姊,我最敬佩 的媛婷學姊總是可以給我這麻煩的小朋友所提出的刁鑽問題一個完美的答覆,而且在 這兩年內實驗上有任何困難,都會給我些好點子。還有程翔學長,在碩二這年給了我 不少建議,尤其是在畢業前的這段日子,讓我可以順利考上台大,也順利畢業。接下 來是天昶學長,不論是實驗還是生活上,除了給了我很多新想法和建議外,在我心情 不好的時候也老是不厭其煩地聽我發牢騷。另外還有奕諄學姊,因為題目相似,不只 是讓我學到了很多實驗上的方法和技巧,還有很多的實驗結果可以參考。 再來是感謝實驗室其他組別的學長姊們。裕國學長上知天文下知地理,對於我不 擅長的非化學領域十分的在行,不管是問問題或是討論實驗總是可以令人豁然開朗。 豪哥學長在實驗室的儀器和環安上幫了我非常大的忙,而且每次出問題都會很熱心的 幫我解決。文鴻學長除了在我帶大學部生化實驗時幫了我很大的忙外,平常就像哥哥 一樣十分地照顧我,給了我很多很有用的觀念。晉源學長在電腦模擬方面幫了我很多, 不論是軟體的使用或是結果的判斷都是我不曾碰過的東西。還有小紅學姊,不管是實 驗問題、生活瑣事都常和我討論外,甚至是做事的觀念和態度都是我學習的目標。小 麵包學姊的實驗態度一直都是我所效法的對象,讓我了解想要有豐富的成果就要肯努 力地做實驗。而 Mili 學姊和 Allen 學長,讓我見識到了許多國外或是不同領域的見聞 和知識。 最後是感謝兩年來的夥伴們。同組的靜婷一直是我最好的拍檔,實驗上互相幫忙、 有問題一起煩惱,常常就這樣解決了很多困難。而書涵和小花不只是在課業上一起研VI 究,在平常生活上除了是需要幫忙時的好救兵外,也是聊聊天調濟心情的好對象。而 即將變成小老大的學弟妹們,首先感謝你們在準備我們口詴時花了不少心力。而對於 OSC 組的怡臻和欣怡,妳們總是為了我們組不辭辛苦的幫忙,感謝妳們默默地接受這 些不人道的虐待,是我能順利完成實驗的幕後功臣,OSC 組以後還要麻煩妳們了。世 穎身為唯一的碩班學弟,感謝你總是幫忙我處理些實驗室的工務雜務,日常生活上也 受到了很多的幫助。欣芳除了總是很溫柔地笑臉迎人、帶給實驗室不錯的氣氛外,也 是聊天的好對象。而在專題生小朋友方面,球球是我最感謝的人,最重要的是我實驗 上純化未知物的救星,而且在其他小事方面總是不辭辛勞地當我的小幫手。小彥除了 一手包辦了實驗室的歡笑,帶給實驗室很多的活力外,在生活上也幫了我不少忙。奕 汝和瑛婷對於實驗室的雜務幫忙了不少,另外妳們的實驗態度也是我一直在學習的地 方。而對於最近新進的碩一學弟妹和專題生,有你們的加入,讓實驗室充滿了新鮮和 活力的氣息,讓我做實驗的精神又為之一振。 當然也要感謝我的家人,總是提供我一個無後顧之憂的環境,並且在我的目標和 方向總是持續地支持,在我遇到困難時絕對會全力幫忙,另外在求學和實驗方面總是 給我很多的建議。 陳之藩的《謝天》一文結尾說到:「要感謝的人太多了,不如就感謝天吧!」我 想,這應該就是我現在心裡的最佳寫照吧!最後在這裡,再次衷心地感謝這兩年來在 我身邊的每一個人,並祝你們也天天精彩、豐富。
VII
Table of Contents
Abstract (Chinese)
... I
Abstract (English) ... III
Acknowledgement
... V
Table of Contents ... VII
List of tables
... XI
List of Figures ... XII
Chapter 1 Introduction ... 1
1.1 The importance of sterols ... 1
1.2 Terpenes, sterols and their biosynthetic pathway ... 1
1.3 Triterpene cyclases family ... 4
1.3.1 The hypothesis of oxidosqualene cyclase ... 6
1.3.2 Squlene-hopene cyclase (SHC) ... 9
1.3.3 Oxidosqualene-lanosterol cyclase (OSC) ... 10
1.3.4 Cycloartenol synthase (CAS) ... 16
1.3.5 β-amyrin synthase (βAS) ... 20
1.3.6 The amino acid sequence alignment ... 22
1.4 Research motive ... 25
1.4.1 The study of Cys703 in SceERG7 ... 25
1.4.2 The study of CAS1 in A. thaliana and PSY in P. sativum ... 28
Chapter 2 Materials and methods ... 30
VIII
2.1.1 Chemicals, reagents and kits ... 30
2.1.2 Bacterial, yeast strains and vectors ... 32
2.1.3 Equipments ... 32
2.1.4 Solutions and mediums ... 33
2.2 Methods ... 37
2.2.1 The construction of recombinant plasmids ... 37
2.2.1.1 Design primers: ... 37
2.2.1.3 Dpn I digest parental DNA template: ... 41
2.2.1.4 Transformation into XL1-Blue and enzyme digestion: ... 41
2.2.1.5 Sequencing analysis ... 42
2.2.2 Transformation and Genetic selections ERG7 mutants ... 42
2.2.2.1 Preparation of competent cell TKW14C2 ... 42
2.2.2.2 Transformation of mutated plasmid into TKW14C2 ... 43
2.2.2.3 Ergosterol supplement ... 43
2.2.3 Extracting lipids and characterizing mutant product profiles ... 44
2.2.3.1 Cell culture and extraction ... 44
2.2.3.2 Silica gel column chromatography ... 44
2.2.3.3 GC-MS column chromatography condition ... 44
2.2.4 Molecular modeling ... 45
Chapter 3 Results and Discussion ... 46
3.1 Functional analysis of S. cerevisiae ERG7 ... 46
3.1.1 Site-saturates mutagenesis of Cys703 ... 46
3.1.2 Product analysis ... 49
3.1.3 Proposed cyclization/rearrangement mechanism ... 51
IX
3.1.5 Product analysis of the double mutant of ERG7F699X/C703I ... 58
3.2 Functional analysis of CAS1 within A. thaliana ... 61
3.2.1 Site-directed alanine scanning mutagenesis ... 61
3.2.2 Product analysis ... 62
3.2.3 Proposed cyclization/rearrangement mechanism ... 63
3.3 Functional analysis of P. sativum PSY ... 66
3.3.1 Site-directed alanine scanning mutagenesis ... 66
3.3.2 Experimental results and phenomena ... 67
Chapter 4 Conclusions
... 71
4.1 Analysis of S. cerevisiae ERG7C703X mutations ... 71
4.2 Analysis of A. thaliana CAS1 mutations ... 72
4.3 Analysis of P. sativum βAS mutations ... 73
Chapter 5 Future prospects ... 74
Appendix ... 75
A.1 Enhance the protein expression by change the vectors ... 75
A.1.1. Construction of the plasmids ... 75
A.1.2. Integrate the genes into the chromosome of TKW14C2 ... 76
A.1.3. Result and discussion ... 77
A.1.4. Future works ... 78
A.2 Construct erg7/erg11 double knockout strain ... 79
A.2.1. The sterol metabolic pathway within S. cerevisiae ... 79
A.2.2. The non-specificity of sterol metabolic enzymes ... 80
X
A.2.4. Result and discussion ... 82 A.2.5. Future works ... 82
XI
List of tables
Table 1.1 Product profiles of AthCAS Ile481, Tyr410 and His477 mutants. ... 18
Table 2.1 QuickChange Site-Directed Mutagenesis Kit PCR composition ... 41
Table 2.2 QuickChange Site-Directed Mutagenesis PCR program ... 41
Table 3.1 The site-saturated mutants of S.c ERG7C703X and their genetic analysis. .. 48
Table 3.2 The product analysis of S. cerevisiae ERG7 C703mut ... 50
Table 3.3 the product pattern of C703, F699 and I705 in S. cerevisiae ... 58
Table 3.4 Product analysis of S. cerevisiae ERG7F699X/C703I double mutants ... 59
Table 3.5 The genetic analysis of A. thiliana CASmut. ... 61
Table 3.6 The product analysis of A. thiliana CASmut. ... 62
Table 3.7 The product analysis of AthCAS mutants compare to their correspounding residue mutants in SceERG7 ... 64
Table 3.8 The genetic analysis of P. sativum PSYmut. ... 66
Table 3.9 The product analysis of P. sativum PSYmut. ... 67
Table 3.10 Compare the product quantity between PSY and CAS1. ... 70
Table A.1 genomic PCR composition ... 77
XII
List of Figures
Figure 1.1 The sterol biosynthetic pathway. ... 3
Figure 1.2 Cyclization of (A) dammarenyl cation, (B) protosteryl cation. ... 5
Figure 1.3 The proposed enzyme models by Johnson ... 8
Figure 1.4 Griffin’s hypothesis model for involvement of electron-rich aromatic side chains from Trp and Tyr residues in the cyclization of oxidosqualene to the protosteryl cation. ... 8
Figure 1.5 Crystal structure of A. acidocaldarius squalene-hopene cyclase. ... 9
Figure 1.6 The cyclization process of squalene-hopene cyclase. ... 10
Figure 1.7 The mechanism of conversion of oxidosqualene into lanosterol. ... 11
Figure 1.8 The proposed model for oxirane ring opening and cyclization initiation. .. 11
Figure 1.9 Substrate analogue and their product derovatoves during the oxidosqualene cyclase cyclization which are suggestive of five-membered C-ring intermediate. ... 12
Figure 1.10 Proposed mechanisms for C-ring expansion and D-ring formation. ... 13
Figure 1.11 The previous study proposed an incorrect C17α stereochemistry of protosteryl cation intermediate required a large side-chain rotation prior to rearrangement to account for the observed stereochemistry at C20. .. 14
Figure 1.12 The evidence for the stereochemistry of protosteryl cation intermediate at C17 prefer a β- rather than an α- orientation at C-17 position by incubation of substrate analogue with OSC. ... 14 Figure 1.13 Human OSC structure. (a) The ribbon diagram of human OSC structure.
