行政院國家科學委員會專題研究計畫 成果報告
肺癌醣化酵素基因異常調控之研究(3/3)
計畫類別: 個別型計畫 計畫編號: NSC93-2314-B-002-012- 執行期間: 93 年 08 月 01 日至 94 年 07 月 31 日 執行單位: 國立臺灣大學醫學院內科 計畫主持人: 余忠仁 報告類型: 完整報告 處理方式: 本計畫可公開查詢中 華 民 國 94 年 12 月 5 日
行政院國家科學委員會補助專題研究計畫
肺癌醣化酵素基因異常調控之研究(3/3)
計畫類別:■ 個別型計畫 □ 整合型計畫
計畫編號:NSC
93-2314-B-002-012
執行期間:93 年 08 月 01 日至 94 年 07 月 31 日
計畫主持人:余忠仁
共同主持人:
計畫參與人員:
成果報告類型(依經費核定清單規定繳交):□精簡報告 ■完整報告
本成果報告包括以下應繳交之附件:
□赴國外出差或研習心得報告一份
□赴大陸地區出差或研習心得報告一份
□出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份
處理方式:除產學合作研究計畫、提升產業技術及人才培育研究計畫、
列管計畫及下列情形者外,得立即公開查詢
□涉及專利或其他智慧財產權,□一年□二年後可公開查詢
執行單位:國立台灣大學醫學院內科
中 華 民 國 九十四 年 十一 月 三十 日
附件一J 成 果 報 告
F期中進度報告
一、中文摘要 黏液素被認為與癌細胞之局部侵犯與遠隔轉移有關,由於癌細胞之黏液素基因表現及 其醣化過程異於正常細胞,有利其脫離局部病灶,並逃避宿主之免疫系統攻擊。在吾人先 前之研究中,已證實肺癌會發生黏液素基因之變異,而此變異與異常醣化過程(涎黏液素之 產生)均與肺癌病患術後之預後有關。同時並發現涎黏液素之發生與致癌基因 neu/ erbB-2 有明顯相關性。吾人先前利用 multiplex PCR 探討肺癌之醣化酵素基因表現(包括 sialyltransferase,STs;與 fucosyltransferase,FucTs),侵犯力高易轉移的細胞株(CL1-5)其 STs 與 FucTs 之表現量高於侵犯力弱的母細胞株(CL1-0、CL1-1)。臨床上,超過 50%肺癌
檢體有 STs 與 FucTs 表現量增加之現象,而表現高量 ST3GalIII 與數種 FucTs(FucTIII、IV、 VI、VII)之肺癌患者其臨床預後明顯較差,術後復發高與長期存活率低。本三年研究計劃 之目的為:(1)廣泛而完整的探討醣化酵素基因(core1-4GnTs、β-1-4GalT I~VI、GlcNAcT I-VI) 在肺癌之表現情形以及與肺癌預後及臨床表現之相關聯性;(2)對於由(1)所選出具臨床重要 性之醣化酵素基因,探討基因表現之調控機轉,本研究將由 ST3GalIII、FucTIII、IV、VI、 VII 著手。本研究建立以 FucTIV、VI 基因轉植的穩定細胞株,此等細胞株會表現出 sLex 醣抗原,與母株不同的腫瘤生長速度,並可能有較高的轉移能力。
關鍵詞:肺癌、醣化酵素基因 Abstract
Mucins and mucin associated antigens are believed to play an important role in both invasion and metastasis of cancer cells. The altered expression of mucin peptides in cancer cells, as well as glycosylation, facilitates tumor cell invasion into blood stream, attachment to endothelial cells and escape from host immuno-surveillance. Our previous studies had demonstrated the prognostic implication of mucin expression for lung cancer. Lung cancers overexpressing sialomucins tend to have higher chance of recurrence and metastasis. The expression of sialomucin is correlated with the overexpression of an oncoprotein, erbB-2/neu, and at least one mucin gene (MUC5AC apomucin). We are than interested about the altered expression of glycosyltransferase genes in lung cancer. Preliminarily, using multiplex PCR to evaluate the expression of sialyltransferase(ST) and fucosyltransferase (FucT) gene families in lung cancer (in >50% of patients), we have demonstrated that overexpression of STs and FucTs are common in lung cancer tissues and cell lines. Daughter cell line (CL1-5) with high
invasiveness and metastasis ability expressed more STs and FucTs than parent cell line (CL1-0
and CL1-1). Clinically, patients bearing tumors with overexpression of ST3GalIII and FucTIII、
IV、VI、VII are liable to cancer recurrence and death. We thus propose a three-year study to comprehensively study the role of glycosyltransferase genes expression in lung cancer. Is up-regulation or down-regulation of certain glycosyl -transferase which leads to expression of cancer-associated sialylated antigens occurs preferentially in lung cancer? Can the altered expression of glycosyltransferase genes serve as a prognostic marker? Specific glycosyltransferase genes considered to be important in lung cancers will be cloned and subjected to further study.
