Elsevier Editorial System(tm) for Ophthalmology Manuscript Draft
Manuscript Number: 2010-181R2
Title: Genome-wide association study of diabetic retinopathy in a Taiwanese population Article Type: Manuscript
Corresponding Author: Prof Fuu-Jen Tsai, Corresponding Author's Institution: First Author: Yu-Chuen Huang, Ph.D.
Order of Authors: Yu-Chuen Huang, Ph.D.; Jane-Ming Lin, MD; Hui-Ju Lin, MD, PhD; Ching-Chu Chen, MD; Shih-Yin Chen, PhD; Chang-Hai Tsai, MD, PhD; Fuu-Jen Tsai, MD, PhD
Abstract: Purpose: Diabetic retinopathy (DR) is a microvascular complication of diabetes with a complex multifactorial pathogenesis. The aim of this study was to identify the susceptibility genes that increase the risk of DR in type 2 diabetes (T2D) and to further elucidate the underlying mechanism of DR pathogenesis.
Design: A case-control study.
Participants: 749 unrelated individuals with T2D (174 with DR and 575 without DR) and 100 non-diabetic controls.
Methods: We conducted a genome-wide association study using Illumina HumanHap550-Duo BeadChips.
Main outcome Measures: Compared with the genotypic distribution of single nucleotide polymorphism (SNPs) between subjects with DR and without DR.
Results: Using statistical models, we selected a total of 12 SNPs with p- -6 that were associated with DR. After controlling for diabetes duration and hemoglobin A1C, nine of the 12 SNPs located on five chromosomal regions were found to associated with DR. Five loci not previously
associated with DR susceptibility were identified in and around the following genes: MYSM1 (Myb-like, SWIRM and MPN domains 1) located on chromosome 1p (odds ratio [OR] = 1.50, 95% confidence interval [CI] = 1.03-2.20); PLXDC2 (plexin domain-containing 2) located on the chromosome 10p (OR=1.67, 95% CI=1.06-2.65); ARHGAP22 (Rho GTPase-activating protein 22) located on chromosome 10q (OR = 1.65, 95% CI = 1.05-2.60) and HS6ST3 (heparan sulfate 6-O-sulfotransferase 3) located on chromosome 13q (OR = 2.33, 95% CI = 1.13-4.77). The SNPs rs13163610 and rs17376456 located in the unknown gene on chromosome 5q were also associated with DR (OR = 3.63, 95% CI = 1.38-9.58). Conclusions: We identified a genetic association for susceptibility to DR in five novel chromosomal regions and PLXDC2 and ARHGAP22, the latter two of which are genes implicated in endothelial cell angiogenesis and increased capillary permeability. These findings suggest unsuspected pathways in the pathogenesis of DR.
Ref.: Manuscript 2010-181R1
Genome-wide association study of diabetic retinopathy in a Taiwanese population
Ophthalmology
Dear Prof Tsai,
I am pleased to inform you that your Manuscript entitled, "Genome-wide association
study of diabetic retinopathy in a Taiwanese population," has been accepted for
publication in Ophthalmology, pending your addressing the following:
Change from:
Five loci not previously associated with DR susceptibility were identified in and
around the following genes: MYSM1 (Myb-like, SWIRM and MPN domains 1)
located on chromosome 1p [odds ratio (OR) = 1.50, 95% confidence interval (CI) =
1.03-2.20];
to:
Five loci not previously associated with DR susceptibility were identified in and
around the following genes: MYSM1 (Myb-like, SWIRM and MPN domains 1)
located on chromosome 1p (odds ratio [OR] = 1.50, 95% confidence interval [CI] =
1.03-2.20)…
Review other instances of use of brackets. Use brackets to represent parentheses
within parentheses and not the other way around. Numerous errors to correct.
Response: We have made this correction in the revised manuscript.
Use commas and not semicolons for consecutive reference callouts (e.g., 6, 7 and
Response: We have made this correction in the revised manuscript.
p. 18 - add date accessed for website.
Response: We have added this information in the text, as follows
Data from the SNP database (dbSNP BUILD 131; http://www.ncbi.nlm.gov/SNP/)
Submit each table as a separate file.
Response: We would like to upload a separate table file while submitting the revised
manuscript.
Define SD, NPDR, SNP ID, MPN, DR (table 2) in tables. Review for others.
Response: We have made this correction in the revised manuscript.
Please do not highlight changes in revision.
With apologies, we currently have very limited space for both print and online pages.
I need to ask authors to help by removing tables or figures since they take so much
room to print. Please submit tables 3 and 4 in PDF format for online only publication.
Submit each table as a separate file. The PDF files will not be typeset or reformatted
in any way by the publisher. Thank you for your understanding and assistance.
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
Genome-wide association study of diabetic retinopathy in a
Taiwanese population
Yu-Chuen Huang, PhD,1,6 Jane-Ming Lin, MD,2,6 Hui-Ju Lin, MD, PhD,2,6 Ching-Chu
Chen, MD,3,6 Shih-Yin Chen, PhD,1,6 Chang-Hai Tsai, MD, PhD,8 and Fuu-Jen Tsai,
MD, PhD1,4,5,7
1
Genetics Center, Department of Medical Research, 2Department of Ophthalmology,
3
Division of Endocrinology and Metabolism, Department of Medicine, 4Department
of Pediatrics, 5Department of Medical Genetics, China Medical University Hospital,
Taichung 404, Taiwan, 6School of Chinese Medicine, 7School of Post Baccalaureate
Chinese Medicine, College of Chinese Medicine, China Medical University,
Taichung 404, Taiwan, 8Asia University, Taichung 413, Taiwan
Funding: This research was supported by China Medical University Hospital,
Taichung, Taiwan (DMR92-076 and DMR93-017), and the National Research
Program for Genomic Medicine from National Science Council, Taipei, Taiwan, as
well as the National Clinical Core for Genomic Medicine at Academia Sinica, Taipei,
Taiwan (NSC96-3112-B-001-010).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
Conflict of Interest: None of the authors have any financial interests to disclose.
Short title: Genome-wide association study of diabetic retinopathy
Corresponding author: Prof. Fuu-Jen Tsai, MD, PhD, Department of Medical
Research, China Medical University Hospital, No. 2, Yuh-Der Road, Taichung 404,
Taiwan.
e-mail: [email protected]
This article contains online-only material. The following should appear
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 Abstract
Purpose: Diabetic retinopathy (DR) is a microvascular complication of diabetes with
a complex multifactorial pathogenesis. The aim of this study was to identify the
susceptibility genes that increase the risk of DR in type 2 diabetes (T2D) and to
further elucidate the underlying mechanism of DR pathogenesis.
Design: A case-control study.
Participants: 749 unrelated individuals with T2D (174 with DR and 575 without DR)
and 100 non-diabetic controls.
Methods: We conducted a genome-wide association study using Illumina
HumanHap550-Duo BeadChips.
Main outcome Measures: Compared with the genotypic distribution of single
nucleotide polymorphism (SNPs) between subjects with DR and without DR.
