國立交通大學
生化工程研究所
博 士 論 文
決定牛乳
β-lactoglobulin-vitamin D complex 之結晶結構並利用胺
基酸定點突變及生物資訊探討其生理與物理生化功能
Physiological and physico-biochemical function of bovine milk
β-lactoglobulin as probed by site-directed mutagenesis, bioinformatics
and its crystal structure complexed with vitamin D
研 究 生: 楊明誌
指導教授: 毛仁淡 講座教授
決定牛乳
β-lactoglobulin-vitamin D complex 之結晶結構並利用胺
基酸定點突變及生物資訊探討其生理與物理生化功能
Physiological and physico-biochemical function of bovine milk
β-lactoglobulin as probed by site-directed mutagenesis, bioinformatics
and its crystal structure complexed with vitamin D
研 究 生:楊明誌 Student:Ming Chi Yang
指導教授:毛仁淡 講座教授 Advisor:Chair Prof. Simon JT Mao
國 立 交 通 大 學
生 化 工 程 研 究 所
博 士 論 文
A Thesis
Submitted to Institute of Biochemical Engineering College of Biological Science and Technology
National Chiao Tung University in partial Fulfillment of the Requirements
for the Degree of PhD in
Biological Science and Technology April 2009
決定牛乳
β-lactoglobulin-vitamin D complex 之結晶結構並利用胺
基酸定點突變及生物資訊探討其生理與物理生化功能
研究生:楊明誌 指導教授:毛仁淡 講座教授
國立交通大學 生化工程研究所 博士班
摘要
β-Lactoglobulin (LG)為牛乳中主要乳清蛋白之一,calyx 公認為維生素
D 的主要結合區,而第二個維生素 D 結合區之存在仍受到許多爭論。在本
研究中,利用螢光光譜分析,得知維生素
D 與 LG 結合比例為 2,表示 LG
可能具有兩個維生素
D 結合區。藉由降低 pH 值及加熱破壞 calyx 結合疏
水性分子之能力,LG 仍具有結合維生素 D 之能力,其結合比例為 1,此
結果證實
LG 除 calyx 以外具有第二個維生素 D 結合區。運用生物資訊程
式(Insight II、Q-SiteFinder、GEMDOCK)發現有兩個區域可能為第二個維
生素
D 結合區,且 GEMDOCK 幫助我們在具有潛力的第二維生素 D 結
合區上尋找額外電子雲密度。藉由改變製備共同結晶的
pH 值及起始維生
素
D/LG 之比率來優化 ligand 佔有率及增加複合體中維生素 D 的電子雲密
度。在
pH 8 及起始維生素 D/LG 之比率為 3 的條件下,製備 LG 與維生
素
D 複合體(complex)結晶,進行同步輻射光束繞射確切得知 LG 上第二個
維生素
D 結合區所在區域。第二維生素 D 結合區位於 LG 分子之 C 端表
面,此結合區具有約
17.91 Å 之長度,而維生素 D 分子約為 12.51 Å,代
表此結合區足以容納維生素
D 分子。第二維生素 D 結合區由α-helix 提供
非極性胺基酸
Phe136、Ala139、Leu140 與β-strand I 提供非極性胺基酸
Ile147,經由疏水性 loop (Ala142、Leu143、Pro144、Met145)連接形成一
個疏水性結合區,促進維生素
D 結合。另一方面,利用定點突變進行系
統化分析發現γ-trun loop 上 Leu143、Pro144、Met145 為 LG 第二維生素 D
結合區上參與結合之重要胺基酸。更進一步支持證據為維生素
D 抑制專
一性辨識γ-trun loop 之單株抗體的免疫反應。眾所皆知,LG 具有中心 calyx
及第二維生素
D 結合區,然而生理上 LG 能攜帶維生素 D 通過腸胃道,
增加維生素
D 吸收之能力至今尚未證實。利用小鼠作為動物模式,起初
證實
LG 為牛乳中攜帶維生素 D 及增加維生素 D 吸收之主要蛋白質。深
入探討加熱對於
LG 增加維生素 D 吸收之影響,發現 LG 具有兩個維生素
D 結合區,餵食老鼠添加維生素 D 之 LG,其血液中維生素 D 濃度於維生
素
D/LG 比率 2 時達到飽和,而 heated LG 只具有第二維生素結合區,餵
食老鼠添加維生素
D 之 heated LG,其血液中維生素 D 濃度於維生素 D/LG
比率
1 時就已達到飽和。因 LG 第二維生素結合區不受加熱破壞,其具有
攜帶維生素
D 及增加維生素 D 吸收之優勢。本研究總結發現於牛乳中添
加維生素
D 將能有效增加維生素 D 為人體吸收。
Physiological and physico-biochemical function of bovine milk
β-lactoglobulin as probed by site-directed mutagenesis,
bioinformatics and its crystal structure complexed with vitamin D
Student::Ming Chi Yang Advisor: Chair Prof. Simon JT Mao
Institute of Biochemical Engineering National Chiao Tung University
Abstract
β-Lactoglobulin (LG) is a major milk whey protein containing primarily a calyx for vitamin D3 binding, although the existence of another site beyond the calyx is controversial.
In this study, using fluorescence spectral analyses, we showed the binding ratio for vitamin D3
to LG to be 2:1 and a ratio of 1:1 when the calyx was “disrupted” by manipulating the pH and temperature, suggesting that a secondary vitamin D binding site existed. The bioinformatic programs (Insight II, Q-SiteFinder, and GEMDOCK) identified the two potential regions for this secondary vitamin D binding site. It was concluded that GEMDOCK can aid in searching for an extra density map around potential vitamin D binding sites. We then optimized the occupancy and enhanced the electron density of vitamin D3 in the complex by
altering the pH and initial ratios of vitamin D3/LG in the cocrystal preparation. Finally, we
identified the secondary site (defined as the exosite) for vitamin D binding using a crystal prepared at pH 8 with a vitamin D3/LG ratio of 3:1. The exosite, however, is near the
with 17.91 Å in length, while the span of vitamin D3 is about 12.51 Å. A remarkable feature
of the exosite is that it combines amphipathic α-helix providing nonpolar residues (Phe136, Ala139, and Leu140) and β-strand I providing a nonpolar (Ile147), which are linked by a hydrophobic loop (Ala142, Leu143, Pro144, and Met145). Thus, the binding pocket of the exosite furnishes strong hydrophobic force to stabilize vitamin D3 binding. On the other
hand, using site-directed mutagenesis, we demonstrate that residues Leu143, Pro144 and Met145 in the γ-turn loop play a crucial role in the binding. Further evidence is provided by the ability of vitamin D3 to block the binding of a specific mAb in the γ-turn loop. LG
contains a central calyx and a second exosite beyond the calyx to bind vitamin D; however, the biological function of LG in transporting vitamin D remains elusive. Using the mouse (n=95) as an animal model, we initially demonstrated that LG is a major fraction of milk proteins responsible for uptake of vitamin D. Most interestingly, dosing mice with LG supplemented with vitamin D3 revealed that native LG containing two binding sites gave a
saturated concentration of plasma 25-hydroxyvitamin D at a dose ratio of 2:1 (vitamin D3/LG),
whereas heated LG containing one exosite (lacking a central calyx) gave a ratio of 1:1. We have demonstrated for the first timethat the exosite of LG has a functional advantage in the transport of vitamin D, indicating that supplementing milk with vitamin D effectively enhances its uptake.