Keywords: Lung cancer, glycosyltransferase 二、緣由與目的
Mucin glycoprotein consists of a protein backbone with many carbohydrate side chains of varying lengths, sequences, compositions and anomeric linkages. They have a very large molecular weight (400 to > 1000 kDa), many O-glycosidically linked carbohydrate side chains which may constitite 50-85% of the total molecular weight, a high content of serine, threonine and proline in the protein backbone structure. The carbohydrate moieties of mucin glycoprotein may provide important biological functions of cells. These include receptor function for growth factors, hormones, toxins, bacteria, and virus lectins, growth regulation, cellular differentiation, homotypic and heterotypic, cell-cell interaction, cell-substratum or cell-basement membrane interactions and various immunological functions.[1] Experimentally, mucins had been demonstrated to promote tumor cell invasion, metastasis and modulate the immune recognition phenomenon of cancer cells.[2] The aberrant expression of mucins and mucin related antigens are poor survival factors in carcinomas arising from various organs, such as colon and breast cancer. Our previous studies had demonstrated the prognostic implication of mucins for lung cancer. [3-5] Lung cancers overexpressing sialomucins (highly sialylated mucins not so heavily expressed in normal airway) tend to have higher chance of recurrence and metastasis. The overexpression of sialomucin correlated with erbB-2/neu oncoprotein overexpression and at least one mucin core peptide (MUC5AC apomucin).
Aberrant glycosylation of mucins is common in cancer cells. Among cancer-associated antigens, sialylated Lewis antigens such as sialyl Lewis x (sLex) and sialyl Lewis a (sLea) have been well characterized. It is well known that the sLex and sLea epitopes produced in cancer cells are mainly carried on mucin O-glycans. The augmented expression of sLex and sLea antigens is frequently observed in some cancerous tissues, including lung cancer. It has been reported that sialyl Lex and sialyl Lea are involved in the process of metastasis, because these compounds serve as ligands for P- and L-selectin expressed on the surface of vascular endothelial cells, and mediate the adhesion of malignant cells to the vascular endothelium[6] Clinical reports also recognized these compounds to be poor prognostic factors for lung cancer and colon cancer.[7,8]
Glycosyltransferase and mucin glycosylation
Glycosyltransferases are enzymes arrayed in Golgi apparatus.. These enzymes transfer glycosyl residues from nucleotide-activated sugar molecules to other carbohydrates or aglycans (peptides or lipids) in a highly efficient and specific way. They work like sequential part of an assembly line with “cooperative sequential specificity”, that the product of one
glycosyltransferase becomes the acceptor substrate for the next glycosyltransferase. By
estimation, there are more than 250 glycosyltransferases, and the assembly of these enzymes on Golgi may be tissue specific, i.e. different tissue may have different glycosylation process on same peptide backbone.[9,10]
Mucin-type glycoproteins are unique in having clusters of large numbers of O-glycans. These O-glycans contain N-acetylgalactosamine residues at reducing ends, which are linked to serine or threonine in a polypeptide. The initiation of O-linked glycosylation occurs through the action of UDP-Gal-Nac: polypeptide N-acetylgalactosaminyltransferase (ppGaN-Tase), which catalyzes the transfer of GalNAc from the nucleotide sugar UDP-GalNAc to the hydroxyl group
of either serine or threonine. [11]
These attached O-glycans can be classified into several different groups according to the core structures subsequently added on GalNAc [12]. In many cells, core 1, Galβ1→3GalNAc, is the major constitute of O-glycans, core 1 oligosaccharides are converted to core 2
oligosaccharides, Galβ1→3 (GalNAcβ1 →6) GalNAc when core 2
β1,6-N-acetylglucosaminyltransferase (C2GnT) is present. [13] In the gastrointestinal tract and breast tissue, oligosaccharides with core 3, GlcNAcβ1→3 GalNAc, can be frequently found. These tissues also contain core 4, Galβ1→4Glcβ1→6(GlcNAcβ1 →3)GalNAc. Core 4 is formed from core 3 by core 4 β1,6-N-acetylglucosaminyltransferase (C4GnT). It has been reported that the amount of core 4 oligosaccharides is reduced in cancer cells, while the amount of core 2 oligosaccharides is maintained or increased.[14] More recently, the increase in the transcript of C2GnT was found to be associated with the progression of colonic carcinoma.[15]
A set of glycosyltransferases are required for the synthesis of cancer-associated sialylated antigens, they are: β1,3 N-acetylglucosaminyltransferase (β1,3GnT), β1,4-galactosyltransferase (β1,4GalT), α2,3-sialyltransferase (ST3Gal), and α1,3-fucosyltransferase (α1,3Fuc-T) for sLex
synthesis, and β1,3GnT, β1,3 GalT, ST3Gal, and α1,4Fuc-T for sLea
synthesis. The genes encoding β1,3GalT and β1,3GnT, which are required for the synthesis of type-1 (Lea
and Leb) and type-2 chains (Lex and Ley), have recently been cloned. The genes encoding β1,4
GalTs(I-VI), ST3GalTs (I-IV), α1,3Fuc-T (III-VI), α1,4FucT (FucTIII) and Core 1-4GnTs had been cloned.
Glycotransferase gene expression and cancer
Detailed mechanisms involved in regulation of glycosyltransferase genes are largely unclear. Most of the glcosyltransferase genes ("glyco-genes") are expressed in an organ- and
tissue-specific manner [16]. For example, the 5' untranslated region of GlcNAc-TV mRNA from various cells showed multiple sequences depending on the cell types [17]. Analysis of the 5' untranslated region revealed the presence of consensus motifs for transcription factors such as TATA box, AP-1/Ets, AP-2, HNF-1, and HP-1. The gene employs a multiple promoter system for transcription, and gene expression may then be regulated in a tissue-specific fashion. More over, multiple GlcNAc-TV transcripts were reported in various cell lines and tissues [18]. Disruption of the GlcNAc-TV gene in mice results in the loss of any detectable GlcNAc-TV activity, as well as β1-6 GlcNAc branching glycans in all tissue examined [19]. GlcNAc-TV homozygous null mice are viable and fertile, but showed enhanced rates of T cell aggregation and cell proliferation. Enhanced adhesion of cells (leukocytes, epithelial cells) to substratum, but reduced cell motility was also noted [20]. GlcNAc-TV catalyzes the attachment of a
β1-6GlcNAc residue, producing branched N-glycans. The high degree of branching of N-glycans appears to be related to the malignant potential of tumor cells. In particular, β1-6 branching of N-glycans is directly linked to tumor metastasis. Transfection of mammary carcinoma cell lines with a GlcNAc-TV expression vector increased metastasis by 4-40 folds [21]. The expression of GlcNAc-TV gene could also be induced by oncogene (e.g., ras-signaling pathway) or viral (e.g., v-src) transfections during hepato-carcinogenesis. In addition, the
GlcNAc-TV activity is also augmented by phorbol esters and TGF-β1. Recently, erbB-2/neu has been shown to be able to stimulate the transcription of GlcNAc-TV through Ras-Raf-Ets signal
transduction pathway [22].
