台大農藝系 遺傳學 601 20000 Chapter 19 slide 1
CHAPTER 7
DNA Mutation, DNA
Repair, and Transposable
Elements
Peter J. Russell
edited by Yue-Wen Wang Ph. D.
Dept. of Agronomy, NTU
台大農藝系 遺傳學 601 20000 Chapter 19 slide 2
Adaptation Versus Mutation
1. A question that received much attention in the early 20
thcentury was whether
mutation causes random variation leading to adaptation, or the environment induces
heritable adaptations.
a. Lamarckism is the doctrine of inheritance of acquired characteristics.
b. The random mutation doctrine says that sometimes chance changes happen to be adaptive, , thus altering phenotype by changing a protein (Figure 7.1).
c. The observation that phage T1-resistant E. coli arise could be interpreted to support either of these theories.
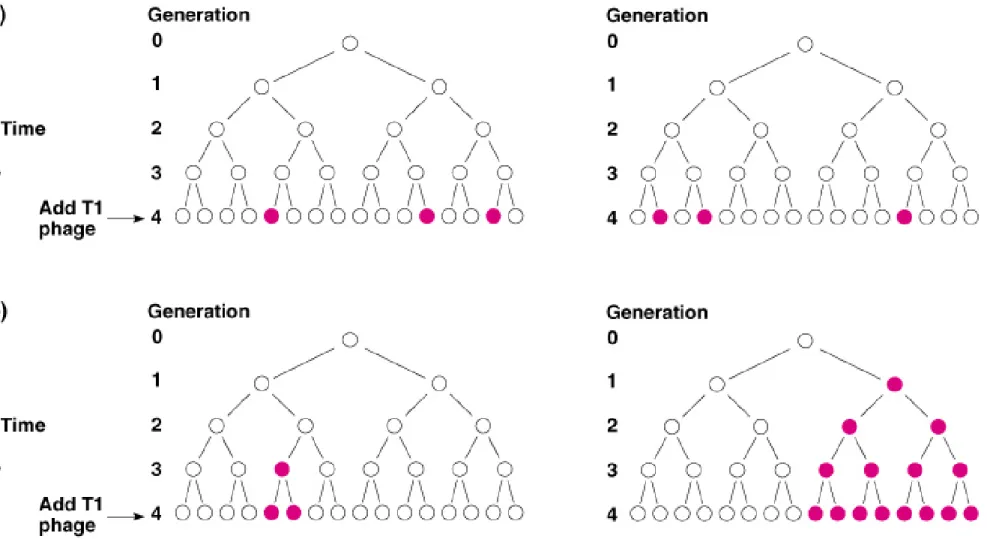
2. Luria and Delbrück (1943) demonstrated that the mutation mechanism is correct, by
doing a fluctuation test (Figure 7.2).
a. An E. coli population that started from one cell would show different patterns of T1 resistance depending on which model is correct.
i. The adaptive theory says that cells are induced to become resistant to T1 when it is added. Therefore, the proportion of resistant cells would be the same for all cultures with the same genetic background.
ii. The mutation theory says that random events confer resistance to T1, whether the phage is present or not. Cultures will therefore show different numbers of T1 resistant cells, depending on when the resistance mutation(s) occurred.
b. Luria and Delbrück observed fluctuating numbers of resistant bacteria from E. coli cultures, indicating that the random mutation model is correct.
台大農藝系 遺傳學 601 20000 Chapter 19 slide 3
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
Fig. 7.2 Representation of a dividing population of T1 phage-sensitive wild-type
台大農藝系 遺傳學 601 20000 Chapter 19 slide 4
Mutations Defined
1. A mutation is a change in a DNA base-pair or a
chromosome.
a. Somatic mutations affect only the individual in which they arise.
b. Germ-line mutations alter gametes, affecting the next generation.
2. Mutations are quantified in two different ways:
a. Mutation rate is the probability of a particular kind of mutation
as a function of time (e.g., number per gene per generation).
b. Mutation frequency is number of times a particular mutation
occurs in proportion to the number of cells or individuals in a
population (e.g., number per 100,000 organisms).
台大農藝系 遺傳學 601 20000 Chapter 19 slide 5
Types of Point Mutations
Animation: Nonsense Mutation and Nonsense Suppressor Mutation
1. There are two general categories of point mutations: base-pair
substitutions and base-pair deletions or insertions.
2. A base-pair substitution replaces 1 base-pair with another. There are
two types (Figure 7.3):
a. Transitions convert a pyrimidine pair to the other
purine-pyrimidine pair (e.g., AT to GC or TA to CG).
b. Transversions convert a purine-pyrimidine pair to a pyrimidine-purine
pair (e.g., AT to TA, or AT to CG).
3. Base-pair substitutions in ORFs are also defined by their effect on the
protein sequence. Effects vary from none to severe.
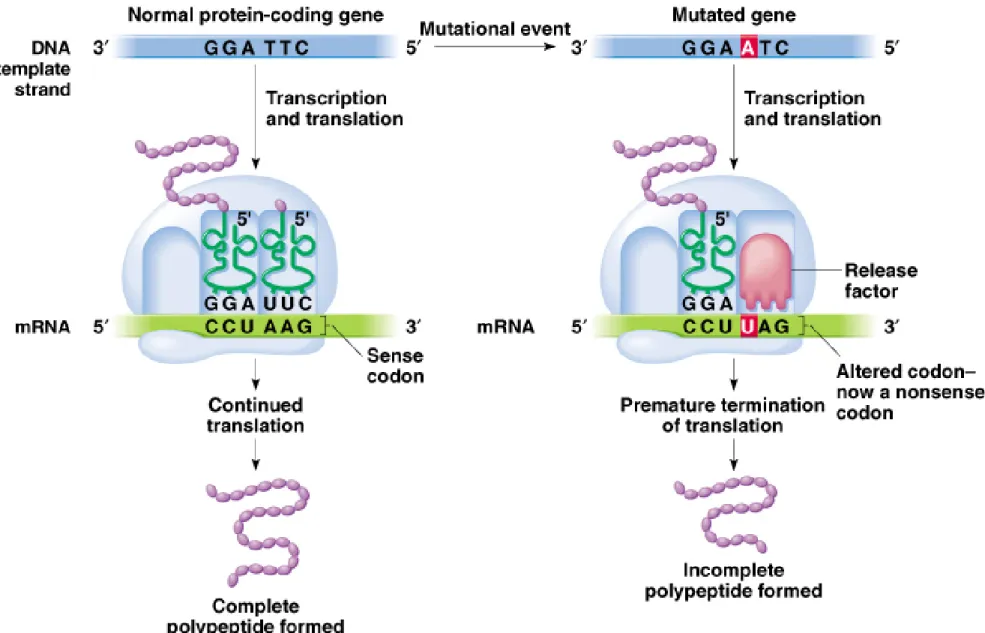
a. Nonsense mutations change a codon in the ORF to a stop
(nonsense) codon, resulting in premature termination of translation,
and a truncated (often nonfunctional) protein (Figure 7.4).
