The fate of ignored stimuli is germane to the under-standing of selective attention. Unattended stimuli can be filtered out completely before perceptual identification is reached (e.g., Broadbent, 1958) or can be processed to the semantic level before being blocked from response selection (e.g., Deutsch & Deutsch, 1963). The phenom-enon of negative priming (Tipper, 1985) sheds light on the fate of ignored stimuli. When an ignored prime stim-ulus becomes the target in the subsequent probe trial (i.e., the ignored repetition [IR] condition), people tend to respond at a slower speed than to a target that is unre-lated to prime stimuli (i.e., the control condition). Nega-tive priming manifests even when an ignored prime dis-tractor (e.g., a picture of a cat) and the subsequent probe target (e.g., the word dog) are in different formats but share a semantic relationship (Tipper & Driver, 1988). The results suggest that the ignored stimuli are processed to the semantic level. The slower response is understood to reflect the working of inhibitory mechanisms in se-lective attention (Tipper & Cranston, 1985).
Depending on which feature of an ignored stimulus is repeated in the probe target, different versions of negative priming can be found in the literature. Identity negative priming occurs when a prime distractor and its probe tar-get share the same identity. Since the first observation of the phenomenon of negative priming (Dalrymple-Alford
& Budayr, 1966), researchers have concentrated only on identity negative priming (e.g., Lowe, 1979; Neill, 1977; Tipper, 1985). Tipper, Brehaut, and Driver (1990) sug-gested that in addition to the traditional select-where, respond-what task, the select-what, respond-where task is also of importance. Through the introduction of a lo-calization task, they showed a slower response to an item at a location that had been occupied by a distractor, even though the two did not share identity information. This is the location negative priming effect, a slower response to a probe target situated at the location that a prime dis-tractor has recently occupied. In addition to these two versions, semantic negative priming had also been stud-ied (e.g., Abad, Noguera, & Ortells, 2003; Fox, 1996). One important characteristic of negative priming is its dependency on the use of repeated stimuli, which has been well documented in the domain of identity negative priming. In a series of experiments, Strayer and his col-leagues (Grison & Strayer, 2001; Malley & Strayer, 1995; Strayer & Grison, 1999) demonstrated that identity neg-ative priming is contingent on repeated stimuli. When nonrepeated stimuli are used as test items, negative prim-ing is often absent, or instead, positive primprim-ing is ob-served. Dark and Schmidt (2000) found that a single episode of semantic priming is insufficient for the man-ifestation of negative priming. In a recent study, Kramer and Strayer (2001) also demonstrated the contingency of identity negative priming on repeated stimuli with both young and old adults as participants. It appears that iden-tity negative priming is difficult to manifest when novel stimuli are used.
Although there is empirical evidence supporting the contingency of stimulus repetition in identity negative priming, no empirical study has been conducted to in-vestigate this contingency in location negative priming. The finding of contingency in location negative priming is important for understanding whether both identity
789 Copyright 2005 Psychonomic Society, Inc.
Parts of this work were presented in November 2002 at the 43rd Annual Meeting of the Psychonomic Society, Kansas City, MO. This study was supported by Grant NSC92-2413-H-002-025 from the National Science Council to the second author. We thank the Ministry of Education Com-mittee for the Promotion and Propagation of the National Language for providing the database from which we selected stimuli. We also thank Hsiao-Chueh Chang for help in data collection. Correspondence concern-ing this article should be addressed to H.-F. Chao or Y.-Y. Yeh, Depart-ment of Psychology, National Taiwan University, No. 1, Sec. 4, Roosevelt Rd., Taipei, Taiwan 106 (e-mail: d90227005@ntu.edu.tw or yyy@ntu. edu.tw).
Location negative priming in identity
discrimination relies on location repetition
HSUAN-FU CHAO and YEI-YU YEH
National Taiwan University, Taipei, Taiwan
Negative priming manifests when a previously ignored stimulus becomes a target. The contingency of identity negative priming on repeated stimuli has been demonstrated, implying a crucial role for dis-tractor competition. In this study, a naming task was used to examine whether location negative prim-ing also relies on the repetition of locations. In Experiment 1, location negative primprim-ing was observed only when a small set of repeated locations was used. Positive priming was found instead when a large set of nonrepeated locations was used. Experiment 2 demonstrated that target-to-distractor distance modulated location priming effects, with priming effects observed only for a far distance. Experiments 3 and 4 demonstrated that the effect of location negative priming increased as locations repeated. Like identity negative priming, location negative priming depends on location repetition.
negative priming and location negative priming follow the same operating principles and whether they are based on the same mechanisms of selective attention (e.g., Tip-per & Cranston, 1985) or memory retrieval (e.g., Neill, Valdes, Terry, & Gorfein, 1992). Christie and Klein (2001) have suggested that location negative priming does not reflect distractor inhibition. Instead, multiple mecha-nisms are involved, including inhibition of return (IOR; Posner & Cohen, 1984) to any stimulated position that has been occupied by a target or a distractor, attention cap-ture by new objects in positions that have not been occu-pied, and memory retrieval. Milliken, Tipper, Houghton, and Lupiáñez (2000) also have proposed that location negative priming shares mechanisms similar to those un-derlying the phenomenon of IOR. They suggested that the manifestation of location negative priming depends on the relative speed of two processes: attention capture by novelty and spatiotemporal integration with existing episodic representation. If location negative priming re-flects processes different from those underlying identity negative priming, they may follow different operating principles.
Several studies have demonstrated the dissociation be-tween these two types of negative priming. For example, although aging affects the manifestation of identity neg-ative priming, it has little effect on location negneg-ative prim-ing (Connelly & Hasher, 1993). Although there have been challenges to this dissociation (e.g., Sullivan & Faust, 1993), a meta-analysis has revealed reduced iden-tity negative priming in older adults, whereas location negative priming remained stable with age (Verhaeghen & De Meersman, 1998). Koshino, Boese, and Ferraro (2000) used regression analysis to infer that different brain areas are responsible for identity negative priming and location negative priming.
The main purpose of this study was to examine whether location negative priming is contingent on location rep-etition. If location negative priming is also contingent on location repetition, location negative priming and iden-tity negative priming share at least one operating princi-ple. Furthermore, the finding of contingency in location negative priming corroborates the proposition that activa-tion level is a determinative factor in inhibitory processes for selective attention or in building durable traces for memory retrieval.
Four experiments were conducted. In Experiment 1, location negative priming was examined by using re-peated locations in one block and testing nonrere-peated lo-cations in another block. Experiments 2A and 2B repli-cated Experiment 1 while examining the impact of target–distractor distance on location negative priming. A novel set of locations was repeatedly used in a block in Experiment 3, with each block divided into quartiles in chronological order. Whether the magnitude of tion negative priming increases as the number of loca-tion repetiloca-tion increases across quartiles in a block was examined. Finally, Experiment 4 replicated Experiment 3 while examining the effect of location repetition on tri-als in which the target location was repeated in the
prime–probe couplet. Through these experiments, the impact of location repetition was examined. It should be noted that stimulus repetition was kept constant by using a small set of stimuli in all the experiments.