Unlike GlcNAc-TV, the molecular basis of most human glycogenes remains elusive. As the genomic regions containing the promoter were isolated in only few genes, and complete
nucleotide sequence of many genes had only recently become available.
Rationale of study design
Expression of the above glyco-genes has also been studied in many other types of cancer, and also demonstrated tissue-specificity. For example, downregulation of FucT VII and
ST3GalIII was noted in colon cancer and cervical cancer [23,24], while the opposite was shown in gastric cancer [25] and lung cancer (as in our preliminary data). Based on the PCR analyzing the CL cell lines, we noticed a successive increased expression of FucTIII, FucTVI and ST3Gal III from cell line with low invasiveness(CL1-0) to high invasiveness(CL1-5). Since the expression
of glycosyltransferases is mainly regulated at the level of transcription, enzyme activity of glycosyltransferases is strongly correlated with mRNA expression, measured by either Northern blot or in situ hybridization [26]. We hypothesize that increase expression (activity) of FucT III, FucT VI and ST3Gal III play important role in lung carcinogenesis.
Both STs (ST3s and ST6s) and FucTs (FucT-III-VII) are key enzymes participating in synthesizing Lewis and sialyl-Lewis antigens (Lea, Leb, Lex, Ley, sLex, sLea). Lewis and
sialyl-Lewis antigens are involved in the process of metastasis, these sialylated and fucosylated compounds serve as the ligands of E-selectin or P-selectin expressed on the surface of vascular endothelial cells. ST3GalIII preferentially synthesize sLea, FucTIII-VII are enzymes catalyzing the last step of synthesis of Lewis and sialyl-Lewis antigens. FucT III is the Lewis enzyme and ubiquitously expressed in the epithelial cells of aerodigestive tracts, and is the only FucT with both α1,3 and α1,4 fucosylation activity to synthesize both α1,3-fucosyl-containing Lex
and sLex and α1,4-fucosyl-containing sLea. FucT VI and FucT VII preferentially synthesize Lex, Ley, and sLex. FucT VI possesses the strongest activity in synthesizing sLex antigen.
ST3GalIII and FucT III are identified as genes of interest. Therefore, we are interested in studying whether the abnormal activation of ST3GalIII and FucT III may contribute to the metastatic phenotype during the progression of cancers. We will clone ST3GalIII and FucT III genes and establish inducible expression system of ST3GalIII and FucT III in transfected cell lines. We propose to transfect ST3GalIII and FucT III sense cDNA into CL1-0 and anti-sense
cDNA to CL1-5. Biochemical or functional change of cells with overexpressed or
down-translated enzymes will be evaluated by flow cytometry (formation of specific glycans, i.e. Lex ,sLex and sLea ), migration/ invasion/metastasis assay, anchorage-independent growth assay,
in vivo tumorigenicity and metastasis. Catalytic activity of glycosyltansferases in tranfected cell
lines will also be measured.
Except few enzymes, the regulatory mechanism of the expression of most
glycosyltransferase are majorly unknown. Limited reports had demonstrated that the expression of certain glycosyltransferase genes, e.g. GlcNAcT V, ST8, FucT VII can be changed by growth factor, phorbolester, or retinoic acid.[27,28] We, therefore, will investigate the effect of several factors (proliferation factors: EGF, phorbol-12-myristate-13-acetate (PMA), differentiation factors: retinoic acid, 8-Br-cAMP, cytokines: TNF, monokines) on the expression of FucT III
and ST3GalIII in CL1-0 and CL1-5 cells, as well as non-malignant airway epithelial cell lines
(HBE and BAES-2B).
Besides ST3s and FucTs, several classes of glycosyltansfease genes may have contribution to tumorigenesis. These genes include core1-4GnTs、β-1-4GalT I~VI、GlcNAcTs (I-VI, especially III and V) [16,29]. Unlike STs and FucTs which modify terminal glycosylation, coreGnTs、 β-1-4GalTs and GlcNAcTs synthesize the core glycan and elongate backbone glycans. Altered expression of these genes may affect cell surface receptors (e.g. cytokine and adhesion receptors) and influence cellular growth and proliferation. To identify candidate genes related to lung carcinogenesis, multiplex RTp-PCR of coreGnTs、β-1-4GalTs and GlcNAcTs will be performed in CL cell lines and in clinical lung cancer specimens.
三、材料與方法
Patients and tumor tissues
From 1992 to 1996, we have collected 309 pairs of lung tumor/nontumor lung tissues. Among them, 235 had the pathology of non-small cell lung cancer. Seventy tissue pairs from patients with detailed clinical data and follow-up history were chosen.