Results: Using statistical models, we selected a total of 12 SNPs with p-values <1 10–6 that were associated with DR. After controlling for diabetes duration and
hemoglobin A1C, nine of the 12 SNPs located on five chromosomal regions were
found to associated with DR. Five loci not previously associated with DR
susceptibility were identified in and around the following genes: MYSM1 (Myb-like,
SWIRM and MPN domains 1) located on chromosome 1p (odds ratio [OR] = 1.50,
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
located on the chromosome 10p (OR=1.67, 95% CI=1.06-2.65); ARHGAP22 (Rho
GTPase-activating protein 22) located on chromosome 10q (OR = 1.65, 95% CI =
1.05–2.60) and HS6ST3 (heparan sulfate 6-O-sulfotransferase 3) located on
chromosome 13q (OR = 2.33, 95% CI = 1.13–4.77). The SNPs rs13163610 and
rs17376456 located in the unknown gene on chromosome 5q were also associated
with DR (OR = 3.63, 95% CI = 1.38–9.58).
Conclusions: We identified a genetic association for susceptibility to DR in five
novel chromosomal regions and PLXDC2 and ARHGAP22, the latter two of which are
genes implicated in endothelial cell angiogenesis and increased capillary permeability.
These findings suggest unsuspected pathways in the pathogenesis of DR.
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 Introduction
Diabetes mellitus, of which more than 95% cases worldwide are attributed to
type 2 diabetes mellitus (T2D), has a complex multifactorial pathogenesis.1 The
devastating complications of diabetes are mostly macro- and microvascular diseases,
which are the major causes of morbidity and early mortality in diabetes.2
Diabetic retinopathy (DR) is the commonest microvascular complication of
diabetes and is a leading cause of new-onset blindness in the working-age population
in the developed countries.3-5 The pathogenesis of DR is believed to be complex and
have multifactorial biochemical causes. Alteration of glucose metabolism is the
primary cause of DR.6,7 Poor glycemic control and prolonged duration of diabetes are
other major risk factors in the development of DR.7,8 In addition, there is increasing
evidence to implicate genetic factors in the susceptibility to DR independent of
glycemic control and the duration of diabetes.9,10
To date, genome-wide linkage studies have been performed to identify
susceptibility loci for DR. In Pima Indians with T2D, these studies have not only
identified the susceptibility loci for diabetes and obesity11 but have also reported weak
evidence for linkage of DR to regions of chromosomes 3 and 9,12 and suggestive
evidence for linkage of DR to chromosome 1p36.13 Furthermore, in a study of
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
most strongly associated with any severity of DR.14 In addition, numerous candidate
gene association studies have shown a significant association between genetic factors
and the development of DR, including the involvement of the following: aldose
reductase (AKR1B1),15-20 which is involved in the polyol pathway and catalyzes the
NADPH-dependent reduction of glucose to sorbitol, and is particularly active under
hyperglycemic conditions;21 vascular endothelial growth factor (VEGF),22-29 which
promotes angiogenesis and is a potent mediator of microvascular permeability;30
transforming growth factor-beta (TGF-),31-34 which plays an important role in stimulating angiogenesis and inhibiting endothelial function in the eye;35 endothelial
nitric oxide synthase (NOS3);26,36-38 nitric oxide synthase 2A (NOS2A);39
erythropoietin40; integrin (ITGA2);41,42 and intercellular cell adhesion molecule 1 (ICAM1)43,44 [for a detailed review, see Abhary et al.45]. Identification of the
specific genetic risk factors for DR susceptibility will contribute to developing new
treatments and help to improve existing screening methods.
A considerable amount of work has focused on dissecting the genetics of
diabetes itself; however, fewer studies have been conducted on the molecular
mechanisms leading to its specific complications such as DR. In order to identify
susceptibility loci that are associated with T2D retinopathy in Taiwanese population,
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
retinopathy and 575 without retinopathy) and 100 non-diabetic controls, and
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 Methods Subjects
The study involved 749 unrelated individuals with T2D over the age of 20 years,
who were recruited from the China Medical University Hospital (CMUH), Taichung,
Taiwan. Subjects were diagnosed using the American Diabetic Association Criteria.
Subjects with type 1 diabetes, gestational diabetes, or maturity-onset diabetes of the
young were excluded from this study. Of this group, 174 T2D subjects were
diagnosed with DR: 102 (58.6%) with non-proliferative diabetic retinopathy (NPDR)
and 72 (41.4%) with proliferative diabetic retinopathy (PDR). All T2D subjects
underwent a complete ophthalmologic examination, including corrected visual acuity,
fundoscopic examination, and fundus photography. DR was graded by an expert
ophthalmologists according to the American Academy of Ophthalmology proposed
international scales for severity of clinical diabetic retinopathy.46 A questionnaire was
designed for collecting information regarding gender, age, age at diagnosis of diabetes,
and ocular history. For each patient, systolic and diastolic blood pressure, waist and
hip circumferences, body mass index (BMI), and hemoglobin A1C (HbA1C) levels
were determined. In addition, 100 non-diabetic controls were randomly selected from
the Taiwan Han Chinese Cell and Genome Bank.47 The criteria for selecting controls
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
HbA1C ranging between 3.4% and 6.0%, and (iii) a BMI <32 kg/m2. This study was
approved by CMUH institutional review boards, and informed consent was obtained
from all study participants.
Genotyping
For the genome-wide association study, genomic DNA was extracted from
peripheral blood mononuclear cells using a PUREGENE DNA isolation kit (Gentra
Systems, Minneapolis, MN). Whole genome genotyping using Illumina
HumanHap550-Duo BeadChips was performed by deCODE genetics, Inc., Reykjavík,
Iceland. Genotype calling was performed using the standard procedure implemented
in BeadStudio, with default parameters suggested by the platform manufacturer.
Quality control of the genotype data was performed by examining several summary
statistics. First, the ratio of loci with heterozygous calls on the X chromosome was
calculated to double check the subject gender. Total successful call rate and the minor
allele frequency of the cases were also calculated for each SNP. SNPs were excluded
if they showed one of the following: (i) no polymorphism; (ii) a total call rate of
<95%; (iii) a minor allele frequency of <5%, or a total call rate of <99%. Genotyping
validation was performed using the Sequenom iPLEX assay (SEQUENOM
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 Statistical analysis
T2D association analysis was carried out to compare allele frequency and
genotype distribution between T2D subjects with and without DR using six
single-point methods: genotype, allele, trend (Cochran-Armitage test), additive,
dominant, and recessive models for each SNP. The most significant test statistic
obtained from the six models was selected. First, we selected SNPs with p-values less
than 10–5 in any of the abovementioned models for subsequent analysis. We then used
a logistic regression model with stepwise selection to determine the significant SNPs.
SNPs with p-values less than = 10–7, a cut-off for the multiple comparison adjusted by Bonferroni correction, were considered to be significantly associated with the traits.