An identity discrimination task was used in this study. Most experiments on location negative priming have used a localization task by prespecifying the mappings of buttons to locations (e.g., Buckolz, Boulougouris, O’Donnell, & Pratt, 2002; Connelly & Hasher, 1993; Neill, Terry, & Valdes, 1994). Because the number of lo-cations used in these previous studies was small, it was possible to assign a particular response button to each location. However, in this study, hundreds of locations were used, and it would have been difficult to assign one-to-one button-to-location mappings. Thus, an identifica-tion task was chosen, with the advantage that there was no need to prespecify the mappings of buttons to locations.
EXPERIMENT 1
With a design conceptually similar to Strayer and Grison’s (1999) Experiment 1, this experiment tested whether location negative priming is contingent on loca-tion repetiloca-tion. Rather than using repeated or nonre-peated stimuli for identification, we used renonre-peated or nonrepeated locations in an identif ication task. Two blocks were used. As in many studies of location nega-tive priming, in one block (repeated condition), only four locations were repetitively used within the block. In the other block (nonrepeated condition), a location was never repeated until it became the location of a prime distrac-tor and then, again, the location of the probe target in the IR condition. This experiment involved a couplet type (IR/control) ⫻ stimulus type (nonrepeated/repeated lo-cation) design. If location negative priming is contingent on location repetition, negative priming should be found only in the repeated condition, and not in the nonre-peated condition.
Method
Participants. Twenty-five undergraduate students at National Taiwan University participated in this experiment for a bonus credit in an introductory psychology course. All the participants were naive as to the purpose of the experiment and had normal or corrected-to-normal vision.
Stimuli. Locations were selected from an invisible 16 ⫻ 16 ma-trix. The matrix was located in the center of the screen. The view-ing distance was about 60 cm, and each cell of the matrix had a width of 1.1º and a height of 1.1º. Each stimulus word was pre-sented in the center of a cell and subtended a visual angle of 0.8º vertically and horizontally. The target word was presented in red, and the distractor word was shown in green.
Four locations were used in the repeated condition. These four lo-cations were randomly selected from the 256 lolo-cations of the ma-trix, under the constraint that each location was not too close to the others selected (see the Appendix for the coordinates of these loca-tions). The other 168 locations were randomly selected to position stimuli in the nonrepeated condition. Finally, another 16 locations were selected for the practice trials in the nonrepeated condition. To minimize the effect of naming difficulty, only four Chinese character words were used as stimuli in the experiment. These four words were of high frequency, ranging from 700 to 1,000 per
mil-lion (Ministry of Education Committee for the Promotion and Prop-agation of the National Language, 1997). These words were also of low visual complexity, with the number of strokes ranging from 6 to 8, so that location effects could be tested with low demand on identity discrimination.
Procedure. The experiment was run by DMDX software (Forster & Forster, 2003). Naming latency was measured with a microphone connected to an audio card. Each participant was tested individu-ally, and the experimenter recorded naming accuracy.
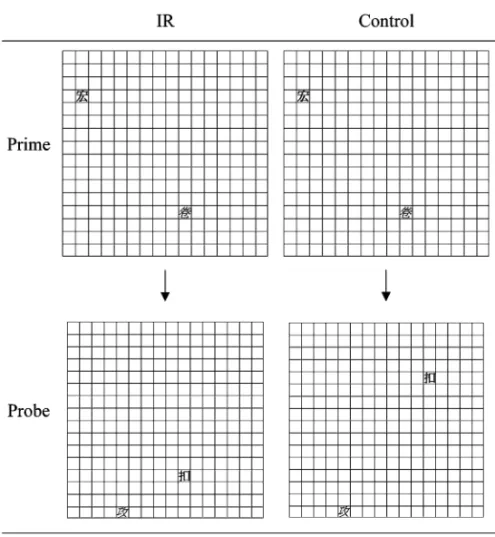
Each block had 48 couplets, in which 24 couplets were IR trials and 24 couplets were control trials. The participants could rest after every 24 couplets. All the couplets within a block were presented in random order. In the control couplets, the targets and the distractors of prime and probe trials occupied different locations. In the IR cou-plets, the location of a prime distractor was the location of the probe target, whereas the prime target and the probe distractor occupied different, unrelated locations. The words of a couplet were different from each other (see Figure 1 for examples of an IR couplet and a control couplet). They were randomly selected in each couplet, with the constraint that each word served equally frequently as prime tar-get, prime distractor, probe tartar-get, or probe distractor. They also ap-peared with equal frequency in each condition. Before each block, the participants received eight practice trials. The order of these two blocks was counterbalanced across participants.
Each trial consisted of one red target word and one green distrac-tor word. The participants were instructed to name the red word while ignoring the green word. Both speed and accuracy were em-phasized. The time course of a trial was as follows. A cross at the center of the screen and a “ding” sound were simultaneously
pre-sented for 300 msec as a warning signal. A stimulus display was sub-sequently presented on the screen until the participant spoke the word or until 3,000 msec had elapsed. The next trial began with a delay of 200 msec after the participant had said the word. To elimi-nate any observable distinction between the prime trial and the probe trial for a couplet, there were no procedural differences between prime and probe trials. That is, the response-to-stimulus interval (RSI) between a prime trial and a probe trial of the same couplet was identical to the RSI between a probe trial and the prime trial of the next couplet. To the participants, there was just a series of displays.
Results and Discussion
A total of 5.73% of the trials were excluded from data analysis because of recording failures (e.g., the voice did not register, or the participant coughed) or errors in re-sponding to the prime display. Thus, both the reaction time (RT) and the error rate reflect the performance in correctly naming probe targets contingent on correct re-sponse to the prior prime targets. One participant’s data were excluded because of a high error rate (40.62%). The averaged median RT and the error rate in each con-dition are presented in Table 1.
Median RTs were analyzed with a repeated measures analysis of variance (ANOVA). Only the interaction of couplet type and stimulus type was significant [F(1,23)⫽ 12.08, MSe⫽ 318.28, p ⬍ .01]. Planned contrasts
re-Figure 1. Illustrations of an ignored repetition (IR) couplet and a control cou-plet. The matrix was invisible during the experiments. Chinese character words were presented in this study. The one in bold (the upper character in each panel) was the target, and the one in italics was the distractor. In the experimental dis-play, the target was in red, and the distractor was in green.
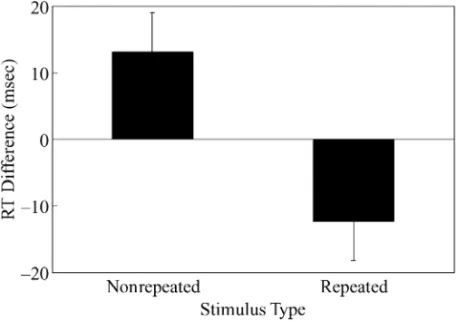
vealed signif icant negative priming in the repeated condition [see Figure 2 for location priming effects; F(1,46)⫽ 4.35, MSe⫽ 415.48, p ⬍ .05]. In contrast, there was a significant positive priming effect in the nonrepeated condition [F(1,46)⫽ 4.91, MSe⫽ 415.48, p⬍ .05]. Error rates were analyzed in the same manner, and no significant effects were observed ( ps⬎ .10).
