Journal
of
Orthopaedic
Research
ELSEVIER
Journal of Orthopaedic Research 23 (2005) 446453www.elsevier.com/locate/orthres
Type I and I1 collagen regulation
of chondrogenic differentiation
by mesenchymal progenitor cells
C.W. Chen
Y.H.
Tsai
b7c,1,W.P. Deng
a,S.N.
Shih
e,
C.L. Fang
d,
J.G. Burch
f,
W.H.
Chen
a,W.F.
Lai
a Institute of Medical Sciences, Taipei Medical University, 250 Wu-Hsing Street. Taipei 110, Taiwan Institute of Biomedical Materials, Taipei Medical University, 2.50 Wu-Hsing Street, Taipei 110, Taiwan Institute of Cellular and Molecular Biology, Taipei Medical University. 250 Wu-Hsing Street, Taipei 110, Taiwan
Department of Pathology, Taipei Medical University, 250 Wu-Hsing Street, Taipei 110, Taiwan Orthopedic Department, Chang-Gung Memorial Hospital, Taoyuan 333, Taiwan
'
Department of Orthodontics, School of Dental Medicine, Nova Southeastern University, Ft. Lauderdale, FL 33328, USA Received 19 December 2003; accepted 3 September 2004Abstract
Chondrogenic differentiation by mesenchymal progenitor cells (MPCs) is associated with cytokines such as transforming growth
factor-beta
1 (TGF-PI) and dexamethasone. Extracellular matrix (ECM) also regulates the differentiation by MPCs. To define
whether ECM plays a functional role in regulation of the chondrogenic differentiation by MPCs, an in vitro model was used. That
model exposed to dexamethasone, recombinant human TGF-@l(rhTGF-@I)
and collagens. The results showed that MPCs incorpo-
rated with dexamethasone and rhTGF-01 increased proliferation and expression of glycosaminoglycan (GAG) after 14 days. Type
11collagen enhanced the GAG synthesis, but did not increase alkaline phosphatase (ALP) activity. When adding dexamethasone
and rhTGF-PI MPCs increased mRNA expression
of
sox9. Incorporation with type
I1
collagen, dexamethasone and rhTGF-Pl,
MPCs induced mRNA expression of aggrecan and enhanced levels of type
I1
collagen, and sox9 mRNA. In contrast, incorporation
with type
I collagen, dexamethasone and rhTGF-PI MPCs reduced levels of aggrecan, and sox9 mRNA, and showed no type
I1
collagen mRNA. Altogether, these results indicate that type I and
I1collagen, in addition to the cytokine effect, may play a func-
tional role in regulating of chondrogenic differentiation by MPCs.
0
2004 Orthopaedic Research Society. Published by Elsevier Ltd. All rights reserved.
Keywords: Chondrogenic differentiation; Collagen; Dexamethasone; Extracellular matrix; Mesenchymal progenitor cells, Sox9; TGF-Dl
Introduction
Cartilage has little capacity for self-repair [lo]. An in-
jured joint predisposed to continued arthritic degenera-
tion [8]. Current therapies for cartilage regeneration
* Corresponding author. Tel.: +886 2 87800225/27361661~5210; E-mail address: [email protected] (W.F. Lai).
'
C.W. Chen and Y.H. Tsai contributed equally to this paper. fax: +886 2 27357714.include placement
of
carbon plugs [5], periosteum [ 141,
or periochondrium [
151.
Autologous chondrocyte trans-
plantation [6,3 13, and subchondral drilling [4,16,26].
Success rates vary. Most methods of therapy have seri-
ous clinical limitations.
Structural, chemical, and mechanical properties of
regenerated cartilage are definitely not normal [43].
The regenerated tissue does not bond to adjacent tissue
[38].
New cartilage undergoes degenerative changes.
After one year this new cartilage is similar to that in
untreated defects [25]. Thus, most repair methods fail
0736-0266/$
-
see front matter 0 2004 Orthopaedic Research Society. Published by Elsevier Ltd. All rights reserved doi: 10.101 6/j.orthres.2004.09.002C. W. Chen et al. I Journal of Orthopaedic Research 23 (2005) 4 4 6 4 5 3 447
to improve cartilage beyond what is found in natural re-
pair of untreated osteochondral defects.
The clinical need for improved treatment options for
patients with cartilage injuries has encouraged scientists
to investigate in vivo implantation of isolated MPCs.
MPCs can undergo differentiation to regenerate either
cartilage [13] or bone [7,44] in defects. Implantation of
MPCs has been demonstrated to affect tissue regenera-
tion. Then different environmental factors can be intro-
duced to trigger differentiation into specific phenotypes.