SNPs with p-values between 10–7 and 10–5 were considered to have suggestive
significant associations. Characteristics and clinical data of subjects with DR and
without DR were compared by Student’s t-test for continuous variables and chi-square
test for categorical variables. Odds ratios (ORs) and 95% confidence intervals (CIs)
were performed by logistic regression and were adjusted with diabetes duration and
HbA1C level. Linkage disequilibrium analysis (D’ and r2) between any two loci were
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 Results
We conducted a genome-wide association study to identify genetic variants for
DR in T2D Han Chinese residing in Taiwan. We genotyped 749 T2D patients (174
with DR and 575 without DR) and 100 non-diabetic controls using Illumina
Hap550duov3 chips. The demographic and clinical characteristics of the above groups
are summarized in Table 1. Subjects without DR were of a significantly younger age
at the time of study and diagnosis, and had a shorter disease duration, lower HbA1C
levels, and lower systolic blood pressure compared with the subjects with DR (Table
1).
DR-associated SNPs were selected from those showing –log10(p-value) 5under the most significant test statistic obtained from any of the six statistical models. We
then used a logistic regression model with stepwise selection to determine the
significant SNPs. As shown in Table 2, twelve SNPs representing seven regions of
five chromosomes were selected from the regression model. The strongest SNP
association with DR (rs17376456) occurred on chromosome 5 [–log10(p-value) =
14.52]. This SNP is located in an intergenic region and has tight linkage
disequilibrium with rs13163610 (D’ = 1, r2
= 0.98). The SNP rs2038823 associated
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
sulfate 6-O-sulfotransferase 3) gene. The remaining five regions, located on
chromosomes 1, 4, and 10, are also associated with DR. The rs2811893 and
rs12092121 SNPs were localized to chromosome 1 in an intronic region of the
MYSM1 (Myb-like, SWIRM and MPN domains 1) gene and are in complete linkage
disequilibrium with each other (D’ = 1, r2 = 1). The rs4470583 SNP is located in an
intergenic region on chromosome 4. In addition, six SNPs located on three regions of
chromosome 10 are associated with DR. The rs12219125 SNP in an intergenic region
is strongly associated with DR [–log10(p-value) = 8.03], and is in strong linkage
disequilibrium with rs1571942 (D’ = 0.99, r2
= 0.96) localized to the PLXDC2 (plexin
domain containing 2) gene. The rs4462262 SNP in an intergenic region is also
strongly associated with DR [–log10(p-value) = 7.04]. Three SNPs (rs4838605,
rs11101355, and rs11101357) are in an intronic region of ARHGAP22 (Rho
GTPase-activating protein 22). Results from a pairwise linkage disequilibrium
analysis revealed complete linkage disequilibrium between rs11101355 and
rs11101357 (D’ = 1, r2 = 1).
Furthermore, the etiology of DR can vary according to duration of diabetes and
the status of glycemic control. Table 3 (available at http://aaojournal.org) shows the
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
with and without DR after controlling for the diabetes duration and HbA1C. The
results show that 9 SNPs representing five regions of four chromosomes remained
significantly associated with DR in the dominant model after controlling for diabetes
duration and HbA1C in the multiple logistic regression analysis. The risk genotypes
were defined subjects with homozygous risk allele (higher allele frequency in subjects
with DR than in subjects without DR). The risk TT genotype of rs2811893 was
associated with a 1.50-fold increase in DR risk under the dominant model (OR = 1.50,
95% CI = 1.03–2.20). Since rs2811893 is in complete linkage disequilibrium with
rs12092121, the AA genotype of rs12092121 was associated with the same 1.50-fold
increase in DR risk. The AA genotypes of rs13163610 and rs17376456 were
associated with a 3.59-fold (95% CI = 1.36–9.47) and 3.63-fold (95% CI = 1.38–9.58)
increase, respectively, in DR risk in the multiple logistic regression model after
controlling for diabetes duration and HbA1C. The SNPs rs4838605, rs11101355, and
rs11101357 lie within the ARHGAP22 gene of chromosome 10 and were associated
with 1.58-, 1.65- and 1.65-fold increases in DR risk, respectively. The SNP rs1571942,
located in the PLXDC2 gene of chromosome 10 was associated with a 1.67-fold (95%
CI = 1.06–2.65) higher risk of developing of DR. The SNP rs12219125 was in strong
linkage disequilibrium with rs1571942 and was associated with a 1.62-fold increase in
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
disequilibrium with rs12219125, it remains unclear which gene it is located in. The
CC genotype of rs2038823 was associated with a 2.33-fold increase in DR risk (95%
CI = 1.13–4.77). After controlling for diabetes duration and HbA1C, there was no
significant relationship between DR and the SNPs rs4470583 and rs4462262 located
on chromosomes 4 and 10, respectively. In addition, we compared the frequency of
genotypes between T2D patients with/without DR and non-diabetic controls. As
shown in Table 3 (available at http://aaojournal.org), there were significant differences
between subjects with DR and non-diabetic controls in rs13163610 and rs17376456
on chromosome 5 as well as for rs2038823 in the HS6ST3 gene on chromosome 13.
There also were significant differences between subjects without DR and non-diabetic
controls with respect to three SNPs in the ARHGAP22 gene on chromosome 10. For
the remaining SNPs, we observed no statistically significant difference between the
patients with/without DR and the controls.
We subsequently stratified the T2D subjects with DR according to DR severity
scales; however, due to the relatively small sample size of the DR subjects after
stratification, we pooled all the NPDR subjects together. We analyzed the 10 SNPs
exhibiting a significant difference (including rs4838605 in the ARHGAP22 gene,
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
DR after controlling for HbA1C and diabetes duration. As shown in Table 4 (available
at http://aaojournal.org), the risk genotypes located in the MYSM1 gene on
chromosome 1 (rs2811893 and rs12092121) were associated with a 1.72-fold increase
in NPDR risk under the dominant model (OR = 1.72, 95% CI = 1.09–2.71) after
controlling for HbA1c and diabetes duration. The AA genotypes of rs13163610 and
rs17376456 located in an unknown gene on chromosome 5 were associated with a
3.48-fold (95% CI = 1.05–11.6) and 3.54-fold (95% CI = 1.07–11.7) increase,
respectively, in NPDR risk. The SNPs rs1571942 and rs12219125, which were in
strong linkage disequilibrium and located in the PLXDC2 gene and an unknown gene,
respectively, on chromosome 10, were associated with a 1.86-fold (95% CI =
1.09–3.18) higher risk of developing NPDR. Furthermore, the CC genotype of
rs2038823 was associated with a 3.15-fold increase in NPDR risk (95% CI =
1.18–8.39). The SNPs rs4838605, rs11101355, and rs11101357 lie within the
ARHGAP22 gene of chromosome 10 and were associated with 2.18-, 2.22-, and
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 Discussion
Here, we describe the results of a genome-wide association study designed to
identify loci associated with the risk of DR in subjects with T2D. Significant
associations were identified in regions of chromosomes 1, 5, 10, and 13 after
controlling for diabetes duration and HbA1C. The results implicate MYSM1, PLXDC2,
ARHGAP22, HS6ST3, and an unknown gene on chromosome 5q as being involved in
the pathogenesis of DR, and particularly, with the exception ARHGAP22, in NPDR,
although the biology underlying these associations remains to be elucidated. None of
these loci have previously been reported to be associated with genes implicated in the
development of DR.