With an identity discrimination task, Experiment 1 demonstrated reliable location priming effects. More-over, the results paralleled the findings in the domain of identity negative priming (e.g., Strayer & Grison, 1999). Location negative priming was observed only when a small set of locations was repeatedly used. When nonre-peated locations were used, positive priming was found. The results implied that when a distractor occupied a lo-cation, the activation of this distractor location increased, leading to significant positive priming. And only when activation of a distractor location was sufficiently high, due to repetitive use of a small set of locations, was this particular location inhibited, and only then did negative priming manifest.
EXPERIMENT 2A
In Experiment 1, the locations of the targets and dis-tractors were randomly selected. Thus, the distance
be-tween the target and the distractor varied trial by trial. However, the target-to-distractor distance may play a role in location negative priming. To evaluate this issue, Experiments 2A and 2B were conducted to replicate Ex-periment 1 and also to investigate the impact of target-to-distractor distance on location priming.
With repeated locations in an identification task, two studies have shown inconsistent results. Location nega-tive priming was observed in Connelly and Hasher’s (1993) Experiment 2, but not in Tipper, Weaver, and Houghton’s (1994) Experiment 2. The difference in target-to-distractor distance used in their experiments may have caused the inconsistency. Whereas Connelly and Hasher used longer target-to-distractor distances (ranging between 2.5º and 9.5º), Tipper et al. (1994) used shorter target-to-distractor distances (ranging between 1.6º and 2.4º, estimated). We suspected that location neg-ative priming in a naming task might occur only when the target and the distractor are far apart.
The hypothesis that location negative priming occurs only in the far target-to-distractor distance may appear counterintuitive. Near distractors ought to create greater interference than far distractors and, hence, larger nega-tive priming. Yet we postulated this possibility in order to account for the discrepancy in previous findings. When an identification task that involves spatial selection is Table 1
Averaged Median Correct Reaction Times (RTs, in Milliseconds) and Error Rates (%) in Each Condition in Experiment 1
Couplet Type
IR Control
RT Error Rate RT Error Rate
Stimulus Type M SD M SD M SD M SD
Nonrepeated 610.0 37.1 0.7 2.1 623.1 47.0 1.1 2.2
Repeated 624.3 42.8 1.1 1.9 612.0 35.8 1.6 2.7
Note—IR, ignored repetition.
Figure 2. Means and standard errors for location priming effects at nonrepeated and repeated locations in Experiment 1.
considered, the localization process must occur prior to stimulus identification. Thus, two levels of selection may be involved: localization and identification. When the target and the distractor are near to each other, spa-tial attention might be directed to the target–distractor re-gion, with little selection involved in localization. When the target and distractor are far apart, spatial attention zooms in on the target region like a lens (Eriksen & St. James, 1986; Eriksen & Yeh, 1985) and ignores the dis-tractor region. The disdis-tractor region was inhibited in the localization process. Thus, location negative priming was observed when the target–distractor distance was far, but not when the distance was near. Only after a target region was selected would distractors within the region near the target create greater interference in identification.
The impact of target-to-distractor distance on location negative priming with nonrepeated locations was studied in Experiment 2A, and the impact of target-to-distractor distance on location negative priming with repeated lo-cations was studied in Experiment 2B. The same partic-ipants served in both experiments, doing Experiment 2A first. With the same participants, the results can be com-pared more directly. Because the locations used in the nonrepeated condition were also used in the later repeated condition and not vice versa, we ran the nonrepeated block first.
A couplet type (IR/control) ⫻ distance type (short/ long) design was used in each experiment. According to the distance hypothesis proposed earlier and the findings obtained in Experiment 1, location negative priming was expected only in the long-distance condition when re-peated locations were used (Experiment 2B). If one con-siders the fact that positive identity priming from non-repeated stimuli has been shown to be less reliable (e.g., Grison & Strayer, 2001; Strayer & Grison, 1999), it would be of interest to investigate whether the positive priming effect found in Experiment 1 occurs only in the long-distance condition. If the positive priming effect is also the result of a selection process, the priming effect should be observed only when the target and the distrac-tor are far apart.
Method
Participants. Nineteen undergraduate students at National Tai-wan University participated in this experiment for a bonus credit in an introductory psychology course. All the participants were naive as to the purpose of the experiment and had normal or corrected-to-normal vision.
Stimuli. The same materials as those in Experiment 1 were used. For an IR couplet, the location of prime distractor/probe target was first randomly selected. In the short-distance condition, the prime target and the probe distractor were then randomly selected from the cells that were within a range of one to three cells (a visual angle ranging from 1.1º to 4.4º). In the long-distance condition, the prime target and the probe distractor were randomly selected from the cells that were within a range of seven to nine cells (a visual angle ranging between 7.7º and 13.2º). Control couplets were constructed in a similar way as IR couplets, with the exception that another prime distractor location was selected under the same constraint. All locations were selected without replacement.
Procedure. The procedure was identical to that in Experiment 1, with two changes: There was a total of 16 couplets in each condi-tion, and the participants could rest after every 32 couplets.
Results and Discussion
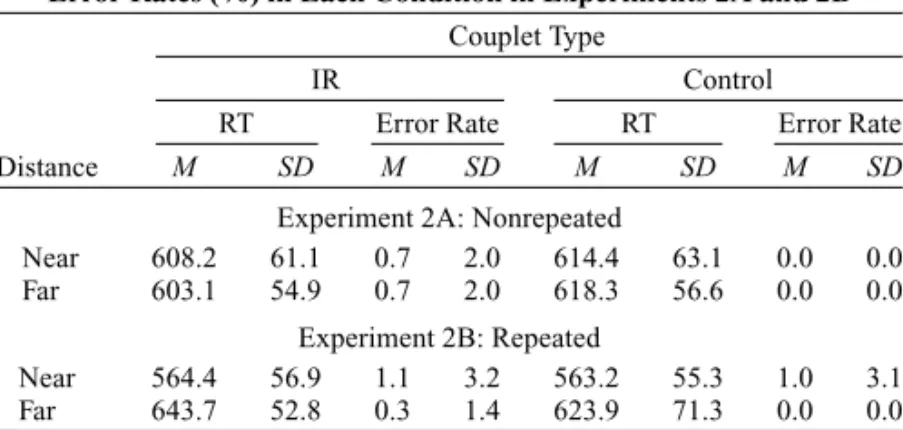
A total of 5.56% of the trials were excluded from data analysis because of recording failures or errors in re-sponding to the prime display. One participant’s data were excluded due to the high error rate (28.13%). The averaged median RT and error rate of each condition are presented in Table 2.
Median RTs were analyzed with an ANOVA with a cou-plet type ⫻ distance type design. Only the main effect of couplet type was significant [F(1,17)⫽ 4.76, MSe⫽ 435.81, p⬍ .05], indicating faster responses in the IR condition. However, planned contrasts revealed that pos-itive priming was significant only in the long-distance condition [see Figure 3 for location priming effects; F(1,34)⫽ 4.20, MSe⫽ 496.84, p ⬍ .05]. Error rates showed a significant main effect of couplet type [F(1,17)⫽ 4.85, MSe⫽ 1.91, p ⬍ .05], with more errors in the IR condition than in the control condition.