誌 謝
在經過五年努力終於完成此博士論文,這個帶著苦楚、沮喪、歡欣、鼓
舞等喜怒哀樂情緒夾雜的博士班求學過程,終於要告一段落了。在途中,
曾因磨難而對於研究生涯產生困惑感,特別要感謝指導教授 毛仁淡講座教
授,給予鼓勵與包容。毛講座的諄諄教誨及日夜叮嚀,在生活、實驗、論
文撰寫上給予學生悉心地指導,明誌都將謹記在心,並獻上最誠摯地感謝。
在博士研究中,誠摯地感謝同步輻射中心生命科學組 陳俊榮教授引領
我一窺蛋白質晶體結構的奧秘,不厭其煩教導我這個門外漢,給予支持令
我由衷感激。與此同時,感謝生物資訊所 楊進木教授引領我進入生物資物
領域,不時地討論以化解彼此研究領域上的鴻溝,常常激發出非常驚奇、
偉大之想法。此論文之英文文法上承蒙核能研究所 劉明毅教授不吝斧正,
使學生許多期刊論文得以發表。除此以外,感謝中央研究院分子生物所 蕭
傳鐙教授於口試及論文撰寫上的建議與指正,許多寶貴意見使我獲益良
多。另感謝清華大學生物資訊與結構生物所 管泓翔學長在蛋白質晶體結構
修正上給予許多協助。同時感謝生物資訊所 葛振寧同學在生物資訊預測方
面給予許多幫忙。
這些日子以來,實驗室如同是我的家,學長姊、同學及學弟妹陪我度過
這酸甜苦辣的五年光陰。首先感謝文亮學長製作許多單株抗體供我實驗上
所需、俊瑩學長與我高談闊論許多偉大的研究方向及學弟妹乃齊、沁紜、
佶穎實驗上的幫忙。另感謝財木、文亮、繼鋒、以祥、怡荏、彰威、宏輝、
俊瑩、珮真等學長姊在我沮喪時給予鼓勵及幫忙,文昭、立品同學在生活
及實驗上的同甘共苦,靜怡、善琦小秘書給予許多的協助,中曦、冠佑、
乃齊、小涵、威延、佩宇、詩璇、佶穎、巾涵、沁紜、俐穎、柏如、宛伶、
伊玟等學弟妹帶來眾多的歡樂。在未來的日子裡,回味起這段充滿喜怒哀
樂的記憶時,仍是充滿感激。
最後,最必須感謝的是雙親(金圳及玉美)及知己朋友(志宏、俊杰、
逸宏、老顧、韶康、詩玉、孟璇、筱婷、小寶貝),謝謝你們在背後默默
地支持我,當我的避風港。每當遇到困難及鬱悶時,替我排憂解難,讓我
不斷地鼓起勇氣,去克服接踵而來的挑戰,如果沒有你們,我將無法順利
完成博士學位。前方的道路雖有荊棘、磨難,有你們的陪伴將使我更有毅
力往前邁進。謹以此論文獻給你們,此為我們共同的榮耀。
Dissertation index
Abstract in Chinese ---I Abstract in English---III Acknowledgements--- V Index --- VII
Overview --- 1
Overall experimental design--- 8
References --- 9
Section 1: The existence and location of secondary vitamin D binding site of LG (Proteins: Structure, Function, and Bioinformatics) ---16
Abstract ---17
Introduction---18
Materials and Methods---21
Results---26 Discussion ---34 Acknowledgments ---42 References ---42 Figure Legends---51 Tables---55 Figures ---58
Section 2: Rational design for crystallization of LG-vitamin D complex (Crystal Growth & Design) ---68
Abstract ---69
Introduction---69
Materials and Methods---73
Results and Discussion---76
Conclusion---86 Acknowledgments ---87 References ---87 Figure Legends---94 Tables---97 Figures --- 100
Abstract --- 107
Introduction--- 108
Results--- 109
Discussion --- 117
Materials and Methods--- 122
Acknowledgments --- 129 References --- 129 Figure Legends--- 135 Tables--- 139 Figures --- 141 Appendix --- 149 Curriculum vitae --- 150 Publications--- 158 Deposited Structure--- 230
Overview
Bovine β-lactoglobulin (LG) is a major whey protein consisting about 10-15% of total proteins in milk.1 Due to its thermally unstable and molten-globule nature, LG has been studied extensively for its physical and biochemical properties in the past 40 years.2-8 The overall folding of LG is remarkably similar to that of the human plasma retinol binding
protein9-12 and human tear lipocalin13, known as members of the lipocalin superfamily. LG comprises of 162 amino-acid residues with two disulfide linkages and one free cysteine. It has predominantly a β-sheet configuration containing nine anti-parallel β-strands from A to I.14-16 Topographically, β-strands A-D form one surface of the barrel (calyx), whereas strands E-H form the other.17-19 The only α-helical structure with three turns is at the COOH-terminus (residues 130-141), which is followed by a β-strand I lying on the outer surface of the calyx.20 The structural and functional relationship of this helical region is not yet clearly defined. The calyx has a remarkable ability to bind hydrophobic molecules such
as retinol,fatty acids and vitamin D.21-24 It seems clear that the binding of fatty acid, retinol, and vitamin D is within the central calyx of the protein; however, the existence of a second
ligand binding site beyond the calyx is a matter of controversy.17
LG is quite sensitive to thermal denaturation, the secondary structure is altered upon
heating with a transition temperature at 70-80 °C. Recently, it has shown that the
and β-strand D of the calyx is directly involved in the unfolding during the thermal
denaturation.8 As a result, the binding of palmitate or retinol to the central calyx is diminished.8 The EF loop of LG is known to act as a gate over the calyx;14 at pH values lower than 6 the loop is in a “closed” position. We manipulated the binding capability by
switching off the gate of calyx at low pH or denaturing the conformation of the calyx by heat
treatment to further substantiate the “two site hypothesis”.
Although the biological functions of LG still remain elusive, some essential functions of
LG, such as cholesterol lowering,7 modulation of immune system,3,17 binding of retinol, fatty acid, and vitamin D,21-24 and prevention of oxidative stress,26,27 have been reported. Biological function of LG in terms of the transport role of LG for retinol, fatty acids, and
vitamin D is still unproven. Intact LG has been demonstrated to resist acid and to be
superpermeable to cross the epithelium cells of the gastrointestinal tract via a receptor.28 The LG therefore may be an effective vehicle in transporting the hydrophobic ligands.
Vitamin D (cholecalciferol), a hormone that regulates bone development, metabolism,
calcium homeostasis, cell growth and cell differentiation, can be obtained through
endogenous pathways (photolysis of vitamin D precursors in the skin).29-31 Other major sources of vitamin D include fortified milk, fish and vitamin D-containing supplements. 29-31 Latitude, season, age, skin color, and skin coverage can all potentially influence the amount of
increased plasma vitamin D concentration is associatedwith decreased incidence of colon,
prostate, and breast cancers, osteoarthritis, autoimmune diseases, and diabetes.33-37 Increased mean 25-hydroxyvitamin D was significantly related to increasing milk
consumption in the summer and winter (P = 0.02 and P= 0.01).38 Because the binding of vitamin D to milk LG is well known, we can test the hypothesis questioning whether LG
enhances the transport of vitamin D of milk.