The C and N termini and several sequence positions are labeled. The inner barrel helices are colored yellow. The bound inhibitor, Ro48-8071
XIII
(black), indicates the location of the active site. (b) The orientation of OSC is shown relative to one leaflet of the membrane, and Ro48-8071 binds in the central active-site cavity. ... 15 Figure 1.14 The difference between cyclization mechanisms of lanosterol synthase
and cycloartenol synthase. ... 16 Figure 1.15 Conservation pattern between CAS1 and ERG7... 17 Figure 1.16 Products formed by cycloartenol synthase mutants. ... 18 Figure 1.17 Proposed mechanism of 2,3-oxidosqualene into β-amyrin and lupeol. .. 20 Figure 1.18 Amino acids alignment. The red words are the CAS1mut and the green
one is ERG7C703 mutation. ... 24 Figure 1.19 The position of Cys703 in the active-site within SceERG7. ... 26 Figure 1.20 The alanine scanning mutation position in the active-site of β-amyrin
synthase within Pisum sativum. ... 29 Figure 3.1 The earlier stage analysis by ergosterol supplement in TKW14C2 strains.
... 47 Figure 3.2 Strategies of product purification and analysis. ... 49 Figure 3.3 Proposed cyclization/rearrangement pathway occurred in the ERG7C703X
site-saturated mutants. ... 53 Figure 3.4 Homology model of Cys703 complex with Phe699, Ile705 and lanosterol.54 Figure 3.5 Homology model of Cys703 mutant complex with Phe699, Ile705 and
lanosterol. The green one is the wild type ERG7, and the blue one is the mutant. (A) Cys703Tyr. (B) Cys703Arg. ... 55 Figure 3.6 Homology model of Cys703 mutant complex with Phe699, Ile705 and
lanosterol. The green one is the wild type ERG7, and the yellow one and the orange one are the mutants. (A) Cys703Ile. (B) Cys703Met. ... 56 Figure 3.7 Proposed mechanism of the production of the 17α product. ... 57
XIV
Figure 3.8 Homology model of double mutant complex with lanosterol. The green one is the single mutation on Cys703, and the blue one is the
F699A/C703I double mutants. ... 60 Figure 3.9 Proposed cyclization/rearrangement pathway occurred in CAS1mut ... 63 Figure 3.10 Homology model of F445, F699 and Y707 in SceERG7 mutant complex
with lanosterol. ... 65 Figure 3.11 Proposed cyclization/rearrangement pathway occurred in PSYmut ... 68 Figure 3.12 Phe552, Tyr259 and Met729 complex with β-amyrin. ... 69 Figure 3.13 Homology model of L734, Y736 and Y739 in PSY complex with
β-amyrin. ... 70 Figure A.1 Sterol metabolism pathway ... 81
1
Chapter 1 Introduction
1.1
The importance of sterols
Sterols play important roles in living cells. They usually process a tetracyclic or pentacyclic ring structure, and they are the product of the cyclization of triterpenoids. Moreover, they are known as steroid alcohols with a hydroxyl group at the C-3 position of the A-ring.
Cholesterol is the most important sterol in vertebrates. It is a primary component of the cell membranes and serves as an intermediate in the biosynthesis of most of the important steroids. Therefore, it is essential to life. Cholesterol circulates in the bloodstream and is synthesized by the liver and several other organs. However, high levels of blood cholesterol have been implicated in the development of arteriosclerosis and in heart attacks that occur when cholesterol-containing plaques block arteries supplying the heart, so some treatments to reduce blood cholesterol are usually required.
Steroid derivatives have various functions as well. For example, many higher plants contain steroid saponins and sapogenins. Diosgenin, one of the steroid sapogenins, is the starting material of industrial interest in the synthesis of several steroids that are used as anti-inflammatory, androgenic, estrogenic, and contraceptive drugs on the market. Furthermore, these compounds are reported to possess cytotoxic, antitumor, antifungal, immunoregulatory, hypoglycemic, and cardiovascular properties.1
1.2
Terpenes, sterols and their biosynthetic pathway
2
molecular formula C5H8. The basic molecular formula of terpenes are multiples of
the isoprene unit (C5H8)n, where n is the number of linked isoprene units.
Triterpenes are derived from six isoprene units, and have a formula C30H48. In some
biosynthetic pathways, they are converted into triterpenoids, a family of oxygen-containing compounds. For example, the acyclic triterpene, squalene, is an important metabolic precursor of sterols.
The sterols biosynthesis process is generally through the mevalonic acid (MVA) pathway. Acetyl-CoA (C2) is condensed into acetoacetyl-CoA (C4) with another
acetyl-CoA subunit via the acetyl-CoA transferase triggered reaction. By 3-hydroxy-3-methylglutaryl-CoA (HMG-CoA) synthase, acetyl-CoA condenses with acetoacetyl-CoA into HMG-CoA (C6). HMG-CoA is converted into mevalonate (C6),
after the catalysis of HMG-CoA reductase. Mevalonate is phosphorylated by three sequential Pi transferred from ATP, yielding 3-phospho-5-pyrophosphate (C6).
Moreover, by undergoing decarboxylation, coupled with dehydration, isoprenoid pyrophosphate (C5) is generated. Isopentenyl pyrophosphate is referred to
isoprenoid in the pathway. By a series of chemical reactions, including isomerization and condensation, a linear molecule, squalene (C30), is produced.
In bacteria, squalene is made via different compounds via various enzymatic reactions. Squalene is largely converted to hopene by squalene-hopene synthase, and can be catalyzed into hopanol or diplopterol via other enzymes. However, in most eukaryotes, the oxidation of squalene by squalene synthase yields an acyclic polyolefin substrate, (3S)-2,3-oxidosqualene (OS), which is the precursor of steroids.
In higher plants, the cyclization of OS can be transformed into cycloartenol, lupeol, and β-amyrin by cycloartenol synthase, lupeol synthase, and β-amyrin synthase, respectively. Furthermore, the OS can also convert into lanosterol by
3
oxidosqualene-lanosterol cyclase, in fungi and animals. After a series of cyclization and/or rearrangement cascades, depending on the forming species, the steroids are formed with diverse structures (Fig. 1.1).
HO Ergosterol Cholesterol Cycloartenol Lanosterol -Amyrin Lupeol Hopene HO HO HO HO HO O (3S)2,3-Oxidosqualene squalene Acetyl-CoA Farnesyl pyrophosphate Fungi Animals Higher plants Higher plants Higher plants Bacteria
Fungi & Animals
Oxidosqualene-Lanosterol Cyclase Cycloartenol Synthase -Amyrin Synthase Lupeol Synthase Squalene-Hopene Cyclase Squalene epoxidase Squalene synthaseERG9
ERG1
ERG7
4
1.3
Triterpene cyclases family
The triterpenoids are a large and structurally diverse group of natural products derived from squalene or other acyclic 30-carbon precursors.2 The unusually complex reaction mechanism and flexible reactant structure generates these ring systems and results in over 100 distinct steroid skeletons. Enzymes that catalyze these reactions are known as triterpene synthases and can be categorized as squalene cyclases or oxidosqualene cyclases, which convert squalene and oxidosqualene to diverse sterols. The enzymatic cyclization of squalene or oxidosqualene to polycyclic triterpenoids is one of the most complicated biochemical reactions to capture the interest of organic chemists and biochemists for over half a century.