The SNP rs2811893 associated with DR, particularly NPDR, is in complete
linkage disequilibrium with rs12092121 and is located in the MYSM1 gene of
chromosome 1p. The MYSM1 gene encodes a JAMM/MPN+ (Jab1/MPN domain
metalloenzyme) domain-containing zinc metalloprotease that acts a deubiquitinylating
enzyme specific for mono-ubiquitinylated histone H2A.49-51 MYSM1 acts by
coordinating histone acetylation and deubiquitination, and destabilizing the
association of linker histone H1 with nucleosomes.49 MYSM1 is located on
chromosome 1p32, and a previous genome-wide linkage study has suggested that the
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
hypodysplasia and associated urinary tract malformations.52 However, the role of
MYSM1 in DR pathogenesis awaits further characterization. In addition to MYSM1
located in chromosome 1p32, a previous study has also reported suggestive evidence
linking DR with a region of chromosome 1p36 in Pima Indians with T2D.13 This
suggests that the shorter arm of chromosome 1 may harbor a number of genes
conferring susceptibility to DR.
In this study, six of the DR-associated SNPs were located in three regions of
chromosome 10. On the basis of the haplotype analysis, we can divide these SNPs
(with the exception of rs4462262) into two major haplotype blocks. The larger of the
two blocks, which includes the SNPs rs4838605, rs11101355, and rs11101357, is
located in the ARHGAP22 gene of chromosome 10q. ARHGAP22 encodes a Rho
family GTPase protein, which is involved in the signal transduction pathway that
regulates endothelial cell capillary tube formation during angiogenesis.53 In addition,
a recent study has suggested that the expression levels of ARHGAP22 play an
important role in determining the mode of tumor cell movement.54 A further recent
study has also suggested that ARHGAP22 may be involved in a novel
insulin-regulated pathway.55 In the present study, ARHGAP22 was the only gene
found to be associated with PDR. Since ARHGAP22 is involved in endothelial cell
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
which is located on chromosome 10p, includes the SNPs rs1571942 and rs12219125.
SNP rs1571942 is located in the PLXDC2 gene and is in strong linkage disequilibrium
with SNP rs12219125, the gene location of which is unknown. In previous studies,
PLXDC2, also called TEM7R (tumor endothelial marker 7-related protein), was
found to be expressed at high levels not only in tumor endothelium but also in the
vessels of some normal tissues, and may thus play a role in tumor angiogenesis.56 A
recent study has also shown that PLXDC2 may be associated with fasting insulin
levels and insulin resistance.57 Furthermore, a recent genome-wide study has shown
that PLXDC2 may be associated with overgrowth in patients with Sotos
syndrome-like features.58 SNP rs4462262 was not significantly associated with DR
after controlling for diabetes duration and HbA1C.
rs2038823, the other SNP associated with DR, particularly NPDR, is located on
chromosome 13q in a gene that encodes HS6ST3. HS6ST3 is a heparan sulfate (HS)
biosynthetic enzyme that modifies HS to generate structures required for the
interactions between HS and a variety of proteins. These interactions are regulated by
a wide variety of biological activities and are implicated in proliferation and
differentiation, adhesion, migration, inflammation, blood coagulation, and other
diverse processes.59 The Steno hypothesis states that in diabetes mellitus, changes in
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
dysfunction and increase capillary permeability.60 Previous studies using different
animal models, namely, Drosophila and mice, have shown that HS6ST3 is involved in
both limb bud development and tracheal branching.61,62
Finally, an additional locus (rs13163610 and rs17376456) is strongly associated
with DR, even after controlling for diabetes duration and HbA1C. However, the locus
is located in an intergenic region on chromosome 5q, and therefore it is uncertain
whether this locus plays a causal role in DR or is merely in linkage disequilibrium
with other functional loci.
According to a recent report by Abhary et al., VEGF and associated genes
involved in the renin-angiotensin system, the polyol pathway, and NO pathways are
the most appropriate candidate genes for studying the pathogenesis of DR.45 Since
both ARHGAP22 and PLXDC2 are involved in endothelial cell
angiogenesis—similar to VEGF, a major mediator of vascular permeability and
angiogenesis—they may play a pivotal role in mediating the development and
progression of DR.29,30 In addition, it is worth noting that the unknown gene on
chromosome 5q had the strongest association with DR and accordingly warrants
further investigation in the future.
The major limitation of this study is the small sample size of the DR patients. In
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
83% for subjects with or without DR for the unknown gene on chromosome 5q,
PLXDC2, and ARHGAP22, respectively. MYSM1 and HS6ST3, nevertheless, lacked
adequate power for subjects with/without DR. However, we have compared the allele
frequencies of these significant SNPs in healthy controls among different ethnic
groups [Data from the SNP database (dbSNP BUILD 131;
http://www.ncbi.nlm.gov/SNP/)]. It is interesting to note that the allele frequencies of
three SNPs located in the ARHGAP22 gene are very different among the different
ethnic groups. Since these three SNPs were in strong linkage disequilibrium with each
other, we selected only the SNP rs11101357 for description. The A and G allele
frequencies are reported, respectively, to be 12.2% and 87.8% in Chinese populations,
4.7% and 95.3% in Japanese populations, 65.3% and 34.7% in European populations,
and 22.9% and 77.1% in sub-Saharan African populations. The allele frequencies
found in the Chinese populations were similar to those seen in our Taiwanese
population (A allele, 13.5%; G allele, 86.5%) in the present study. The allele
frequency is very different among Africans and the Japanese, and particularly in
Europeans. Therefore, future studies with a larger number of subjects or subjects from
different ethnic backgrounds will be necessary in order to determine whether these
findings can be replicated. As yet, we have been unable to find any evidence to link a
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
DR. In order to determine whether subjects with the risk genotypes of SNPs are more
susceptible to developing DR, it will be necessary to undertake a long-term follow-up
of T2D subjects without DR who carry these risk genotypes and to evaluate the
effects of these genes on the development of DR.
In conclusion, the findings of this study contribute to our understanding of the
genetic susceptibility of DR in type 2 diabetes. The novel DR risk loci occur in
ARHGAP22 and PLXDC2 genes that are implicated in endothelial cell angiogenesis
and increased capillary permeability. Although these findings require further study for
confirmation, including a different cohort or larger sample size, it is interesting to
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 References
1. Prokopenko I, McCarthy MI, Lindgren CM. Type 2 diabetes: new genes, new
understanding. Trends Genet 2008;24:613-21.
2. He Z, King GL. Microvascular complications of diabetes. Endocrinol Metab
Clin North Am 2004;33:215-38.