The present dissertation is divided as three sections:
1. The existence and location of secondary vitamin D binding site of LG 2. Rational design for crystallization of LG-vitamin D complex
3. Evidence for LG involvement in vitamin D transport in vivo
Section 1: The existence and location of secondary vitamin D binding site of LG
The existence of a second vitamin D binding site beyond the calyx is controversial.17 It has been postulated that there are two binding sites for vitamin D based on the work of
Swaisgood and de Wolf39-43 using fluorescence binding assays. Nevertheless, the location of the secondary binding site remote from the calyx has been implicated,9,10 but has not yet been identified by the crystal structure of bovine LG with vitamin D2.17 We first conducted a
ligand binding assay using the fluorescence changes by retinol, palmitic acid, and vitamin D3
the maximal binding stoichiometry of vitamin D3 with LG was 2:1, whereas that of retinol or
palmitic acid was 1:1, suggesting that there was another binding site for the vitamin D3
molecule. Second, we manipulated the binding capability by switching off the gate of calyx
at low pH (2-6)14 to further substantiate the “two site hypothesis”. As expected, the binding capability of retinol and palmitic acid diminished under this condition, but it retained the
vitamin D binding (30% of the maximal binding) with a vitamin D3 to LG stoichiometry of
1:1. We also used a strategy to denature the conformation of the calyx by heat treatment and
conducted the binding assay after the calyx was thermally “disrupted”. Under this condition,
heated LG retained 40% of the vitamin D3 binding even at 100 °C heating for 16 min and its
vitamin D3 binding stoichiometry was found to be 1:1. These binding experiments suggest
that there is a secondary site for vitamin D3 binding, which is thermally resistant and distinct
from the calyx. To confirm the hypothesis that a second vitamin D3 binding site exists, we
determined the crystal structure of LG-vitamin D3 complex and attempted to identify, localize,
and characterize such a site. In this section of the thesis, we report a second vitamin D3
binding site identified by synchrotron X-ray diffraction (at 2.4 Å resolution). The second
binding site (defined as an exosite) is near the surface at the C-terminus (residues 136-149) containing part of an α-helix and a β-strand I with 17.91 Å in length, while the span of vitamin D3 is about 12.51 Å. A remarkable feature of the exosite is that it combines an
β-strand I providing another nonpolar residue (Ile147). They are linked by a hydrophobic loop (Ala142, Leu143, Pro144, and Met145). Thus, the binding pocket furnishes strong
hydrophobic force to stabilize vitamin D3 binding. This finding provides a new insight into
the exosite may provide another route for the transport of vitamin D in vitamin D fortified
dairy products.
Section 2: Rational design for crystallization of LG-vitamin D complex
Based on vitamin D3 binding assay of LG treated at various pH and temperature, we
concluded that a thermally stable exosite beyond the calyx exists for vitamin D binding.
We attempted to identify the regions of LG that might be available for the interaction with
vitamin D using well-known bioinformatics programs (Insight II,44 Q-SiteFinder,45 and GEMDOCK46). The Insight II44 is based on the size of surface cavities of a given protein without a specified ligand; it searches for the location and extent of the pocket according to
the geometric criteria. The Q-SiteFinder,45 however, defines a binding pocket only by energy calculations using a methyl probe for van der Waals interactions with a given protein.
The GEMDOCK46 is a more accurate docking program to dock a specific ligand with a given protein based on a nonbiased search for their interactions. Following the analyses of Insight
II, Q-SiteFinder, and GEMDOCK cross-docking, we were able to narrow the region of potential binding sites to two sites which were located near the C-terminal α-helical region.
secondary vitamin D binding site. Furthermore, we cocrystallized the LG-vitamin D3
complex which was prepared at pH 7 with a vitamin D3/LG ratio of 2 according to the
previous crystallographic study.47 We found a weak extra electron density that was located near the C-terminal α-helical region. With respect to cocrystallization, the maximum occupancy of the ligand should provide a better opportunity in growing high-quality crystals
of the ligand-protein complex. In this section of thesis, we reported a rationally designed
approach for preparing the complex of LG and vitamin D3 at various pH and vitamin D3/LG
ratios to optimize the occupancy of vitamin D3 and improve the electron density of the
secondary binding site. Finally, we identified an exosite for vitamin D binding to be located near the α-helix and β-strand I of LG using a crystal prepared at pH 8 with a vitamin D3/LG
ratio of 3:1. The strategy in practice may be useful for future identification of a
ligand-binding site in a given protein.
Section 3: Evidence for LG involvement in vitamin D transport in vivo
In the previous section, we proposed that this exosite contains a unique inverse γ-turn loop (residues 143-145 or Leu-Pro-Met), located between the α-helix and β-strand I, that is essential in forming a pocket to bind vitamin D according to the crystal structure of
We further tested this hypothesis using a mAb specific for this γ-turn region (selected from a battery containing 900 mAbs) as a probe, and then determined whether vitamin D might
interfere with the binding between LG and the mAb. We provide several observations that might support the idea that the γ-turn loop plays a role in the interaction between the exosite and vitamin D. First, substituting each Leu143 and Met145 with charged amino acids
resulted in a substantially decreased binding affinity of vitamin D3 (4.7-fold to 8.1-fold, which
is consistent with our previous crystallographic study showing the side chains of these two
residues to be directly involved in vitamin D binding. Second, substituting centered Pro144
by Ala also significantly attenuated the binding affinity (4.6-fold). Because Pro144 has no
direct contact with vitamin D3, we suggest that it might be essential to maintain the
conformation of the loop structure. Third, binding of vitamin D3 to LG attenuated the
recognition of a γ-turn loop specific mAb (1D8F8). Although the binding of vitamin D to milk LG is well known,17,39,42 whether it enhances the transport of vitamin D of milk is still unproven. We have demonstrated for the first time that LG is a fraction responsible for the
uptake of vitamin D3 from milk, using the mouse (n=95) as an animal model. Most
interestingly, we showed that native LG containing both the calyx and exosite had an efficacy
in vitamin D3 uptake almost twice that of heated LG containing a single exosite. Our study
therefore provides new insights concerning the value of supplementing dairy products with
Overall experimental design
Cloning LG into pQE-30 Fluorescence binding assay Isolate LG by Affinity column CD spectrum Site-directed Mutagenesis Isolate Milk Fraction Feeding Mouse Analyze serum vitamin D Predicting Pocket Docking Grow CrystalsCollect X-ray Data
Molecular Replacement Refine Model Analyze secondary Expression and Purification of rLG
Analyze S-S linkage of LG (trypsin
References
1. Hambling SG, MacAlpine AS & Sawyer L (1992) Beta-lactoglobulin. In Advanced Dairy
Chemistry I (Fox PF eds) pp. 141-190. Elsevier, Amsterdam.
2. Sawyer L & Kontopidis G (2000) The core lipocalin, bovine beta-lactoglobulin. Biochim
Biophys Acta 1482, 136-148.
3. Marshall K (2004) Therapeutic applications of whey protein. Altern Med Rev 9, 136-156.
4. Sava N, Van der Plancken I, Claeys W & Hendrickx M (2005) The kinetics of
heat-induced structural changes of beta-lactoglobulin. J Dairy Sci 88, 1646-1653.
5. Chen WL, Huang MT, Liu HC, Li CW & Mao SJT (2004) Distinction between dry and
raw milk using monoclonal antibodies prepared against dry milk proteins. J Dairy Sci 87,
2720-2729.
6. Song CY, Chen WL, Yang MC, Huang JP & Mao SJT (2005) Epitope mapping of a
monoclonal antibody specific to bovine dry milk: involvement of residues 66-76 of strand
D in thermal denatured beta-lactoglobulin. J Biol Chem 280, 3574-3582.