The findings of Experiment 1 were partially repli-cated. There was location positive priming, rather than location negative priming, in RTs for nonrepeated loca-tions. Moreover, the priming effect was reliable only in the long-distance condition. This finding is consistent with the distance hypothesis; that is, the priming effect of distractor location in an identity discrimination task occurs when the target and the distractor are located far apart. However, error rates showed location negative priming in the IR condition, regardless of the target– distractor distance. The observation that RT was shorter, although the error rate was higher, in the IR condition was not the result of a speed –accuracy tradeoff, because the overall correlation between these two measures was low (r⫽ ⫺.16).
EXPERIMENT 2B Method
Participants. The participants completed this experiment after finishing Experiment 2A.
Stimuli. The same materials as those in Experiment 1 were used. For the short-distance condition, four locations were preselected to repeat throughout the experiment. The vertical and horizontal dis-tance between one location and the other was 3 cells (a visual angle of 3.3º) in the short-distance condition. The vertical and horizontal distance between one location and the other was 11 cells (a visual angle of 12.1º) in the long-distance condition. The exact coordi-nates of these locations can be found in the Appendix.
Locations of an IR couplet and a control couplet were randomly selected from these preselected locations with replacement. In a control couplet, no item shared the same location. In an IR couplet, the prime distractor and the probe target shared the same location. Procedure. The procedure was identical to that in Experiment 2A, with one exception. Because the participants may have adopted a zooming strategy in localizing a target from a far distractor, since long-distance conditions were more salient when being mixed with short-distance conditions in a block, the two distance conditions
were tested in separate blocks. The results could clarify those for Experiment 1 with a different test context.
Results and Discussion
A total of 4.78% of the trials were excluded from data analysis because of recording failures, or errors in re-sponding to the prime display. The participant whose data were excluded from Experiment 2A, due to a high error rate, was also excluded in this experiment. The averaged median RT and error rate of each condition are presented in Table 2.
Median RTs were analyzed with an ANOVA with a cou-plet type⫻ distance type design. The main effect of cou-plet type was significant [F(1,17)⫽ 5.25, MSe⫽ 380.32, p⬍ .05]. The main effect of distance was also significant [F(1,17)⫽ 57.25, MSe⫽ 1,541.95, p ⬍ .01]. Moreover, the two-way interaction was significant [F(1,17)⫽ 4.40, MSe⫽ 356.11, p ⬍ .05]. Planned contrasts revealed sig-nif icant location negative priming only in the long-distance condition [F(1,34)⫽ 9.64, MSe⫽ 949.03, p ⬍
.01]. Figure 3 shows the priming effects. Error rates did not show any significant results ( ps⬎ .10).
The results replicated those for Experiment 1, demon-strating location negative priming with repeated loca-tions. Moreover, the influence of target-to-distractor dis-tance supported the disdis-tance hypothesis. Location negative priming in an identification task could be found only when the distractor was located away from the target. One possibility is that near distractors generated a sup-pression region, so that items within this region were suppressed.1That is, suppression may have occurred in both the control and the IR conditions. If this were the case, we should find a performance difference in the control condition between near and far manipulations. The results of Experiment 2A did not show any differ-ence, and the results of Experiment 2B showed shorter RTs in the responding to control trials when distractors were near to targets.
Although the distance effect may be surprising, it was also found in another spatial task. In a recent study of a
Figure 3. Means and standard errors for location priming effects at nonre-peated and renonre-peated locations in Experiments 2A and 2B.
Table 2
Averaged Median Correct Reaction Times (RTs, in Milliseconds) and Error Rates (%) in Each Condition in Experiments 2A and 2B
Couplet Type
IR Control
RT Error Rate RT Error Rate
Distance M SD M SD M SD M SD
Experiment 2A: Nonrepeated
Near 608.2 61.1 0.7 2.0 614.4 63.1 0.0 0.0
Far 603.1 54.9 0.7 2.0 618.3 56.6 0.0 0.0
Experiment 2B: Repeated
Near 564.4 56.9 1.1 3.2 563.2 55.3 1.0 3.1
Far 643.7 52.8 0.3 1.4 623.9 71.3 0.0 0.0
Simon-like spatial congruence effect, the positive con-gruent effect could be found when a study letter was far from the screen center, but not when it was near (Zhang & Johnson, 2004). Because the participants in their study were responding to the color, it appears that distance mattered when overt responses to stimulus locations were not required.
Taken together, Experiments 2A and 2B replicated Experiment 1 and further demonstrated the impact of target- to-distractor distance on location priming effects. However, different procedures were used in these two ex-periments: Near and far trials were intermixed in Exper-iment 2A but were presented in separate blocks in Ex-periment 2B. Such a manipulation might introduce some differences between these two experiments. For example, it might be easier to attend to a small region in Experi-ment 2B than in ExperiExperi-ment 2A, because targets and dis-tractors were always near in a block. Nevertheless, the findings raised the possibility that the results of Experi-ment 1 might be affected by the target-to-distractor dis-tance. It would be necessary to examine the effect of loca-tion repetiloca-tion while keeping target-to-distractor distances constant across experimental conditions. We adopted this method in Experiment 3.
EXPERIMENT 3
The objective of this experiment was to demonstrate the transition in priming effects as distractor locations accumulate activation through repetition, as shown in Strayer and Grison’s (1999) Experiment 2 on identity negative priming. In this experiment, their method was adopted by presenting four blocks. Each block had its own unique set of four locations. Moreover, each block was divided into four quartiles, resulting in a couplet type (IR/control) ⫻ quartile (1/2/3/4) design. In the first quartile of a block, each location was novel, with no prior appearance. After repetitive use of the same set of locations across the four quartiles, activation of locations in the fourth quartile would be expected to be quite high. Location negative priming should manifest only in later quartiles, as has been shown with identity negative prim-ing (Strayer & Grison, 1999).
With this design, the evidence should be much stronger in support of the contingency of location negative
prim-ing on location repetition, because only one set of loca-tions was used in each block to accumulate activation across quartiles. In Experiment 1, different sets of stim-ulus locations were used in different blocks. Results might be due to different target-to-distractor distances in the repeated condition and in the nonrepeated condition. As Experiments 2A and 2B demonstrated, distance was also an important factor. Thus, it was necessary to con-duct an experiment keeping target-to-distractor distances constant between the repeated condition and the nonre-peated condition.
Method
Participants. Eighteen undergraduate students at National Tai-wan University participated in this experiment for a bonus credit in an introductory psychology course. All the participants were naive as to the purpose of the experiment and had normal or corrected-to-normal vision.
Stimuli. The same materials as those in Experiment 1 were used. Four sets of four locations were selected, under the constraint that the locations in each set should not be too close to each other, in the range of 6–13 cells apart. The exact coordinates of these locations can be found in the Appendix.
One set of locations was used in each block. Locations of an IR couplet and a control couplet were randomly selected from the as-signed set of locations, with replacement. In a control couplet, no items shared the same location. In an IR couplet, the prime distrac-tor and the probe target shared the same location.