3. Caldwell RB, Bartoli M, Behzadian MA, et al. Vascular endothelial growth
factor and diabetic retinopathy: pathophysiological mechanisms and treatment
perspectives. Diabetes Metab Res Rev 2003;19:442-55.
4. Moss SE, Klein R, Klein BE. The 14-year incidence of visual loss in a diabetic
population. Ophthalmology 1998;105:998-1003.
5. Taylor HR, Keeffe JE. World blindness: a 21st century perspective. Br J
Ophthalmol 2001;85:261-6.
6. Brownlee M. Biochemistry and molecular cell biology of diabetic complications.
Nature 2001;414:813-20.
7. Jerneld B, Algvere P. Relationship of duration and onset of diabetes to
prevalence of diabetic retinopathy. Am J Ophthalmol 1986;102:431-7.
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
macrovascular and microvascular complications of type 2 diabetes (UKPDS 35):
prospective observational study. BMJ 2000;321:405-12.
9. Hallman DM, Huber JC, Jr., Gonzalez VH, et al. Familial aggregation of
severity of diabetic retinopathy in Mexican Americans from Starr County, Texas.
Diabetes Care 2005;28:1163-8.
10. Rema M, Saravanan G, Deepa R, Mohan V. Familial clustering of diabetic
retinopathy in South Indian Type 2 diabetic patients. Diabet Med 2002;19:910-6.
11. Hanson RL, Ehm MG, Pettitt DJ, et al. An autosomal genomic scan for loci
linked to type II diabetes mellitus and body-mass index in Pima Indians. Am J
Hum Genet 1998;63:1130-8.
12. Imperatore G, Hanson RL, Pettitt DJ, et al. Sib-pair linkage analysis for
susceptibility genes for microvascular complications among Pima Indians with
type 2 diabetes. Pima Diabetes Genes Group. Diabetes 1998;47:821-30.
13. Looker HC, Nelson RG, Chew E, et al. Genome-wide linkage analyses to
identify Loci for diabetic retinopathy. Diabetes 2007;56:1160-6.
14. Hallman DM, Boerwinkle E, Gonzalez VH, et al. A genome-wide linkage scan
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
diabetes from Starr County, Texas. Diabetes 2007;56:1167-73.
15. dos Santos KG, Canani LH, Gross JL, et al. The -106CC genotype of the aldose
reductase gene is associated with an increased risk of proliferative diabetic
retinopathy in Caucasian-Brazilians with type 2 diabetes. Mol Genet Metab
2006;88:280-4.
16. Santos KG, Tschiedel B, Schneider J, et al. Diabetic retinopathy in
Euro-Brazilian type 2 diabetic patients: relationship with polymorphisms in the
aldose reductase, the plasminogen activator inhibitor-1 and the
methylenetetrahydrofolate reductase genes. Diabetes Res Clin Pract
2003;61:133-6.
17. Wang Y, Ng MC, Lee SC, et al. Phenotypic heterogeneity and associations of
two aldose reductase gene polymorphisms with nephropathy and retinopathy in
type 2 diabetes. Diabetes Care 2003;26:2410-5.
18. Fujisawa T, Ikegami H, Kawaguchi Y, et al. Length rather than a specific allele
of dinucleotide repeat in the 5' upstream region of the aldose reductase gene is
associated with diabetic retinopathy. Diabet Med 1999;16:1044-7.
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
gene with retinopathy but not with nephropathy or neuropathy in Japanese
patients with Type 2 diabetes mellitus. Diabet Med 1999;16:744-8.
20. Ko BC, Lam KS, Wat NM, Chung SS. An (A-C)n dinucleotide repeat
polymorphic marker at the 5' end of the aldose reductase gene is associated with
early-onset diabetic retinopathy in NIDDM patients. Diabetes 1995;44:727-32.
21. Kador PF, Kinoshita JH. Role of aldose reductase in the development of
diabetes-associated complications. Am J Med 1985;79:8-12.
22. Abhary S, Burdon KP, Gupta A, et al. Common sequence variation in the
VEGFA gene predicts risk of diabetic retinopathy. Invest Ophthalmol Vis Sci
2009;50:5552-8.
23. Kangas-Kontio T, Vavuli S, Kakko SJ, et al. Polymorphism of the manganese
superoxide dismutase gene but not of vascular endothelial growth factor gene is
a risk factor for diabetic retinopathy. Br J Ophthalmol 2009;93:1401-6.
24. Churchill AJ, Carter JG, Ramsden C, et al. VEGF polymorphisms are associated
with severity of diabetic retinopathy. Invest Ophthalmol Vis Sci 2008;49:3611-6.
25. Uthra S, Raman R, Mukesh BN, et al. Association of VEGF gene
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 Genet 2008;29:11-5.
26. Suganthalakshmi B, Anand R, Kim R, et al. Association of VEGF and eNOS
gene polymorphisms in type 2 diabetic retinopathy. Mol Vis
2006;12:336-41.:336-41.
27. Awata T, Kurihara S, Takata N, et al. Functional VEGF C-634G polymorphism
is associated with development of diabetic macular edema and correlated with
macular retinal thickness in type 2 diabetes. Biochem Biophys Res Commun
2005;333:679-85.
28. Ray D, Mishra M, Ralph S, et al. Association of the VEGF gene with
proliferative diabetic retinopathy but not proteinuria in diabetes. Diabetes
2004;53:861-4.
29. Awata T, Inoue K, Kurihara S, et al. A common polymorphism in the
5'-untranslated region of the VEGF gene is associated with diabetic retinopathy
in type 2 diabetes. Diabetes 2002;51:1635-9.
30. Aiello LP, Avery RL, Arrigg PG, et al. Vascular endothelial growth factor in
ocular fluid of patients with diabetic retinopathy and other retinal disorders. N
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
31. Prasad P, Tiwari AK, Kumar KM, et al. Association of TGFbeta1, TNFalpha,
CCR2 and CCR5 gene polymorphisms in type-2 diabetes and renal insufficiency
among Asian Indians. BMC Med Genet 2007;8:20.:20.
32. Buraczynska M, Baranowicz-Gaszczyk I, Borowicz E, Ksiazek A. TGF-beta1
and TSC-22 gene polymorphisms and susceptibility to microvascular
complications in type 2 diabetes. Nephron Physiol 2007;106:69-75.
33. Wong TY, Poon P, Chow KM, et al. Association of transforming growth
factor-beta (TGF-beta) T869C (Leu 10Pro) gene polymorphisms with type 2
diabetic nephropathy in Chinese. Kidney Int 2003;63:1831-5.
34. Beranek M, Kankova K, Benes P, et al. Polymorphism R25P in the gene
encoding transforming growth factor-beta (TGF-beta1) is a newly identified risk
factor for proliferative diabetic retinopathy. Am J Med Genet 2002;109:278-83.
35. Pascal MM, Forrester JV, Knott RM. Glucose-mediated regulation of
transforming growth factor-beta (TGF-beta) and TGF-beta receptors in human
retinal endothelial cells. Curr Eye Res 1999;19:162-70.