7. Chen WL, Hwang MT, Liau CY, Ho JC, Hong KC & Mao SJT (2005) β-Lactoglobulin is a Thermal Marker in Processed Milk as Studied by Electrophoresis and Circular Dichroic
8. Chen WL, Liu WT, Yang MC, Hwang MT, Tsao JH & Mao SJT (2006) A novel
conformation-dependent monoclonal antibody specific to the native structure of
beta-lactoglobulin and its application. J Dairy Sci 89, 912-921.
9. Papiz MZ, Sawyer L, Eliopoulos EE, North AC, Findlay JB, Sivaprasadarao R, Jones TA,
Newcomer ME & Kraulis PJ (1986) The structure of beta-lactoglobulin and its similarity
to plasma retinol-binding protein. Nature 324, 383-385.
10. Monaco HL, Zanotti G, Spadon P, Bolognesi M, Sawyer L & Eliopoulos EE (1987)
Crystal structure of the trigonal form of bovine beta-lactoglobulin and of its complex with
retinol at 2.5 A resolution. J Mol Biol 197, 695-706.
11. Sawyer L, Papiz MZ, North ACT & Eliopoulos EE (1985) Structure and function of bovine β-lactoglobulin. Biochem Soc Trans 13, 265-266.
12. Newcomer ME, Jones TA, Aqvist J, Sundelin J, Eriksson U, Rask L & Peterson PA (1984)
The three-dimensional structure of retinol-binding protein. EMBO J 3, 1451-1454.
13. Redl B (2000) Human tear lipocalin. Biochim Biophys Acta 1482, 241-248.
14. Qin BY, Bewley MC, Creamer LK, Baker HM, Baker EN & Jameson GB (1998)
Structural basis of the Tanford transition of bovine beta-lactoglobulin. Biochemistry 37,
15. Qin BY, Bewley MC, Creamer LK, Baker EN & Jameson GB (1999) Functional
implications of structural differences between variants A and B of bovine
beta-lactoglobulin. Protein Sci 8, 75-83.
16. Kuwata K, Hoshino M, Forge V, Era S, Batt CA & Goto Y (1999) Solution structure and
dynamics of bovine beta-lactoglobulin A. Protein Sci 8, 2541-2545.
17. Kontopidis G, Holt C & Sawyer L (2004) Invited review: beta-lactoglobulin: binding
properties, structure, and function. J Dairy Sci 87, 785-796.
18. Adams JJ, Anderson BF, Norris GE, Creamer LK & Jameson GB (2006) Structure of
bovine beta-lactoglobulin (variant A) at very low ionic strength. J Struct Biol 154,
246-254.
19. Bello M, Pérez-Hernández G, Fernández-Velasco DA, Arreguín-Espinosa R &
García-Hernández E (2008) Energetics of protein homodimerization: effects of water
sequestering on the formation of beta-lactoglobulin dimer. Proteins 70, 1475-1487.
20. Uhrinova S, Smith MH, Jameson GB, Uhrin D, Sawyer L & Barlow PN (2000) Structural
changes accompanying pH-induced dissociation of the beta-lactoglobulin dimer.
Biochemistry 39, 3565-3574.
21. Wu SY, Perez MD, Puyol P & Sawyer L (1999) beta-lactoglobulin binds palmitate within
22. Kontopidis G, Holt C & Sawyer L (2002) The ligand-binding site of bovine
beta-lactoglobulin: evidence for a function? J Mol Biol 318, 1043-1055.
23. Considine T, Singh H, Patel HA & Creamer LK (2005) Influence of binding of sodium
dodecyl sulfate, all-trans-retinol, and 8-anilino-1-naphthalenesulfonate on the
high-pressure-induced unfolding and aggregation of beta-lactoglobulin B. J Agric Food
Chem 53, 8010-8018.
24. Konuma T, Sakurai K & Goto Y (2007) Promiscuous binding of ligands by
beta-lactoglobulin involves hydrophobic interactions and plasticity. J Mol Biol 368,
209-218.
25. Nagaoka S, Futamura Y, Miwa K, Awano T, Yamauchi K, Kanamaru Y, Tadashi K &
Kuwata T (2001) Identification of novel hypocholesterolemic peptides derived from
bovine milk beta-lactoglobulin. Biochem Biophys Res Commun 281, 11-17.
26. Chevalier F, Chobert JM, Genot C & Haertle T (2001) Scavenging of free radicals,
antimicrobial, and cytotoxic activities of the Maillard reaction products of
beta-lactoglobulin glycated with several sugars. J Agric Food Chem 49, 5031-5038.
27. Liu HC, Chen WL & Mao SJT (2007) Antioxidant nature of bovine milk
28. Fluckinger M, Merschak P, Hermann M, Haertlé T & Redl B (2008)
Lipocalin-interacting-membrane-receptor (LIMR) mediates cellular internalization of
beta-lactoglobulin. Biochim Biophys Acta 1778, 342-347.
29. Holick MF (2003) Vitamin D: A millenium perspective. J Cell Biochem 88, 296-307.
30. Vieth R (2005) The pharmacology of vitamin D, including fortification strategies. In
Vitamin D vol. 2 (Feldman D, Pike JW & Glorieux FH, eds), pp. 995–1015. Elsevier.
31. Bortman P, Folgueira MA, Katayama ML, Snitcovsky IM & Brentani MM (2002)
Antiproliferative effects of 1,25-dihydroxyvitamin D3 on breast cells: a mini review. Braz
J Med Biol Res 35, 1-9.
32. Webb AR (2006) Who, what, where and when-influences on cutaneous vitamin D
synthesis. Prog Biophys Mol Biol 92, 17-25
33. Giovannucci E (2005) The epidemiology of vitamin D and cancer incidence and mortality:
a review (United States). Cancer Causes Control 16, 83-95.
34. Cui Y & Rohan TE (2006) Vitamin D, calcium, and breast cancer risk: a review. Cancer
Epidemiol Biomarkers Prev 15, 1427-1437.
35. Knight JA, Lesosky M, Barnett H, Raboud JM & Vieth R (2007) Vitamin D and reduced
risk of breast cancer: a population-based case-control study. Cancer Epidemiol
36. Lappe JM, Travers-Gustafson D, Davies KM, Recker RR & Heaney RP (2007) Vitamin D
and calcium supplementation reduces cancer risk: results of a randomized trial. Am J Clin
Nutr 85, 1586-1591.
37. Holick MF (2007) Vitamin D deficiency. N Engl J Med 357, 266-281.
38. Sahota H, Barnett H, Lesosky M, Raboud JM, Vieth R & Knight JA (2008) Association of
vitamin D related information from a telephone interview with 25-hydroxyvitamin D.
Cancer Epidemiol Biomarkers Prev 17, 232-238.
39. Wang Q, Allen JC & Swaisgood HE (1997) Binding of vitamin D and cholesterol to
beta-lactoglobulin. J Dairy Sci 80, 1054-1059.
40. Wang Q, Allen JC & Swaisgood HE (1997) Binding of retinoids to beta-lactoglobulin
isolated by bioselective adsorption. J Dairy Sci 80, 1047-1053.
41. Wang Q, Allen JC & Swaisgood HE (1998) Protein concentration dependence of
palmitate binding to beta-lactoglobulin. J Dairy Sci 81, 76–81.
