Regulation of ammonium-induced proline accumulation in detached rice
leaves
Chuan Chi Lin and Ching Huei Kao*
National Taiwan University, Department of Agronomy, Taipei, Taiwan, Republic of China; *Author for correspondence (e-mail: [email protected])
Received 11 July 2000; accepted in revised form 8 August 2000
Key words: Ammonium, Oryza sativa, Proline accumulation
Abstract
Accumulation of proline in response to NH4Cl was studied in detached leaves of rice (Oryza sativa cv. Taichung
Native 1). Increasing concentrations of NH4Cl from 50 to 200 mM progressively increased proline content and
this was correlated with the increase in ammonium content. Proline accumulation induced by NH4Cl was related
to proteolysis, an increase in ornithine-␦-aminotransferase activity, a decrease in proline dehydrogenase activity, and a decrease in proline utilisation and could not be explained by NH4Cl-induced modification in⌬
1
-pyrroline-5-carboxylate reductase activity. The content of glutamic acid was decreased by NH4Cl, whereas the increase in
arginine and ornithine contents was found to be associated with the increase in proline content in NH4Cl-treated
detached rice leaves.
Abbreviations: FW – fresh weight, GABA –␥-aminobutyric acid, OAT – ornithine-␦-aminotransferase, P5C – ⌬1-pyrroline-5-carboxylate, P5CR –⌬1-pyrroline-5-carboxylate reductase, PDH – proline dehydrogenase
Introduction
The ammonium ion is a central intermediate in the metabolism of plants. Ammonium has been shown to accumulate in leaves subjected to water stress, when exposed to excess Cu or excess Cd, and during dark-induced leaf senescence (Chen and Kao 1998; Chen et al. 1997; Chien and Kao 2000; Lin and Kao 1998; Postitus and Jacobi 1976; Thomas 1978). Proline has been shown to accumulate in plants subjected to vari-ous types of stress (Aziz et al. 1998; Bassi and Sharma 1993; Charest and Phan 1990; Chen and Kao 1995; Guerrier 1995; Lutts et al. 1999; Madan et al. 1995; Mattioni et al. 1997; Savoure et al. 1997; Wu et al. 1995; Yang et al. 1999, 2000) and during dark-induced leaf senescence (Chou et al. 1990; Wang et al. 1982). Proline accumulation is often considered to be involved in stress resistance mechanisms, although its precise role continues to be controversial (Hare et al. 1999; Yoshiba et al. 1997). Based on data in the literature, Rabe (1990) postulated that any stress
con-dition causing reduced growth or impaired plant health will result in ammonium accumulation early in the stress period and suggested that the detoxification process in which excess ammonium in the cells is re-moved results in the accumulation of nitrogen-con-taining compounds, such as putrescine and proline. In previous studies, we demonstrated that exogenous NH4Cl and methionine sulfoximine, which caused an
accumulation of ammonium in detached rice leaves, increased proline content (Yang and Kao 1999).
Proline accumulation in plant tissues has been sug-gested to be the result of (a) a decrease in proline degradation, (b) an increase in proline biosynthesis, (c) a decrease in protein synthesis or proline utilisa-tion, and (d) hydrolysis of proteins (Charest and Phan 1990; Hare et al. 1999; Yoshiba et al. 1997). In plants, proline is synthesised from glutamic acid via⌬1
-pyr-roline-5-carboxylate (P5C) by two enzymes, P5C synthetase and P5C reductase (P5CR) (Yoshiba et al. 1997). It has been shown from labelling experiments that ornithine can also serve as a precursor to proline
biosynthesis in higher plants (Brown and Fowden 1966; Chiang and Dandekar 1995; Coleman and He-garty 1957). The isolation of cDNA encoding orni-thine-␦-aminotransferase (OAT) in higher plants (De-launey and Verma 1993; Roosens et al. 1998) sug-gests that OAT participates in proline biosynthesis by producing P5C from ornithine and ␣-ketoglutarate. Arginine can also contribute to proline biosynthesis, and the pathway from arginine proceeds via ornithine as a result of catalytic activity of arginase (Brown and Fowden 1966; Lingnowski and Splittstoessor 1971). Proline is metabolised to glutamic acid via P5C by two enzymes, proline dehydrogenase (PDH) and P5C dehydrogenase (Yoshiba et al. 1997).