Procedure. The procedure was identical to that in Experiment 1, with the following changes. First, there were 8 couplets in each quar-tile of a block, with IR in 4 and control in the other 4 couplets. All the couplets within a quartile of a block were presented randomly. Pooling these couplets from the four blocks resulted in a total of 16 couplets in each quartile for the IR condition and also for the con-trol condition. Second, there were eight practice trials before the ex-periment. Third, the participants could rest after every 32 couplets.
Results and Discussion
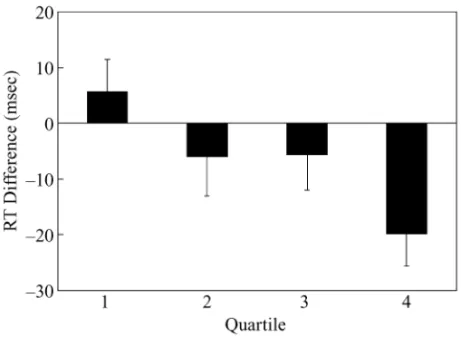
A total of 3.56% of the trials were excluded from data analysis because of recording failure or errors in respond-ing to the prime display. The averaged median RT and error rate for each condition are presented in Table 3. Median RTs were analyzed with an ANOVA with a couplet type ⫻ quartile design. The main effect of cou-plet type was marginally significant [F(1,17)⫽ 3.73, MSe⫽ 394.82, p ⫽ .067]. More important, the inter-action was significant [F(3,51)⫽ 2.96, MSe⫽ 334.08, p⬍ .05]. Planned contrasts revealed significant negative
Table 3
Averaged Median Correct Reaction Times (RTs, in Milliseconds) and Error Rates (%) in Each Condition in Experiment 3
Couplet Type
IR Control
RT Error Rate RT Error Rate
Quartile M SD M SD M SD M SD
1 625.3 56.6 0.0 0.0 631.1 53.8 0.7 2.1
2 629.3 54.8 1.1 2.4 623.3 56.8 0.7 2.0
3 624.8 54.0 1.1 2.4 619.2 55.0 0.3 1.4
4 630.4 59.6 0.4 1.5 610.6 60.0 1.5 3.6
priming in the fourth quartile [see Figure 4 for location priming effects; F(1,68)⫽ 10.11, MSe⫽ 349.27, p ⬍ .01]. Priming effects were not significant in the first three quartiles ( ps⬎ .10). Error rates did not show any significant effects ( ps⬎ .10).
The results of Experiment 3 further confirmed that lo-cation negative priming occurred only when lolo-cations were repeated many times. Such a finding could be ex-plained by assuming the importance of activation in neg-ative priming. When a set of locations was initially in-troduced, the activation level of these locations might have been low. Thus, location negative priming could not be found in the first three quartiles. Since these locations were repeated across the quartiles, activation level might have gradually increased (Strayer & Grison, 1999). As a result, location negative priming was obtained in the fourth quartile.
EXPERIMENT 4
This experiment was conducted to test the effect of IOR (Posner & Cohen, 1984) in the paradigm of location negative priming and also to test the impact of location repetition on IOR. The results of the previous experi-ments in this study showed that location negative prim-ing was contprim-ingent on location repetition, suggestprim-ing that the same processes mediate both identity negative priming and location negative priming. Christie and Klein (2001) have raised the possibility that location negative priming is due to IOR to a stimulated location, rather then reflecting the operation of distractor inhibi-tion. They argued that the reason that an IOR effect has not been observed when the target location has been re-peated in the prime–probe couplet is that confounds have existed in previous studies of location negative priming, so that the contribution of IOR was masked. A major confound, they suggested, has been the use of a biased design where the probability of target repetition was
usu-ally higher than chance level. The participants hence sponded more quickly to the previous target location, re-sulting in positive priming or null results when priming counteracted IOR. Eliminating this bias, they found a cost when target location repeated from the prime to the probe trial. The present experiment incorporated Christie and Klein’s design to test how location repetition influences the effect of target repetition and distractor repetition. This experiment followed Christie and Klein’s (2001) design. A condition of target repetition was incorporated to test whether there was a cost or a benefit in this con-dition. Moreover, prime–probe location contingency was eliminated so that the influence of IOR could not be masked. Seven types of prime–probe location repetition were used (type of repetition): the D-to-T condition, the T-to-D condition, the switch condition, the T-to-T condi-tion, the D-to-D condicondi-tion, the intact condicondi-tion, and the control condition. In the D-to-T condition, which is the traditional IR condition, the location of the probe target was the location of the prime distractor. In the T-to-D condition, where greater interference from the probe dis-tractor was expected (Neumann & DeSchepper, 1991; Stadler & Hogan, 1996), the location of a probe distrac-tor was the location of a prime target. The switch condi-tion was the combinacondi-tion of the D-to-T condicondi-tion and the T-to-D condition—that is, the target and the distractor locations were exchanged from prime to probe. In the T-to-T condition, the location of a probe target repeated the location of a prime target. Either positive priming (Neumann & DeSchepper, 1991; Stadler & Hogan, 1996) or negative priming (Christie & Klein, 2001) was ex-pected. Both Neumann and DeSchepper (1991) and Stadler (1996) and Hogan studied identity priming, whereas Christie and Klein studied location priming effects. In the D-to-D condition, the location of the probe distractor repeated the location of the prime distractor. The intact condition was the combination of the T-to-T condition and the D-to-D condition—that is, the target and distractor Figure 4. Means and standard errors for location priming effects in
locations remained the same from prime to probe. Finally, the control condition was the baseline for the other condi-tions to be compared with. When unspecified, the other location in the display was unrelated to the prime’s loca-tion. Because there were four locations in each block, the ratio of these seven conditions was 2:2:1:2:2:1:2, re-spectively. Thus, the probability of the probe target and the distractor appearing at all possible locations was equal in a given block.
To achieve this design, more trials were required in a segment in this experiment than in Experiment 3. Thus, there were two segments (first half and second half ), in-stead of four quartiles, in a given block. By the combina-tion of Christie and Klein’s (2001) method and the method used in Experiment 3, the impact of location repetition on naming a probe target situated at the prime target location and at the prime distractor location could be evaluated.
Method
Participants. Twenty-two undergraduate students at National Tai-wan University participated in this experiment for a bonus credit in an introductory psychology course. All the participants were naive as to the purpose of the experiment and had normal or corrected-to-normal vision. Data from 2 participants were excluded because of recording failures.
Stimuli. The same sets of stimuli as those selected in Experiment 3 were used.
Procedure. The procedure was identical to that in Experiment 3, with the following changes. First, there were 2 D-to-T, 2 T-to-D, 2 T-to-T, 2 D-to-D, 2 control, 1 intact, and 1 switch couplet in each segment of a block. All the couplets in a segment were presented randomly. Pooling these couplets from the four blocks resulted in a total of 8 couplets for the conditions that repeated twice in each seg-ment and a total of 4 couplets in each segseg-ment for the switch con-dition and the intact concon-dition. Second, the participants could rest after every 24 couplets.