There is a general mechanism among these enzymes, involving: 1. Binding of the polyolefinic substrate in a folded conformation.
2. Initiation of the reaction by protonation of a double bond (in squalene scaffold) or an epoxide (as shown in OS scaffold).
3. Ring formation.
4. Skeletal rearrangement by methyl and hydride shifts in some cases. 5. Termination by deprotonation or addition of water.3
However, there are still many differences in their mechanisms. The enormous skeletal diversity found among the triterpenes is the result of different cyclization reactions that lead to varied carbocyclic skeletons, following by different methyl and hydride shifts and a different cation-quenching step.
The substrate in these enzymes is pre-folded into different conformations. In squalene-hopene cyclase SHC, the binding substrate squalene is only pre-folded into the all-chair conformation. However, there are two folding types within oxidosqualene cyclases, chair-chair-chair and chair-boat-chair conformations,
5
respectively.
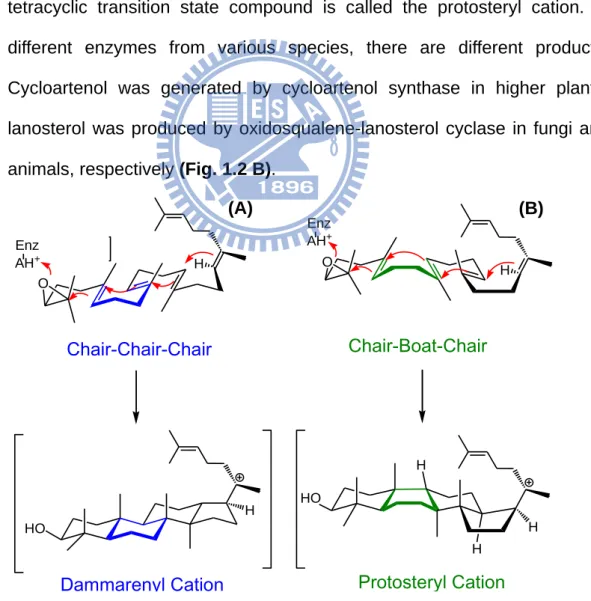
1. Chair-chair-chair conformation:
This conformation has been found in higher plants, including lupeol synthase and β-amyrin synthase. The B-ring was pre-folded in chair form. When cyclization occurs, OS is converted into dammarenyl cation first. After the entire reaction, it generates the product, lupeol or β-amyrin (Fig. 1.2 A). 2. Chair-boat-chair conformation:
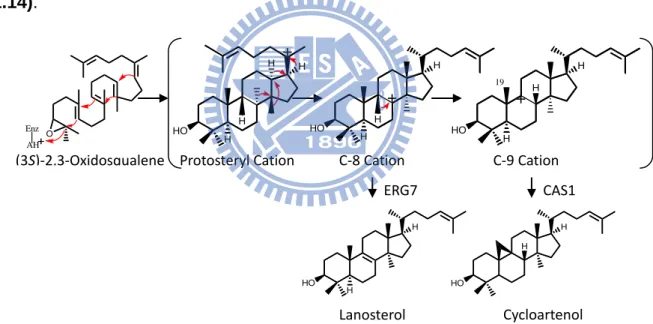
It is another kind of substrate folding manner when the B-ring is pre-folded in the boat form in the enzymatic active site during the cyclization process. The tetracyclic transition state compound is called the protosteryl cation. In different enzymes from various species, there are different products. Cycloartenol was generated by cycloartenol synthase in higher plants; lanosterol was produced by oxidosqualene-lanosterol cyclase in fungi and animals, respectively (Fig. 1.2 B).
HO H H HO O Enz AH+ O Enz AH+ Chair-Chair-Chair Dammarenyl Cation Chair-Boat-Chair Protosteryl Cation H H H H
Figure 1.2 Cyclization of (A) dammarenyl cation, (B) protosteryl cation.
6
1.3.1
The hypothesis of oxidosqualene cyclase
The family of oxidosqualene cyclases extensively exists in organisms. These different enzyme systems convert the oxidosqualene, as a substrate, into various polycyclic triterpenes. The relationship between the enzyme structure and cyclization mechanism is extremely interesting and diverse.
Woodward and Bloch first proposed in 1953 that in the cholesterol biosynthesis pathway, lanosterol was produced after the cyclization of squalene, followed by rearrangement.4 In 1958 and 1965, Maudgal and Cornforth groups provided evidence for 1.2-methyl and hydride shifts during lanosterol formation by incorporation experiments.5-6 Corey and van Tamelen showed that 2,3-oxidosqualene is more efficiently incorporated in sterol synthesis than squalene, the demonstrated intermediacy of 2,3-oxidosqualene in lanosterol biosynthesis.7-8 In addition, van Tamelen also showed that the nonenzymatic cyclization of 2,3-oxidosqualene resulted in truncated cyclization to produce a tricyclic product, suggesting that direct enzymatic control is necessary for the prevention of the chemical tendency in the formation of the five-membered C-ring, and for emergence of the biologically required six-membered C-ring.9 In 1975, Barton confirmed that eukaryotic oxidosqualene cyclases accepted only (3S)-, but not (3R)-, enantiomers of 2,3-oxidosqualene as a substrate in the formation of lanosterol, demonstrating the highly substrate-specific property of oxidosqualene cyclases compared to squalene cyclases.10 Guy Ourisson and his co-workers proposed the possible molecular evolution from the primitive squalene cyclases to oxidosqualene cyclases in higher organisms.11 Furthermore, Corey and Matsuda showed that oxirane cleavage and cyclization of the A-ring are concerted and essential for electrophilic activation of the oxirane function.12-13
In previous research, oxidosqualene was stable in the neutral condition at room temperature for the whole day. A stronger acid such as trichloroacetic acid is
7
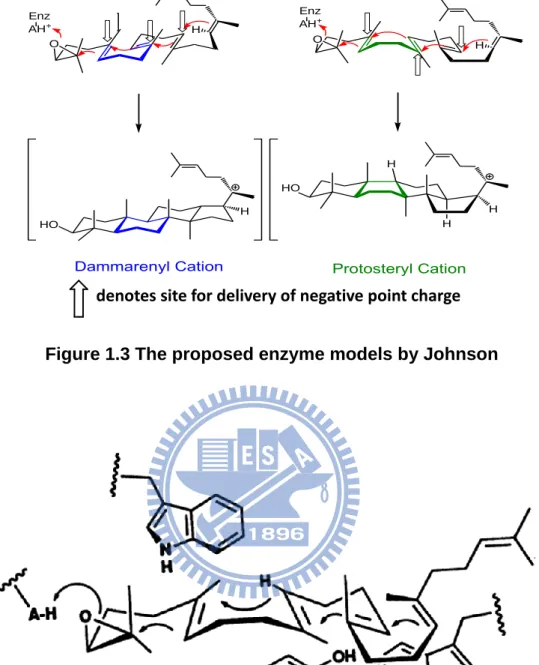
required for epoxide ring activation to promote the cyclization step.14 Then, many unstable cation intermediates were thus formed in the cyclization process. Two hypotheses that showed how the enzyme worked to stabilize the high-energy cation intermediates were proposed. In 1987, Johnson proposed the “cation-stabilizing auxiliary” model which supposed that the Lewis acid residues on the active site provide a proton for initiation of the epoxide group, and a number of anionic sites in the cyclase enzyme which further led to the cation generation and the formation of the proper ring system. These axial negative charge residues would face toward the transition states or the intermediates for the cation stabilization, and also facilitate the ring formation of B-boat/chair ring. Therefore the B-boat ring could be promoted by the delivery of a point charge to the α-face at pro-C-8, and lowering the activation energy of the boat form rather than that of chair skeleton (Fig. 1.3).15-16 In addition, Griffin and co-workers proposed the “aromatic hypothesis” model from which the electron-rich aromatic side chain, such as Trp and Tyr, might stabilize the positively charged transition states or high-energy intermediates during the process of cyclization and rearrangement steps (Fig. 1.4).17 In this hypothesis, the aromatic residues play the role of the anions group, just like that proposed in the “Johnson model”. These cation-π interactions are common features in enzyme-substrate complexes.
8
Figure 1.4 Griffin’s hypothesis model for involvement of electron-rich aromatic side chains from Trp and Tyr residues in the cyclization of oxidosqualene to the protosteryl cation.