42. Wang Q, Allen JC & Swaisgood HE (1999) Binding of lipophilic nutrients to
beta-lactoglobulin prepared by bioselective adsorption. J Dairy Sci 82, 257-264.
43. Muresan S, van der Bent A & de Wolf FA (2001) Interaction of beta-lactoglobulin with
small hydrophobic ligands as monitored by fluorometry and equilibrium dialysis:
44. Xiao L, Cui X, Madison V, White RE & Cheng KC (2002) Insights from a
three-dimensional model into ligand binding to constitutive active receptor. Drug Metab
Dispos 30, 951-956.
45. Laurie AT & Jackson RM (2005) Q-SiteFinder: an energy-based method for the
prediction of protein-ligand binding sites. Bioinformatics 21, 1908-1916.
46. Yang JM & Chen CC (2004) GEMDOCK: a generic evolutionary method for molecular
docking. Proteins 55, 288-304.
47. Qin BY, Creamer LK, Baker EN & Jameson GB (1998) 12-Bromododecanoic acid binds
Section 1: The existence and location of secondary
vitamin D binding site of LG (Proteins: Structure,
Abstract
β-lactoglobulin (LG), one of the most investigated proteins, is a major bovine milk protein with a predominantly β structure. The structural function of the only α-helix with three turns at the C-terminus is unknown. Vitamin D3 binds to the central calyx formed by the
β-strands. Whether there are two vitamin D binding-sites in each LG molecule has been a subject of controversy. Here, we report a second vitamin D3 binding site identified by
synchrotron X-ray diffraction (at 2.4 Å resolution). In the central calyx binding mode, the aliphatic tail of vitamin D3 clearly inserts into the binding cavity, where the 3-OH group of
vitamin D3 binds externally. The electron density map suggests that the 3-OH group
interacts with the carbonyl of Lys60 forming a hydrogen bond (2.97 Å). The second binding site, however, is near the surface at the C-terminus (residues 136-149) containing part of an α-helix and a β-strand I with 17.91 Å in length, while the span of vitamin D3 is about 12.51 Å.
A remarkable feature of the second exosite is that it combines an amphipathic α-helix providing nonpolar residues (Phe136, Ala139, and Leu140) and a β-strand providing a nonpolar (Ile147) and a buried polar residue (Arg148). They are linked by a hydrophobic loop (Ala142, Leu143, Pro144, and Met145). Thus, the binding pocket furnishes strong hydrophobic force to stabilize vitamin D3 binding. This finding provides a new insight into
the interaction between vitamin D3 and LG, in which the exosite may provide another route
for the crystal structure of LG-vitamin D3 complex described in this work have been
deposited in the PDB (access code 2GJ5).
(Keywords: fluorescence ligand binding assay, crystallography, localized alternative vitamin D binding site, thermal denaturation, amphipathic helix)
Introduction
Bovine β-lactoglobulin (LG) is a major whey protein in milk to an extent of about 50%.1
Because of its thermally unstable and molten-globule nature, LG has been studied extensively for its physical and biochemical properties in the past 40 years.2-6 Although the biological functions of the protein still remain elusive, some essential functions of LG, such as cholesterol lowering, modulation of immune system, transport of retinol, fatty acid, and vitamin D,7-9 and prevention of oxidative stress,10,11 have been reported.
Several crystal forms of bovine LG have been described.12-21 Of these, lattices X and Z (Space group P1 and P3221) have been investigated at a low resolution.14 A high resolution
study of another crystal form, lattice Y, has yielded a chain trace and a preliminary model.14 The overall folding turns out to be remarkably similar to that of the human plasma retinol binding protein15,16,22,23 and human tear lipocalin,24 known as members of the lipocalin superfamily. As shown in Figure 1A, LG comprises of 162 amino acid residues with two
containing nine antiparallel β-strands from A to I.18,19,25 Topographically, β-strands A-D
form one surface of the barrel (calyx), whereas β-strands E-H form the other.
The only α-helical structure with three turns is at the COOH-terminus (residues 130-141), which is followed by a β-strand I lying on the outer surface of the calyx.26 The structural
and functional relationship of this helical region is not yet clearly defined. Studies on the crystal structure of LG-retinol complex at 2.5 Å resolution by Monaco et al. have pointed out that there is a surface pocket consisting of almost completely hydrophobic residues near the helical region.16
The remarkable ability of the calyx to bind hydrophobic molecules, such as retinol, fatty acids, and vitamin D (Figure 1B),27-29 has recently been reviewed by Kontopidis et al.9 It seems clear that the binding of fatty acid, retinol, and vitamin D is within the central calyx of the protein; however, the existence of a second ligand binding site beyond the calyx is a matter of controversy.9 Interestingly, an early study has suggested that a hydrophobic pocket, formed by the α-helix and the surface of the barrel, also exists.16 This surface pocket,
limited by Phe136 and followed by residues 139-143 (Ala-Leu-Lys-Ala-Leu), has been suspected to potentially bind retinol;16 however, later studies by X-ray diffraction did not reveal that retinol could occupy this site.9,29 Using cis-parinaric acid as a ligand, Dufour et
al.30 have suggested that this ligand is bound in this hydrophobic pocket. It remains unclear
also controversial.9,31 It has been postulated that there is another binding site, in addition to the calyx, for vitamin D based on the work of Swaisgood and de Wolf31-35 by using biochemical binding assays. Subsequently, they also suggested that one vitamin D is bound in that hydrophobic site. Nevertheless, the location of the secondary binding site remote from the calyx has been implicated,15,16 but has not yet been identified by the crystal structure of bovine LG with vitamin D2.9
Spectroscopic studies and thermodynamic analysis of the calorimetric signal have demonstrated that irreversible unfolding of the LG structure occurs upon thermal treatment above its transition temperature, 65-70 °C.36 Recently, we have shown that the conformational changes of LG are rapid and extensive at temperatures above the transition,5 and β-strand D of the calyx is directly involved in the unfolding during the thermal denaturation.4 As a result, the binding of palmitate or retinol to the central calyx is diminished. In the present study, we also demonstrated that the maximal binding ratios of vitamin D3 to LG were 2:1, similar to that established by Wang et al.31 Our next strategy
was to denature the conformation of the calyx by heating at 100 °C for 16 min; under this condition the calyx pocket was thermally “removed”. We then tested whether the thermally denatured LG was able to bind vitamin D3, palmitate, and retinol. Interestingly, only
which is thermally independent. To confirm the hypothesis that a second vitamin D3 binding
site exists, we determined the crystal structure of LG-vitamin D3 complex and attempted to
identify, localize, and characterize such a site.
Materials and methods
Materials
LG was purified from raw milk using saturated ammonium sulfate (40%) followed by a G-150 column chromatography of the upper fraction as described previously.3 All-trans retinol, palmitic acid, vitamin D3 (cholecalciferol), and N-acetyl-L-tryptophanamide were
purchased from Sigma-Aldrich (St. Louis, MO).