Chondrogenic differentiation has been induced in
vitro by cytokines TGF-PI and dexamethasone in
MPCs derived from bone marrow of human [46] or ani-
mal models [19,27,45]. ECM has been demonstrated to
regulate the chondrogenic or osteogenic differentiation
induced by MPCs [18,29]. How the mechanisms of
ECM affect MPC differentiation are still not well under-
stood. This study was undertaken to further identify
how collagen regulated rhTGF-b 1’s and dexametha-
sone’s effects on chondrogenic differentiation. Differen-
tial expression of MPC-induced glycosaminoglycan
(GAG)
and alkaline phosphatase (ALP) exposed to dif-
ferent concentrations of dexamethasone, rhTGF-Pl, and
collagens was measured by spectrophotometry. The
levels of mRNA expression of Sox9, aggrecan, and type
I1 collagen were found to be identifiers
of
certain mech-
anisms of chondrogenic differentiation by MPCs.
Materials and methods
Subjects
Consenting bone marrow donors were selected from patients admitted to the Orthopedic Section of Taipei Municipal Chung-Hsin Hospital, Taipei, Taiwan. None had endocrine disease or was receiving hormone replacement therapy. Bone marrow was obtained from femur fracture site by proximal femur aspiration during surgical treatment procedures.
Isolation and cultivation of MPCs
MPCs were isolated from rabbit and human bone marrow. They were mixed with sodium-heparin, and diluted with equal five volumes of PBS. The cell suspension was fractionated on a Percoll gradient (40% initial density, Phamacia). The MPC-enriched interface fraction was collected and cultured in Dulbecco’s modified Eagle medium with 1 g/ml glucose (DMEM/LG, Sigma D5523), 10% fetal bovine serum, l00UIml penicillin, 100 pglml streptomycin, and 0.25 pglrnl fungizone under normal conditions. The medium was changed every four days. During the primary culture, adherent cells formed colonies that were passaged when cells proliferated subconfluently. Second- and third- passage cells were selected to identify the mechanisms of differentiation. mRNA extraction and reverse transcriptase polymerase chain reaelion ( R T - P C R )
Total RNA harvested from subconfluent monolayer cultures (approximately1 O6 MPCs) was extracted using T R I Z O I ~ Reagent (Invitrogen life technologies, Carlsbad, CA). Extracted RNA was dis- solved in sterilized ddHlO and stored at -80°C. Reverse transcription was performed with SuperScript” 111 (Invitrogen life technologies) and
Oligo d(T)12-18 primer. Four micrograms of RNA was added into a final solution of 21pl l0mM dNTP mix, 10 X RT buffer, 25mM MgCI2, 0.1 M DTT, RNase Inhibitor and RNase H. Six pg of RT products were used in PCR amplification in a final 50pl solution con- taining 2.5 mM dNTP, 25 mM MgC12, upstreamldownstream primers and Taq DNA polymerase (Invitrogen life technologies). Following an initial denaturation at 95°C for 5min, the DNA was amplified in the Touchgene Gradient (TECHINE, UK). Thirty-five cycles of 1 min at 94°C were applied for denaturation and 1 min at 72°C for extension. A final extension was applied at 72°C for 5min. PCR prod- ucts were visualized on 1% agarose gel (Agarose I, AMRESCO, Ohio) and stained with EtBr. PCR products were confirmed by size verifica- tion and analyzed using FloGel-I (Fluorescent Gel Image System, TOPBIO). Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was used as an internal control. Primer sets were following:
( I )
Sox9-5’-GGCAGCTGTGAACTGGCCA-3’
(sense primer) and 5’- GCACACGGGGAACTTGTCC-3’ (antisense primer) which gives a 408 bp product;(2) COL2A 1 -5’-CACGCAGAAGTTCACCAAGAA-3‘ (sense primer) and 5’-CTGCAGGATCAGCCATGGTAGA-3’ (antisense primer) which gives a 501 bp products;
(3)Aggracan-5‘-TGAGGAGTGGAACAAGTACC-3’(sense
pri-mer) and S’-GGAGGTGGTAAlTGCAGGGAACA-3’ (antisense
primer) which gives 466 bp product;
(4) GADPH-5’-GCTCTCCAGAACATCATCCCTGCC-3‘ (sense pri-
mer) and 5’-CGTTGTCATACCAGGAAATGAGCTT-3‘ (anti-
sense primer) which gives 346 bp product. Preparation of type I and II collagens
Type I and I1 collagens were prepared as previously described in this laboratory [20,21]. Collagen was heat-denatured at 95°C for 30 min, and analyzed using SDS-PAGE. After breaking the peptide chain of a-subunits, the triple-helical structure of collagen was denatured.