Surgical specimens of tumors and the adjacent uninvolved lung tissue will be obtained from patients with lung cancer at the time of resection. All patients should have non-small cell lung cancer confirmed by histological diagnosis. Preoperative staging work-up included chest radiograph, fiberoptic bronchoscopy with brushing/washing cytology and biopsy, sputum cytology, computed tomography of thorax and abdomen, and bone scanning. Computed tomography of the brain is not a routine staging procedure. All patients have been judged preoperatively to have resectable disease, and all undergo a complete resection of the tumor. The resection will be judged complete if all known tumorous tissue is completely removed, resection margins are microscopically free of the tumor, and the area or nodes proximal to the involved area or nodes is microscopically free of tumor. After excision, tumor samples and the uninvolved lung tissues were collected immediately, snap frozen in isopentane at –60°C placed in sterile jar and stored at –70°C until processed. Specimens used for formalin fixation and for OCT embedding are collected separately from specimens used for RNA isolation. The resected lung and lymph nodes were subjected to routine surgical pathological examination.
Representative sections required for staging and histologic classification were generously taken. Sections of 4µm thickness were routinely stained with hematoxylin-eosin. Histologic classification was based on World Health Organization criteria. The final staging of each patient was pathologic, according to the international staging system for lung tumors.
Laser capture microdissection and RNA extraction
For microdissection, a laser microscope system (PixCell II™ LCM System, Arcturus Engineering, Inc., Mountain View, CA) is used. Areas containing tumor cells or cells from normal lung tissues for control are identified on unmounted H&E-stained section and are visualized through video monitor. CapSureTM film carriers are positioned over the cells of interest and capture the cells after a short pulse of low power infrared laser. The CapSureTM film carriers, with the attached cells, are lifted and placed into Eppendorf tubes and subjected for RNA extraction.
Multiplex PCR for glyco-genes
The expression of selected glyco-genes or gene family in tissues will be evaluated using multiplex PCR [31]. CDNA is synthesized from the isolated total RNA by reverse transcription in 20 µl reactions containing 0.5µg of random primers, 200 U of superscript RTse, 2µg of total RNA, 4 µl of 5X RT buffer, 0.1mM each dNTP, 20 units Rnasin, and 6.5 µl DEPCed water. Each tube is incubated at room temperature for 10 min and then at 42°C for 45min, heated to 90°C for 10 mink, and then quick-chilled on ice. CDNA clones of each genes will be used as positive control. The PCR primers are designed based on the cDNA sequence of specific gene, and are selected with Primer 3 (a web-based primer designed, Whitehead Institute for
Biomedical Research). In designing the primers for multiplex PCR, the possibility of primer dimerization is minimized by cross-examination of the mismatches in the sequences of each possible paur of all the primers chosen. Primers with optimal annealing temperatures of 56-60°C are chosen to prevent mismatches, and primers with higher temperatures are chosen for shorter PCR products. Each PCR is performed in a 100-µl reaction mixtuer containing 10 µl RT reaction mixture, 1X PCR buffer, 0.1mM of each dNTP, 2.5 units of Taq polymerase, adjusted
concentration of each primers, and subjected to 28-30 cycles of PCR reaction. The PCR reaction products are then electrophoresed in a 3% agarose gel and stained with ethidium bromide, and the size of each cDNA product is determined by comparison to size marker (100bp DNA ladder).
Quantitaiton of multiplex RT-PCR products
After amplification, the PCR products were electrophoresed in 3% agarose gel, stained in 5 µg/ml ethidium bromide solution for 30 min, and then destained. The signal intensity of amplified native and mutated products was directly measured and digitized by IS-1000 digital imaging system (Alpha Innotech Incorp., San Leandro, CA). To evaluate the relative levels of expression of the target genes in multiplex RT-PCRs, the value of the internal standards (β2-microglobulin, GAPDH, or β-actin) in each test tube is used as the baseline gene expression of that sample, and the relative value is calculated for each of the target genes amplified in that reaction. These values are then used to compare expression across samples tested.
Cloning of cDNA of Fucosyltransferase IV and VI genes
Ten µg of total RNA from cell line or tissues with specific glycogene expression will be reverse transcribed to cDNA by 200 units Superscript RTase (BRL), 5 nmole dNTP (dATP, dTTP, dCTP, dGTP), 0.2 µmole dithiothreitol, 4 µl 5X reaction buffer, 20 units RNasin (Promega), and DEPC-treated distilled, deionized water in 20 µl reaction. One µg cDNA will be used in 1 PCR reaction. The PCR will be amplified by using specific primers for the gene and thermostable Puf DNA polymerase with proofreading activity. The PCR fragments generated will be modified by A-tailed with Taq DNA polymerase and ligated to pGEM-T Easy Vector (Promgea). The recombinant will be tranformed into E.coli JM 109 High efficiency Competnet Cells (Promega). The recombinant clones will be identified by blue/white selection. The plasmids from the selected clones will be isolated by Mini-M (Viogene) plasmid minipre procedure.
DNA sequencing
The construct will be isolated from plasmid clones, dideoxy sequencing will be performed on double-stranded DNA using an autosequencer (model ABI 377, Perkin Elmer).
Subcloning the glycogene from pGEM-T Easy Vector to pTRE2 Vector
The correct clone identifed by sequencing will be sublone to pTRE2 Vector for expression in cell line with low or no expression of target gene. pGEM-T Easy with insert of Slug cDNA and pTRE2 Vector will be digested by Not I. The cDNA and digested pTRE2 Vector will be ligated with T4 DNA ligase. The recombinant will be transformed into competent XL-1 Blue cell. Colonies will be selected for amplification and plasmids will be isolated. Then the plasmids will be HindIII and Not I, the sense direction of cDNA inserted in the pTRE2 vector will be identified.