Ligand binding to LG
LG stock solution was prepared in 0.01M phosphate buffered solution, pH 8.0 (PB). Retinol, palmitate, and vitamin D3 were prepared using absolute ethanol and purged with
nitrogen and stored at -80 °C in the dark. All the binding assays described below were conducted at 24 °C. The ligand binding assay of LG was measured by fluorescence emission techniques similar to that previously described.4,31,32 In general, the binding of retinol to LG was measured by extrinsic fluorescence emission of a retinol molecule at 470 nm using excitation at 287 nm, whereas binding of palmitate or vitamin D3 to LG was measured by the
Fluorescence spectra were recorded with a fluorescence spectrophotometer (Hitachi F-4500; Tokyo, Japan). For the titration experiment, 5 μM of native LG was instantly incubated with various proportions of retinol or vitamin D3 (0.625-25 μM) at pH 8.0. For palmitate
(2.5-100 μM), 20 μM of native LG was used. A solution of N-acetyl-L-tryptophanamide with an absorbance at 287 nm - equal to that of the protein - served as a blank. The change in fluorescence of this solution with titration caused by an inner filter effect was corrected as described by Cogan et al.37 The change in fluorescence intensity at 332 nm or 470 nm was assumed to depend on the amount of protein-ligand complex, which allowed the calculation of a, the fraction of unoccupied ligand-binding sites on the protein: a = (F - Fsat)/(F0 - Fsat).
Here F is the fluorescence intensity at a certain titration ratio, Fsat is the corrected fluorescence
intensity of LG solution with its sites saturated, and F0 is the initial corrected fluorescence
intensity. These data were then used to construct a plot of PTa versus RTa/(1 - a) according
to the equation: PTa = (1/n)[RTa/(1 - a)] - Kdapp/n, where PT is the total protein concentration,
n is the number of binding sites per molecule, RT is the total ligand concentration, and Kdapp
is the apparent dissociation constant.
To study the effect of pH on the binding capacity, native LG between 5 and 20 μM was instantly incubated with retinol, vitamin D3, or palmitate between 5 and 20 μM. To
incubated with retinol, palmitate, or vitamin D3 at pH 8.0. The final concentration of ethanol
in the reaction mixture was kept less than 3% (vol/vol) for all the experiments mentioned above. The ligand binding ability of LG was calculated as described previously.4,31 For the titration curve experiment, the data were expressed as the percentage of emission of LG that had the maximal binding ratio. For the heat denaturation experiment, the data were expressed as the percentage relative to native LG. All the data were collected in triplicate determinations.
Circular dichroism spectrum
For the circular dichroism (CD) spectral measurements, each sample (0.5 mg/mL) was heated in 20 mM Tris, pH 8.0.4,38 The CD spectra were recorded on a spectropolarimeter (Jasco-J715; Tokyo, Japan) at 24 °C over wavelength ranges from 200 to 250 nm, and recorded at a scan speed of 20 nm/min. All spectra were measured twenty times in a cuvette with a path length of 1.0 mm. Each LG sample (100 μL) was preheated at 50, 60, 70, 80, 90, and 100 °C for 16 min and instantly stopped in a 20 °C water bath before an immediate measurement.
Crystallization
Purified LG was concentrated to 20 mg/mL in 20 mM Tris, pH 8.0. Vitamin D3 stock
solution made up as 50 mM in ethanol was added to LG solution to give a molar ratio of 3:1 and incubated for 3 h at 37 °C. Precipitation immediately occurred when LG and vitamin D3
were mixed, and the solution drops became clear on the slides after 3-4 days. Crystallization of the LG-vitamin D3 complex was achieved using the hanging-drop vapor-diffusion method
at 18 °C with 2 μL hanging drops containing equal amounts of LG-vitamin D3 complex and a
reservoir solution (0.1M HEPES containing 1.4M trisodium citrate dehydrate, pH 7.5). Crystals 0.1-0.2 mm long grew after 7 days.
Crystallographic data collection and processing
The crystals were mounted on a Cryoloop (0.1-0.2 mm), dipped briefly in 20% glycerol as a cryoprotectant solution, and frozen in liquid nitrogen. X-ray diffraction data at 2.4 Å resolution were collected at 110 K using the synchrotron radiation on the beamlines BL12B2 at SPring-8 (Harima, Japan) and BL13B at NSRRC (Hsinchu, Taiwan). The data were processed using the HKL2000 program.39 The crystals belong to the space group P3221 with
unit cell dimensions of a = b = 53.78 Å and c = 111.573 Å. There is one molecule per asymmetric unit according to an estimated solvent content in a reasonable region. Details of the data statistics are given in a table in the text.
Crystal structure determination and refinement
The structure of the LG-vitamin D3 was determined by molecular replacement40 as
implemented in CNS v1.141 using the crystal structure of bovine LG (PDB code 2BLG)18 as a search model. The LG molecule was located in the asymmetric unit after rotation and
The composite omitted electron density maps with coefficients |2Fo - Fc| were calculated and
visualized using O v7.0,42 and the model was rebuilt and adjusted iteratively as required. Throughout the refinement, a random selection (8%) of the data was placed aside as a “free data set”, and the model was refined against the rest of the data with F ≥0 as a working set.43-45 The monomer protein model was initially refined by rigid-body refinement using the data from 15.0 to 3.0 Å resolution, for which the group temperature B values were first restrained at 20 Å2. This refinement was followed by simulated annealing using a slow cooling protocol with a starting temperature of 2500 K, provided in CNS, applied to all data between 15.0 and 2.4 Å. The bulk solvent correction was then applied, and group B factors were adjusted. After several cycles of positional and grouped B factor refinement interspersed with interactive modeling, the R-factor for the LG-vitamin D3 complex decreased
to about 28% with the Rfree around 36%. Two elongated extra electron densities with one
LG molecule were clearly visible and recognized as the vitamin D3 in σA-weighted |Fo - Fc|
difference maps. Two vitamin D3 molecules were then adjusted and well fitted into the
density map. The refinement then proceeded with another cycle of simulated annealing with a slow cooling, starting at a temperature of 1000 K. The vitamin D3 molecules were
adjusted iteratively according to the omitted electron density maps. Finally, water molecules were added using the program CNS v1.1.
The final model of LG and vitamin D3 complex contains 1272 nonhydrogen protein
atoms for the monomer LG, 28 atoms for one vitamin D3 molecule, and 38 water molecules.
The refinement statistics are given in the text. The correctness of stereochemistry of the model was verified using PROCHECK.46 The calculations of r.m.s. deviations from ideality47 for bonds, angles, and dihedral and improper angles performed in CNS showed satisfactory stereochemistry. In a Ramachandran plot,48 all main chain dihedral angles were in the most favored and additionally allowed regions except for Tyr99.
Coordinates
Atomic coordinates for the crystal structure of LG-vitamin D3 complex described in this
work have been deposited in the PDB (access code 2GJ5).
Results
Binding of LG to retinol, palmitate, and vitamin D3
Using the titration method previously established by Wang et al.,31,32 we show that the maximal binding of vitamin D3 with LG was achieved at a 2:1 ratio; whereas the binding for
retinol or palmitate remained to be 1:1 (Figure 2). This result is similar to that reported previously by Wang et al.31,32 and tends to support a notion that LG binds two vitamin D3
Effect of pH on LG binding to retinol, palmitate, and vitamin D3
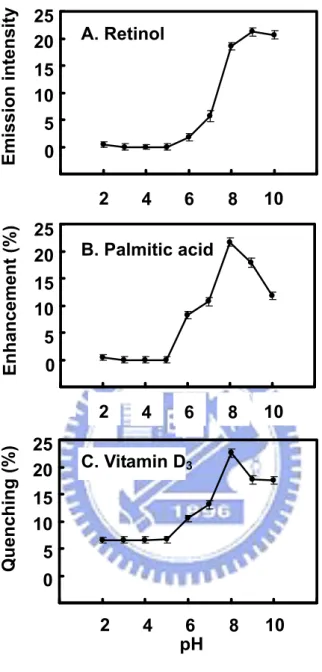
It has been postulated that pH plays a crucial role in controlling the opening of the calyx to allow the entrance of LG ligands. At low pH or below the Tanford transition (about pH 6), the EF loop (the calyx cap) is closed, disallowing the binding of the ligands. To explore whether there is another vitamin D3 binding site that may not be affected by the Tanford
transition, we monitored the binding of vitamin D3 at various pH while using retinol or
palmitate as a reference. A notable transition of vitamin D3 binding to LG was found to
occur between pH 6.0 and 8.0 (Figure 3), similar to that of retinol and palmitate. The binding to vitamin D3 or palmitate was decreased to some extent at pH 9-10 (Figure 3). This
could be due to the protonated state of Lys69 inside the calyx being neutralized at a high pH as suggested previously.4,29 It is of interest to note that unlike retinol and palmitate; LG at a pH between 2 and 6 still retains about 35% of the maximal binding for vitamin D3 (Figure
3C). The data imply that there is a possible secondary binding site for vitamin D3 that is
independent of the calyx.