台大農藝系 遺傳學 601 20000 Chapter 19 slide 6
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
台大農藝系 遺傳學 601 20000 Chapter 19 slide 7
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
台大農藝系 遺傳學 601 20000 Chapter 19 slide 8
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
台大農藝系 遺傳學 601 20000 Chapter 19 slide 9 b. Missense mutations have a base-pair change resulting in a different mRNA codon,
and therefore a different amino acid in the protein.
c. Phenotypic effects may or may not occur, depending on the specific amino acid change.
i. Neutral mutations change a codon in the ORF, but the resulting amino acid substitution produces no detectable change in the function of the protein (e.g., AAA to AGA substitutes arginine for lysine. The amino acids have similar properties, so the protein’s function may not be altered).
ii. Silent mutations occur when the mutant codon encodes the same amino acid as the wild-type gene, so that no change occurs in the protein produced (e.g.,
AAA and AAG both encode lysine, so this transition would be silent).
4. Deletions and insertions can change the reading frame of the mRNA
downstream of the mutation, resulting in a frameshift mutation.
a. When the reading frame is shifted, incorrect amino acids are usually incorporated. b. Frameshifts may bring stop codons into the reading frame, creating a shortened
protein.
c. Frameshifts may also result in read-through of stop codons, resulting in a longer protein.
d. Frameshift mutations result from insertions or deletions when the number of affected base pairs is not divisible by three.
台大農藝系 遺傳學 601 20000 Chapter 19 slide 10
Reverse Mutations and Suppressor Mutations
1. Point mutations are divided into two classes based on their effect on phenotype:
a. Forward mutations change the genotype from wild type to mutant.
b. Reverse mutations (reversions or back mutations) change the genotype from mutant to wild-type or partially wild-type.
i. A reversion to the wild-type amino acid in the affected protein is a true reversion.
ii. A reversion to some other amino acid that fully or partly restores protein function is a partial reversion.
2. Suppressor mutations occur at sites different from the original mutation, and
mask or compensate for the initial mutation without actually reversing it.
Suppressor mutations have different mechanisms depending on the site at
which they occur.
a. Intragenic suppressors occur within the same gene as the original mutation, but at a different site. Two different types occur:
i. A different nucleotide is altered in the same codon as the original mutation. ii. A nucleotide in a different codon is altered (e.g., an insertion frameshift is
台大農藝系 遺傳學 601 20000 Chapter 19 slide 11 b. Intergenic suppressors occur in a different gene (the suppressor gene) from the
original mutation. Many work by changing mRNA translation.
i. Each suppressor gene works on only one type of nonsense, missense or frameshift mutation.
ii. A given suppressor gene suppresses all mutations for which it is specific.
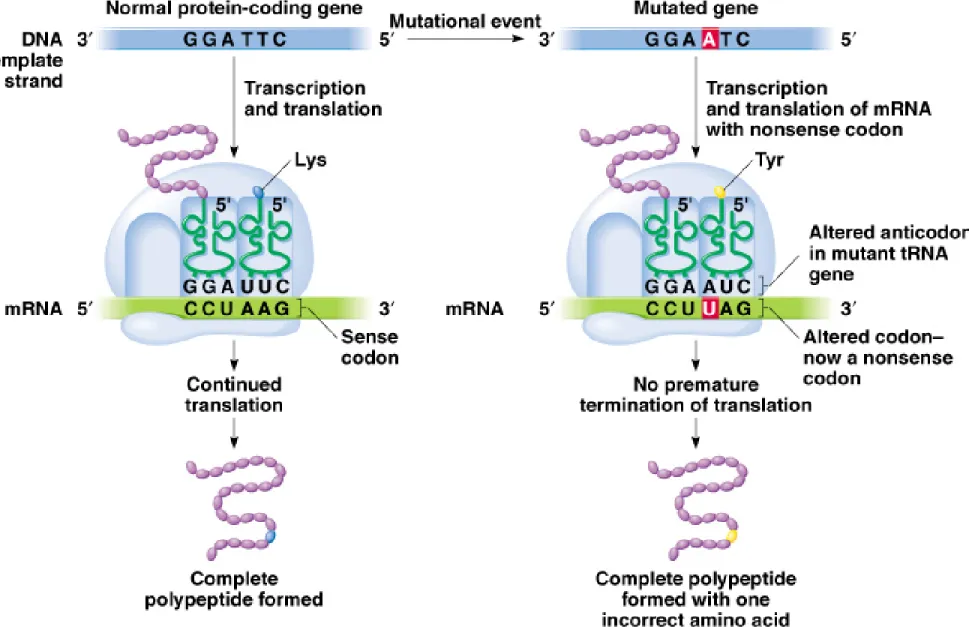
iii. Suppressor genes often encode tRNAs that recognize stop codons and insert an amino acid, preventing premature termination of translation.
(1) Full or partial function of the polypeptide may be restored.
(2) The effect depends on how compatible the substituted amino acid is with protein function.
iv. Nonsense suppressors fall into three classes, one for each stop codon (UAG, UAA and UGA) (Figure 19.5).
v. Typical tRNA suppressor mutations are in redundant tRNA genes, so the wild-type tRNA activity is not lost.
vi. Nonsense suppression occurs by competition between release factors and suppressor tRNAs.
(1) UAG and UGA suppressor tRNAs do well in competition with release factors.
(2) UAA suppressor tRNAs are only 1–5% efficient.
vii. Suppression by a tRNA occurs at all of its specific stop codons (e.g., UGA or UAG), not just the mutant one. This may produce read-through proteins, but they are not as common as expected, possibly due to tandem stop codons (e.g., UAGUGA).
台大農藝系 遺傳學 601 20000 Chapter 19 slide 12
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
Fig. 7.5 Mechanism of action of an intergenic nonsense suppressor mutation that
台大農藝系 遺傳學 601 20000 Chapter 19 slide 13
Spontaneous and Induced Mutations
1. Most mutations are spontaneous, rather than
induced by a mutagen.
台大農藝系 遺傳學 601 20000 Chapter 19 slide 14
Spontaneous Mutations
1. All types of point mutations can occur spontaneously, during S, G
1and
G
2phases of the cell cycle, or by the movement of transposons.
2. The spontaneous mutation rate in eukaryotes is between 10
-4-to-10
-6per
gene per generation, and in bacteria and phages 10
-5-to-10
-7/
gene/generation.
a. Genetic constitution of the organism affects its mutation rate.
i. In Drosophila, males and females of the same strain have similar
mutation rates.
ii. Flies of different strains, however, may have different mutation
rates.
b. Many spontaneous errors are corrected by the cellular repair systems,
and so do not become fixed in DNA.