Results and Discussion
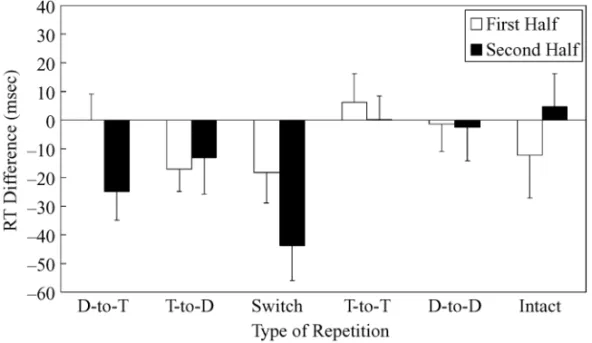
A total of 2.63% of the trials were excluded from data analysis because of recording failures or errors in re-sponding to the prime display. The averaged median RT and error rate for each condition are presented in Table 4. Following Christie and Klein’s (2001) method, sepa-rate F tests were conducted to examine the difference in median RTs between one experimental condition and its control condition (see Figure 5 for location priming ef-fects). Responses in the D-to-T condition were slower
than those in the control condition in the second half [F(1,19) ⫽ 6.27, MSe ⫽ 989.11, p ⬍ .05], but not in the first half ( p⬎ .10). By conducting a 2 (D-to-T vs. control) ⫻ 2 (first vs. second) ANOVA, this observation was sup-ported by the significant two-way interaction [F(1,19)⫽ 4.34, MSe⫽ 715.90, p ⬍ .05]. Responses in the T-to-D condition were significantly slower than those in the control condition in the first half [F(1,19)⫽ 4.82, MSe⫽ 605.41, p⬍ .05], but not in the second half ( p ⬎ .10). The 2 (T-to-D vs. control) ⫻ 2 (first vs. second) ANOVA revealed no significant interaction ( p⬎ .20). Responses in the switch condition were marginally slower than those in the control condition in the first half [F(1,19)⫽ 2.90, MSe⫽ 1,140.70, p ⫽ .10] and were significantly slower in the second half [F(1,19)⫽ 13.42, MSe⫽ 1,432.74, p⬍ .005]. The 2 (switch vs. control) ⫻ 2 (first vs. second) ANOVA revealed no significant interaction ( p⬎ .10). Other comparisons were not significant ( ps⬎ .10). Error rates were also analyzed in the same manner, with no sig-nificant differences (α ⫽ .05) observed.
The contingency of location negative priming on lo-cation repetition was replicated. Responses in the D-to-T condition were slower than those in the control condition only in the second half (24.9 msec), but not in the first half (0.0 msec). Interestingly, the enhanced interference effect in the T-to-D condition, in which the probe dis-tractor location was the prime target’s location, was mar-ginally influenced by location repetition. RTs were sig-nif icantly longer than those in the control condition in the first half (17.1 msec) and also showed a trend (13.0 msec) in the second half. The priming effect in the switch condition appears to be equal to or slightly larger than the summation of the negative priming effect and the enhanced interference effect (see also Chao, Yeh, & Yang, 2003; Stadler & Hogan, 1996; Yeh & Chao, 2004), with a difference of 18.2 msec in the first half and a dif-ference of 43.9 msec in the second half.
The T-to-T condition (and also the intact condition) did not differ from the control condition. Thus, neither the prediction of positive priming (Neumann & DeSchepper, 1991; Stadler & Hogan, 1996) nor the prediction of neg-ative priming (Christie & Klein, 2001) was supported. The null result perhaps was due to a balance of a facili-tatory effect and an inhibitory effect of location repeti-tion. Because participants may need to move their eyes
Table 4
Averaged Median Correct Reaction Times (in Milliseconds) and Error Rates (%) in Each Condition in Experiment 4
Type of Repetition
D-to-T T-to-D Switch T-to-T D-to-D Intact Control
Half M SD M SD M SD M SD M SD M SD M SD Reaction Times 1st 604.8 76.1 621.9 71.8 623.0 62.1 598.6 62.3 606.3 74.0 617.1 77.0 604.9 72.6 2nd 625.7 71.2 613.7 58.0 644.6 82.5 600.5 67.3 603.5 72.6 596.2 77.0 600.8 82.2 Error Rates 1st 0.6 2.8 1.3 4.1 0.0 0.0 0.6 2.8 0.0 0.0 1.3 5.6 1.5 4.8 2nd 1.9 4.6 0.0 0.0 1.3 5.6 1.3 3.8 0.0 0.0 0.0 0.0 1.3 3.8
to fixate on the target in a large space in order to name it, their eyes may still fixate on the target location in the T-to-T condition, given that the RSI duration from re-sponse to the prime target to the initiation of the probe trial was only 200 msec. Because of eye fixation, posi-tive priming occurred to mask the IOR effect.2 Alterna-tively, the stimulus onset asynchrony (SOA) in the pres-ent experimpres-ent (500 msec plus varied RTs) may not be sufficient for the inhibitory effect to manifest when the task demand was to identify a Chinese character. It has been shown that IOR with a discrimination task mani-fested at a longer SOA than did IOR with a detection task (e.g., Lupiáñez, Milán, Tornay, Madrid, & Tudela, 1997).
GENERAL DISCUSSION
The results of this study reliably demonstrated that lo-cation negative priming in an identity discrimination task also depends on location repetition, as stimulus rep-etition is key in identity negative priming (Grison & Strayer, 2001; Malley & Strayer, 1995; Strayer & Gri-son, 1999). When locations had not been repeated many times, negative priming was not observed, with null re-sults in the first quartile of Experiment 3 and in the first half of Experiment 4 and positive priming effects in Ex-periments 1 and 2. Moreover, both a location positive priming effect and a location negative priming effect oc-curred only when a target and a distractor were far apart. When a target and a distractor were near, priming effects were not observed.
Because both location and identity negative priming are contingent on repeated stimuli, it is likely that the same process underlies both types of negative priming, whether it is a central inhibitory mechanism in the frontal lobe (see Koshino et al., 2000) or the retrieval process instigated by a familiar stimulus. The ways in which the
attention and the memory views of negative priming would explain these results will be explored.
First, to explain the contingency of negative priming on the presence of probe distractors, Tipper and Cranston (1985) suggested that the knowledge of the need for se-lection in probe trials drives the maintenance of a selec-tion state. When a selecselec-tion state is absent, the inhibitory process is not maintained, and negative priming cannot manifest. It is likely that the high level of competition from distractors encourages maintenance of a selection state. As a result, negative priming is contingent on re-peated stimuli.
Second, even when a selection state is in operation, the inhibitory process may not function when a prime distractor is not competitive. When a distractor location appears the first time, it receives activation from the ex-posure. However, the transient activation from one pre-sentation appears to be insufficient to compete with the target, and hence, inhibition does not function (Malley & Strayer, 1995; Strayer & Grison, 1999). Without the op-eration of the inhibitory process, the transient activation provides a higher baseline when a distractor becomes the probe target. In the nonrepeated condition, in which the probe distractor location is also novel, the probe target with a higher baseline of activation can win the race in selection, according to Houghton and Tipper’s (1994) model of inhibitory process in selective attention. As lo-cations repeat in a quartile or in a block, the activation level at these locations increases, and the distractors be-come strong competitors against the target. The inhibitory process operates to protect the target, and location nega-tive priming manifests.