Analysis of chondrogenic differentiation in monolayer culture
Rabbit and human MPCs were seeded into six-well plates with 10% FBS and 50 pg/ml ascorbic acid in DMEMILG. After four days of cul- tivation, the medium of rabbit MPCs was changed with dexametha- sone (10-7M, Sigma D2915, St. Louis), rhTGF-PI (0.1, I, and IOngl ml, R & D Systems 240-B, Minneapolis, MN), and collagen (10, and 100 pg/ml). Six wells were employed for each condition. Differentiation was analyzed by GAG synthesis and ALP activity after 14 days. GAG synthesis and ALP activity were measured using spectrophotometry of the absorption of O D m and OD405 respectively after alcian-blue stain- ing [l]. The medium of human MPCs, after four days of cultivation, was changed with dexamethasone (10-7M), rhTGF-PI (1 ng/ml), and collagen (100pglml). Three wells were employed for each condition. The mRNA for aggrecan, type I1 collagen, and sox9 was analyzed using RT-PCR as described above.
Structural effects of collagen on chondrogenic diflerentiation by MPCs
To further identify the regulatory effect of collagen’s structure on chondrogenic differentiation, native and denatured collagens were tested in cell cultivation. Each collagen solution (2mglml) was heated at 95°C for 30min to denature the peptide chains of the collagen subunits. Degradation of collagen and the triple-helical structure was analyzed using 5% SDS-PAGE. Native or denatured collagen was individually added to the medium at a concentration of IOOpglml each. The expres- sion of GAG by MPCs in each condition was measured by alcian-blue assay.
Statistical analysis
Expressions of each GAG and ALP from chondrocyte cultures ex- posed to different concentrations of dexamethasone, rhTGF-PI, and collagens were analyzed using Mann-Whitney test. Dexamethasone-,
448 C. W. Clien et al. I Journul of Orthopaedic Research 23 (2005) 446-453 rhTGF-P1-1, and collagen-treated and untreated control cultures at
different concentrations were also compared. Data are reported as the mean f. SD.
P
< 0.05 was considered statistically significant.Results
Type I and II collagen regulated rhTGF-PI and
dexamethasone effects on GAG expression of rabbit
MPCs
Significantly increased GAG expression was found in
the culture supplemented with 10 ng/ml rhTGF-01.
GAG expression was 2.6 k
0.2
times greater than those
of
untreated cells. However, GAG synthesis did not in-
crease with 0.1 and 1 ng/ml concentrations of rhTGF-PI.
Treatment with lop7 M dexamethasone alone increased
GAG expression 2.25
f
0.18 times in rabbit MPCs com-
pared to the untreated group. The increased rate of
GAG expression remained the same as in the untreated
group after adding 0.1 and 1 ng/ml concentrations of
rhTGF-Pl.
A
synergistic effect was found when incor-
porated with both 10ng/ml rhTGF-01 and lop7 M dexa-
methasone. The maximal GAG expression was
3.75
f
0.32
times higher than that
of
control cells (Fig.
1 (A)).
Type
I1
collagen alone at the concentration of 10
pg/ml did not significantly change GAG expression,
whereas at the concentration
of
100
pg/ml, GAG expres-
sion increased 2.5 times. Synergistic increases in GAG
expression by MPCs to be up to 5.8 times when adding
lOOpg/ml type I1 collagen (Fig. l(B)). When adding
either 10pg/ml or lOOpg/ml concentration of type I1
collagen (10 or lOOpg/ml), the osteogenic marker ALP
activity of MPCs did not significantly increase. In-
creases in ALP activity were only 1.2 and 1.4 times,
respectively, higher than the control. With treatment
of
10ng/ml TGF-01 and 10p7M dexamethasone, ALP
expression increased 2.4 times. Further addition of type
TGF-beta 1 (nglml) Type I I Collagen (microgramlml)
(A) GAG, glycosaminoglycan; DEX, dexamethasone; TGF, hTGF-pl (B) GAG, glycosaminoglycan; DEX, dexamethasone; TGF, rhTGF-pl
(c)
ALP, alkaline phosphatase; DEX, dexamethasone; TGF, rhTGF-p1 (D) GAG, glycosaminoglycanFig. I. GAG and ALP expression of MPCs exposed to varying concentrations of rhTGF-PI, DEX, and collagen, measured by spectrophotometry. Means f S.D; (n = 6). (A) TGF-PI and dexamethasone induced an increase in GAG by MPCs. An asterisk indicates a significant (p < 0.05) difference in DEX compared to the untreated control. A diamond indicates a significant (p < 0.05) difference among different TGF-PI concentrations. (B) Type I1 collagen up regulated GAG synthesis with and without dexamethasone. An asterisk indicates a significant (p < 0.05) difference in DEX compared to the Untreated control. A diamond indicates a significant (p < 0.05) difference among different type I1 collagen concentrations. (C) Type I1 collagen, in contrast to dexamethasone, did not significantly increase ALP activity by mesenchymdl progenitor cells. An asterisk indicates a significant
(p < 0.05) difference in TGF-PI and DEX compared to the untreated control. (D) Only type I1 collagen significantly increased GAG synthesis, but not cell proliferation, whereas type I collagen did not change GAG synthesis or cell proliferation. An asterisk indicates a significant (p < 0.05) difference in type I1 collagen compared to type I collagen and the untreated control.