台大農藝系 遺傳學 601 20000 Chapter 19 slide 15
DNA Replication Errors
1. DNA replication errors can be either point mutations, or small insertions or
deletions.
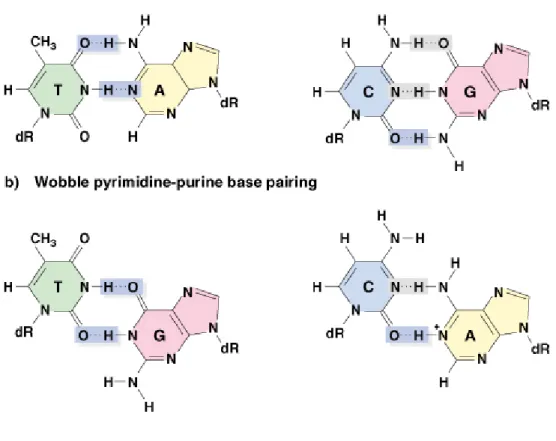
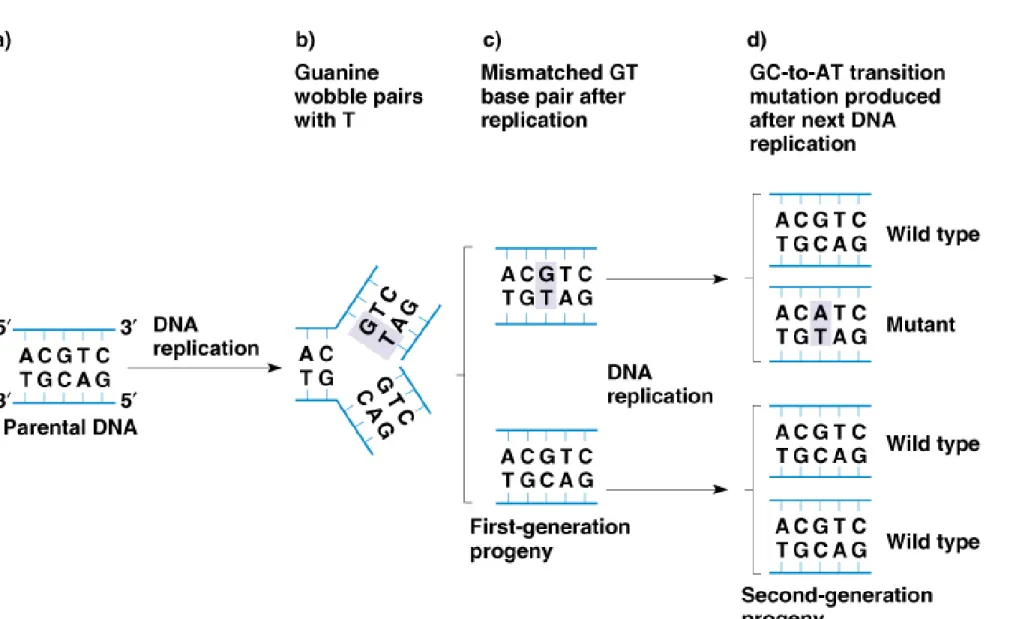
2. Base-pair substitution mutations can result from “wobble” pairing. Bases are
normally in the keto form, but sometimes can undergo tautomeric shift to
form the abnormal (enol) form. The enol form can hydrogen bond with an
incorrect partner due to different spatial positioning of the atoms involved in
H–bonding (Figure 7.6). An example is a GC-to-AT transition (Figure 7.7):
a. During DNA replication, G could wobble pair with T, producing a GT pair.
b. In the next round of replication, G and A are likely to pair normally,
producing one progeny DNA with a GC pair, and another with an AT pair.
c. GT pairs are targets for correction by proofreading during replication, and
by other repair systems. Only mismatches uncorrected before the next round
of replication lead to mutations.
台大農藝系 遺傳學 601 20000 Chapter 19 slide 16
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
台大農藝系 遺傳學 601 20000 Chapter 19 slide 17
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
Fig. 7.7 Production of a mutation as a result of a mismatch caused by wobble base
台大農藝系 遺傳學 601 20000 Chapter 19 slide 18
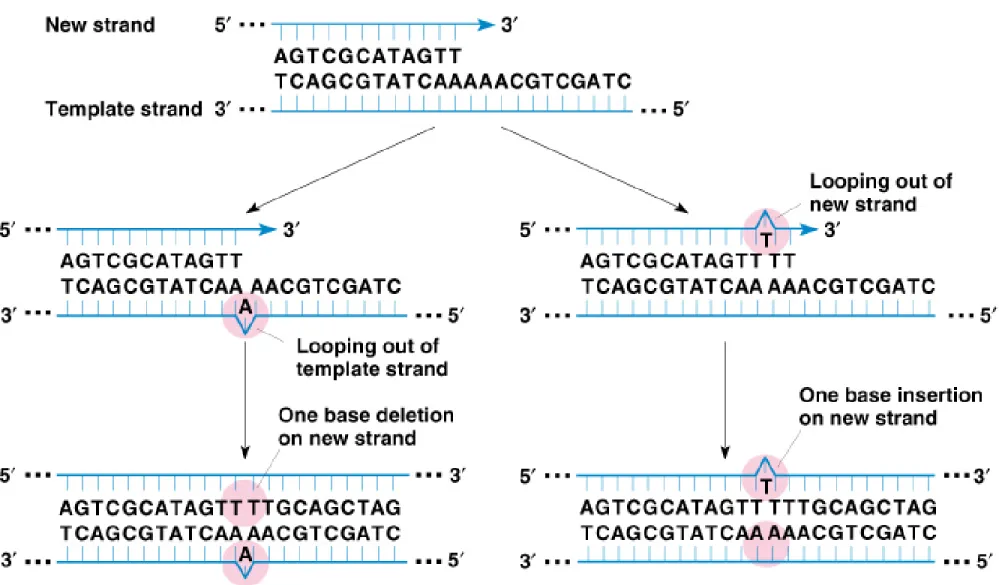
3. Additions and deletions can occur spontaneously during
replication (Figure 7.8).
a. DNA loops out from the template strand, generally in a run of
the same base.
b. DNA polymerase skips the looped out bases, creating a deletion
mutation.
c. If DNA polymerase adds untemplated base(s), new DNA looping
occurs, resulting in additional mutation.
d. Insertions and deletions in structural genes generate frameshift
mutations (especially if they are not multiples of three).