Studying the effect of ammonium on enzyme ac-tivities involved in proline biosynthesis and degrada-tion could provide valuable informadegrada-tion on the physi-ological significance of its accumulation. However, to our knowledge, no such study has been undertaken. Neither do we know whether three amino acids (glutamic acid, ornithine and arginine) involved in the proline biosynthesis pathways are limiting factors for proline induced by excess ammonium. This paper re-ports the results of an investigation into the regula-tion of proline accumularegula-tion in detached rice leaves exposed to NH4Cl.
Materials and methods Plant material
Rice (Oryza sativa L., cv. Taichung Native 1) was culutured as described previously (Lin et al. 1999). Briefly, rice seedlings were planted on a stainless net floating on half-strength Johnson’s modified nutrient solution (Johnson et al. 1957) in a 500 ml beaker. The nutrient solution (pH 4.8) was replaced every three days. Rice plants were grown for 12 days in a green-house, where natural light was provided and the tem-perature was controlled at 30 °C during the day and at 25 °C at night. The apical 3 cm of the third leaf was used for the experiment. A group of 10 segments was floated in a Petri dish containing 10 ml of test solution. Incubation was at 27 °C in the light (40
mol m−2s−1). All the experiments were repeated
four times and yielded results consistent with the data presented here.
Determinations of proline and other amino acids
Proline was extracted and its concentration deter-mined by the method of Bates et al. (1973). Leaf seg-ments were homogenised with 3% sulfosalicylic acid and the homogenate was centrifuged at 3,000 g for 20 min. The supernatant was treated with acetic acid and acid ninhydrin, boiled for 1 h and then absor-bance at 520 nm was determined. Contents of proline are expressed as mol g−1
FW. For determination of glutamic acid, glutamine, arginine, ornithine, ␥ -ami-nobutyric acid (GABA), and total amino acids, leaf samples were extracted with 2% sulfosalicylic acid and the homogenate was centrifuged at 15,000 g for 20 min. The supernatant was used directly for amino acid analysis. Amino acid analysis was done using an amino acid analyzer (Beckman 6300, California, USA) and contents of amino acids are expressed as nmol g−1 FW. For protein determination, leaf
seg-ments were homogenised in 50 mM sodium phos-phate buffer (pH 6.8). The extracts were centrifuged at 17,600 g for 20 min, and the supernatants were used for determination of protein by the method of Bradford (1976). Protein content is expressed as mg g−1FW.
Enzyme assay
A similar extraction procedure was used for P5CR and PDH, which is mainly based on the procedure described by Lutts et al. (1999). Detached rice leaves were homogenised in a prechilled mortar and pestle with 50 mM Tris-HCl buffer (pH 7.4) containing 7 mM MgCl2, 0.6 M KCl and 3 mM EDTA. The
ho-mogenate was centrifuged at 15,000 g for 20 min. The supernatant was desalted by Sephadex G-25 column before the assay of P5CR and PDH. The solution used for extraction of OAT was 100 mM potassium phos-phate buffer (pH 7.4) containing 1 mM pyridoxal-5⬘-phosphate, 1 mM EDTA and 10 mM 2-mercaptoetha-nol. The extract was centrifuged at 12,000 g. The supernatant was desalted by Sephadex G-25 column before the assay of OAT. All the extraction procedures were done at 4 °C.