C. W. Chen et al. I Journal of Orthopaedic Research 23 (2005) 446453 449
I1
collagen ALP caused the same increased expres-
sion of 2.4 times (at 10pg/ml concentration) and 2.7
times (at 100 pg/ml concentration). Thus, type I1 colla-
gen did not significantly regulate the ALP activity of
MPCs with
or
without TGF-P1 and dexamethasone
(Fig. l(C)).
Incorporated with lOOpg/ml
of
type I1 collagen, the
population of MPCs was reduced by 40%. However,
GAG expression increased 2.5 times. In contrast, with
the addition of 100 pg/ml of type I collagen, the popula-
tion and GAG expression of MPCs appeared to not sig-
nificantly change (Fig. l(D)).
Type
I
and II collagen regulated rhTGF-Bl und
dexamethasone effects on aggrecan,
Sox9
and collagen
mRNA expression
RT-PCR using RNA obtained from human MPCs
showed no signal for aggrecan, type
I1
collagen, and
Sox9. An mRNA expression
of
type
I1
collagen and
Sox9 was induced when treated with dexamethasone.
With dexamethasone and rhTGF-P 1 MPCs increased
mRNA expression of type
I1
collagen and Sox9. In addi-
tion to dexamethasone and rhTGF-Pl, type
I1
collagen
induced mRNA expression
of
aggrecan and enhanced
(A)
GAG, glycosaminoglycan;
DEX,
dexamethasone;
TGF, rhTGF-Pl
(6)
Fig. 2. Structural effects of collagen on chondrogenic differentiation by MPCs. (A) Native type I1 collagen remarkably increased 4.3 times GAG synthesis, the increase of GAG was reduced to 2.9 times with the denatured type I1 collagen. Noted no significant difference of GAG synthesis among groups of native type I collagen, denatured type I and I1 collagens, and dexamethasone and rhTGF-PI treated. (B) SDS-polyacrylamide gel electrophoresis of native and denatured collagens. Note the clearly identifiable bands of type I native collagen in lane 1 and of type I1 collagen in lane 3. The denatured type I collagen showed a smear phenomenon in lane 2, and small fragments of denatured type I1 collagen was showed in lane 4.
450 C. W. Chen et al. I Journal of Orihopaedic Research 23 (2005) 446453
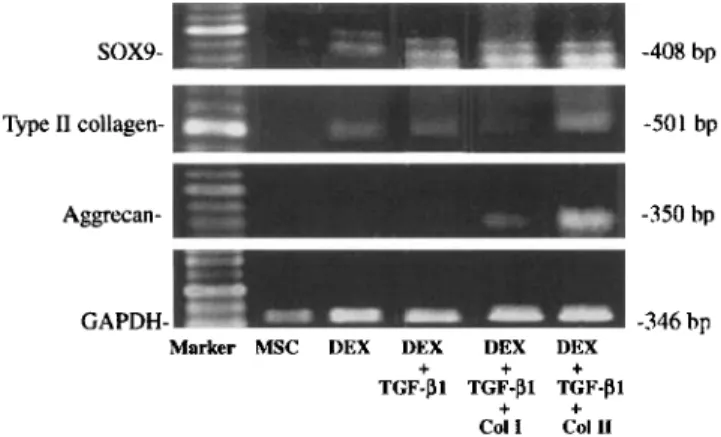
SOX9- -408 bp
Type I1 collagen- -501 bp
Aggrecan- -350 bp
GAPDH- -346 bp
Marker MSC DEX DEX VEX DEX
+
+ ++ +
TGF-PI TGF-PI TGF-PI
C O l 1 Call1
Fig. 3. Type I and I1 collagens regulated rhTGF-BI and dexameth- asone effects on aggrecan, sox9 and collagen mRNA expression. Total RNA was isolated from MPCs and used for PT-PCR to detect expression of type 11 collagen and aggregan. Expression of GADPH was used as control. Lane 1: DNA standards; lane 2: MPCs showed no signal for aggrecan, type I1 collagen, and sox9; lane 3: With dexamethasone MPCs induced mRNA expression of type I1 collagen and sox9; lane 4 With dexamethasone and rhTGF-01 MPCs increased mRNA expression of type I1 collagen and sox9; lane 5: Cultured with type I collagen, dexamethasone and rhTGF-PI MPCs showed no type I1 collagen mRNA; lane 6: Cultured with type I1 collagen. dexameth- asone and rhTGF-01 MPCs induced mRNA expression of aggrecan and enhanced levels of sox9 and type I1 mRNA.
levels of Sox9 mRNA. In contrast, incorporation with
type
I
collagen, dexamethasone and rhTGF-Pl MPCs
reduced levels of aggrecan, and Sox9 mRNA, showed
no type I1 collagen mRNA (Fig. 3).