台大農藝系 遺傳學 601 20000 Chapter 19 slide 19
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
Fig. 7.8 Spontaneous generation of addition and deletion mutants by DNA looping-out errors during replication
台大農藝系 遺傳學 601 20000 Chapter 19 slide 20
4. Spontaneous chemical changes include depurination and deamination of
particular bases, creating lesions in the DNA.
a. Depurination removes the purine (A or G) from DNA by breaking the
bond with its deoxyribose in the backbone.
i. Depurination is common.
ii. If not repaired before the next round of replication, it will result in
a random base at that site.
b. Deamination removes an amino group from a base (e.g., cytosine to
uradil) (Figure 7.9).
i. Uracil is an abnormal base in DNA, and it will usually be repaired.
ii. If uracil is not replaced, it will pair with an A during replication,
resulting in a CG-to-TA transition.
iii. Both prokaryotic and eukaryotic DNA have small amounts of
5-methylcytosine (5
mC) in place of the normal C.
(1) Deamination of 5
mC produces T.
(2) T is a normal nucleotide in DNA, so it is not detected by repair
mechanisms.
(3) Deamination of 5
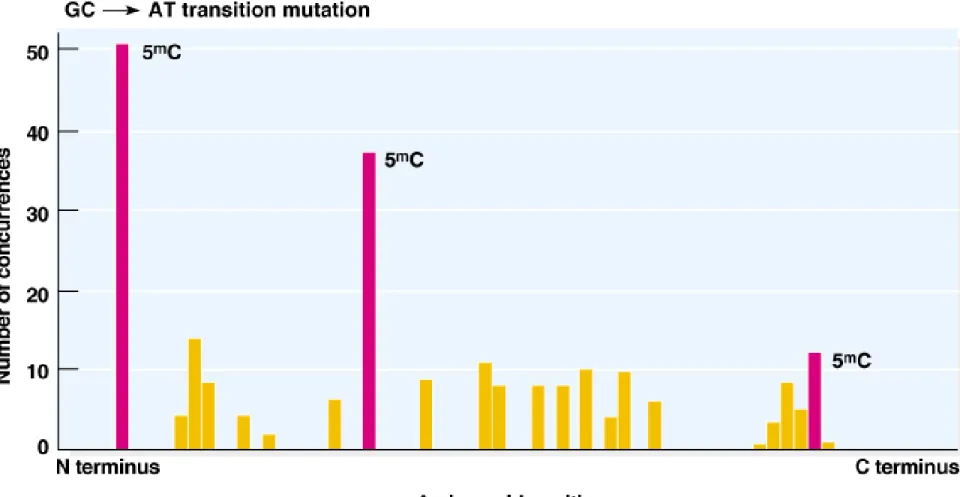
mC results in CG-to-TA transitions.
(4) Locations of 5
mC in the chromosome are often detected as
台大農藝系 遺傳學 601 20000 Chapter 19 slide 21
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
Fig. 7.9 Deamination of cytosine to uracil (a); deamination of 5-methylcytosine to
台大農藝系 遺傳學 601 20000 Chapter 19 slide 22
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
Fig. 7.x Distribution of spontaneous GC-to-AC transition mutations to stop codons
台大農藝系 遺傳學 601 20000 Chapter 19 slide 23
Induced Mutations
1. Exposure to physical mutagens plays a role in genetic research, where they are used to increase mutation frequencies to provide mutant organisms for study.
2. Radiation (e.g., X rays and UV) induces mutations.
a. X rays are an example of ionizing radiation, which penetrates tissue and collides with molecules, knocking electrons out of orbits and creating ions.
i. Ions can break covalent bonds, including those in the DNA sugar-phosphate backbone. ii. Ionizing radiation is the leading cause of human gross chromosomal mutations.
iii. Ionizing radiation kills cells at high doses, and lower doses produce point mutations. iv. Ionizing radiation has a cumulative effect. A particular dose of radiation results in the
same number of mutations whether it is received over a short or a long period of time. b. Ultraviolet (UV) causes photochemical changes in the DNA.
i. UV is not energetic enough to induce ionization.
ii. UV has lower-energy wavelengths than X rays, and so has limited penetrating power. iii. However, UV in the 254–260 nm range is strongly absorbed by purines and pyrimidines,
forming abnormal chemical bonds.
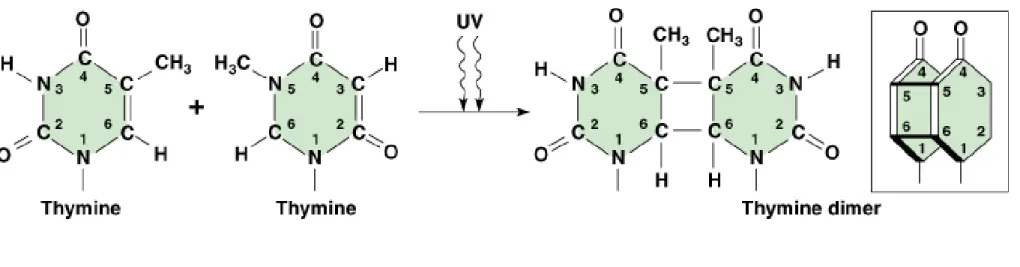
(1)A common effect is dimer formation between adjacent pyrimidines, commonly thymines (designated T^T) (Figure 7.10).
(2)C^C, C^T and T^C dimers also occur, but at lower frequency. Any pyrimidine dimer can cause problems during DNA replication.
(3)Most pyrimidine dimers are repaired, because they produce a bulge in the DNA helix. If enough are unrepaired, cell death may result.
台大農藝系 遺傳學 601 20000 Chapter 19 slide 24
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
台大農藝系 遺傳學 601 20000 Chapter 19 slide 25
Chemical Mutagens
Animation: Mutagenic Effects of 5BU
1. Chemical mutagens may be naturally occurring, or synthetic. They form
different groups based on their mechanism of action:
a. Base analogs depend upon replication, which incorpocates a base with
alternate states (tautomers) that allow it to base pair in alternate ways,
depending on its state.
i. Analogs are similar to normal nitrogen bases, and so are incorporated
into DNA readily.
ii. Once in the DNA, a shift in the analog’s form will cause incorrect
base pairing during replication, leading to mutation.
iii. 5-bromouradil (5BU) is an example. 5BU has a bromine residue
instead of the methyl group of thymine (Figure 7.11).
(1) Normally 5BU resembles thymine, pairs with adenine and is
incorporated into DNA during replication.
(2) In its rare state, 5BU pairs only with guanine, resulting in a
TA-to-CG transition mutation.
(3) If 5BU is incorporated in its rare form, the switch to its normal
state results in a CG-to-TA transition. Thus 5BU- induced
mutations may be reverted by another exposure to 5BU.
iv. Not all base analogs are mutagens, only those that cause base- pair
changes (e.g, AZT is a stable analog that does not shift).
台大農藝系 遺傳學 601 20000 Chapter 19 slide 26
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
台大農藝系 遺傳學 601 20000 Chapter 19 slide 27
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
台大農藝系 遺傳學 601 20000 Chapter 19 slide 28 b. Base-modifying agents can induce mutations at any stage of the cell cycle. They work
by modifying the chemical structure and properties of the bases. Three types are (Figure 7.12):
i. Deaminating agents remove amino groups. An example is nitrous acid (HNO2 ),
which deaminates G, C and A.