P5CR was assayed by a NADH dependent P5CR reaction (Madan et al. 1995). The assay mixture con-tained 0.06 mM NADH, 0.15 mM P5C, 120 mM po-tassium phosphate buffer, 2 mM dithiothreitol, and the enzyme extract. The reaction was started by the addition of P5C and the decrease in absorbance was followed at 340 nm. P5CR is expressed as units g−1
FW (one unit is defined as a decrease in 1 A340 per
min) or units mg−1protein (one unit is defined as a
decrease in 0.01 A340per min). OAT activity was
sayed according to Vogel and Kopac (1960). The as-say mixture contained 0.2 ml enzyme extract and 0.8 ml 100 mM potassium phosphate buffer (pH 8.0) con-taining 50 mM L-ornithine, 20 mM ␣-ketoglutarate and 1 mM pyridoxal-5⬘-phosphate. The reaction me-dium was incubated at 37 °C for 30 min. The reaction was stopped by adding 0.5 ml trichloroacetic acid (10%) and the colour was developed by incubating the reaction mixture with 0.5 ml o-aminobenzalde-hyde (0.5%) in ethanol (95%) for 1 h. After centri-fugation at 12,000 g for 10 min, the clear supernatant fraction was taken to measure the absorbance at 440 nm. OAT is expressed as units g−1 FW (one unit is
defined as an increase in 1 A440per h) or units mg −1
protein (one unit is defined as an increase in 0.01 A440
per h). PDH was assayed by following the NAD+
re-duction at 340 nm in a 0.15 M Na2CO3-HCl buffer
(pH 10.3) containing 13 mM L-proline and 1.5 mM NAD+(Lutts et al. 1999). PDH is expressed as units
g−1FW (one unit is defined as an increase in 1 A 340
per min) or units mg−1protein (one unit is defined as
an increase in 0.01 A340per min).
Determination of proline utilisation
For proline utilisation, detached rice leaves were pre-treated with 50 mM ornithine for 3 h (since addition of ornithine has been observed to be more effective than that of glutamic acid or arginine in increasing proline content in rice leaves (Yang et al. 1999)) to increase the endogenous proline content and then transferred to distilled water and 100 mM NH4Cl for
8 h in the light. Proline content was then determined. The decline in proline content was considered as a measure of proline is utilisation (Yang et al. 1999). Determination of ammonium
Ammonium was extracted by homogenising leaf seg-ments in 0.3 mM sulphuric acid (pH 3.5). The homo-genate was centrifuged for 10 min at 39,000 g and the supernatant was used for the determination of am-monium as described previously (Lin and Kao 1998).
Results and discussion
Proline content in detached rice leaves increased with the increase of NH4Cl concentrations (Figure 1).
Pro-line content increased 4- to 5-fold in detached leaves treated with 100 mM NH4Cl for 12 h (Figures 1 and
2). To be sure that the proline accumulation described was related to leaf ammonium content, ammonium concentrations were determined in detached rice leaves treated with various concentrations of NH4Cl.
Ammonium content in NH4Cl-treated detached rice
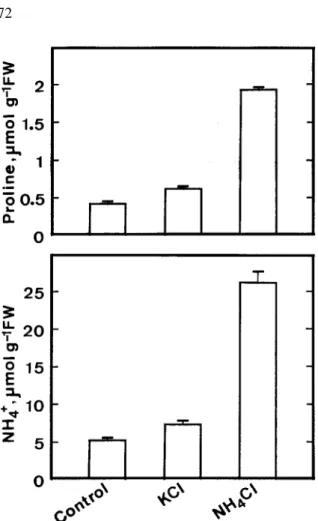
leaves increased with increasing concentrations of NH4Cl (Figure 1). Since treatment of 100 mM KCl
only resulted in a slight increase in ammonium and proline contents (Figure 2), it seemed that the in-crease in proline content in NH4Cl-treated detached
rice leaves was mainly associated with the increase in ammonium content rather than of Cl−.
Figure 1. Effect of NH4Cl on ammonium and proline contents in
detached rice leaves. All measurements were made 12 h after treat-ment in the light. Vertical bars represent SE (n = 4).