Strong competition may arise from the statistical prop-erties involved in the assigning of target and distractor locations. Reder, Weber, Shang, and Vanyukov (2003) investigated location negative priming by varying the Figure 5. Means and standard errors for location priming effects in each condition in
frequency of a distractor at each location to be never, rare, medium, or frequent. They found larger location negative priming in the probe trials when a target occu-pied a rare distractor location, as compared with the fre-quent and medium conditions. More important, a rare distractor location led to greater interference in the prime trials. The greater interference suggested that the rare distractor location was more competitive in the selection process. The rare distractor location was competitive, perhaps, for two reasons: its relative salience from its low frequency and its higher activation. As was described in the methodology section, the absolute number of dis-tractors appearing in each location was manipulated across conditions, whereas the absolute number of targets in each location was constant, except in the never condi-tion. If one assumes that activation comes from being a target location and inhibition from being a distractor lo-cation, the rare location received the same amount of ac-tivation but less inhibition. Thus, acac-tivation of the rare location could be higher than that of the frequent and medium locations.
Third, probe competition might also play a role in neg-ative priming, because probe distractor locations in the four experiments were also varied with location repeti-tion. Houghton and Tipper’s (1994) model implicated a role for probe competition by proposing that negative priming occurs because the target’s activation level is low from inhibition, as compared with the probe dis-tractor. Yeh and Chao (2004) demonstrated the impor-tance of probe competition in identity negative priming. With a small set of stimuli, they either grouped a target and two distractors with a rectangle or did not group stimuli in probe trials. Negative priming manifested only when the target and its distractors were grouped together. Thus, activation of probe distractors modulated by per-ceptual grouping was critical to the manifestation of neg-ative priming.
One result defies the inhibition view of negative prim-ing. Neill et al.’s (1994) study showed that location negative priming could occur without the presence of probe distractors. However, a recent study by Buckolz, Boulougouris, and Khan (2002) demonstrated significant but reduced location negative priming when the probe distractor was removed. Furthermore, with the fore-knowledge that no distractor existed in the probe trial, lo-cation negative priming was eliminated with the removal of the probe distractor. Thus, strategic factors may mod-ulate the role of a probe distractor in negative priming. Clearly, we assumed the involvement of a selection pro-cess in negative priming when using the models from se-lective attention to account for the contingency on loca-tion repetiloca-tion. Yet negative priming can occur when prime displays contain only one stimulus (e.g., Christie & Klein, 2001; Milliken, Joordens, Merikle, & Seiffert, 1998). Furthermore, the necessity of a selection process can be challenged if location negative priming simply re-flects IOR (Christie & Klein, 2001; Milliken et al., 2000).3 It is for future research to verify whether location negative
priming is simply a derivative of IOR. We operationally define selection as selective processing of a stimulus when distractors are absent and concurrently suppressing distractors when distractors are highly competitive.
Alternatively, memory retrieval has been proposed to account for negative priming (Neill, 1997; Neill & Mathis, 1998; Neill et al., 1992). This theoretical view can also ac-count for the results of our experiments. Neill and Joor-dens (2002) suggested that there are two processes under-lying the processing of distractors. One is the retrieval of a memory representation, which can lead to negative priming when it contains inappropriate information for the current behavioral goal, and the other is repetition, which can result in positive priming. These two forces counteract each other. When nonrepeated items of low activation are used, their activation can still be increased, and hence, stimulus repetition can produce positive priming. Because positive priming masks negative prim-ing, novel stimuli fail to produce negative priming. When repeated stimuli of high activation are used, their activation cannot be increased further, and positive prim-ing is weak. Because the weak positive primprim-ing cannot fully mask negative priming, negative priming mani-fests. Thus, the importance of stimulus repetition in neg-ative priming was explained. In response to this postula-tion, Strayer, Drews, and Albert (2002) suspected that positive priming could not mask negative priming. Fur-thermore, they found no evidence for implicit perceptual priming of distractors, failing to support the existence of positive priming. It is still uncertain whether there is a positive priming effect that can mask negative priming. Thus, the results of the present study could be explained with an inhibition view, a memory retrieval view, or an integrated view that incorporates both inhibition and memory retrieval mechanisms (e.g., Tipper, 2001).
In summary, this study demonstrated that location negative priming in an identity discrimination task was contingent on location repetition, similar to what has been found in identity negative priming. This finding suggests that a common mechanism underlies both loca-tion negative priming and identity negative priming. The results of this study, however, left the contribution of IOR in location negative priming an open question. Be-cause our participants might have learned the contin-gency in the assignment of target location and, hence, fixated their eyes on the target location when it repeated at the same position, it is for future research to resolve the puzzle.
REFERENCES
Abad, M. J. F., Noguera, C., & Ortells, J. J. (2003). Influence of prime–target relationship on semantic priming effects from words in a lexical-decision task. Acta Psychologica, 113, 283-295.
Broadbent, D. E. (1958). Perception and communication. Oxford: Pergamon.
Buckolz, E., Boulougouris, A., & Khan, M. (2002). Influence of probe-trial selection on the location negative priming effect.
Cana-dian Journal of Experimental Psychology, 56, 273-282.
Disengaging the negative priming mechanism in location tasks.
Eu-ropean Journal of Cognitive Psychology, 14, 207-225.
Chao, H.-F., Yeh, Y.-Y., & Yang, T.-Y. (2003). Interference and inhi-bition of distractors. Chinese Journal of Psychology, 45, 361-377. Christie, J., & Klein, R. M. (2001). Negative priming for spatial lo-cations? Canadian Journal of Experimental Psychology, 55, 24-38. Connelly, S. L., & Hasher, L. (1993). Aging and the inhibition of spa-tial location. Journal of Experimental Psychology: Human
Percep-tion & Performance, 19, 1238-1250.
Dalrymple-Alford, E. C., & Budayr, B. (1966). Examination of some aspects of the Stroop color–word test. Perceptual & Motor
Skills, 23, 1211-1214.
Dark, V. J., & Schmidt, P. A. (2000). Conceptual activation of distrac-tors during selection is not sufficient to produce negative priming.
Psychonomic Bulletin & Review, 7, 126-132.
Deutsch, J. A., & Deutsch, D. (1963). Attention: Some theoretical considerations. Psychological Review, 70, 80-90.
Eriksen, C. W., & St. James, J. D. (1986). Visual attention within and around the field of focal attention: A zoom lens model. Perception &
Psychophysics, 40, 225-240.
Eriksen, C. W., & Yeh, Y.-Y. (1985). Allocation of attention in the vi-sual field. Journal of Experimental Psychology: Human Perception
& Performance, 11, 583-597.
Forster, K. I., & Forster, J. C. (2003). DMDX: A Windows display program with millisecond accuracy. Behavior Research Methods,
In-struments, & Computers, 35, 116-124.
Fox, E. (1996). Cross-language priming from ignored words: Evidence for a common representational system in bilinguals. Journal of
Mem-ory & Language, 35, 353-370.
Grison, S., & Strayer, D. L. (2001). Negative priming and perceptual fluency: More than what meets the eye. Perception & Psychophysics, 63, 1063-1071.