Effect of heating on LG binding to retinol, palmitate, and vitamin D3
The pH titration experiment described above was unable to yield an accurate explanation of the existence of another binding site for vitamin D3. Our previous work showed that
thermally denatured LG (heated to 100 °C for 5 min) was unable to bind to retinol and palmitate because of the unfolding of the calyx.4 In the next experiment, our strategy was to
thermally “remove” the calyx and then test whether heated LG retained an activity allowing vitamin D3 binding. Figure 4 shows a notable and sharp decrease in retinol, palmitate, and
vitamin D3 binding to LG heated between 70 °C and 80 °C over time. The change of
binding is consistent to the molten-globule nature of LG, which correlates to its transition temperature.4 At temperatures above 80 °C, the protein lost its binding ability to retinol and palmitate in a time-dependent fashion, but it still retained 40% of the binding to vitamin D3
even after being heated at 100 °C for 16 min (Figure 4C). The heated LG (100 °C for 16 min) was further titrated with the binding of vitamin D3 in excess. Figure 4D reveals that
there was about 42% of maximal binding of vitamin D3 relative to that using native LG.
There was no fluorescence change while titrating with retinol or palmate (data not shown). Remarkably, a maximal stoichiometry of 1:1 was observed between the denatured LG and vitamin D3. Thus, it suggests that a thermally stable site exists in LG to bind vitamin D3.
Comparison of binding affinity of LG to retinol, palmitate, and vitamin D3
We further determined the binding affinities of LG for retinol, palmitate, and vitamin D3
using the method previously described by Wang et al.31,32 The fluorescence data obtained for retinol, palmitate, and vitamin D3 binding to native LG are shown in Figure 2 (right
panels), while the vitamin D3 binding to heated LG (100 °C for 16 min) is shown in Figure
vitamin D3 to native LG using this method was about 5 nM (Kdapp= 4.74 ± 0.37 nM) and
appears to be 5-10 times greater than that of retinol and palmitate. On the other hand, the binding affinity of vitamin D3 to heated LG was attenuated at about 45 nM (Kdapp= 45.67 ±
3.12 nM), but is within the same order as that between native LG and retinol or palmitate. Because heating LG also induces the aggregation of LG,5 the overall attenuated binding affinity of the “secondary site” indicates a structural change in the second binding site in heated LG. Nevertheless, the data temptingly suggest that the putative second binding site is somewhat heat resistant.
Overall crystal structure of LG-vitamin D3 complex
We have clarified that there are two vitamin D binding sites on LG according to ligand binding assay performed in solution. Protein crystallography was used to locate the secondary vitamin D binding site of LG. There are several crystal forms of bovine LG that have been well reported, including the triclinic (lattice X), orthorhombic (lattice Y), and trigonal (lattice Z) forms belonging to space groups P1, C2221, and P3221, respectively.12-21
The crystal of the LG-vitamin D3 complex we obtained was found to be a trigonal (lattice Z)
space group P3221 based on the data of 2.4 Å resolution. The final model comprised of 161
residues and its refinement statistics of the complex are given in Table 2. The discrepancy indices for R and Rfree are 23.86% and 26.12%, respectively. The LG-vitamin D3 complex
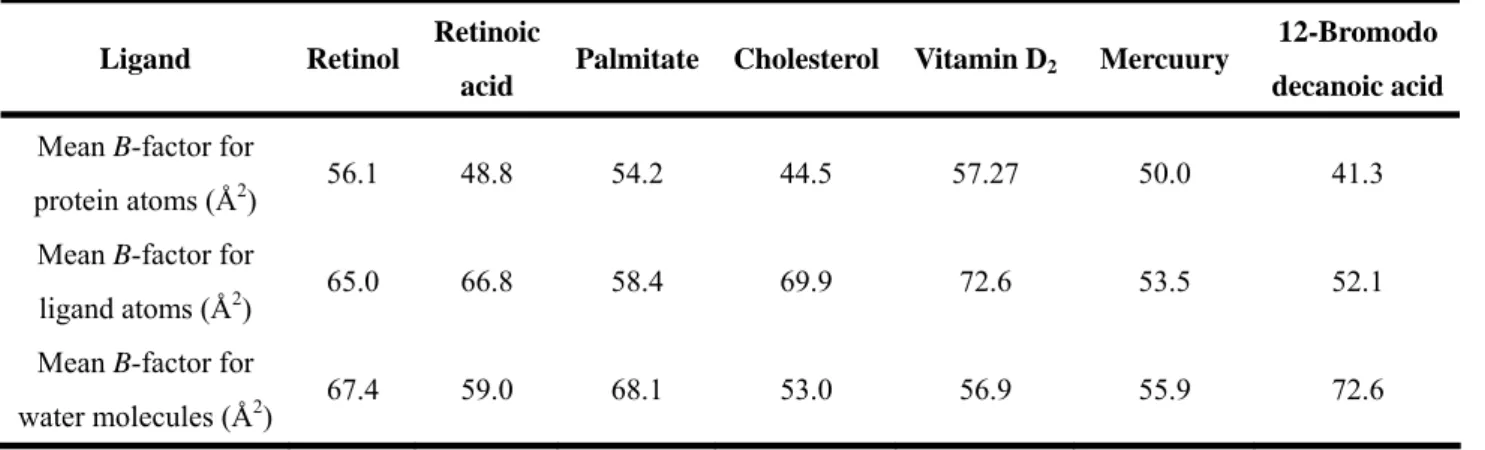
bond length and bond angle deviation from ideality 0.007 Å and 1.264° respectively (Table 2). A Ramachandran plot reveals that only one residue is in the disallowed regions (Tyr99), which arises in most of the lipocalin family as a result of the γ-turn associated with the sequence “TDY” (residues 97-99). The average temperature factor for all protein atoms is 55.02 Å2, which is considered to be inadequate for established crystallographic standards. This may be explained by LG having nearly 25% of the residues located around flexile surface loops and NH2- and COOH-terminal regions. Previously published statistics of LG
complexes9,27,29 show a B-factor range of 41.3-57.27 (Table 3), which accommodates for the elevated values acquired in this study that seem to deviate from the norm.
The overall topology of LG is similar to that previously described15-19 with a well-defined antiparallel β-sheet structure and flexile loops connecting the secondary structure elements. The loops AB, CD, EF, and GH are more flexile than the others, consistent with the observations from other reported crystal structures.15-19 The EF loop that acts as a flap is in the open position of the central calyx, which is expected when vitamin D3 is
present.