It is well established that glutamic acid, ornithine and arginine can all contribute to the accumulation of proline in plant tissues (Chiang and Dandekar 1995). In previous studies, we reported that exogenous ap-plication of ornithine or arginine was effective in in-creasing the proline content in detached rice leaves (Yang et al. 1999). In contrast, proline content could not be increased in detached rice leaves by the addi-tion of glutamic acid up to 50 mM (Yang et al. 1999). It seems that detached rice leaves contained sufficient amounts of glutamic acid. Thus, it is of great interest to know the effect of NH4Cl on the content of
orni-thine, arginine and glutamic acid contents in detached rice leaves. Table 1 shows that NH4Cl treatment
re-sulted in an increase in ornithine and arginine con-tents in detached rice leaves and a decrease in glutamic acid content. These observations suggest that high contents of endogenous ornithine and argi-nine are associated with NH4Cl-induced proline
ac-cumulation in detached rice leaves. Also shown in Table 1 is that NH4Cl treatment increases glutamine
and GABA contents. It seems that the decreased con-tent of glutamic acid by NH4Cl may result from it
being metabolised to GABA (Ireland and Lea 1999). The increase in glutamine content and the decrease in glutamic acid in NH4Cl-treated detached rice leaves
also suggest that NH4Cl inhibits the conversion of
glutamine to glutamic acid, a step catalysed by glutamic acid synthase (Ireland and Lea 1999). The results of Table 1 show that protein content decreases and total amino acids increases in NH4Cl-treated
de-tached rice leaves, indicating that hydrolysis of pro-teins in NH4Cl-treated detached rice leaves is also
responsible for the accumulation of proline.
To determine the role of biosynthetic pathways for proline accumulation caused by NH4Cl, the effect of
NH4Cl on OAT and P5CR activities was examined.
The results are shown in Figure 1. OAT activity, ex-pressed either on per g FW or per mg protein basis, increased with increase of concentrations of NH4Cl.
However, NH4Cl had no effect on P5CR activity in
detached rice leaves. Thus, the increase in OAT ac-tivity may have contributed, to some extent, to the el-evated content of proline by NH4Cl. Increase in OAT
activity, along with an increase in the content of pro-line has also been reported in wheat under cold stress (Charest and Phan 1990), and in Brassica juncea un-der salt stress (Madan et al. 1995). NH4Cl could also
have an effect on the bifunctional enzyme P5C syn-thetase involved in the synthesis of P5C from glutamic acid (Hare et al. 1999; Yoshiba et al. 1997). However, for unknown reasons, we were unable to detect any P5C synthetase activity in crude extracts. Therefore, this enzyme was not considered in the present work.
The enzyme PDH is reported to catalyse proline oxidation (Hare et al. 1999; Yoshiba et al. 1997). In the present investigation, NH4Cl treatment resulted in
a decrease in PDH activity in detached rice leaves (Figure 3). This result suggests that proline oxidation (or degradation) contributes to proline accumulation in detached rice leaves exposed to NH4Cl.
Recently, we demonstrated that proline in detached rice leaves exposed to water stress is less utilised than in the water controls (Yang et al. 2000). It is possible that less utilisation of proline may also contribute to the accumulation of proline in detached rice leaves caused by NH4Cl. The results reported in Table 2
demonstrated that this is, in fact, correct.
Figure 2. Effect of KCl and NH4Cl on ammonium and proline
contents in detached rice leaves. Detached rice leaves were treated with distilled water, 100 mM KCl or 100 mM NH4Cl for 12 h in
In conclusion, proline accumulation, in detached leaves, caused by ammonium is related to protein hy-drolysis, an increase in ornithine and arginine con-tents, an increase in OAT activity, a decrease in PDH activity, and a decrease in proline utilisation. In the present investigation, the regulation of proline accu-mulation caused by ammonium was studied using de-tached rice leaves. It is not known whether similar mechanism is operated in intact rice leaves. Thus, ammonium-regulated proline accumulation in intact leaves of rice plants merits future investigation.
Acknowledgements
This work supported by grant NSC 89-2313-B002-111 from the National Science Council of the Repub-lic of China.
References
Aziz A., Martin-Tanguy J. and Larher F. 1998. Stress-induced changes in polyamine and tyramine levels can regulate proline accumulation in tomato leaf discs treated with sodium chloride. Physiol. Plant. 104: 195–202.
Bassi R. and Sharma S.S. 1993. Changes in proline content accom-panying the uptake of zinc and copper by Lemna minor. Ann. Bot. 72: 151–154.