Transient Transfection of Cell lines with glycogenes
In a six-well or 35 mm tissue culture plate, seed ~2x105 human bronchial epithelial (HBE) cells or NIH-3T3 cells per well in 2 ml of suitable mediums. Incubate the cells at 37°C in a CO2
incubator until the cells are 60-80% confluent. Prepare the following solutions: Solution A: For each transfection, dilute 2 µg of DNA into 100 µl serum-free medium. Solution B: For each transfection, dilute 4-16 µl of LIPOFECTlN™ Reagent (GIBCO BRL) into100 µl serum-free medium. Combine the two solutions, mix gently, and incubate at room temperature for15-45 min. Wash the cells once with 2 ml of serum-free medium. Add 0.8 ml serum-free medium to each tube containing the lipid-DNA complexes. Mix gently, overlay diluted complex solution onto washed cells. Incubate the cells for 5 h at 37°C in a CO2 incubator, later change medium to
serum-containing (3T3) or growth factors (HBE) containing. Replace medium at 18-24 h following start of transfection. After a 48 h transfection, cells are lysed and assayed for luciferase and β-galactosidase activities. Luciferase activity is then normalized with β-galactosidase activity. All transfection will be done in triplicate and the results are then averaged.
Development of stable Tet-Off cell lines
Cells will be transfected pTet-Off vector (Clontech) with LipofectAMINE reagent (Gobco-BRL). Host cells 5x105 will be seeded on 60 mm dishes and incubated overnight at 37℃ overnight. Before the addition of DNA-liposome complex, the cells should reach a condluency of 50-60%. The DNA-liposome complexes will be prepared as followed. Five µg of plasmid DNA will be diluted to 300 µl in RPMI-1640 serum free medium. Twenty µg of cationic liposomes will be diluted to 300 µl in RPMI-1640 serum free medium. The DNA and liposome solution will be mixed and stand at room temperature for 30 min. Then add 2400 µl serum free medium to DNA-liposome complex. After addition of DNA-liposome complex, the cells will be incubated for 16 hours at 37°C, at 5 % CO2. At the end of incubation, the medium will be replaced with
RPMI-1640 medium supplemented with 10% fetal bovine serum (FBS) and 2mM L-glutamine. After the growth in nonselective medium for 24-48 hours, G418 selection will be imposed (500 µg/ml; Geneticin; GiboBRL). After 3 weeks, G418-resistant cells will be isolated using cloning cylinders and maintained in the medium containing G418. A number of single-cell clones will be expanded and cultured. The picked clones will be screening by transient transfection with pTRE2-Luc for clones with low background and high induction of luciferase in response to Doxycycline (Sigma) 1 µg/ml.
First year
The expression of N-acetylglucosamin- ylransferase (MGAT, or GnT) I~V was evaluated in 49 paired tissues of lung cancer patients. The enzymes have broad acceptor activity for both N- and “mucin-type” O-glycan branching activity. There were 30 adenocarcinoma, and 19 squamous cell carcinoma. Stage distribution was: 20 stage I, 10 stage II, 17 stage III; 9 T1, 36 T2, 2 T3, 2 T4; 24 N0, 10N1,14 N2, 1 N3.
Overexpression of MGAT(GnT)s in lung cancer is not rare: MGAT I (57.1%), MGAT II (69.4%), MGATIV (40.8%), MGAT V (36.7%).
The correlation between MGATs expression and the clinical features or outcome of lung cancer patients reveals that only the expression MGAT1 has correlation with histology (78.9% of squamous cell carcinoma overexpressed MGAT1, vs 43.3% of adenocarcinoma, P=0.014). Patients bearing tumors with MGAT1 overexpression tended to have earlier disease relapse and cancer death..
Second year
A549 and CL1-0 cell lines were chosen as they expressed less FucT IV and VI in all the cell lines. (Fig.1). pTet-off control vector were transfected into A549 and CL1-0 cell lines, screened with G418. Transient transfected pTRE2-Luc and pRL-Tkwere tested for efficiency of transient trnsfection. The PCR fragments of full coding regions of FUT4 and FUT6 genes (Fig. 2) were inserted into pTRE2 -HA –hyg to form pTRE2 –HA-Hyg-FUT4 and pTRE2 –HA-Hyg-FUT6. Both plasmids were transfected into to A549-tetoff and CL1-0-tetoff cell lines. Stable cell lines were selected by hygromycin. Expression of RNA and protein of fucosyltransferases were screened by RT-PCR (Fig. 3) and anti-HA antibody.