(1) HNO2 deaminates guanine to produce xanthine, which has the same base
pairing as G. No mutation results.
(2) HNO2 deaminates cytosine to produce uradil, which produces a CG-to-TA
transition.
(3) HNO2 deaminates adenine to produce hypoxanthine, which pairs with
cytosine, causing an AT-to-GC transition.
(4) Mutations induced by HNO2 can revert with a second treatment.
ii. Hydroxylating agents include hydroxylamine (NH2OH).
(1) NH2OH specifically modifies C with a hydroxyl group (OH), so that it pairs
only with A instead of with G.
(2) NH2OH produces only CG-to-TA transitions, and so revertants do not occur
with a second treatment.
(3) NH2OH mutants, however, can be reverted by agents that do cause
TA-to-CG transitions (e.g., 5BU and HNO2).
iii. Alkylating agents are a diverse group that add alkyl groups to bases. Usually alkylation occurs at the 6-oxygen of G, producing O6-allcylguanine.
(1) An example is methylmethane sulfonate (MMS), which methylates G to produce O6-alkyl G.
台大農藝系 遺傳學 601 20000 Chapter 19 slide 29
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
台大農藝系 遺傳學 601 20000 Chapter 19 slide 30
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
台大農藝系 遺傳學 601 20000 Chapter 19 slide 31
c. Intercalating agents insert themselves between adjacent bases in
dsDNA(Figure. 7.13). They are generally thin, plate-like hydrophobic
molecules.
i. At replication, a template that contains an intercalated agent will
cause insertion of a random extra base.
ii. The base-pair addition is complete after another round of
replication, during which the intercalating agent is lost.
iii. If an intercalating agent inserts into new DNA in place of a
normal base, the next round of replication will result in a deletion
mutation.
iv. Point deletions and insertions in ORFs result in frameshift
mutations. These mutations show reversion with a second
treatment.
台大農藝系 遺傳學 601 20000 Chapter 19 slide 32
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
台大農藝系 遺傳學 601 20000 Chapter 19 slide 33
Site-Specific in vitro Mutagenesis of DNA
1. Recombinant DNA technology allows genes to be
mutated at specific positions, and then introduced
back into cells.
2. This allows study of genes of unknown function
and sequences used to regulate gene expression.
台大農藝系 遺傳學 601 20000 Chapter 19 slide 34
Chemical Mutagens in the Environment
Animation: Ames Test Protocol
1. A wide variety of chemicals exist in our environment, and many can have mutagenic effects that can lead to genetic diseases and cancer. Examples include:
a. Drugs b. Cosmetics c. Food additives d. Pesticides
e. Industrial compounds
f. Chemical warfare agents such as mustard gas
2. The Ames test is a simple and inexpensive screen for potential carcinogens. It assays the reversion rate of mutant strains of Salmonella typhimurium back to wild-type (Figure 7.14).
a. Histidine (his) auxotrophs are tested for reversion in the presence of the chemical, by plating on media lacking the amino acid histidine.
i. Different his tester strains are available, to test for base-substitution and frameshift mutations.
ii. Liver enzymes (the S9 extract) are mixed with the test chemical to determine whether the liver’s detoxification pathways convert it to a mutagenic form.
iii. More revertants in the region of the test chemical than in the untreated control indicates that it may be a mutagen, and further tests are indicated.
b. The Ames test is used routinely to screen industrial and agricultural chemicals, and shows a good, but not perfect, correlation between mutagens and carcinogens.
c. The test can be made quantitative to produce a dose-response curve, allowing comparison of relative mutagenicity of different chemicals.
台大農藝系 遺傳學 601 20000 Chapter 19 slide 35
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
Fig. 7.14 The Ames test for assaying the potential mutagenicity of chemicals
台大農藝系 遺傳學 601 20000 Chapter 19 slide 36
Detecting Mutation
1. Mutants are often studied by geneticists.
Generally, mutation in haploid organisms is
readily detected, while recessive mutations in
diploid organisms are more difficult to
characterize. The problem is compounded in
humans, where controlled crosses cannot be done.
2. For some organisms, especially microorganisms,
台大農藝系 遺傳學 601 20000 Chapter 19 slide 37
Visible Mutations
1. Some mutations affect the appearance of an
organism (e.g., Drosophila eyes or wing-shape,
coat color in animals, colony size in yeast, plaque
morphology of phages).
台大農藝系 遺傳學 601 20000 Chapter 19 slide 38
Nutritional Mutations
1. Auxotrophic mutants are easily detected for
microorganisms that normally can grow on minimal
medium, using methods that have been developed for
selection and screening.
2. An example is replica plating. Cells are first grown on
supplemented medium, and then the colonies transferred
to minimal medium, as well as to a control plate of
supplemented medium. Colonies that grow on
supplemented, but not minimal, media are selected for
further study.
台大農藝系 遺傳學 601 20000 Chapter 19 slide 39
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
Fig. 7.15 Replica-plating technique to screen for mutant strains of a colony-forming microorganism
台大農藝系 遺傳學 601 20000 Chapter 19 slide 40
Conditional Mutations
1. Mutations in some genes (e.g., DNA and RNA
polymerases) are usually lethal, so these genes are
studied by isolating conditional mutations.
2. Heat sensitivity is a common conditional
mutation, in which a normal protein is produced
at permissive temperature, and a nonfunctional
protein results at the nonpermissive temperature.
Screening is generally by replica plating and
台大農藝系 遺傳學 601 20000 Chapter 19 slide 41
Resistance Mutations
1. Microorganisms like E. coli and yeast are easily
screened for resistance to viruses, chemicals or
drugs, because resistant cells will grow when
wild-type cells will not.
台大農藝系 遺傳學 601 20000 Chapter 19 slide 42
Repair of DNA Damage
1. Both prokaryotes and eukaryotes have
enzyme-based DNA repair systems that prevent mutations
and even death from DNA damage.
2. Repair systems are grouped by their repair
mechanisms. Some directly correct, while others
excise the damaged area and then repair the gap.
台大農藝系 遺傳學 601 20000 Chapter 19 slide 43
Direct Reversal of DNA Damage
1. DNA polymerase proofreading corrects most of the incorrect nucleotide insertions that occur during DNA synthesis, which stalls until the wrong nucleotide is replaced with a correct one.
a. The role of 3’-to-5’ exonuclease activity is illustrated by mutator mutations in E. coli, which confer a much higher mutation rate on the cells that carry them.
b. The mutD gene, encoding the e subunit of DNA polymerase III, is an example. Cells mutant in mutD are defective in proofreading.