Bates L.S., Waldren S.P. and Teare I.D. 1973. Rapid determination of free proline for water-stress studies. Plant Soil. 39: 205–207. Bradford M.M. 1976. A rapid and sensitive method for the quan-titation of microgram quantities of protein utilising the princi-ple of protein-dye binding. Anal. Biochem. 72: 248–254. Brown D.H. and Fowden L. 1966. Metabolism of␦-acetylornithine
in two Leguminous species. Phytochemistry. 5: 887–892. Charest C. and Phan C.T. 1990. Cold acclimation of wheat:
Prop-erties of enzymes involved in proline metabolism. Physiol. Plant. 80: 159–168.
Chen L.-M. and Kao C.H. 1998. Relationship between ammonium accumulation and senescence of detached rice leaves caused by excess copper. Plant Soil. 200: 169–173.
Chen S.L. and Kao C.H. 1995. Cd induced changes in proline con-tent and peroxidase activity in roots of rice seedlings. Plant Growth Regul. 17: 67–71.
Table 1. Contents of glutamate, ornithine, arginine, GABA, protein and total amino acids in detached rice leaves incubated in distilled water
or NH4Cl.
Treatment Protein (mg g−1FW) Amino acids (nmol g−1FW)
Total amino acids Glutamine Glutamate Ornithine Arginine GABA H2O 60.55 ± 1.05 20120 ± 775 1383 ± 88 6898 ± 215 17 ± 1 287 ± 50 382 ± 142 NH4Cl 50.29 ± 0.74 78533 ± 1169 32877 ± 1183 3179 ± 110 55 ± 8 1088 ± 149 2653 ± 933
Detached rice leaves were treated with distilled water or NH4Cl (100 mM). All measurements were made 12 h after treatment. Means ± SE
(n = 4).
Figure 3. Effect of NH4Cl on the activities of OAT, P5CR and
PDH in detached rice leaves. All measurements were made 12 h after treatment in the light. Vertical bars represent SE (n = 4).
Table 2. Proline content in ornithine-pretreated detached rice
leaves incubated in distilled water and NH4Cl
Treatment Proline,mol g−1FW
H2O, 3 h 0.77 ± 0.17
Ornithine, 3 h 27.13 ± 0.68
Ornithine, 3 h→ H2O, 8 h 13.78 ± 0.45 Ornithine, 3 h→ NH4Cl, 8 h 19.12 ± 0.66
Detached rice leaves were pretreated with 50 mM ornithine for 3 h in the light and then incubated in distilled water or NH4Cl (100
Chen S.J., Hung K.T. and Kao C.H. 1997. Ammonium accumula-tion is associated with senescence of rice leaves. Plant Growth Regul. 21: 195–201.
Chiang H.-H. and Dandekar A.M. 1995. Regulation of proline ac-cumulation in Arabidopsis thaliana (L.) Heynh during devel-opment and in response to desiccation. Plant Cell Environ. 18: 1280–1290.
Chien H.-F. and Kao C.H. 2000. Accumulation of ammonium in rice leaves in response to excess cadmium. Plant Sci. 156: 111– 115.
Chou I.T., Chen C.T. and Kao C.H. 1990. Regulation of proline accumulation in detached rice leaves. Plant. Sci. 70: 43–48. Coleman R.G. and Hegarty M.P. 1957. Metabolism of D,
L-orni-thine-214C in normal and potassium-defficient barley. Nature.
179: 376–377.
Delauney A.J. and Verma D.P.S. 1993. Proline biosynthesis and osmoregulation in plants. Plant. J. 4: 215–223.
Guerrier G. 1995. Effect of salt-stress on proline metabolism in calli of Lycopersicon esculentum, Lycopersicon pennellii, and their interspecific hybrid. Can. J. Bot. 73: 1939–1946. Hare P.D., Cress W.A. and van Staden J. 1999. Proline synthesis
and degradation: a model system for elucidating stress-related signal transduction. J. Exp. Bot. 333: 413–434.
Ireland A.J. and Lea P.J. 1999. The enzymes of glutamine, glutamate, asparagine, and aspartate metabolism. In: Singh B.K. (ed.), Plant Amino Acids. Mercel Dekker Inc, New York, pp. 49–109.