36. Awata T, Neda T, Iizuka H, et al. Endothelial nitric oxide synthase gene is
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
37. Ezzidi I, Mtiraoui N, Mohamed MB, et al. Endothelial nitric oxide synthase
Glu298Asp, 4b/a, and T-786C polymorphisms in type 2 diabetic retinopathy.
Clin Endocrinol (Oxf) 2008;68:542-6.
38. Chen Y, Huang H, Zhou J, et al. Polymorphism of the endothelial nitric oxide
synthase gene is associated with diabetic retinopathy in a cohort of West
Africans. Mol Vis 2007;13:2142-7.:2142-7.
39. Warpeha KM, Xu W, Liu L, et al. Genotyping and functional analysis of a
polymorphic (CCTTT)(n) repeat of NOS2A in diabetic retinopathy. FASEB J
1999;13:1825-32.
40. Tong Z, Yang Z, Patel S, et al. Promoter polymorphism of the erythropoietin
gene in severe diabetic eye and kidney complications. Proc Natl Acad Sci U S A
2008;105:6998-7003.
41. Petrovic MG, Hawlina M, Peterlin B, Petrovic D. BglII gene polymorphism of
the alpha2beta1 integrin gene is a risk factor for diabetic retinopathy in
Caucasians with type 2 diabetes. J Hum Genet 2003;48:457-60.
42. Matsubara Y, Murata M, Maruyama T, et al. Association between diabetic
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
43. Kamiuchi K, Hasegawa G, Obayashi H, et al. Intercellular adhesion molecule-1
(ICAM-1) polymorphism is associated with diabetic retinopathy in Type 2
diabetes mellitus. Diabet Med 2002;19:371-6.
44. Liu L, Yu Q, Wang H, et al. Association of intercellular adhesion molecule 1
polymorphisms with retinopathy in Chinese patients with Type 2 diabetes.
Diabet Med 2006;23:643-8.
45. Abhary S, Hewitt AW, Burdon KP, Craig JE. A systematic meta-analysis of
genetic association studies for diabetic retinopathy. Diabetes 2009;58:2137-47.
46. Wilkinson CP, Ferris FL, III, Klein RE, et al. Proposed international clinical
diabetic retinopathy and diabetic macular edema disease severity scales.
Ophthalmology 2003;110:1677-82.
47. Pan WH, Fann CS, Wu JY, et al. Han Chinese cell and genome bank in Taiwan:
purpose, design and ethical considerations. Hum Hered 2006;61:27-30.
48. Barrett JC, Fry B, Maller J, Daly MJ. Haploview: analysis and visualization of
LD and haplotype maps. Bioinformatics 2005;21:263-5.
49. Zhu P, Zhou W, Wang J, et al. A histone H2A deubiquitinase complex
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
regulation. Mol Cell 2007;27:609-21.
50. Cope GA, Suh GS, Aravind L, et al. Role of predicted metalloprotease motif of
Jab1/Csn5 in cleavage of Nedd8 from Cul1. Science 2002;298:608-11.
51. Verma R, Aravind L, Oania R, et al. Role of Rpn11 metalloprotease in
deubiquitination and degradation by the 26S proteasome. Science
2002;298:611-5.
52. Sanna-Cherchi S, Caridi G, Weng PL, et al. Localization of a gene for
nonsyndromic renal hypodysplasia to chromosome 1p32-33. Am J Hum Genet
2007;80:539-49.
53. Katoh M, Katoh M. Identification and characterization of ARHGAP24 and
ARHGAP25 genes in silico. Int J Mol Med 2004;14:333-8.
54. Sanz-Moreno V, Gadea G, Ahn J, et al. Rac activation and inactivation control
plasticity of tumor cell movement. Cell 2008;135:510-23.
55. Larance M, Rowland AF, Hoehn KL, et al. Global phosphoproteomics identifies
a major role for AKT and 14-3-3 in regulating EDC3. Mol Cell Proteomics
2010;9:682-94.
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
markers are conserved in mice and humans. Cancer Res 2001;61:6649-55.
57. Dupuis J, Langenberg C, Prokopenko I, et al. New genetic loci implicated in
fasting glucose homeostasis and their impact on type 2 diabetes risk. Nat Genet
2010;42:105-16.
58. Visser R, Gijsbers A, Ruivenkamp C, et al. Genome-wide SNP array analysis in
patients with features of sotos syndrome. Horm Res Paediatr 2010;73:265-74.
59. Habuchi H, Tanaka M, Habuchi O, et al. The occurrence of three isoforms of
heparan sulfate 6-O-sulfotransferase having different specificities for hexuronic
acid adjacent to the targeted N-sulfoglucosamine. J Biol Chem
2000;275:2859-68.
60. Deckert T, Feldt-Rasmussen B, Borch-Johnsen K, et al. Albuminuria reflects
widespread vascular damage. The Steno hypothesis. Diabetologia
1989;32:219-26.
61. Kamimura K, Fujise M, Villa F, et al. Drosophila heparan sulfate
6-O-sulfotransferase (dHS6ST) gene. Structure, expression, and function in the
formation of the tracheal system. J Biol Chem 2001;276:17014-21.
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
heparan sulfate O-sulfotransferases and regional differences in heparan sulfate
Five previously unknown susceptibility regions related to diabetic retinopathy in a genome-wide association study were identified, which possibly indicate novel pathways in the pathogenesis of this disease.