Figure 1.3 The proposed enzyme models by Johnson
HO H H HO O Enz AH+ O Enz AH+
Dammarenyl Cation Protosteryl Cation
H
H
H
H
9
1.3.2
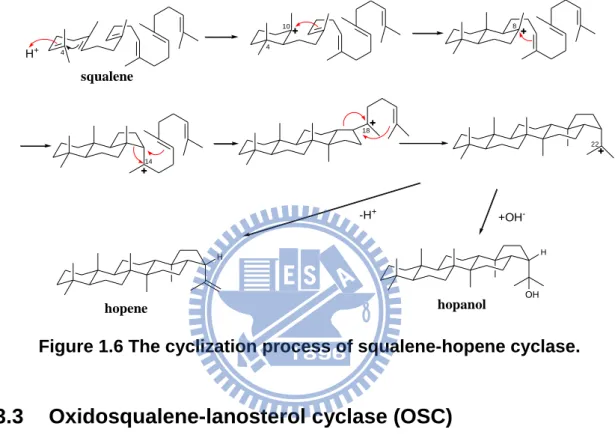
Squalene-hopene cyclase (SHC)
Squalene-hopene cyclase (EC 5.4.99.17) is a homodimeric enzyme which is organized in two groups of α-helical domains, which together form a dumbbell-shaped molecule (Figure 1.5). It contains 631 amino acids per subunit, and the molecular masses are about 71.5 kDa each.
In 1997, the X-ray crystal structure of Alicyclobacillus acidocaldarius SHC was first reported at a 2.9 Å resolution, later refined to 2.0 Å resolution, in 1999.18-19 Moreover, Reinert et al. reported another X-ray crystal structure from which the squalene-hopene cyclase was cocrystalized with 2-azasqualene and its resolution is 2.13 Å in 2004.20 These structures combined with the biological studies, provided a more mechanistic insight into squalene-hopene cyclases and oxidosqualene cyclases.
Squalene-hopene cyclase converts squalene to the pentacyclic hopene skeleton in prokaryotes. It binds squalene in the all-chair conformation, and initiates the cyclization cascade by protonating the terminal double bond. The cyclization reaction produces the 6.6.6.6.5-fused pentacyclic hopanyl C-22 cation, which Figure 1.5 Crystal structure of A. acidocaldarius squalene-hopene cyclase.
10
undergoes either proton elimination or addition of water to produce a 5:1 mixture of hopene and hopanol without the rearrangement step (Figure 1.6).21 However, the bacterial SHCs displayed very low substrate specificity. They can cyclize not only the natural substrate, but both enantiomers of oxidosqualene and regular polyprenols.
1.3.3
Oxidosqualene-lanosterol cyclase (OSC)
Oxidosqualene-lanosterol cyclase (EC 5.4.99.7), also called lanosterol synthase, is widely found in animals and fungi. It converts oxidosqualene into the tetracyclic product, lanosterol. Lanosterol is the precursor of ergosterol in fungi and cholesterol in animals, both them are important components of the cell membrane.
In Saccharomyces cerevisiae, cyclases are encoded from erg7, as a membrane protein. The protein contains 731 amino acids and the molecular mass is approximately 83 kDa. Because of the difficulty in purifying membrane proteins, the crystal structure of S. c oxidosqualene-lanosterol cyclases (SceERG7) enzyme has not been possible until recently. In order to understand the structure-function relationships of oxidosqualene cyclase-catalyzed reactions in depth, site directed /
4 squalene H+ 4 10 14 8 18 22 H H OH hopene hopanol +OH --H+
11
saturated mutagenesis, coupled with genetic selection and products analysis, for illustrating the functional importance of these mutated residues, participating in the carbon cationic intermediates stabilization of the reaction cascade were carried out.
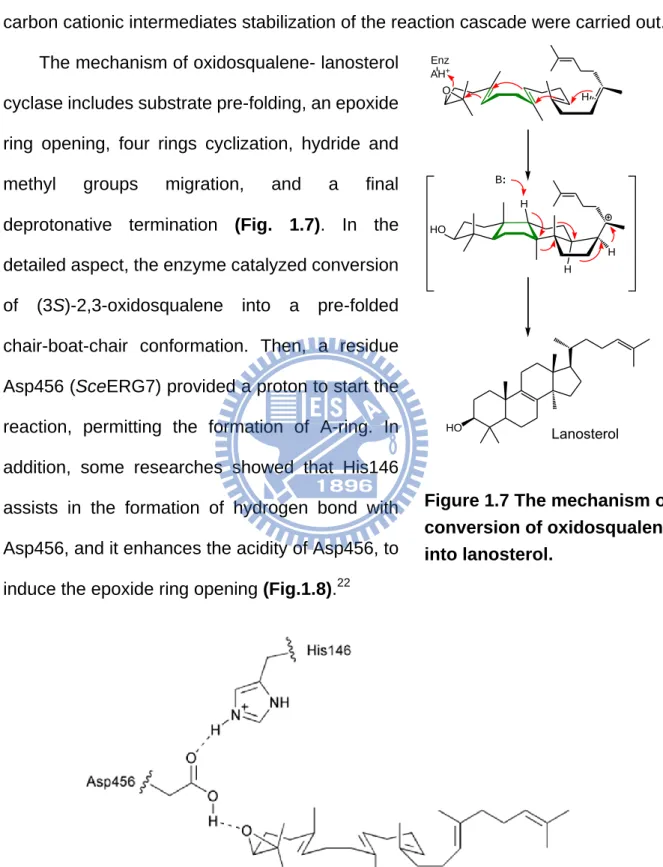
The mechanism of oxidosqualene- lanosterol cyclase includes substrate pre-folding, an epoxide ring opening, four rings cyclization, hydride and methyl groups migration, and a final deprotonative termination (Fig. 1.7). In the detailed aspect, the enzyme catalyzed conversion of (3S)-2,3-oxidosqualene into a pre-folded chair-boat-chair conformation. Then, a residue Asp456 (SceERG7) provided a proton to start the reaction, permitting the formation of A-ring. In addition, some researches showed that His146 assists in the formation of hydrogen bond with Asp456, and it enhances the acidity of Asp456, to induce the epoxide ring opening (Fig.1.8).22
Figure 1.8 The proposed model for oxirane ring opening and cyclization initiation. HO H H HO O Enz AH+ Lanosterol H H B:
Figure 1.7 The mechanism of conversion of oxidosqualene into lanosterol.
12
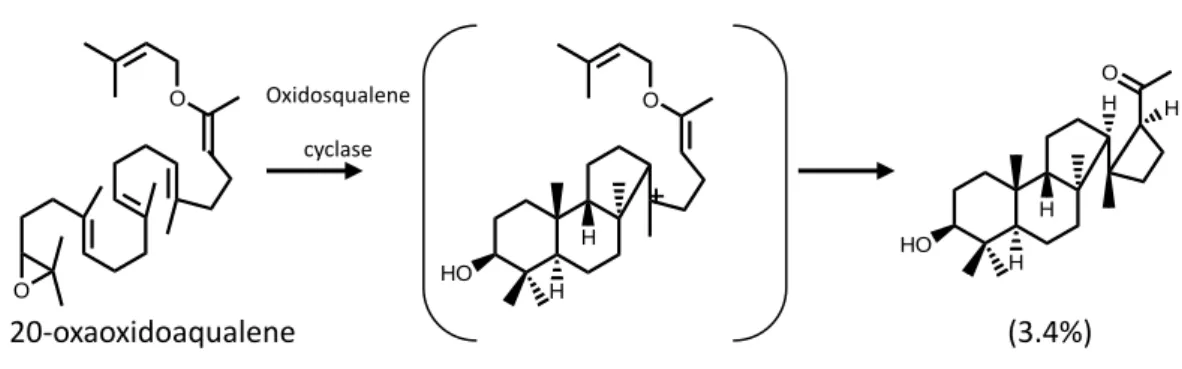
The formation of the B-boat ring probably occurs very rapidly when the positive charge appears on the C-6 of the A-ring. The closure of the C-ring is puzzling because the direct formation of the six-membered C-ring would represent the anti-Markovnikov form. In 1995, E. J. Corey and his co-workers first proposed the five-membered C-ring Markovnikov cyclization against the direct formation of a six-membered C-ring. By using oxidosqualene analogues, 20-oxaoxidosqualene was reacted with oxidosqualene cyclase, and the resulting products suggested that there was a five-membered C-ring intermediate appearing through an intramolecular reaction (Fig. 1.9).23 Moreover, incubation of additional substrate analogues within oxidosqualene cyclases produced 6-6-5 cyclization products, providing more evidence for a five-membered C-ring closure followed by a ring expansion.24 In the theoretical calculation, Hess indicated that the 6-6-5-tricyclic cation was the first intermediate during the formation of the protosteryl cation, and five-membered C-ring expansion and formation of the D-ring were concerted (Fig. 1.10).25 In comparison, for the SHC study, Gao showed no and fewer intermediates of tricyclic and tetracyclic cations, suggesting that the formation of C, D and E-ring were concerted.26 O O 20-oxaoxidoaqualene HO H H O HO H H O H H (3.4%) Oxidosqualene cyclase
Figure 1.9 Substrate analogue and their product derivatives during the oxidosqualene cyclase cyclization which are suggestive of
13
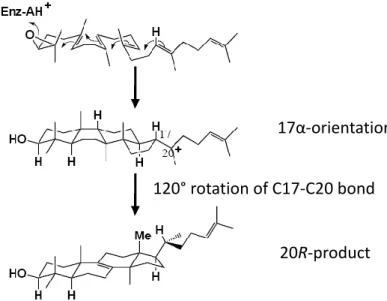
Early studies of the sterol biosynthesis assumed that the conversion of oxidosqualene into lanosterol proceed via a protosteryl cation where its cationic sidechain at C-17 is α-oriented.27 After a hydride migration of this 17α-protosteryl
cation from C-17 to C-20, an un-natured 20S configuration, but not the natural 20R skeleton product, was thus generated. Another exception for the mechanistic illustration was suggested to explaining the backbone rotation at C20, where a 120° rotation of C17-C20 bond occurs via the hydride migration in order to produce the 20R skeleton of lanosterol, whereas only 60° rotation is required to produce the unnatural 20S configuration (Fig. 1.11).28 In recent years, it had been proposed that the 17α-protosteryl cation may not be the intermediate. In 1992, Corey proposed the evidence that stereochemistry at C-17 of the protosteryl cation prefers a β- rather than an α- orientation by the enzyme-catalyzed cyclization of 20-oxaoxidosqualene (Fig. 1.12). This result overthrows the assumption that the large rotation around C17-C20 bond occurs prior to the actual rearrangement.24,28 Moreover, the 17β-protosteryl cation was also demonstrated by the catalysis of
Figure 1.10 Proposed mechanisms for C-ring expansion and D-ring formation.