2. UV-induced pyrimidine dimers are repaired using photoreactivation (light repair).
a. Near UV light (320–370 nm) activates photolyase (product of the phr gene) to split the dimer.
b. Photolyases are found in prokaryotes and simple eukaryotes, but not in humans.
3. Damage by alkylation (usually methyl or ethyl groups) can be removed by specific DNA repair enzymes.
a. For example, O6-methylguanine methyltransferase (from the ada gene) recognizes O6
-methylguanine in DNA, and removes the methyl group, restoring the base to its original form.
b. A similar system repairs alkalated thymine.
4. Mutations of repair enzyme genes increase the organism’s rate of spontaneous mutations.
台大農藝系 遺傳學 601 20000 Chapter 19 slide 44
Base Excision Repair
1. Base excision repair uses a repair glycosylase
enzyme to recognize and remove damaged bases.
a. Bond between base and deoxyribose is cleaved by the
glycosylase.
b. Other enzymes cleave sugar-phosphate from
backbone, leaving a gap in the DNA.
c. Repair DNA polymerase and DNA ligase use the
opposite strand as template to fill the gap.
台大農藝系 遺傳學 601 20000 Chapter 19 slide 45
Repair Involving Excision of Nucleotides
1. A repair system, which does not require light, was discovered
in 1964. It is called dark repair, the excision repair system, or
the nucleotide excision repair (NER) system.
a. In E. coli, NER corrects pyrimidine dimers and other damage-induced distortions of the DNA helix.
b. The proteins required are UvrA, UvrB, UvrC and UvrD (encoded by genes of the same name) (Figure 7.16).
c. A complex of two UvrA and one UvrB proteins slides along the DNA. When it encounters a helix distortion, the UvrA subunits dissociate, and a UvrC binds the UvrB at the lesion.
d. When UvrBC forms, the UvrC cuts 4–5 nucleotides from the lesion on the 3’ side, and eight nucleotides away on the 5’ side. Then UvrB is released and UvrD binds the 5’ cut end.
e. UvrD is a helicase that unwinds the region between the cuts, releasing the short ssDNA, while DNA polymerase I fills the gap and DNA ligase seals the backbone.
f. In yeast and mammalian systems, about 12 genes encode proteins involved in excision repair.
台大農藝系 遺傳學 601 20000 Chapter 19 slide 46
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
Fig. 7.16 Nucleotide excision repair (NER) of pyrimidine dimmer and other
台大農藝系 遺傳學 601 20000 Chapter 19 slide 47
2. Methyl-directed mismatch repair recognizes mismatched base pairs, excises the
incorrect bases and then carries out repair synthesis.
a. In E. coli, initial stages involve products of the mutS, mutL and mutH genes (Figure 7.17).
i. MutS binds the mismatch, and determines which is the new strand by its lack of methylation.
ii. MutL and MutH bind unmethylated GATC sequences (site of methylation in E.
coli) and bring the GATC close to the mismatch by binding MutS.
iii. MutH then nicks the unmethylated GATC site, the mismatch is removed by an exonuclease and the gap is repaired by DNA polymerase III and ligase.
b. Eukaryotes also have mismatch repair, but it is not clear how old and new DNA strands are identified.
i. Four genes are involved in humans, hMSH2 (homologous to E. coli mutS), and
hMLH1, hPMS1 and hPMS2 (all homologous to mutL).
ii. All of these are mutator genes, and mutation in any 1 of them confers
台大農藝系 遺傳學 601 20000 Chapter 19 slide 48
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
台大農藝系 遺傳學 601 20000 Chapter 19 slide 49
3. Translesion DNA synthesis is used to allow a cell to survive when specific
base-pairing cannot occur. Survival is usually at the cost of incurring new
mutations.
a. Bacteria use a system called SOS. In E. coli SOS is controlled by two genes, lexA and recA. Mutants in either of these genes have their SOS response permanently turned on.
i. When no DNA damage is present, LexA represses transcription of about 17 genes with products involved in various types of DNA repair.
ii. Sufficient DNA damage activates the RecA protein, which stimulates LexA to autocleave, removing repression of the DNA repair genes.
iii. After damage is repaired, RecA is inactivated, and newly synthesized LexA again represses the DNA repair genes.
b. The SOS system is a mutagenic bypass synthesis system.
i. DNA polymerase for translesion synthesis is made in SOS response. It replicates over and past the lesion.
ii. Nucleotides are incorporated at the lesion that may not match wild type, leading to mutations.
台大農藝系 遺傳學 601 20000 Chapter 19 slide 50
Human Genetic Diseases Resulting from DNA
Replication and Repair Mutations
1. Many human genetic disorders result from gene
mutations, and are included in the OMIM website
(http://www3.ncbi.nlm.nih.gov/OMIM).
2. Some human genetic diseases resulting from defects in
DNA replication or repair are listed in Table 7.1.
Xeroderma pigmentosum is an example (Figure 7.18).
a. The disorder occurs in homozygotes for a mutation in a repair
gene.
b. Affected individuals are photosensitive, and portions of skin
exposed to light show intense pigmentation and warty growths
that may become malignant.
c. The defect is in excision repair, and the inability to repair
radiation damage to DNA often results in malignancies.
台大農藝系 遺傳學 601 20000 Chapter 19 slide 52
General Features of Transposable Elements
1.
Transposable elements are divided into two classes on the basis of their
mechanism for movement:
a. Some encode proteins that move the DNA directly to a new position or replicate the DNA to produce a new element that integrates elsewhere. This type is found in both prokaryotes and eukaryotes.
b. Others are related to retroviruses, and encode reverse transcriptase for making DNA copies of their RNA transcripts, which then integrate at new sites. This type is found only in eukaryotes.
2.
Transposition is nonhomologous recombination, with insertion into DNA
that has no sequence homology with the transposon.
a. In prokaryotes, transposition can be into the cell’s chromosome, a plasmid or a phage chromosome.
b. In eukaryotes, insertion can be into the same or a different chromosome.
3.
Transposable elements can cause genetic changes, and have been involved in
the evolution of both prokaryotic and eukaryotic genomes. Transposons may:
a. Insert into genes.
b. Increase or decrease gene expression by insertion into regulatory sequences.
c. Produce chromosomal mutations through the mechanics of transposition.
台大農藝系 遺傳學 601 20000 Chapter 19 slide 53
Transposable Elements in Prokaryotes
1. Prokaryotic examples include:
a. Insertion sequence (IS) elements.
b. Transposons (Tn).
台大農藝系 遺傳學 601 20000 Chapter 19 slide 54
Insertion Sequences
Animation: Insertion Sequences in Prokaryotes
1. IS elements are the simplest transposable elements found in
prokaryotes, encoding only genes for mobilization and insertion of its
DNA. IS elements are commonly found in bacterial chromosomes and
plasmids.