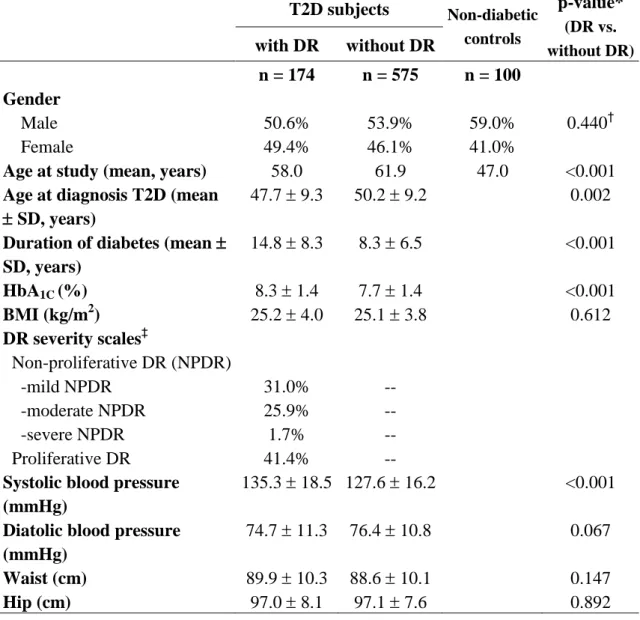
Table 1. Characteristics and clinical profiles of the study subjects
T2D, type 2 diabetes; DR, diabetic retinopathy; SD, standard deviation; HbA1C, hemoglobin A1C; NPDR, non-proliferative DR
*Student’s t-test; †Chi-square test; ‡According to the American Academy of Ophthalmology proposed international scales for severity of clinical diabetic retinopathy T2D subjects Non-diabetic controls p-value* (DR vs. without DR) with DR without DR n = 174 n = 575 n = 100 Gender Male 50.6% 53.9% 59.0% 0.440† Female 49.4% 46.1% 41.0%
Age at study (mean, years) 58.0 61.9 47.0 <0.001 Age at diagnosis T2D (mean
SD, years)
47.7 9.3 50.2 9.2 0.002 Duration of diabetes (mean
SD, years) 14.8 8.3 8.3 6.5 <0.001 HbA1C (%) 8.3 1.4 7.7 1.4 <0.001 BMI (kg/m2) 25.2 4.0 25.1 3.8 0.612 DR severity scales‡ Non-proliferative DR (NPDR) -mild NPDR -moderate NPDR -severe NPDR 31.0% 25.9% 1.7% -- -- -- Proliferative DR 41.4% --
Systolic blood pressure (mmHg)
135.3 18.5 127.6 16.2 <0.001 Diatolic blood pressure
(mmHg)
74.7 11.3 76.4 10.8 0.067 Waist (cm) 89.9 10.3 88.6 10.1 0.147
Hip (cm) 97.0 8.1 97.1 7.6 0.892
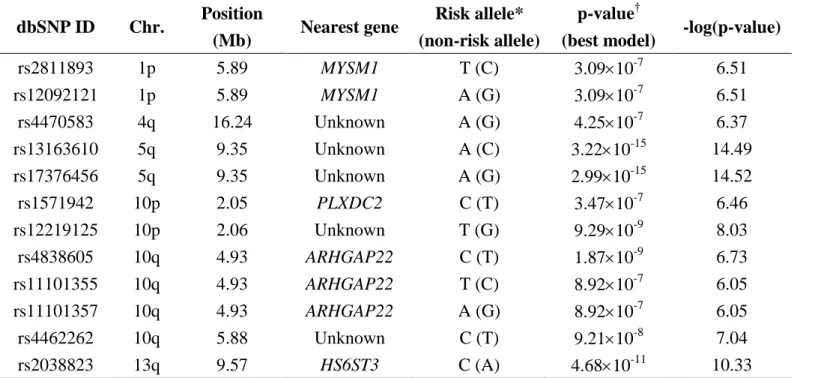
Table 2. Summary of the SNPs associated with diabetic retinopathy in type 2 diabetes
dbSNP ID, SNP database identification; Chr, chromosome; MYSM1, Myb-like, SWIRM and MPN domains 1; PLXDC2, plexin domain-containing 2; ARHGAP22, Rho GTPase-activating protein 22; HS6ST3, heparan sulfate 6-O-sulfotransferase 3
*Risk allele: the allele with higher frequency in subjects with diabetic retinopathy (DR) compared with subjects without DR
† P-value (best model): p-value of the most significant statistic obtained from six models: genotype, allele, trend, additive, dominant, and recessive mo
dbSNP ID Chr. Position
(Mb) Nearest gene
Risk allele* (non-risk allele)
p-value†
(best model) -log(p-value)
rs2811893 1p 5.89 MYSM1 T (C) 3.0910-7 6.51 rs12092121 1p 5.89 MYSM1 A (G) 3.0910-7 6.51 rs4470583 4q 16.24 Unknown A (G) 4.2510-7 6.37 rs13163610 5q 9.35 Unknown A (C) 3.2210-15 14.49 rs17376456 5q 9.35 Unknown A (G) 2.9910-15 14.52 rs1571942 10p 2.05 PLXDC2 C (T) 3.4710-7 6.46 rs12219125 10p 2.06 Unknown T (G) 9.2910-9 8.03 rs4838605 10q 4.93 ARHGAP22 C (T) 1.8710-9 6.73 rs11101355 10q 4.93 ARHGAP22 T (C) 8.9210-7 6.05 rs11101357 10q 4.93 ARHGAP22 A (G) 8.9210-7 6.05 rs4462262 10q 5.88 Unknown C (T) 9.2110-8 7.04 rs2038823 13q 9.57 HS6ST3 C (A) 4.6810-11 10.33 Table 2
Table 3. Genotypic distribution among type 2 diabetics with and without retinopathy and non-diabetic controls, and adjusted odds ratios of the diabetic retinopathy susceptibility SNPs in type 2 diabetic with and without retinopathy
Chr, chromosome; dbSNP ID, SNP database identification; T2D, type 2 diabetes; DR, diabetic retinopathy; aOR, adjusted odds ratio; CI, confidence interval
*Adjusted odds ratio after controlling diabetes duration and HbA1C; †
Significant differences between T2D subjects with DR and non-diabetic control; ‡Significant differences between T2D subjects without DR and