14
20,21-dehydrooxidosqualene.29
The final step of lanosterol synthesis is the rearrangement of the protosteryl cation. After two hydride and two methyl groups shift (H-17α→20, H-13α→17α, CH3-14β→13β, CH3-8α→14α), this is followed by deprotonation on C-8 / C-9 to
Figure 1.11 The previous study proposed an incorrect C17α stereochemistry of protosteryl cation whereby the intermediate required a large side-chain rotation prior to rearrangement to account for the observed stereochemistry at C20.
17α-orientation
20R-product 120° rotation of C17-C20 bond
Figure 1.12 The evidence for the stereochemistry of protosteryl cation intermediate at C17 preferring a β- rather than an α- orientation at C-17 position by incubation of substrate analogue with OSC.
15
generate the product lanosterol.
In 2004, Thoma and co-workers solved the X-ray structure of human OSC which is in complex with lanosterol.30 Human OSC is a monomer that consists of two barrel domains that are connected by loops and three smaller β-structures, and the large active-site cavity is located in the center of the molecule between domains 1 and 2. It is a monotopic membrane protein that attached to the membrane from one side, and the membrane-inserted surface consists of a plateau with 25 Å in diameter and a channel that leads to the active-site cavity. The channel is considered to permit oxidosqualene, as a substrate, to enter the hydrophobic active site and have the separated role from that of the putative active-site cavity. Achievement of the substrate passage is constructed either by a change in the side chain of the residue such as Tyr237, Cys233 and Ile524 or by rearrangement of the strained loops from 516 to 524 and from 697 to 699 (Fig. 1.13). Because of the human OSC crystal structure, the relationship between the functional residues and cyclization mechanism could be understood in depth.
Figure 1.13 Human OSC structure. (a) The ribbon diagram of human OSC structure. The C and N termini and several sequence positions are labeled. The inner barrel helices are colored yellow. The bound inhibitor, Ro48-8071 (black), indicates the location of the active site. (b) The orientation of OSC is shown relative to one leaflet of the membrane, and Ro48-8071 binds in the central active-site cavity.
16
1.3.4
Cycloartenol synthase (CAS)
Cycloartenol synthase (EC 5.4.99.8) is the sterol precursor in higher plants and its skeleton is similar to lanosterol. The reaction of converting oxidosqualene into cycloartenol is very similar to that catalyzed by lanosterol synthase. Moreover, cycloartenol synthase has 759 amino acids and the molecular mass is approximately 86 kDa. Although there are 400 amino acids that are different, shown in the results of sequence alignment between CAS and OSC, all steps in cycloartenol and lanosterol synthase are identical with the exception of the final deprotonation reaction. Cycloartenol synthase forms the cyclopropyl ring and abstracts a proton from C-19, whereas lanosterol synthase removes a different proton and forms lanosterol (Fig. 1.14).
Phytosterols, such as campesterol and sitosterol, are biosynthesized from cycloartenol. Phytosterols, also called as plant sterols, are a group of naturally occurring steroid alcohols synthesized by plants. In addition to cholesterol-lowering effects, phytosterols have been suggested to possess anti-inflammatory, antibacterial, antifungal, antiulcerative, and antitumor activities.31
Cycloartenol synthase was first cloned from Arabidopsis thaliana (AthCAS1), and
(3S)-2,3-Oxidosqualene O Enz AH H HO H H H Protosteryl Cation H HO H H C-8 Cation H HO H H 19 C-9 Cation H HO H Lanosterol H HO H Cycloartenol ERG7 CAS1
Figure 1.14 The difference between cyclization mechanisms of lanosterol synthase and cycloartenol synthase.
17
expressed and characterized in the yeast lanosterol synthase system in 1993.32 Furthermore, because the cyclopropyl ring formation in the cycloartenol biosynthesis is thermodynamically unfavorable relative to that of the lanosterol formation, some of amino acid differences are probably specifically required for inducing cyclopropyl ring formation and excluding the formation of more energetically favored products by the cycloartenol synthase. The site-direct mutagenesis study of cycloartenol synthase (AthCAS1) showed some important residues such as Tyr410, His477, and Ile481. 33 These residues are highly-conserved within cycloartenol synthase in many plant species, but they maintain Thr, Cys or Gln, and Val in the corresponding positions of ERG7 from animals and fungi (Fig. 1.15). Speculation on this phenomenon, these residues may promote the formation of cyclopropyl rings within AthCAS1; in other words, mutations at these positions may permit the lanosterol formation (Table 1.1).
Ile481 is conserved in all cycloartenol synthases, whereas Val is present in lanosterol synthase at this position. The γ-methyl of Ile481 might promote cycloartenol formation by preventing the rotation of the intermediate cation through steric interactions with C-2 and the two axial methyl groups at the A-ring. Removing the γ-methyl group with an Ile481Val substitution resulted in 25% lanosterol production in addition to production of cycloartenol and parkeol. In addition, Ile481 may also be involved in assisting in proper substrate folding, as well as for the cyclization reaction. Mutation of Ile481 to smaller residues such as Ala and Gly has led to achilleol A and camelliol C production (Fig. 1.16).34
Figure 1.15 Conservation pattern between CAS1 and ERG7 AthCAS1 QGYNG 412 TADHGWPISDCT 485
DdiCAS1 QGYNG 365 TVDHGWPISDCT 437 SceERG7 MGTNG 386 TKTQGYTVADCT 458 SpoERG7 RGTNG 381 NITQGYTVSDTT 453 HsaERG7 QGTNG 383 TLDCGWIVSDCT 457 RnoERG7 QGTNG 384 TLDCGWIVADCT 458
18
Figure 1.16 Products formed by cycloartenol synthase mutants. AthCAS1mut Cycloartenol Lanosterol Parkeol 9β-lanosta-
7,24-dien-3β-ol Achilleol A Camelliol C
Wild type 99 1 I481L 83 1 16 I481V 54 25 21 I481A 12 54 15 13 6 I481G 17 23 4 44 12 Y410T 65 2 33 H477N 88 12 H477Q 22 73 5 I481V/Y410T 75 1 24 I481V/H477N/Y410T 78 22 I481V/H477Q/Y410T 78 22 I481V/H477N 99 1
Table 1.1 Product profiles of AthCAS Ile481, Tyr410 and His477 mutants.