2. IS elements were first identified in E. coli’s galactose operon,
wheresome mutations’ were shown to result from insertion of a DNA
sequence now called IS1 (Figure 7.19)
3. Prokaryotic IS elements range in size from 768bp to over 5kb. Known
E. coli IS elements include IS1 which is 768bp long and present in 4–
19 copies on the E. coli chromosome, IS2, and IS10R.
4. The ends of all sequenced IS elements show inverted terminal repeats
(IRs) of 9–41bp. IS1 has 23bp of nearly identical sequence.
台大農藝系 遺傳學 601 20000 Chapter 19 slide 55
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
台大農藝系 遺傳學 601 20000 Chapter 19 slide 56
5. Random integration of IS elements may:
a. Disrupt coding sequences or regulatory regions.
b. Alter expression of nearby genes by the action of IS element promoters.
c. Cause deletions and inversions in adjacent DNA.
d. Serve as a site for crossing-over between duplicated IS elements.
6. When an IS element transposes:
a. Transposition requires transposase, an enzyme encoded by the IS element.
b. Transposase recognizes the IR sequences to initiate transposition at a
frequency characteristic for each IS element (range is 105 to 107 per
generation).
c. IS elements insert into the chromosome without sequence homology
(illegitimate recombination) at target sites (Figure 7.20).
i. A staggered cut is made in the target site, and the IS element inserted.
ii. DNA polymerase and ligase fill the gaps, producing small direct
台大農藝系 遺傳學 601 20000 Chapter 19 slide 57
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
Fig. 7.20 Schematic of the integration of an IS element into chromosomal DNA
台大農藝系 遺傳學 601 20000 Chapter 19 slide 58
Transposons
1. Transposons are similar to IS elements, but carry additional genes, and
have a more complex structure. There are two types of prokaryotic
transposons:
a. Composite transposons carry genes (e.g., antibiotic resistance) flanked
on both sides by IS elements (IS modules).
i. The IS elements are of the same type, and called ISL (left) and ISR
(right).
ii. ISL and ISR may be in direct or inverted orientation to each other.
iii.Transposition of composite transposons results from the IS
elements, which supply transposase and its recognition signals, the
IRs (Figure 7.21).
(1) Tn10’s transposition is rare (
< 10
-7/molecule/generation).
(2) Transposons, like IS elements, produce target site
duplications (e.g., a 9-bp duplication for Tn10).
台大農藝系 遺傳學 601 20000 Chapter 19 slide 59
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
台大農藝系 遺傳學 601 20000 Chapter 19 slide 60
b. Noncomposite transposons also carry
genes (e.g., drug resistance) but do not
terminate with IS elements.
i. Transposition proteins are encoded in
the central region.
ii. The ends are repeated sequences (but
not IS elements).
iii. Noncomposite transposons cause
target site duplications (like composite
transposons).
iv. An example is Tn3, which produces a
5-bp duplication upon insertion.
台大農藝系 遺傳學 601 20000 Chapter 19 slide 61
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
台大農藝系 遺傳學 601 20000 Chapter 19 slide 62
2. Models have been generated for transposition:
a. Cointegration is an example of the replicative transposition that
occurs with Tn3 and its relatives (Figure 7.22).
i. Donor DNA containing the Tn fuses with recipient DNA.
ii. The Tn is duplicated, with one copy at each donor-recipient DNA
junction, producing a cointegrate.
iii. The cointegrate is resolved into two products, each with one copy
of the Tn.
b. Conservative (nonreplicative) transposition is used by Tn10, for
example. The Tn is lost from its original position when it transposes.
3. Transposons cause the same sorts of mutations caused by IS elements:
a. Insertion into a gene disrupts it.
b. Gene expression is changed by adjacent Tn promoters.
c. Deletions and insertions occur.
台大農藝系 遺傳學 601 20000 Chapter 19 slide 63
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
Fig. 7.22 Cointegration model for transposition of a transposable element by
台大農藝系 遺傳學 601 20000 Chapter 19 slide 64
IS Elements in the E. coli F factor
1. The E. coli F factor is an episome, able to
integrate into the bacterial chromosome and
capable of self-replication.
a. The F plasmid contains four IS elements: two copies
of IS3, one of IS2, and one of γδ(gamma-delta).
b. The E. coli chromosome also has copies of these
insertion sequences in different sites and orientations.
c. Homologous recombination moves the F factor into
台大農藝系 遺傳學 601 20000 Chapter 19 slide 65
Transposable Elements in Eukaryotes
1. Rhoades (1930s) working with sweet corn, observed
interactions between two genes:
a. A gene for purple seed color, the Al locus. Homozygous mutants
(a/a) have colorless seeds.
b. A gene on a different chromosome, Dt (dotted) that causes seeds
with genotype a/a Dt/-- to have purple dots.
i. Dt appears to mutate the a allele back to the Al wild-type in
regions of the seed, producing a dotted phenotype.
ii. The effect of the Dt allele is dose dependent.
(1) One dose gave an average of 7.2 dots per seed.
(2) Two doses gave an average of 22.2 dots/seed.
(3) Three doses gave an average of 121.9 dots/seed.
c. Rhoades interpreted Dt as a mutator gene.
2. McClintock (1940s-50s), working with corn (Zea mays)
proposed the existence of “controlling elements” that
regulate other genes and are mobile in the genome.
台大農藝系 遺傳學 601 20000 Chapter 19 slide 66
3. The genes studied by both Rhoades and McClintock have turned out to
be transposable elements, and many others have been identified in
various eukaryotes.
a. Most studied are transposons of yeast, Drosophila, corn and humans.
b. Their structure is very similar to that of prokaryotic transposable
elements.
c. Eukaryotic transposable elements have genes for transposition and
integration at a number of sites, as well as a variety of other genes.
d. Random insertion results from non-homologous recombination, and
means that any chromosomal gene may be regulated by a transposon.
4. The result of transposon insertion into a chromosome will depend on the
properties of the transposon, with possible effects including:
a. Chromosome mutations such as duplications, deletions, inversions,
translocations, or breakage.
b. Disruption of genes to produce a null mutation (gene is nonfunctional).
c. Activation or repression of adjacent genes by disrupting a cellular
台大農藝系 遺傳學 601 20000 Chapter 19 slide 67
Transposons in Plants
Animation: Transposable Elements in Plants
1. Plant transposons also have IR sequences, and generate short direct
target site repeats.
2. Several families of transposons have been identified in corn, each with
characteristic numbers, types and locations.
a. Each family has two forms of transposon. Either can insert into a gene
and produce a mutant allele.
i. Autonomous elements, which can transpose by themselves. Alleles
produced by an autonomous element are mutable alleles, creating
mutations that revert when the transposon is excised from the gene.
ii. Nonautonomous elements, which lack a transposition gene and
rely on the presence of another transposon to supply the missing
function. Mutation by these elements is stable (except when an
autonomous element from the family is also present).