T2D subjects Chr. (nearest gene) dbSNP ID Risk allele Genotype (dominant model) with DR n (%) without DR n (%) Non-diabetic controls n (%) aOR (95% CI)* (DR vs. without DR) rs2811893 T CC+CT 88 (50.6) 334 (58.3) 61 (61.0) 1.00 (Ref) 1p (MYSM1) TT 86 (49.4) 239 (41.7) 39 (39.0) 1.50 (1.03-2.20) rs12092121 A GG+GA 88 (50.6) 334 (58.3) 61 (61.0) 1.00 (Ref) 1p (MYSM1) AA 86 (49.4) 239 (41.7) 39 (39.0) 1.50 (1.03-2.20) rs4470583 A GG 145 (83.3) 484 (84.2) 84 (84.0) 1.00 (Ref) 4q (Unknown) GA+AA 29 (16.7) 91 (15.8) 16 (16.0) 1.16 (0.70-1.92) rs13163610† A CC+CA 5 (2.9) 60 (10.5) 13 (13.0) 1.00 (Ref) 5q (Unknown) AA 169 (97.1) 514 (89.5) 87 (87.0) 3.59 (1.36-9.47) rs17376456† A GG+GA 5 (2.9) 62 (10.8) 13 (13.0) 1.00 (Ref) 5q (Unknown) AA 169 (97.1) 513 (89.2) 87 (87.0) 3.63 (1.38-9.58) rs1571942 C TT 128 (73.6) 481 (83.7) 82 (82.0) 1.00 (Ref) 10p (PLXDC2) CT +CC 46 (26.4) 94 (16.3) 18 (18.0) 1.67 (1.06-2.65) rs12219125 T GG 129 (74.1) 481 (83.7) 82 (82.0) 1.00 (Ref) 10p (Unknown) GT+TT 45 (25.9) 94 (16.3) 18 (18.0) 1.62 (1.02-2.58) rs4838605‡ C TT 130 (74.7) 479 (83.7) 73 (73.0) 1.00 (Ref) 10q (ARHGAP22) CT+CC 44 (25.3) 93 (16.3) 27 (27.0) 1.58 (1.00-2.52) rs11101355‡ T CC 128 (73.6) 480 (83.8) 74 (74.0) 1.00 (Ref) 10q (ARHGAP22) CT+TT 46 (26.4) 93 (16.2) 26 (26.0) 1.65 (1.05-2.60) rs11101357‡ A GG 128 (73.6) 480 (83.8) 74 (74.0) 1.00 (Ref) 10q (ARHGAP22) AG+AA 46 (26.4) 93 (16.2) 26 (26.0) 1.65 (1.05-2.60) rs4462262 C TT+TC 14 (8.0) 75 (13.0) 11 (11.0) 1.00 (Ref) 10q (Unknown) CC 160 (92.0) 500 (87.0) 89 (89.0) 1.54 (0.79-2.99) rs2038823† C AA+AC 13 (7.5) 76 (13.3) 17 (17.0) 1.00 (Ref) 13q (HS6ST3) CC 161 (92.5) 497 (86.7) 83 (83.0) 2.33 (1.13-4.77) Table 3
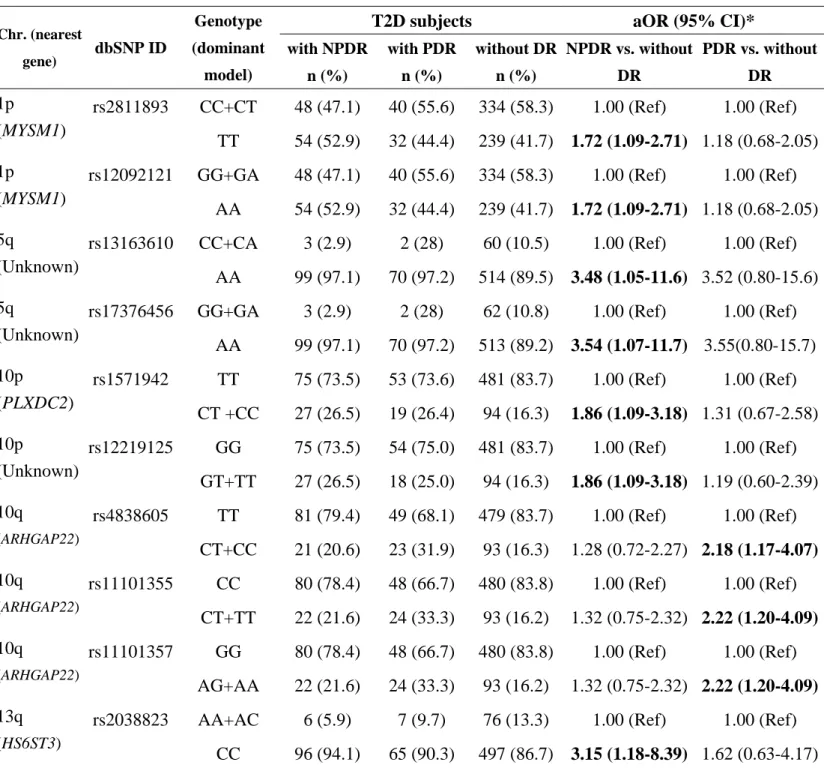
Table 4. Genotypic distribution and adjusted odds ratios of the diabetic retinopathy (DR) susceptibility SNPs in non-proliferative DR and proliferative DR
Chr, chromosome; dbSNP ID, SNP database identification; T2D, type 2 diabetes; DR, diabetic retinopathy; NPDR, non-proliferative DR; PDR, proliferative DR; aOR, adjusted odds ratio; CI, confidence interval; MYSM1, Myb-like, SWIRM and MPN domains 1; PLXDC2, plexin domain-containing 2; ARHGAP22, Rho GTPase-activating protein 22; HS6ST3, heparan sulfate 6-O-sulfotransferase 3
*Adjusted odds ratio after controlling diabetes duration and HbA1C
T2D subjects aOR (95% CI)*
Chr. (nearest gene) dbSNP ID Genotype (dominant model) with NPDR n (%) with PDR n (%) without DR n (%) NPDR vs. without DR PDR vs. without DR rs2811893 CC+CT 48 (47.1) 40 (55.6) 334 (58.3) 1.00 (Ref) 1.00 (Ref) 1p (MYSM1) TT 54 (52.9) 32 (44.4) 239 (41.7) 1.72 (1.09-2.71) 1.18 (0.68-2.05)
rs12092121 GG+GA 48 (47.1) 40 (55.6) 334 (58.3) 1.00 (Ref) 1.00 (Ref) 1p
(MYSM1)
AA 54 (52.9) 32 (44.4) 239 (41.7) 1.72 (1.09-2.71) 1.18 (0.68-2.05)
rs13163610 CC+CA 3 (2.9) 2 (28) 60 (10.5) 1.00 (Ref) 1.00 (Ref) 5q
(Unknown)
AA 99 (97.1) 70 (97.2) 514 (89.5) 3.48 (1.05-11.6) 3.52 (0.80-15.6)
rs17376456 GG+GA 3 (2.9) 2 (28) 62 (10.8) 1.00 (Ref) 1.00 (Ref) 5q (Unknown) AA 99 (97.1) 70 (97.2) 513 (89.2) 3.54 (1.07-11.7) 3.55(0.80-15.7) rs1571942 TT 75 (73.5) 53 (73.6) 481 (83.7) 1.00 (Ref) 1.00 (Ref) 10p (PLXDC2) CT +CC 27 (26.5) 19 (26.4) 94 (16.3) 1.86 (1.09-3.18) 1.31 (0.67-2.58) rs12219125 GG 75 (73.5) 54 (75.0) 481 (83.7) 1.00 (Ref) 1.00 (Ref) 10p (Unknown) GT+TT 27 (26.5) 18 (25.0) 94 (16.3) 1.86 (1.09-3.18) 1.19 (0.60-2.39) rs4838605 TT 81 (79.4) 49 (68.1) 479 (83.7) 1.00 (Ref) 1.00 (Ref) 10q (ARHGAP22) CT+CC 21 (20.6) 23 (31.9) 93 (16.3) 1.28 (0.72-2.27) 2.18 (1.17-4.07) rs11101355 CC 80 (78.4) 48 (66.7) 480 (83.8) 1.00 (Ref) 1.00 (Ref) 10q (ARHGAP22) CT+TT 22 (21.6) 24 (33.3) 93 (16.2) 1.32 (0.75-2.32) 2.22 (1.20-4.09) rs11101357 GG 80 (78.4) 48 (66.7) 480 (83.8) 1.00 (Ref) 1.00 (Ref) 10q (ARHGAP22) AG+AA 22 (21.6) 24 (33.3) 93 (16.2) 1.32 (0.75-2.32) 2.22 (1.20-4.09)
rs2038823 AA+AC 6 (5.9) 7 (9.7) 76 (13.3) 1.00 (Ref) 1.00 (Ref) 13q
(HS6ST3)
CC 96 (94.1) 65 (90.3) 497 (86.7) 3.15 (1.18-8.39) 1.62 (0.63-4.17) Table 4