Lanosterol HO O (3S)2,3-Oxidosqualene Cycloartenol 9β-lanosta-7,24-dien-3β-ol HO HO Achilleol A HO Camelliol C H Parkeol HO HO HO Protosteryl cation H H H
19
Tyr410 and His257 participate in an H-bonding network positioned near the C-19 methyl group for the deprotonation reaction.33 Tyr410 is present in all cycloartenol synthases, but animal and fungal oxidosqualene-lanosterol synthase maintain Thr at the corresponding position. The AthCAS1Tyr410Thr mutant forms 65% lanosterol, along with 9β-lanosta-7,24-dien-3β-ol and parkeol. Removing the aromatic ring of Tyr410 decreases the steric bulk above the intermediate cation. Because the hydroxyl group in Thr is closer to its α-carbon than that in Tyr, the polar groups of Tyr410Thr, Tyr532, and His257 were repositioned in the Tyr410Thr mutant. This combination of steric and electronic changes abolishes the cycloartenol synthesis and allows deprotonation of
C-8/C-9 lanosterol cation to form lanosterol, parkeol, and
9β-lanosta-7,24-dien-3β-ol.35
His477 is not in the active site, but is a second-sphere residue that affects the product profile through the interactions with the side chain of Tyr410.33 His477 is strictly conserved in the known cycloartenol synthase, whereas lanosterol synthases maintain either Gln or Cys. The AthCAS1His477Gln mutant has the polar functionality moved toward C-11, consequently resulting in more parkeol production than lanosterol. The AthCAS1His477Asn mutant formed lanosterol by positioning the basic group near the C-9/C-8, but also produced parkeol due to a close enough distance to C-11.36
The double mutant of CAS1I481V/ Y410T formed lanosterol more accurately than either single mutant alone. However the triple mutant (His477Asn/Gln, Ile481Val, and Tyr410Thr) did not promote the lanosterol synthesis because the hydroxyl group of Thr is too distant to interact with the amide group of Asn or Gln residues. The His477Asn Ile481Val double mutant is the most accurate example for the enzyme mutation to generate lanosterol.36
20
1.3.5
β-amyrin synthase (βAS)
β-amyrin synthase originates from several plant species, such as Pisum sativum and Panax ginseng, and it is also a member of phytosterols biosynthesis pathway. The biosynthesis mechanism of β-amyrin is much different from that of lanosterol and cycloartenol. In the beginning, oxidosqualene is pre-folded into a chair-chair-chair conformation, and then forms the dammarenyl cation intermediate. Next, the D-ring expansion and E-ring cyclization occur, producing the 6-6-6-6-5 pentacyclic lupenyl cation. The lupenyl cation then converts to lupeol by deprotonation by lupeol synthase. But for β-amyrin synthase, the E-ring expands to continue the rearrangement cascade. Through the last deprotonation step, the β-amyrin is generated.
Figure 1.17 Proposed mechanism of 2,3-oxidosqualene converted into β-amyrin and lupeol.
AH+ Enz O HO HO HO HO H B: H H HO (3S)-2,3-oxidosqualene Dammarenyl cation Chair-chair-chair conformation Lupenyl cation -amyrin Oleanyl cation HO Lupeol LUS PSY
21
Since the biogenetic isoprene rule proposed by Eschenmoser, Ruzicka, Arigoni, and Jeger in 1955,27 the cyclization of (3S)-2,3-oxidosqualene into β-amyrin has fascinated organic chemists for over a half century. To investigate the catalytic motifs within cyclases that form the dammarenyl cation, the Ebizuba group first generated chimeras of the A. thaliana lupeol synthase and the Panax ginseng β-amyrin synthase.37
They determined the function of the portions of cyclase by using a domain swapping strategy. It was observed that only relatively small portions of the protein can control the generation of lupeol or β-amyrin. One chimera in which only one fourth of the protein was β-amyrin sequence made four times as much β-amyrin as it did lupeol, and a mixed PCR method further confirmed the important region of chimeras. From an alignment analysis, it showed that the Trp259 within Panax ginseng β-amyrin synthase (PNY) and the Leu256 within Olea europa lupeol synthase (OEW) might control the product specificity.38 Therefore, the authors constructed the PNYTrp259Leu and OEWLeu256Trp mutants and conducted the product analysis. Lupeol was twice as abundant as β-amyrin in the product profile of the PNYTrp259Leu mutant, whereas β-amyrin was the major product for the OEWLeu256Trp mutant. These results indicated that this position plays a critical role in directing either β-amyrin or lupeol formation. Furthermore, the authors created the PNYTyr261His mutant and the experimental results showed that Tyr261 stabilizes one of the cationic intermediates formed after the dammarenyl cation.22,37-38 In addition, the experimental results for Pisum sativum β-amyrin synthase (PSY) showed that the expansion of the D-ring could take place in the absence of the terminal double bond, so that the formation of anti-Markovnikov six-membered D-ring is independent of the terminal π-electrons. Thereby, the aromatic residues within the putative active site might play a crucial role for the ring expansion reaction.21
22
1.3.6
The amino acid sequence alignment
The amino acid sequence alignment of the enzyme cyclases could provide much information and led us understand these cyclases in depth. In evolutionary history, some highly-conserved residues indicated that they may play some specific function and it is necessary for the organisms. Therefore, it would be retained in various species. However, some other residues were not conserved or even deleted through the variation of evolution in many species. The information of these differences helped us to figure out the cyclized mechanism of enzyme cyclases. Moreover, their similar structures, stereoselectivity, and the catalytic mechanism were also investigated.39 The relationship between the function and structure thus were deeply realized.
In order to understand the functional residues within the putative active site of triterpene cyclases, the program of Clustal W had been used to produce multiple sequence alignment of the following enzymes: H. sapiens ERG7: P48449, S. cerevisiae ERG7: P38604, A. thaliana CAS1: NP_178722, P. sativum PSY: BAA97558, A. acidocaldarius SHC: BAA25185. All of them were obtained from the Protein Data Bank (PDB) in NCBI, and the result of sequence alignment is shown below.
H.sapiens ERG7 MTEGTCLRRRGGPYKTEPATDLG--RWRLN-CERGRQTWTYLQDER---AGREQT 49 S.cerevisiae ERG7 MTEFYSDTIG---LPKTDPR--LWRLRTDELGRESWEYLTPQQ---AANDPP 44 A.thaliana CAS1 MWKLKIAEGGS-PWLRTTNNHVGRQFWEFDPNLGTPEDLAAVEEARKSFSDNRFVQKHSA 59 P.sativum PSY MWRLKIAEGGNDPYLFSTNNFVGRQTWEYDPEAGSEEERAQVEEARRNFYNNRFEVKPCG 60 A.acidocaldarius SHC ---