Johnson C.M., Stout P.R., Broyer T.C. and Carlton A.B. 1957. Comparative choline requirements of different plant species. Plant. Soil. 8: 337–353.
Lin J.-N. and Kao C.H. 1998. Water stress, ammonium, and leaf senescence in detached rice leaves. Plant Growth Regul. 26: 165–169.
Lin J.-N., Wang J.-W. and Kao C.H. 1999. Effect of abscisic acid and water stress on the senescence of detached rice leaves. Biol. Plant. 42: 313–316.
Lingnowski E.M. and Splittstoessor W.E. 1971. The change in argi-nine levels and metabolism of urea and ornithine in Cucubita
moschatta seedlings. Physiol. Plant. 25: 225–229.
Lutts S., Majerus V. and Kinet J.M. 1999. NaCl effects on proline metabolism in rice (Oryza sativa L.) seedlings. Physiol. Plant. 105: 450–458.
Madan S., Nainawatee H.S., Jain R.K. and Chowdhury J.B. 1995. Proline and proline metabolising enzymes in in-vitro selected NaCl-tolerant Brassica juncea L. under salt stress. Ann. Bot. 76: 51–57.
Mattioni C., Lacerenza N.G., Troccoli A., De Leonardis A.M. and Di Fonzo N. 1997. Water and salt stress-induced alterations in proline metabolism of Triticum durum seedlings. Physiol. Plant. 101: 787–792.
Postitus C. and Jacobi G. 1976. Dark starvation and plant metabo-lism. VI. Biosynthesis of glutamic acid dehydrogenase in de-tached leaves of Cucurbita maxima. Z. Pflanzenphysiol. 78: 133–140.
Rabe E. 1990. Stress physiology: The functional significance of the accumulation of nitrogen-containing compounds. J. Hort. Sci. 65: 231–243.
Rena A.B. and Splittstoesser W.E. 1975. Proline dehydrogenase and pyrroline-5-carboxylate reductase from pumpkin cotyle-dons. Phytochemistry. 14: 657–661.
Roosens N.H.C.J., Thu T.T., Iskandar H.M. and Jacobs M. 1998. Isolation of the ornithine-␦-aminotransferase cDNA and effect of salt stress on its expression in Arabidopsis thaliana. Plant. Physiol. 117: 263–271.
Savoure A., Hua X.-J., Bertauche N., van Montagu M. and Ver-bruggen N. 1997. Abscisic independent and abscisic acid-dependent regulation of proline biosynthesis following cold and osmotic stresses in Arabidopsis thaliana. Mol. Gen. Genet. 254: 104–109.
Thomas H. 1978. Enzymes of nitrogen mobilization in detached leaves of Lolium temulentum during senescence. Planta. 142: 161–169.
Vogel R.H. and Kopac M.J. 1960. Some properties of ornithine-␦ -transaminase from Neurospore. Biochem. Biphys. Acta. 37: 539–540.
Wang C.Y., Cheng S.H. and Kao C.H. 1982. Senescence of leaves VII. Proline accumulation in senescing excised leaves. Plant. Physiol. 69: 1348–1349.
Wu J.-T., Chang S.J. and Chou T.-L. 1995. Intracellular proline ac-cumulation in algae exposed to copper and cadmium. Bot. Bull. Acad. Sin. 36: 69–93.
Yang C.-W. and Kao C.H. 1999. Ammonium in relation to proline accumulation in detached rice leaves. Plant Growth Regul. 30: 139–144.
Yang C.-W., Lin C.C. and Kao C.H. 1999. Endogenous ornithine and arginine contents and dark-induced proline accumulation in detached rice leaves. J. Plant. Physiol. 155: 665–668. Yang C.-W., Wang J.-W. and Kao C.H. 2000. The relation between
accumulation of abscisic acid and proline in detached rice leaves. Biol. Plant. 43: 301–304.
Yoshiba Y., Kiyosue T., Nakashima K., Yamaguchi-Shinozaki K. and Shinozaki K. 1997. Regulation of levels of proline as an osmolyte in plants under water stress. Plant. Cell Physiol. 38: 1095–1102.