Structural efsrcts of collagen on chondrogenic
differentiation induced by MPCs
To determine whether the structure of collagen affects
chondrogenic differentiation, native and denatured col-
lagens were introduced to MPCs. With pretreatment
using rhTGF-PI and dexamethasone, MPCs increased
GAG synthesis 4.3 times after adding native type
I1
col-
lagen. The enhancement of GAG synthesis was down-
graded to 2.9-fold increase, when denatured type
I1
collagen was added instead.
In
contrast, no significant
difference was noted between the increase in GAG syn-
thesis in the presence of native (2.5 times) or denatured
type I collagen (2.1 times) (Fig. 2(A)). Intact al(I), a2(I),
and al(I1) bands were clearly identified with native type
1 and I1 collagens. Contrarily, a smear phenomenon was
noted with denatured type I and I1 collagens (Fig. 2(B)).
These results indicate that type I1 collagen-triggered
chondrogenic differentiation is type-specific, and corre-
lates with collagen’s native triple-helical structure.
Type 11 and I collagen efects on morphological changes
in MPCs treated with dexamethasone and rh TGF-Pl
in rnonolayer cultures
MPC cultures of
all
six wells showed a phenomenon
of contact inhibition with fibroblast-like morphology
after 14 days (Fig. 4(A)). A mild pile-up was noted
in MPCs of all six wells when supplemented with
rhTGF-1 and dexamethasone (Fig. 4(D)). In all cultures
incorporating type I collagen, cells showed a shift to
elongated spindle-shaped fibroblastic appearance inde-
pendent of the absence or presence of
TGF-(31
and dexa-
methasone (Fig. 4(B) and (E)).
Incorporated with type I1 collagen, MPCs of all six
wells showed more cuboidal-, and less spindle-shaped
morphology (Fig. 4(C)). Semi-transparent fibrous like
substance was noted on cells. Cells predominantly ap-
peared rectangular after pretreatment with type
I1
colla-
gen, dexamethansone and rhTGF-(3 1. Cell-collagen
matrix aggregates were found in four of six wells (Fig.
4(F)). Abundant GAG, which found in the cartilage-like
cell-collagen matrix aggregate was then identified by
alcian-blue staining (Fig. 4(G)).
Discussion
Bone marrow-derived MPCs are highly proliferative,
multipotential cells that have been considered ideal cells
for use in repair of injured cartilage and fractures of
bone. It is known that repair tissue arises from differen-
tiation of local MPCs. Both periosteum and bone mar-
row contain these cells, which preserve the ability to
differentiate into both chondrocytes and osteoblasts
[ 11,24,28]. Chondrogenic differentiation can be trig-
gered, if the environmental factors such as ECM and
local cytokines are facilitative. Local MPCs enable
accumulation, proliferation, and terminal differentiation
into hypertrophic chondrocytes. If environmental fac-
tors are not facilitative; however, local MPCs differenti-
ate into fibrochondrocytes and form fibrocartilage
[9,381.
In addition to the potential for multidifferentiation,
MPCs are relatively easy to extract from bone marrow
and expand in culture. Development of an in vitro chon-
drogenic differentiation model of marrow-derived MPCs
presents an opportunity to explore the extracellular ma-
trix’s guiding effects on chondrogenesis that is of poten-
tial therapeutic utility.
Previous study showed that TGF-(31 induces prolifer-
ation of osteoblasts, chondrocytes, and mesenchymal
stem cells [40]. The present study also showed an
increase in the proliferation of mesenchymal progeni-
tors. Additionally, other study have shown that syn-
thesis of ECM was found to be enhanced by TGF-PI
[39]. Similarily, this study also showed a consistency
in this regard, in that TGF-PI increased synthesis of
GAG in MPCs.
TGF-PI controls the morphology
and differentiation of epithelial cells [47, 3,301. The
morphological changes in MPCs induced by TGF-P1
in this research were elongation and extension of cell
shapes.
C. W. Chen et al. I Journul yf Ortliopardic Research 23 (2005) 446453 45 1
Fig. 4. Type I and I1 collagen effects on morphological changes in MPCs treated with dexamethasone and rhTGF-PI in monolayer culture ( 1 0 0 ~ ) . (A and D) MPCs appeared fibroblast-like morphology and mild pile-up 14 days after cultured with dexamethasone and TGF-PI. Incorporation with extracellular type I collagen cells showed an elongated spindle-shaped fibroblastic appearance in the absence (B) or presence (E) of rhTGF-PI and dexamethasone. (C) Cells showed more cuboidal-, and less spindle-shaped morphology after pretreatment with type I1 collagen. (F) Cells appeared predominantly rectangular after pretreated with type I1 collagen in addition to dexamethansone, and rhTGF-PI. (G) Note abundant G A G in the cartilaginous-like cell-collagen matrix aggregate (alcian-blue staining).