H.sapiens ERG7 GLEAYALGLDTKNYFKDLPKAH---TAFEGALN----GMTFYVGLQAED-GHWTGDY 98 S.cerevisiae ERG7 STFTQWLLQDPK-FPQPHPERNKHSPDFSAFDACHN----GASFFKLLQEPDSGIFPCQY 99 A.thaliana CAS1 DLLMRLQFSRENLISPVLPQVKIEDTDDVTEEMVETTLKRGLDFYSTIQAHD-GHWPGDY 118 P.sativum PSY DLLWRFQVLRENNFKQTIGGVKIEDEEEITYEKTTTTLRRGTHHLATLQTSD-GHWPAQI 119 A.acidocaldarius SHC ---MAEQLVEAPAYARTLDRAV---EYLLSCQKDE-GYWWGPL 36
23
H.sapiens ERG7 GGPLFLLPGLLITCHVAR---IPLPAGYREEIVRYLRSVQLP-DGGWGLHIEDKSTVFGT 154 S.cerevisiae ERG7 KGPMFMTIGYVAVNYIAG---IEIPEHERIELIRYIVNTAHPVDGGWGLHSVDKSTVFGT 156 A.thaliana CAS1 GGPMFLLPGLIITLSITGALNTVLSEQHKQEMRRYLYNHQNE-DGGWGLHIEGPSTMFGS 177 P.sativum PSY AGPLFFMPPLVFCVYITGHLDSVFPPEHRKEILRYIYCHQNE-DGGWGLHIEGHSTMFCT 178 A.acidocaldarius SHC LSNVTMEAEYVLLCHILDR----VDRDRMEKIRRYLLHEQRE-DGTWALYPGGPPDLDTT 91
H.sapiens ERG7 ALNYVSLRILGVGPDDP---DLVRARNILHKKGGAVAIPSWGKFWLAVLNVYSWEGLNTL 211 S.cerevisiae ERG7 VLNYVILRLLGLPKDHP---VCAKARSTLLRLGGAIGSPHWGKIWLSALNLYKWEGVNPA 213 A.thaliana CAS1 VLNYVTLRLLGEGPNDG-DGDMEKGRDWILNHGGATNITSWGKMWLSVLGAFEWSGNNPL 236 P.sativum PSY ALNYICMRILGEGPDGGEDNACVRARNWIRQHGGVTHIPSWGKTWLSILGVFDWLGSNPM 238 A.acidocaldarius SHC IEAYVALKYIGMSRDEE---PMQKALRFIQSQGGIESSRVFTRMWLALVGEYPWEKVPMV 148
H.sapiens ERG7 FPEMWLFPDWAPAHPSTLWCHCRQVYLPMSYCYAVRLSAAEDPLVQSLRQELYVEDFASI 271 S.cerevisiae ERG7 PPETWLLPYSLPMHPGRWWVHTRGVYIPVSYLSLVKFSCPMTPLLEELRNEIYTKPFDKI 273 A.thaliana CAS1 PPEIWLLPYFLPIHPGRMWCHCRMVYLPMSYLYGKRFVGPITSTVLSLRKELFTVPYHEV 296 P.sativum PSY PPEFWILPSFLPMHPAKMWCYCRLVYMPMSYLYGKRFVGPITPLILQLREELHTEPYEKI 298 A.acidocaldarius SHC PPEIMFLGKRMPLNIYEFGSWARATVVALSIVMSRQPVFPLPERARVP--ELYETDVPPR 206
H.sapiens ERG7 DWLAQRNNVAPDELYTPHSWLLRVVYALLNLYEHHHS---AHLRQRAVQKLYEHIVA 325 S.cerevisiae ERG7 NFSKNRNTVCGVDLYYPHSTTLNIANSLVVFYEKYLRN---RFIYSLSKKKVYDLIKT 328 A.thaliana CAS1 NWNEARNLCAKEDLYYPHPLVQDILWASLHKIVEPVLMRWPG-ANLREKAIRTAIEHIHY 355 P.sativum PSY NWTKTRHLCAKEDIYYPHPLIQDLIWDSLYIFTEPLLTRWPFNKLVRKRALEVTMKHIHY 358 A.acidocaldarius SHC RRGAKGG---GGWIFDALDRALHGYQKLSVHP---FRRAAEIRALDWLLE 250
H.sapiens ERG7 DDRFTKSISIGPISKTINMLVRWYVDGPASTAFQEHVSRIPDYLWMGLDGMKMQGTNGSQ 385 S.cerevisiae ERG7 ELQNTDSLCIAPVNQAFCALVTLIEEGVDSEAFQRLQYRFKDALFHGPQGMTIMGTNGVQ 388 A.thaliana CAS1 EDENTRYICIGPVNKVLNMLCCWVED-PNSEAFKLHLPRIHDFLWLAEDGMKMQGYNGSQ 414 P.sativum PSY EDENSRYLTIGCVEKVLCMLACWVED-PNGDAFKKHIARVPDYLWISEDGMTMQSF-GSQ 416 A.acidocaldarius SHC RQAGDGSWGGIQPPWFYALIALKILDMTQHPAFIKGWEGLELYGVELDYGGWMFQASISP 310
H.sapiens ERG7 IWDTAFAIQALLEAGGHHRPEFSSCLQKAHEFLRLSQVP-DNPPDYQKYYRQMRKGGFSF 444 S.cerevisiae ERG7 TWDCAFAIQYFFVAGLAERPEFYNTIVSAYKFLCHAQF---DTECVPGSYRDKRKGAWGF 445 A.thaliana CAS1 LWDTGFAIQAILATNLVE--EYGPVLEKAHSFVKNSQVLEDCPGDLNYWYRHISKGAWPF 472 P.sativum PSY EWDAGFAVQALLATNLIE--EIKPALAKGHDFIKKSQVTENPSGDFKSMHRHISKGSWTF 474 A.acidocaldarius SHC VWDTGLAVLALRAAGLPAD---HDRLVKAGEWLLDRQIT--VPGDWAVKRPNLKPGGFAF 365
24
H.sapiens ERG7 STLDCGWIVSDCTAEALKAVLLLQEK--CPHVTEHIPRERLCDAVAVLLNMRNPD----G 498 S.cerevisiae ERG7 STKTQGYTVADCTAEAIKAIIMVKNSPVFSEVHHMISSERLFEGIDVLLNLQNIGSFEYG 505 A.thaliana CAS1 STADHGWPISDCTAEGLKAALLLSKVP-KAIVGEPIDAKRLYEAVNVIISLQNAD----G 527 P.sativum PSY SDQDHGWQVSDCTAEGLKCCLLLSLLP-PEIVGEKMEPERLFDSVNLLLSLQSKK----G 529 A.acidocaldarius SHC QFDNVYYPDVDDTAVVVWALNTLRLPD---ERRRRDAMTKGFRWIVGMQSSN----G 415
H.sapiens ERG7 GFATYETKRGGHLLELLNPSEVFGDIMIDYTYVECTSAVMQALKYFHKRFPEHRAAEIRE 558 S.cerevisiae ERG7 SFATYEKIKAPLAMETLNPAEVFGNIMVEYPYVECTDSSVLGLTYFHKYF-DYRKEEIRT 564 A.thaliana CAS1 GLATYELTRSYPWLELINPAETFGDIVIDYPYVECTSAAIQALISFRKLYPGHRKKEVDE 587 P.sativum PSY GLAAWEPAGAQEWLELLNPTEFFADIVVEHEYVECTGSAIQALVLFKKLYPGHRKKEIEN 589 A.acidocaldarius SHC GWGAYDVDNTSDLPNHIPFCDFG--EVTDPPSEDVTAHVLECFG---SFGYDDAWK 466
H.sapiens ERG7 TLTQGLEFCRRQQRADGSWEGSWGVCFTYGTWFGLEAFACMGQTYRDGTACAEVSRACDF 618 S.cerevisiae ERG7 RIRIAIEFIKKSQLPDGSWYGSWGICFTYAGMFALEALHTVGETYEN---SSTVRKGCDF 621 A.thaliana CAS1 CIEKAVKFIESIQAADGSWYGSWAVCFTYGTWFGVKGLVAVGKTLKN---SPHVAKACEF 644 P.sativum PSY FIFNAVRFLEDTQTEDGSWYGNWGVCFTYGSWFALGGLAAAGKTYTN---CAAIRKGVKF 646 A.acidocaldarius SHC VIRRAVEYLKREQKPDGSWFGRWGVNYLYGTGAVVSALKAVGIDTREP----YIQKALDW 522
H.sapiens ERG7 LLSRQMADGGWGEDFESCEERRYLQSA--QSQIHNTCWAMMGLMAVRHPDIE--AQERGV 674 S.cerevisiae ERG7 LVSKQMKDGGWGESMKSSELHSYVDSE--KSLVVQTAWALIALLFAEYPNKE--VIDRGI 677 A.thaliana CAS1 LLSKQQPSGGWGESYLSCQDKVYSNLDGNRSHVVNTAWAMLALIGAGQAEVDRKPLHRAA 704 P.sativum PSY LLTTQREDGGWGESYLSSPKKIYVPLEGNRSNVVHTAWALMGLIHAGQSERDPTPLHRAA 706 A.acidocaldarius SHC VEQHQNPDGGWGEDCRSYEDPAYAGKG--ASTPSQTAWALMALIAGGRAESE--AARRGV 578
H.sapiens ERG7 RCLLEKQLPNGDWPQENIAG-VFNKSCAISYTSYRNIFPIWALGRFSQLYPERALAGHP 732 S.cerevisiae ERG7 DLLKNRQEESGEWKFESVEG-VFNHSCAIEYPSYRFLFPIKALGMYSRAYETHTL---- 731 A.thaliana CAS1 RYLINAQMENGDFPQQEIMG-VFNRNCMITYAAYRNIFPIWALGEYRCQVLLQQGE--- 759 P.sativum PSY KLLINSQLEQGDWPQQEITG-VFMKNCMLHYPMYRDIYPLWALAEYRRRVPLP--- 758 A.acidocaldarius SHC QYLVETQRPDGGWDEPYYTGTGFPGDFYLGYTMYRHVFPTLALGRYKQAIERR--- 631
Figure 1.18 Amino acids alignment. The red words are the CAS1mut and the green one is ERG7C703 mutation.
25
1.4
Research motive
The oxidosqualene cyclases family and their sterol products have fascinated scientists for more than half of a century. Not only biochemists, but also organic chemists and physical chemists, even pharmaceutical scientists and doctors are conducting research topic on this family of molecules. The goals of these studies are to understand the enzyme mechanisms, to make artificial enzymes to synthesize specific products, to find inhibitors to against bacteria or fungi, and to produce drugs to lower cholesterol levels or to kill cancer cells for various therapies.
The mechanism of the oxidosqualene cyclases activity involves cyclization and rearrangement. In fact, only one enzyme and one step is needed during the entire process of this amazing organic reaction. In addition, a few amino acid alterations could make a large stereochemical change on the product profiles. Currently, however, it is still troublesome to synthesize a compound when the processes are complicated, and involve a mass of chemicals. Studies on mechanisms in this area of chemistry could lead us to understand the relationship between the active-site amino acids and the appearances of products. In the future, synthesizing a complicated compound by using a few artificial enzymes is a goal worth achieving.
1.4.1 The study of Cys703 in SceERG7
In previous study,40 the cysteine-modifying agent, 5,5-dithio-bis-(2-nitrobenzoic) acid (DTNB), was used to assess the possible role of cysteine residues in the enzymatic function of bovine liver OSC activity. The results show that there are two cysteine residues that play important roles in catalytic function or conformation in active site of ERG7.