DMSO (-) 5% MgCl2(mM) 1 2 4 1 2 4 T306 RT N257 DNA MgCl2(mM) 1 2 4 1 2 4 1593bp 1080bp FUT4 FUT6 DMSO (-) 5% MgCl2(mM) 11 22 44 11 22 44 T306 RT N257 DNA MgCl2(mM) 11 22 44 11 22 44 1593bp 1080bp FUT4 FUT6 NC I-H 35 8 CL 1-0 CL 1-1 CL 1-5 F4 NC I-H 15 7 NC I-H 52 0 NC I-H 29 2 A5 49 po siti ve NC I-H 32 2 Squamous Adenocarcinoma Mar ker FUT4 NC I-H 35 8 CL 1-0 CL 1-1 CL 1-5 F4 NC I-H 15 7 NC I-H 52 0 NC I-H 29 2 A5 49 po siti ve NC I-H 32 2 Squamous Adenocarcinoma Mar ker NC I-H 35 8 CL 1-0 CL 1-1 CL 1-5 F4 NC I-H 15 7 NC I-H 52 0 NC I-H 29 2 A5 49 po siti ve NC I-H 32 2 Squamous Adenocarcinoma Mar ker FUT4 NC I-H 358 CL 1-0 CL 1-1 CL 1-5 F4 NC I-H 15 7 NC I-H 52 0 NC I-H 29 2 A5 49 po siti ve NC I-H 32 2 Squamous Adenocarcinoma Mar ker FUT6 NC I-H 358 CL 1-0 CL 1-1 CL 1-5 F4 NC I-H 15 7 NC I-H 52 0 NC I-H 29 2 A5 49 po siti ve NC I-H 32 2 Squamous Adenocarcinoma Mar ker NC I-H 358 CL 1-0 CL 1-1 CL 1-5 F4 NC I-H 15 7 NC I-H 52 0 NC I-H 29 2 A5 49 po siti ve NC I-H 32 2 Squamous Adenocarcinoma Mar ker FUT6 con trol 1 2227 cont 1 2 4 5 ro l 4 14 CL1-0 Tetoff A12 con trol 1 2227 cont 1 2 4 5 ro l 4 14 CL1-0 Tetoff A12 Fig. 1 Fig. 2 Fig. 3
Third year
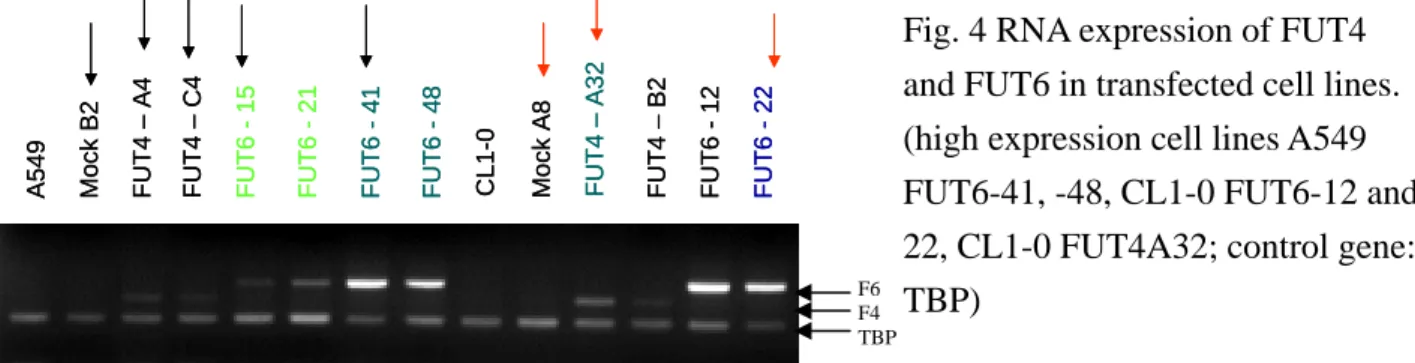
A549 and CL1-0 cell lines were transfected with FUCT4 and FUCT6 genes. Overexpression of FUT4 or FUT6 mRNA was confirmed in these cell lines by multiplex RT-PCR. (Fig.4) Cell lines with low or high expression of FUT4 or FUT6 mRNAs were identified.
A549 Mo ck B 2 FUT6 -1 5 FUT6 -2 1 FUT6 -4 1 FUT6 -4 8 FUT4 – A 4 FUT4 – C 4 CL 1-0 Mock A 8 FUT4 – A 32 FUT4 – B 2 FUT6 -1 2 FUT6 -2 2 A549 Mo ck B 2 FUT6 -1 5 FUT6 -2 1 FUT6 -4 1 FUT6 -4 8 FUT4 – A 4 FUT4 – C 4 CL 1-0 Mock A 8 FUT4 – A 32 FUT4 – B 2 FUT6 -1 2 FUT6 -2 2
The transfected cell lines expressed sLex antigen and ligands of E-selectin as detected by flow cytometry using E-selectin human IgG γ chain and anti-sLex antibody. (Fig 5 A & B).
Fig. 4 RNA expression of FUT4 and FUT6 in transfected cell lines. (high expression cell lines A549 FUT6-41, -48, CL1-0 FUT6-12 and 22, CL1-0 FUT4A32; control gene: TBP)
F6 F4 TBP
Fig. 5 (A) FACscan using E-selectin human IgG γ chain, shows that cell lines with FUT6 high expression expressed ligands recognized by E-selectin; (B) FACscan using E-selectin human IgG γ chain, shows that cell lines with FUT6 high expression expressed antigen recognized by antisLex antibody.