Chondrogenesis of chondroprogenitors can be stim-
TGF-P1 further enhances the process
of
the synthesis
ulated in the presence of the dexamethasone [12].
of extracellular matrix. Levels of mRNA for type
I1
Chondrogenesis was induced by dexamethasone. That
collagen and Sox9 were also increased when treated
was revealed by increased GAG expression. There
with dexamethasone in this study. This was correlated
was a synergistic effect with TGF-P1.
This
study imply
with previous study that an enhancement of
Sox
that dexamethasone may be the fundamental factor
mRNA by dexamethasone was showed in chondrocytes
that triggers chondrogenic differentiation, whereas
[36].
452 C.
W.
Chen et al. I Journal of Orthopaedic Research 23 (2005) 446453The regulatory effect of collagen seems to be corre-
lated with its triple-helical structure. Our data showed
that enhancement of
GAG
synthesis by native type
I1
collagen (4.3 times) was significantly downgraded when
using denatured type
I1
collagen (2.9 times). These find-
ings agree with previous studies that denatured type
I1
collagen dimilished native type
I1
collagen effect on
chondrocyte regulation [33,4 1,421.
Exogenous type I1 collagen could maintain the phe-
notype of chondrocytes. This increased the syntheses
of type I1 collagen and
GAG
[37,32,33]. Cell-matrix
interactions via cell surface receptors transduce extracel-
lular signals inwards to regulate the cell phenotype [34].
In this research by adding type
I1
collagen the semi-
transparent fibrous-like substance became noted on
MPCs. This indicated the type
I1
collagen receptor
might be induced on cell surfaces. Integrin a2p1, a major
receptor for type
I1
collagen, plays important roles
during chondrogenic differentiation by MPCs [ 17,231.
Binding affinity and signaling of type
I1
collagen are
mediated by integrin receptor [35]; however, the signal-
ing of specific domains of
a
or
p
integrins for type 11 col-
lagen is not well understood. Further study will identify
the specific receptor for type
I1
collagen, their down-
stream signals, and how it regulates chondrogenic differ-
entiation of MPCs.
Type I collagen, in contrast to cartilage-specific type
I1
collagen, exists ubiquitously in bone, tendon, cornea,
and skin and acts as a structural protein in mammals
[22]. During cartilage repair of large osteochondral de-
fects, bone marrow-derived MPCs are induced to differ-
entiate into fibroblasts or osteoblasts within fibrin clots
containing fibronectin and type
I
collagen [2,38]. Consis-
tent with previous research, type
I
collagen treated
MPCs did not increase GAG expression, but induced
an elongated spindle-shaped fibroblastic appearance.
Therefore, type
I
collagen matrix may provide the suit-
able microenvironment, which is better for fibrogenic
differentiation. This matrix-guided mesenchymal pro-
genitor cell differentiation
in
situ predominantly induces
regeneration towards fibro-cartilage formation.
The current study substantiates the concept that
chondrogenic differentiation by MPCs can be synergisti-
cally triggered by cytokines and further regulated by the
ECM.
A
comprehensive understanding of the regulation
of chondrogenic differentiation by MPCs may enable the
intentionally engineering of cartilage development
in vitro. Subsequent results may yield further informa-
tion for the purpose of a direct repair of large cartilage
defects.
Acknowledgments
This work was supported by the National Science
Council, Taiwan, R.O.C. under project no. NSC91-
2314-B-038-033. The authors thank Prof C.Y. Yeh for
statistical advice, and Ching-Jin Tu and Ching-Fu Liao
for their technical help with histological preparations.
References
[I] Bjornsson S. Simultaneous preparation and quantitation of proteoglycans by precipitation with alcian blue. Anal Biochem
1993;210:282-91.
[2] Blair HC, Zaidi M, Schlesinger PH. Mechanisms balancing skeletal matrix synthesis and degradation. Biochem J 2002; 36432941.
[3] Boland S, Boisvieux-Ulrich E, Houcine 0, Baeza-Squiban A, Pouchelet M, Schoevaert D, et al. TGF beta 1 promotes actin cytoskeleton reorganization and migratory phenotype in epithelial tracheal cells in primary culture. J Cell Sci 1996;109(Pt 9):2207-19. [4] Bouwmeester PS, Kuijer R, Homminga GN, Bulstra SK, Geesink
RG. A retrospective analysis of two independent prospective cartilage repair studies: autogenous perichondrial grafting versus subchondral drilling 10 years post-surgery. J Orthop Res 2002; 20:267-73.