26
Furthermore, in the sequence alignment of SceERG7, Cys703 is a highly-conserved residue in the oxidosqualene cyclases family. By homology modeling, it is a secondary sphere residue in the active-site, where the Phe699 locates between Cys703 and the substrate. The minimum distance from Cys703 to the lanosterol molecule within the SceERG7 modeling structure is 7.01 Å , while the distance between Cys703 and Phe699 is only 3.28 Å . Therefore, the mutation on Cys703 might change the situation of Phe699, and influence the mechanism of cyclization and rearrangement (Fig. 1.19). Some further studies showed the importance of second-sphere residues. The best example is the His477 within AthCAS1, which is hydrogen bonded to Tyr410, thus it is essential for cycloartenol biosynthesis.33
27
According to the griffin aromatic hypothesis, the role of aromatic residues is considered to stabilize the highly-energy cation intermediate. The aromatic residues, which are within the putative active site, include Tyr99, Trp232, His234, Trp390, Trp443, Phe445, Tyr510, Phe699, and Tyr707. Among them, Phe699 affects the C-17 cation directly. The mutation on the Phe699 residue could produce many truncated compounds that reflect the unstable actuality of the C-17 cation. Therefore, the mutation on the Cys703 residue might indirectly influence the Phe699 residue to cause similar results.
On the other hand, the argument regarding the B-chair/boat ring and the 17α/17β mechanistic transitions has occurred lately in evolutionary history.41
The multiple products of oxidosqualene cyclases including the B-chair ring with the 17α/17β skeleton and B-boat ring with the 17β structure were isolated. Fascinatingly, no B-boat compounds with the 17α skeleton have found been in nature.42
So the method could be a means to understand the stereochemistry of enzymatic control. Through the mutation on the residue Cys703 within SceERG7, the steric effect close to the C-17 cation might change. By analyzing its coresponding products, the relationship of structure-function might be understood in better detail.
28
1.4.2 The study of CAS1 in A. thaliana and PSY in P. sativum
To understand the entire stereochemical mechanism of oxidosqualene cyclases, not only the information acquired from the mutation on Cys703 within SceERG7, but also the information by comparing the similarities and dissimilarities between cycloartenol synthase (CAS) and β-amyrin synthase (βAS) are needed. Either the CAS or βAS are found in higher plants. Some species, such as Arabidopsis thaliana, contain both of CAS and βAS, with distinct stero-chemical control. Many differences exist between CAS and βAS, even in the same species. CAS triggers the substrate into the chair-boat-chair conformation, forming the protosteryl cation intermediate and producing the final tetracyclic compound. βAS catalyzes the OS to go though the chair-chair-chair dammarenyl cation to form the pentacyclic product. In addition, there are some mutational studies between lupeol synthase and β-amyrin synthase as mentioned earlier, to illustrate their stero-chemical selectivity.
On the other hand, detailed information about amino acids in the active-site is critical to understand the whole mechanism of one enzyme. Just like the lanosterol cyclase from S. cerevisiae, the complete alanine-scanning of the active-site had been done by our laboratory. Our data indicated that the residue plays an important role in catalytic reaction of the enzyme, while an amino acid mutated to alanine produced novel products or lost the enzyme activity dramatically.
According to the view as mentioned above, to construct a complete site-directed alanine-scanning experiment is necessary. So based on the previous study19, many important residues were chosen in the SHC active-site. Fifteen amino acids from the SHC active-site had been chosen to find their corresponding residues in AthCAS1 and PsaPSY, then the change of the product profile was analyzed in each mutant (Fig. 1.20). The residues in CAS1 (or PSY) are V368(V371), L372(L374), W416(W418), F472(F474), C484(C486), E548(E550), F550(F552),
29
I553(I555), W610(W612), Y616(Y618), F726(F728), C730(C732), I732(L734), Y734(Y736) and Y737(Y739). By mutating the residues, more information could be obtained. Based on the given information, it is possible to preliminarily predict the stereochemical mechanism of enzyme-mediated cyclization, to propose a foundation for the further studies.
Figure 1.20 The alanine-scanning mutation position in the active-site of β-amyrin synthase within Pisum sativum.
30
Chapter 2 Materials and methods
2.1
Materials
2.1.1
Chemicals, reagents and kits
Purchase from Sigma:
Acetic anhydride Adenine Ampicillin sulfate D-Sorbitol Ergosterol Glucose Histidine Lysine Methionine Pyridine Tryptophan UracilPurchase from Merck:
95%Ethanol Acetic acid Acetone Anisaldehyde Dichloromethane Ether31 Ethyl acetate Glycerol Hemin Chloride Hexane Methanol Potassium hydroxide Pyrogallol Silica gel Sodium sulfate Sulfonic Acid TLC plate Tween 80
Purchase from USB:
Agarose-LEBromophenol blue
Purchase from DIFCO:
BactoTM AgarLB Broth, Miller Trypton
Yeast Extract
Yeast Nitrogen Base w/o amino acid
Purchase from other companies:
32
Dimethyl sulfoxide (MP Biomedicals) DNA 10Kb Ladder (Bio Basic Inc., Tanwan) dNTP Set, 100mM Solutions (GE Healthcare) G418 (Gibco)
GFXTMPCR DNA and Gel Band Purification Kit (GE Healthcare) Plasmid Miniprep Purification Kit (GeneMark)
Primers (Bio Basic Inc.)
QuikChange Site-Directed Mutagenesis Kit (Stratagene Inc., La Jolla, CA) Restriction enzymes (New England BioLabs Inc.)
SYBR® Green I (Roche)
2.1.2
Bacterial, yeast strains and vectors
Escherichia coli XL1-Blue (Novagen)
TKW14C2 (a yeast stain, MATa or MATα ERG7Δ:: LEU2 ade2-101 his3-Δ200 leu2-Δ1 lys2-801 trp1-Δ63 ura3-52 hem1Δ::KanR)
Vector pRS313 (a shuttle vector with selection marker His3, New England BioLabs) Vector pRS314 (a shuttle vector with selection marker Trp1, New England BioLabs)
2.1.3
Equipments
ABI PRISM® 3100 Genetic Analyzer (Applied Biosystems) AllegraTM 21R Centrifuge (Beckman Coulter)
Avanti® J0E Centrifuge (Beckman Coulter) Centrifuges 5415R (eppendorf)
Colling Circulator Bath Model B401L (Firstek Scientific) DU 7500 UV-Vis Spectrophotometer (Beckman Coulter) DurabathTM Water Bath (Baxter)
33
Electrophoresis Power Supply EPS 301 (GE healthcare)
GeneAmp® PCR System 9700 Thermal Cycler (Applied Biosystems) Hoefer® HE 33 Mini Horizontal Submarine Unit (GE Healthcare)
Kodak Electrophoresis Documentation an Analysis System 120 (Kodak) Orbital Shaking Incubator Model-S302R (Firstek Scintific)
Pulse Controller (BioRad)
Rotary Vaccum Evaporator N-N Series (EYELA) SteritopTM 0.22μm Filter Unit (Millipore)
2.1.4
Solutions and mediums
Ampicillin stock solution (100 mg/mL)
Dissolve 1 g ampicillin sulfate in 10 mL ddH2O. Filter through 0.22 μm pore size
filter and store at -20 °C.
50X TAE buffer
Dissolve Tris base 242 g, acetic acid 57.1 ml, and 0.5 M EDTA in 1 L dH2O and
adjust to pH 8.5.
50X ALTHMU solution
1 g Adeine, 1.5 g Lysine, 1 g Tryptophan,, 1 g Hisidine, 1 g Methonine, 1 g Uracil dissolved in 500 ml dH2O and sterilized.
50X ALHMU solution
1 g Adeine, 1.5 g Lysine, 1 g Hisidine, 1 g Methonine, 1 g Uracil dissolved in 500 ml dH2O and sterilized.
34
50X ALTMU solution
1 g Adeine, 1.5 g Lysine, 1 g Tryptophan, 1 g Methonine, 1 g Uracil dissolved in 500 ml dH2O and sterilized.
50% Glucose solution
500 g glucose dissolved in 1 L dH2O and sterilized.
80% Glycerol solution
80 mL glycerol added in 20 mL dH2O and sterilized.
LB medium
25 g LB Broth dissolved in 1 L dH2O and sterilized. Store at 4 °C.
LB plate
25 g LB Broth and 20 g BactoTM Agar dissolved in 1 L dH2O and sterilized. The
sterile LB agar was poured and dispersed in Petri dishes before it coagulates. Store at 4 °C.
G418 stock solution (1g/mL)
Dissolve 500 mg G418 in 500 μl sterile dH2O. Store in darkness at 4 °C.
YNB medium
6.7 g Yeast nitrogen base dissolved in 1 L dH2O and sterilized.
1 M sorbitol solution