Houghton, G., & Tipper, S. P. (1994). A model of inhibitory mecha-nisms in selective attention. In D. Dagenbach & T. H. Carr (Eds.),
In-hibitory processes in attention, memory, and language (pp. 53-112).
San Diego: Academic Press.
Koshino, H., Boese, G. A., & Ferraro, F. R. (2000). The relationship be-tween cognitive ability and positive and negative priming in identity and spatial priming tasks. Journal of General Psychology, 127, 372-382. Kramer, A. F., & Strayer, D. L. (2001). Influence of stimulus
repeti-tion on negative priming. Psychology & Aging, 16, 580-587. Lowe, D. G. (1979). Strategies, context, and the mechanism of response
inhibition. Memory & Cognition, 7, 382-389.
Lupiáñez, J., Milán, E. G., Tornay, F. J., Madrid, E., & Tudela, P. (1997). Does IOR occur in discrimination tasks? Yes, it does, but later. Perception & Psychophysics, 59, 1241-1254.
Malley, G. B., & Strayer, D. L. (1995). Effect of stimulus repetition on positive and negative identity priming. Perception &
Psycho-physics, 57, 657-667.
Milliken, B., Joordens, S., Merikle, P. M., & Seiffert, A. E. (1998). Selective attention: A reevaluation of the implications of negative priming. Psychological Review, 105, 203-229.
Milliken, B., Tipper, S. P., Houghton, G., & Lupiáñez, J. (2000). Attending, ignoring, and repetition: On the relation between negative priming and inhibition of return. Perception & Psychophysics, 62, 1280-1296.
Ministry of Education Committee for the Promotion and Prop-agation of the National Language (1997). The frequency
statis-tics of the words and phrases in the compiled materials of the Con-cise Mandarin Dictionary. Taipei, Taiwan: Ministry of Education.
Neill, W. T. (1977). Inhibitory and facilitatory processes in selective attention. Journal of Experimental Psychology: Human Perception
& Performance, 3, 444-450.
Neill, W. T. (1997). Episodic retrieval in negative priming and repeti-tion priming. Journal of Experimental Psychology: Learning,
Mem-ory, & Cognition, 23, 1291-1305.
Neill, W. T., & Joordens, S. (2002). Negative priming and multiple
repetition: A reply to Grison and Strayer (2001). Perception &
Psychophysics, 64, 855-860.
Neill, W. T., & Mathis, K. M. (1998). Transfer-inappropriate process-ing: Negative priming and related phenomena. In D. L. Medin (Ed.),
The psychology of learning and motivation: Advances in research and theory (Vol. 38, pp. 1-44). San Diego: Academic Press.
Neill, W. T., Terry, K. M., & Valdes, L. A. (1994). Negative priming without probe selection. Psychonomic Bulletin & Review, 1, 119-121. Neill, W. T., Valdes, L. A., Terry, K. M., & Gorfein, D. S. (1992). Persistence of negative priming: II. Evidence for episodic trace re-trieval. Journal of Experimental Psychology: Learning, Memory, &
Cognition, 18, 993-1000.
Neumann, E., & DeSchepper, B. G. (1991). Costs and benefits of tar-get activation and distractor inhibition in selective attention. Journal
of Experimental Psychology: Learning, Memory, & Cognition, 17,
1136-1145.
Posner, M. I., & Cohen, Y. (1984). Components of visual orienting. In H. Bouma & D. G. Bouwhuis (Eds.), Attention and performance X:
Control of language processes (pp. 531-556). Hillsdale, NJ: Erlbaum.
Reder, L. M., Weber, K., Shang, J., & Vanyukov, P. M. (2003). The adaptive character of the attentional system: Statistical sensitivity in a target localization task. Journal of Experimental Psychology: Human
Perception & Performance, 29, 631-649.
Stadler, M. A., & Hogan, M. E. (1996). Varieties of positive and neg-ative priming. Psychonomic Bulletin & Review, 3, 87-90.
Strayer, D. L., Drews, F. A., & Albert, R. W. (2002). Negative prim-ing and stimulus repetition: A reply to Neill and Joordens (2002).
Perception & Psychophysics, 64, 861-865.
Strayer, D. L., & Grison, S. (1999). Negative identity priming is con-tingent on stimulus repetition. Journal of Experimental Psychology:
Human Perception & Performance, 25, 24-38.
Sullivan, M. P., & Faust, M. E. (1993). Evidence for identity inhibi-tion during selective atteninhibi-tion in old adults. Psychology & Aging, 8, 589-598.
Tipper, S. P. (1985). The negative priming effect: Inhibitory priming by ignored objects. Quarterly Journal of Experimental Psychology, 37A, 571-590.
Tipper, S. P. (2001). Does negative priming reflect inhibitory mecha-nisms? A review and integration of conflicting views. Quarterly
Journal of Experimental Psychology, 54A, 321-343.
Tipper, S. P., Brehaut, J. C., & Driver, J. (1990). Selection of mov-ing and static objects for the control of spatially directed action.
Jour-nal of Experimental Psychology: Human Perception & Performance, 16, 492-504.
Tipper, S. P., & Cranston, M. (1985). Selective attention and priming: Inhibitory and facilitatory effects of ignored primes. Quarterly
Jour-nal of Experimental Psychology, 37A, 591-611.
Tipper, S. P., & Driver, J. (1988). Negative priming between pictures and words in a selective attention task: Evidence for semantic pro-cessing of ignored stimuli. Memory & Cognition, 16, 64-70. Tipper, S. P., Weaver, B., & Houghton, G. (1994). Behavioural goals
determine inhibitory mechanisms of selective attention. Quarterly
Journal of Experimental Psychology, 47A, 809-840.
Verhaeghen, P., & De Meersman, L. (1998). Aging and the negative priming effect: A meta-analysis. Psychology & Aging, 13, 435-444. Yeh, Y.-Y., & Chao, H.-F. (2004). Probe distractors can influence neg-ative priming by perceptual grouping. Perception & Psychophysics, 66, 208-218.
Zhang, J. X., & Johnson, M. K. (2004). A memory-based, Simon-like, spatial congruence effect: Evidence for persisting spatial codes.
Quarterly Journal of Experimental Psychology, 57A, 419-436.
NOTES 1. We thank John Christie for suggesting this. 2. We thank John Christie for providing this idea.
APPENDIX
Coordinates of the Repeated Locations Used in Each Experiment Coordinates Experiment 1 (2, 13), (5, 1), (10, 4), (12, 12) Experiment 2B Short (7, 7), (10, 7), (7, 10), (10, 10) Long (3, 3), (3, 14), (14, 3), (14, 14) Experiment 3 Block 1 (1, 9), (6, 1), (15, 4), (11, 10) Block 2 (5, 7), (12, 3), (16, 10), (7, 14) Block 3 (2, 4), (9, 7), (11, 14), (4, 14) Block 4 (6, 4), (12, 6), (13, 16), (5, 11) Note—The coordinates of the cell in the lower left corner are (0, 0). The first number refers to cell number on the horizontal axis, and the second number refers to cell number on the vertical axis.
(Manuscript received September 15, 2003; revision accepted for publication October 4, 2004.)