[5] Brittberg M, Faxen E, Peterson L. Carbon fiber scaffolds in the treatment of early knee osteoarthritis. A prospective 4-year followup of 37 patients. Clin Orthop 1994:155-64.
[6] Brittberg M, Lindahl A, Nilsson A, Ohlsson C, Isaksson 0, Peterson L. Treatment of deep cartilage defects in the knee with autologous chondrocyte transplantation. N Engl J Med 1994;33 1:88%95.
[7] Bruder SP, Kurth AA, Shea M, Hayes WC. Jaiswal N, Kadiyala S. Bone regeneration by implantation of purified, culture- expanded human mesenchymal stem cells. J Orthop Res 1998;
16: 15542.
[8] Buckwalter JA. Articular cartilage injuries. Clin Orthop 2002: [9] Buckwalter JA, Mankin HJ. Articular cartilage: degeneration and osteoarthritis, repair, regeneration, and transplantation. Instr Course Lect 1998;47:487-504.
[lo] Buckwalter JA, Mankin HJ. Articular cartilage 11: degeneration and osteoarthrosis, repair, regeneration and transplantation. J Bone Joint Surg Am 1997;79:612-32.
[I I] Goshima J, Goldberg VM, Caplan AI. The origin of bone formed in composite grafts of porous calcium phosphate ceramic loaded with marrow cells. Clin Orthop 1991:27483.
[I21 Grigoriadis AE, Heersche JN, Aubin JE. Analysis of chondro- progenitor frequency and cartilage differentiation in a novel family of clonal chondrogenic rat cell lines. Differentiation 1996; 60:299-307.
131 Hiraki Y, Shukunami C, Iyama K, Mizuta H. Differentiation of chondrogenic precursor cells during the regeneration of articular cartilage. OA & Cartilage 2001;9(Suppl A):S102-8.
141 Hoikka VE, Jaroma HJ, Ritsila VA. Reconstruction of the patellar articulation with periosteal grafts, 4-year follow-up of 13 cases. Acta Orthop Scand 199061:369.
151 Homminga GN, Bulstra SK. Bouwmeester PS, van der Linden AJ. Perichondral grafting for cartilage lesions of the knee. J Bone Joint Surg Br 1990;72:1003-7.
[16] Hunt SA, Jazrawi LM, Sherman OH. Arthroscopic management of osteoarthritis of the knee. J Am Acad Orthop Surg 2002;
10:35&63.
[I71 Hynes RO. Integrins: versatility, modulation, and signaling in cell adhesion. Cell 1992;69:11-2.
[I81 Ikeuchi M, Dohi Y, Horiuchi K, Ohgush iH, Noshi T, Yoshikawa T, et al. Recombinant human bone morphogenetic protein-2 promotes osteogenesis within atelopeptide type 1 collagen solution 21-37.
C. W. Chen et al. I Journal of Orthopaedic Reseurch 23 (2005) 4 4 6 4 5 3 453 by combination with rat cultured marrow cells. J Biomed Mater
Res 2002;60:61-9.
[I91 Johnstone B, Hering T, Caplan A, Goldberg V, J U V. In vitro chondrogenesis of bone marrow-derived mesenchymal progenitor cells. Exp Cell Res 1998;238:265-72.
[20] Lai WF, Tsai YH, Chan WP, Yang WC, Pham W. New bone implant material. US Patent 2003;0045942Al.
[21] Lai WF, Tang JR, Chen CT. Fabrication of a cartilage implant. US Patent 2003;0152556Al.
[22] Linsenmayer T. Collagen. New York: Plenum Press: 1991. pp. 45-78.
[23] Loeser RFS S, Tan L, Goldring MB. Integrin expression by primary and immortalized human chondrocytes: evidence of a differential role for albland a2bl integrins in mediating chon- drocyte adhesion to type I1 and
VI collagen. OA & Cartilage
2000;8:96 105.[24] Maniatopoulos CSJ, Melcher AH. Bone formation in vitro by stromal cells obtained from bone marrow of young adult rats. Cell Tissue Res 1988;254317-30.
[25] Messner K. Lohmander LS, Gillquist J. Neocartilage after artificial cartilage repair in the rabbit: histology and proteoglycan fragments in joint fluid. J Biomed Mater Res 1993;27:949-54. [26] Minas T, Nehrer S. Current concepts in the treatment of articular
cartilage defects. Orthopedics 1997;20:525-38.
[27] Mizuta H, Sanyal A, Fukumoto T, Fitzsimmons J, Matsui N, Bolander M, et al. The spatiotemporal expression of TGF-beta1 and its receptors during periosteal chondrogenesis in vitro. J Orthop Res 2002;20:562-74.