FUT6 clone dose
500 600 700 800 900 0 100 200 300 400 500 600 700 21 48 41 2(-∆∆Ct) sL e X G e oM ean A B
Fig.6. Correlation between sLex antigen expression and FUCT6 mRNA expression qutitated by real-time RTPCR
The growth rate of transfected cell lines are close to mock cell lines. The migration rate of each cell lines are also close to those of mock cell lines. However, transfected cell lines had lower capability in invasion to matrigel, with a decrease attachment ability to collagen IV matrix protein. CL1-0 FUT6 clone -20.0 0.0 20.0 40.0 60.0 80.0 100.0 120.0 0 20 40 60 hours re duc ti o n % CL1-0 m ock A8 FUT6 3 FUT6 5 FUT6 12 FUT6 22 ne ga tive control
invasion assay (matrigel)
-50.00 0.00 50.00 100.00 150.00 200.00 250.00
CL1-0 mock CL1-0 FUT6 -3 CL1-0 FUT6 -5 CL1-0 FUT6 -12 CL1-0 FUT6 -22
cel l n um be r m ig rat ed CL1-0 CollagenIV CL1 -0 CL1 -0 m ock CL1-0 FU T4 A32 CL1 -0 FUT 4 B2 CL1-0 FU T6 1 2 CL1-0 FU T6 22 0 1000 2000 BSA CollagenIV CL1-0 Vitronectin CL1 -0 CL1-0 moc k CL 1-0 FUT4 A32 CL 1-0 F UT4 B2 CL1 -0 FUT 6 1 2 CL1 -0 F UT 6 2 2 0 5000 10000 15000 BSA Vitronectin CL1-0 Fibronectin CL 1-0 CL 1-0 m ock CL 1-0 FU T4 A32 CL1-0 FUT4 B2 CL1 -0 FU T6 1 2 CL1 -0 FU T6 22 0 5000 10000 15000 BSA Fibronectin A B C D E
Fig 7 CL1-0 stable clone with FUT6 gene. (A) the growth rate is the same as parental line and mock, (B) Matrigel invasion assay. Transfection clone is less invasive to matrigel. (C-E) Matrix attachment assay with collagen IV, vitronectin and
fibronectin. Transfected cell lines showed various adhesive ability.
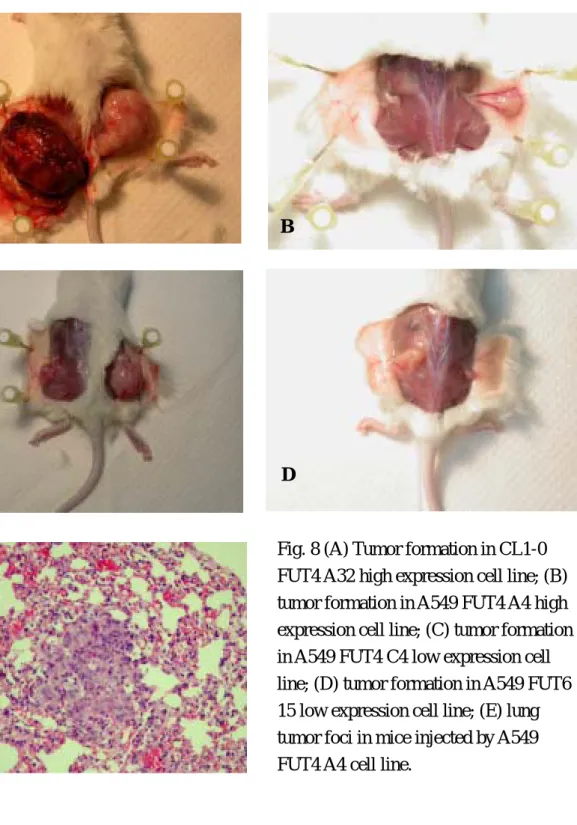
105 cells were injected subcutaneously into SCID mice, the formation of subcutaneous tumor was measured on a weekly base. The animals were sacrificed 8 weeks after the injection of tumor. Subcutaneous tumor, lung, and liver were sent for pathology. Transfected cell lines showed higher tumorigenicity than mock cell lines. One clone (FUT4) generates spontaneous lung metastasis. (Fig 8)
A B
C D
E
Fig. 8 (A) Tumor formation in CL1-0 FUT4 A32 high expression cell line; (B) tumor formation in A549 FUT4 A4 high expression cell line; (C) tumor formation in A549 FUT4 C4 low expression cell line; (D) tumor formation in A549 FUT6 15 low expression cell line; (E) lung tumor foci in mice injected by A549 FUT4 A4 cell line.
五、結論
We demonstrated that besides glycosyltransferases specific for terminal glycosyltaion, such as fucosyltransferases and sialyltransferases, enzymes specific for O-glycosylation branching may play certain role in lung caner carcinogenesis or tumor progression. Stable transfection of two fucosyltransferases (FUCT4 and FUCT6) into lung adenocarcinoma cell lines, A549 and CL1-0 will lead to expression of sLex antigen on cell surface and change the behavior of caner cells and potentially, lead to a highly metastatic clone.
六、參考文獻
1. Gendler SJ and Spicer AP. Epithelial mucin genes. Ann Rev Physiol. 1995;57:607-34. 2. Van Klinken BJW, Dekker J, Buller HA, and Enerhand AWC. Mucin gene structure and
expression: protection vs. adhesion. Am J Physiol. 1995;269:G613-27.
3. Yu CJ, Yang PC, Shew JY, Hong JM, Yang SC, Lee YC, Lee LN, Luh KT, Wu CA. Mucin mRNA expression in lung adenocarcinoma cell lines and tissues.Oncology 1996;53:118-26. 4. Yu CJ, Yang PC, Shun CT, Lee YC, Kuo SH, Luh KT. Overexpression of MUC5 genes is
associated with early postoperative metastasis in non-small cell lung cancer. Int J Cancer 1996; 69:457-65,
5. Yu CJ, Yang PC, Lee YC, Sew JY, Kuo SH, Luh KT. Sialomucin expression is associated with erbB-2 oncoprotein overexpression, early recurrence and cancer death in non-small cell lung cancer. Am J Respir Crit Care Med. 1997;155:1419-27.
6. Shimizu Y and Shaw S. Mucins in the mainstream. Nature 1993;366:630-1.
7. Ogawa Ji, Sano A, Koide S, and Shohtsu A. Relation between recurrence and expression of proliferating cell nuclear antigen, sialyl LewisX, and sialyl Lewis(a) in lung cancer. J Thorac
Cardiovas Surg 1994;108:329-36.