台大農藝系 遺傳學 601 20000 Chapter 19 slide 68
3. Multiple genes control corn color, and classical genetics indicates that a
mutation in any of these genes leads to a colorless kernel. McClintock
studied the unstable mutation that produces spots of purple pigment on
white kernels (Figure 7.23).
a. She concluded that spots do not result from a conventional mutation,
but from a controlling element (now Tn).
b. A corn plant with genotype c/c will have white kernels, while C/-- will
result in purple ones.
i. If a reversion of c to C occurs in a cell, that cell will produce purple
pigment, and hence a spot.
ii. The earlier in development the reversion occurs, the larger the
spot.
台大農藝系 遺傳學 601 20000 Chapter 19 slide 69
iii. McClintock concluded that the c allele resulted from insertion of a
“mobile controlling element” into the C allele. (Figure 7.24)
(1) The element is Ds (dissociation), now known to be a
nonautonomous transposon.
(2) Its transposition is controlled by Ac (activator), an
autonomous transposon.
c. McClintock’s evidence of transposable elements did not fit the
prevailing model of a static genome. More recent studies have
confirmed and characterized the elements involved, the Ac-Ds system.
i. The autonomous Ac element is 4,563bp, with short inverted terminal
repeats and one gene (transposase). Insertion generates an 8-bp target
site duplication.
ii. The nonautonomous Ds elements vary in length and sequence, but
all have the same terminal IRs as Ac, and most are deleted or
台大農藝系 遺傳學 601 20000 Chapter 19 slide 70
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
台大農藝系 遺傳學 601 20000 Chapter 19 slide 71
iii. Ac transposes only during chromosome replication, by a
conservative transposition mechanism (Figure 7.25). There
are two possible results of Ac transposition, depending on
whether the target DNA has replicated or not.
(1) If Ac transposes during replication into a replicated
target site, its chromatid’s donor site will be empty since
that copy of Ac has inserted elsewhere. In the
homologous donor site on the other chromatid, a copy
will remain. There is no net increase in copies of Ac.
(2) Transposition to an unreplicated chromosome site also
leaves one donor site empty (and the other with a copy of
Ac). The DNA into which Ac inserts will then be
replicated, resulting in a net gain of one copy of Ac.
iv. Replication of Ds is the same, except that the
台大農藝系 遺傳學 601 20000 Chapter 19 slide 72
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
台大農藝系 遺傳學 601 20000 Chapter 19 slide 73
Ty Elements in Yeast
1. Ty element is diagrammed in Figure 7.26:
a. It is 5.9 kb including 2 terminal direct repeats of 334 bp, the long terminal repeats (LTR) or deltas (δ).
b. Each delta contains a promoter and transposase recognition sequences.
c. Ty elements encode one 5.7 kb mRNA beginning at the delta 5’ promoter (Figure 20.14).
d. There are two ORFs in the mRNA, designated TyA and TyB, encoding two different proteins.
e. Ty copy number varies between yeast strains, with an average of about 35.
2. Ty elements also share similarities with retroviruses, ssRNA viruses that replicate
via dsDNA intermediates.
a. Ty elements transpose by making an RNA copy of the integrated DNA sequence, then making DNA using reverse transcriptase. This DNA can integrate at a new
chromosomal site. Evidence for this includes:
i. An experimentally introduced intron in the Ty element (which normally lacks
introns) was monitored through transposition. The intron was removed, indicating an RNA intermediate.
ii. Ty elements encode a reverse transcriptase.
iii. Virus-like particles containing Ty RNA and reverse transcriptase activity occur. b. Ty elements are referred to as retrotransposons.
台大農藝系 遺傳學 601 20000 Chapter 19 slide 74
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
台大農藝系 遺傳學 601 20000 Chapter 19 slide 75
Drosophila transposons
1. It is estimated that 15% of the Drosophila genome is mobile!
The P element is an example. P elements vary from full-length
autonomous elements through shorter nonautonomous versions that
lack the transposase gene (Figure 7.27).
2. P elements are used experimentally to transfer genes into the germ
line of Drosophila embryos. For example (Figure 7.28):
a. The wild-type rosy
+(ry
+) gene was inserted into a P element,
cloned in a plasmid, and microinjected into a mutant ry/ry
strain.
b.Insertion of the recombinant P element into the recipient
chromosome introduced the ry
+allele, and produced wild-type
台大農藝系 遺傳學 601 20000 Chapter 19 slide 76
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
Fig. 7.27 Structure of the autonomous P transposable element found in
Drosophila
台大農藝系 遺傳學 601 20000 Chapter 19 slide 77
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
Fig. 7.28 Illustration of the use of P elements to introduce genes into the
Drosophila
台大農藝系 遺傳學 601 20000 Chapter 19 slide 78
Human Retrotansposons
1. Retrotransposons in humans include:a. LINEs (long interspersed sequences), repeated sequences >5kb interspersed with unique-sequence DNA up to 35kb. LINEs occur as:
i. Autonomous elements with functional transposition genes. ii. Nonautonomous derivatives with internal deletions.
b. SINEs (short interspersed sequences), 100–400bp repeats interspersed with unique-sequence DNA 1–2kb in length.
2. LINEs make up about 20% of the human genome. L1 is the best studied type.
a. L1 elements are about 5% of the human genome. The full-length element (6.5kb) is not abundant, and most L1 elements are deleted versions.
b. The full-length L1 element contains a large ORF with homology to known reverse transcriptases. Experimentally, the L1 ORF can substitute for the yeast Ty reverse transcriptase gene.
c. L1 elements transpose via RNA, but unlike other retrotransposons, do not have LTRs, so use a different mechanism for transposition.
d. Clinically, cases of hemophilia have been shown to result from newly transposed L1 insertions into the gene for factor VIII (required for normal blood clotting). This
insertional mutagenesis has been found in families where a child has L1 in the factor VIII gene, but neither parent does.
台大農藝系 遺傳學 601 20000 Chapter 19 slide 79 3. SINEs lack genes for transposition, and so are nonautonomous, requiring enzymes
encoded by LINEs in order to move. The Alu family is a very abundant type of SINE.
a. Alu sequences (named for the restriction site contained in their sequences) are about 300bp, repeated 300,000–500,000 times in the genome. Alu sequences make up to 3% of total human DNA.
b. A human case of a genetic disease, neurofibromatosis, provides some evidence. i. Neurofibromas (tumorlike growths on the body) result from an
autosomal dominant mutation.
ii. In a patient’s DNA, an unusual Alu sequence was detected in one of the introns of the neurofibromatosis gene.
iii. The resulting longer transcript is incorrectly processed, removing an exon from the mRNA and producing a nonfunctional protein.
iv.Neither parent had this Alu sequence in the neurofibromatosis gene. v. Divergent Alu sequences made it possible to track this particular version to an insertion event in the germ line of the patient’s father.