[28] Nakahara H, Bruder SP, Haynesworth SE, Holecek JJ, Baber MA, Goldberg VM, et al. Bone and cartilage formation in diffusion chambers by subcultured cells derived from the perios- teum. Bone 1990;11:181-8.
[29] Nicoll S, Wedrychowska A, Smith N, Bhatnagar R. Modulation of proteoglycan and collagen profiles in human dermal fibroblasts by high density micromass culture and treatment with lactic acid suggests change to a chondrogenic phenotype. Connect Tissue Res 200 1;42:5949.
[30] Petersen B, Yee CJ, Bowen W, Zamegar R, Michalopoulos GK. Distinct morphological and mito-inhibitory effects induced by TGF-beta 1, H G F and EGF on mouse, rat and human hepato- cytes. Cell Biol Toxicol 1994;10:219-30.
[3 I] Peterson L. Articular cartilage injuries treated with autologous chondrocyte transplantation in the human knee. Acta Orthop Belg 1996;62(Suppl 1):19&200.
[32] Qi WN, Scully SP. Effect of type I1 collagen in chondrocyte response to TGF-beta 1 regulation. Exp Cell Res 1998;241: 142-50.
[33] Qi WN, Scully SP. Extracellular collagen modulates the regulation of chondrocytes by transforming growth factor-beta 1. J Orthop Res 1997;15:483-90.
[34] Saoncella S, Echtermeyer F, Denhez F, Nowlen JK, Mosher DF, Robinson SD, et al. Syndecan-4 signals cooperatively with
integrins in a Rho-dependent manner in the assembly of focal adhesions and actin stress fibers. Proc Natl Acad Sci USA 1999;96:2805-10.
[35] Scully SPL
JW,
Ghert PMA, Qi W. The role of the extracellular matrix in articular chondrocyte regulation. Clin Orthop [36] Sekiya I, Koopman P, Tsuji K, Mertin S, Harley V, YamadaY,
et al. Dexamethasone enhance SOX9 expression in chondrocytes. J Endocrinol2001:169:573-9.
[37] Shakibaei M, De Souza P, Merker HJ. Integrin expression and collagen type I1 implicated in maintenance of chondrocyte shape in monolayer culture: an immunomorphological study. Cell Biol Int 1997;21: 115-25.
[38] Shapiro F, Koide S , Glimcher MJ. Cell origin and differentiation in the repair of full-thickness defects of articular cartilage. J Bone Joint Surg Am 1993;75:532-53.
[39] Silberstein GB, Strickland P, Coleman S, Daniel CW. Epithelium- dependent extracellular matrix synthesis in transforming growth factor-beta 1-growth-inhibited mouse mammary gland. J Cell Biol 1990; 1 10:2209- 19.
[40] Spom MB, Roberts AB, Wakefield LM, Assoian RK. Trans- forming growth factor-beta biological function and chemical structure. Science 1986;233:5324.
[4 I] Stoop R, Buma P, van der Kraan PM, Hollander AP, Billinghurst RC, Meijers TH, et al. Type I I collagen degradation in articular cartilage fibrillation after anterior cruciate ligament transection in rats. OA & Cartilage 2001;9:308-15.
[42] Tuckwell DS, Ayad S, Grant ME, Takigawa M, Humphries MJ. Conformation dependence of integrin-type I1 collagen binding. Inability of collagen peptides to support alpha 2 beta 1 binding, and mediation of adhesion to denatured collagen by a novel alpha 5 beta I-fibronectin bridge. J Cell Sci 1994;lO7(4):993-l005. [43] Wakitani S, Goto T, Pineda SJ, Young RG, Mansour JM, Caplan
AI, et al. Mesenchymal cell-based repair of large, full-thickness defects of articular cartilage. J Bone Joint Surg Am 199476: 579-92.
[44] Werntz JR, Lane JM, Burstein AH, Justin R, Klein R, Tomin E. Qualitative and quantitative analysis of orthopedic bone rege- neration by marrow. J Orthop Res 1996;14:85-93.
[45] Worster A, Brower-Toland B, Fortier L, Bent S, Williams J, Nixon A. Chondrocytic differentiation of mesenchymal stem cells sequentially exposed to transforming growth factor-beta 1 in monolayer and insulin-like growth factor-I in a three-dimensional matrix. J Orthop Res 2001;19:73849.
[46] Yo0 J, Barthel T, Nishimura K, Solchaga L, Caplan A, Goldberg V, et al. The chondrogenic potential of human bone-marrow- derived mesenchymal progenitor cells. J Bone Joint Surg Am 1998;801745-57.
[47] Zavadil J, Bitzer M, Liang D, Yang YC, Massimi A, Kneitz S, et al. Genetic programs of epithelial cell plasticity directed by transforming growth factor-beta. Proc Natl Acad Sci USA 2001;98:668691.