8. Ogawa Ji, Sano A, Inoue H, and Koide S. Expression of Lewis-related antigen and prognosis in stage I non-small cell lung cancer. Ann Thorac Surg 1995;59:412-5.
9. Van den Steen P, Rudd PM, Dwek RA, and Opdenakker G. Concepts and principles of O-linked glycosylation. Crit Rev Biochem Mol Biol 1998;33: 151-208.
10. Kleene R and Berger EG. The molecular and cell biology of glycosyltransferases. Biochem
Biophys Acta 1993;1154:283-325.
11. Ten Hagen KG, Hagen FK, Balys MM, Beres TM, Van Wuyckhuyse B, and Tabak LA. Cloning and expression of a novel, tissue specifically expressed member of the
UDP-GalNAc: polypeptide N-acetylgalactosaminyltransferase family. J Biol Chem 1998;273:27749-54.
12. Yeh JC, Ong E, and Fukuda M. Molecular cloning and expression of a novel beta-1, 6-N-acetylglucosaminyltransferase that forms core 2, core 4, and I branches. J Biol Chem 1999;274: 3215-21.
13. Fukuda M, Carlsson SR, Klock JC, and Dell A. Structures of O-linked oligosaccharides isolated from normal granulocytes, chronic myelogenous leukemia cells, and acute myelogenous leukemia cells. J Biol Chem 1986;261:12796-806.
14. Yang JM, Byrd JC, Siddik BB, Chung YS, Okuno M, Sowa M, Kim YS. Mtta KL, and Brockhausen I. Alterations of O-glycan biosynthesis in human colon cancer tissues.
15. Shimodaira K, Nakayama J, Nakayama N, Hasebe O, Katsuyama T, and Fukuda M.
Carcinoma -associated expression of core 2 beta-1,6-N-acetylglucosaminyltransferase gene in human colorectal cancer: role of O-glycans in tumor progression. Cancer Res
1997;5201-6.
16. Taniguchi N, Jain SK, Takahashi M, et al. Glycosyltransferases: cell surface remodeling and regulation of receptor tyrosine kinase-induced signaling. Pure Appl. Chem. 1999;71:719-28. 17. Saito H, Gu J, Nishikawa A, Ihara Y, Fujii J, Kohgo Y and Taniguchi N. Organization of the
human N-acetylglucosaminyltransferase V gene. Eur J Biochem 1995;233:18-26. 18. Perng GS, Shoreibah M, Margitich I, Pierce M and Fregien N. Expression of
N-acetyl-glucosaminyltransferase V mRNA in mammalian tissues and cell lines.
Glycobiology 1994;4: 867-71.
19. Granovsky M. Fode C. Warren CE. Campbell RM. Marth JD. Pierce M. Fregien N. Dennis JW. GlcNAc-transferase V and core 2 GlcNAc-transferase expression in the developing mouse embryo. Glycobiology. 5:797-806, 1995
20. Dennis JW. Granovsky M. Warren CE. Protein glycosylation in development and disease.
BioEssays. 1999;21:412-21.
21. Seberger PJ. Chaney WG. Control of metastasis by Asn-linked, beta1-6 branched oligosaccharides in mouse mammary cancer cells. Glycobiology 1999;9:235-41. 22. Chen L, Zhang W, Freigen N, and Pierce M. The her-2/neu oncogene stimulates the
transcription of N-acetylglucosaminyltransferase V and expression of its cell surface oligosaccharide products. Oncogene 1998; 17:2087-93.
23. Geβner P, Riedl S, Quentmaier A, Kemmner W. Enhanced activity of CMP-NeuAc:Galβ1-4 GlcNAc:α2,6-sialyltransferase in metastasizing human colorectal tumor tissue and serum of tumor patients. Cancer Lett 1993;75:143-9.
24. Wang PH, Li YF, Juang CM, et al. Altered mRNA expression of sialyltransferase in squamous cell carcinoma of the cervix. Gynecol Oncol 2001;83:121-7.
25. Petretti T, Schulz B, Schlag PM, Kemmner W. Altered mRNA expression of
glycosyltransferases in human gastric carcinomas. Biochem Biophys Acta 1999;1428:209-18. 26. Harduin-Lepers A, Krzewinski-Recchi MA, Hebbar M, et al. Sialyltransferases and breast
cancer. Recent Res Dev Cancer 2001;3:111-26.
27. Liu F, Qi HL, Chen HL. Regulation of differentiation-and proliferation-inducers of Lewis antigens, a-fucosyltransferase and metastatic potential in hepatocarcinoma cells. Br J Cancer 2001;84:1556-63.
28. Seidenfaden R, Hildebrandt H. Retinoic acid-induced changes in polysialyltransferase mRNA expression and NCAM polysialylation in human neuroblastoma cells. J Neurobiol 2001;46:11-28.
29. Dennis JW, Granovsky M, Warren CE. Protein glycosylation in development and disease.
BioEssays 1999;21:412-21.
30. Chu YW, Yang PC, Yang SC, et al. Selection of invasive and metastatic subpopulation from a human lung adenocarcinoma cell line. Am J Respir Cell Mol Biol 1997;17:353-60.
31. Wei Q, Xu X, Cheng L, Legerski RJ, Ali-Osman F. Simultaneous amplification of four DNA repair genes and β-actin in human lymphocytes by multiplex reverse transcriptase-PCR.
32. Yu D, Wang SS, Dulki KM, Tsai CM, Nicolson GL, Hung MC. C-erbB2/neu overexpression enhances metastatic ppotential of human lung cancer cells by induction of