Space-filling drawings of the LG-vitamin D3 complex show that there are two domains
for vitamin D3 binding (Figure 5). One vitamin D3 molecule inserts almost perpendicularly
α-helix and β-strand I (Figure 1) as shown in Figure 6. The B-factor for the vitamin D3
molecules in the calyx and the second site are 39.19 and 46.90, respectively (Table 2). Proximity of these values to the published data9,27,29 reveals the rigidity of vitamin D3 bound
to LG (Table 3). For the second site, vitamin D3 is close to the surface of LG with 17.91 Å
in length, while the span of vitamin D3 is about 12.51 Å. Under this orientation, the A ring
or the aliphatic tail following the C/D rings of vitamin D3 interacts with the β-strand I or the
α-helix of LG, respectively (Figure 6). We putatively defined this second vitamin D3
binding site as an exosite.
Figure 6 depicts that the bulk of the electron density is sufficient to cover the entire extent of vitamin D3 in the calyx (in stereo view) as well as that in the exosite. The aliphatic
tail of vitamin D3 (C17-27) is oriented inside the calyx with the 3-OH group of vitamin D3
near the outside of the pocket.
Superimposing the current LG-vitamin D3 and previously described LG models (PDB
codes 1BSQ) in the region of the calyx (Figure 7A) reveals that the movement of the external EF “gate loop” of LG-vitamin D3 is quite similar to that of the LG-retinol complex18,29 (data
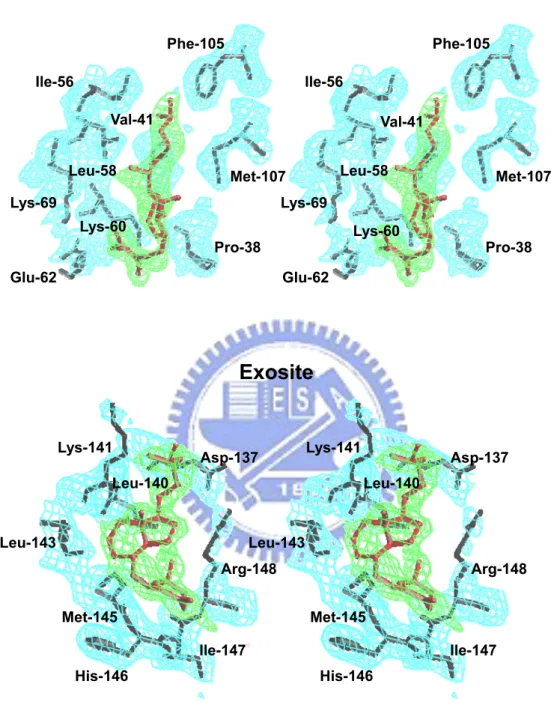
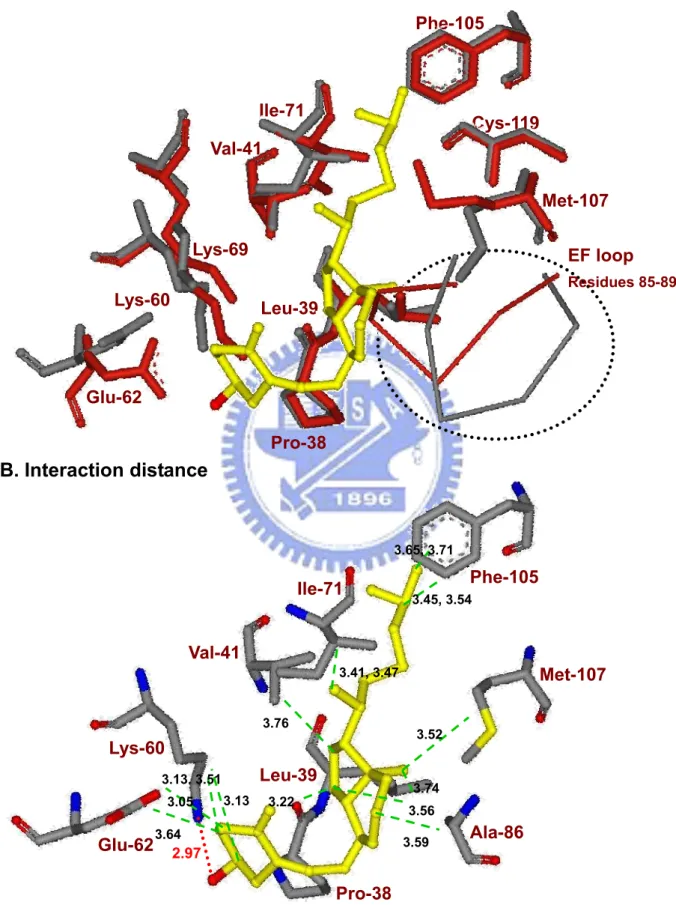
not shown). Some side chains, such as Lys60, Glu62, Phe105, and Met107, require significant reposition to make room for vitamin D3 insertion into the calyx. The atoms of
vitamin D3 near or at the “mouth” of the calyx possess higher values of B-factors suggesting
shortest interaction distance is hydrogen bonding between the 3-OH group of vitamin D3 and
Lys60 (2.97 Å) of LG; whereas the only hydrogen bond involving retinol binding is that to Glu62.29 Hydrophobic interaction and the distances between the carbons of vitamin D3 and
Pro38, Leu39, Val41, Ile71, Ala86, Phe105, and Met107 of LG are also displayed.
With respect to the second binding site for vitamin D3, it appears that vitamin D3 is
bound to a surface pocket between the COOH-terminal α-helix and β-strand I (residues 136-149). The current LG-vitamin D3 and previously described native LG models in this
region (PDB codes 1BSQ) are superimposed and shown in Figure 8A. Notably, there is not much conformational change near the exosite of LG upon the vitamin D3 binding. Figure 8B
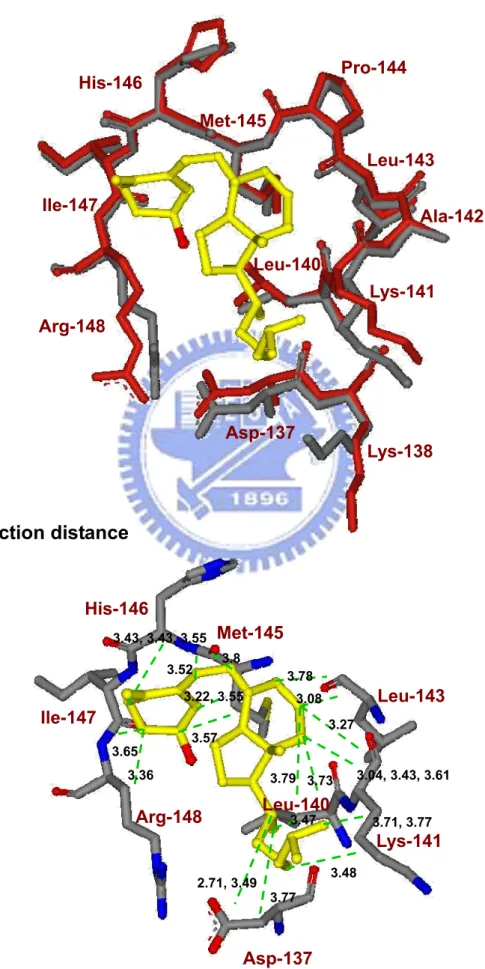
shows the distance between vitamin D3 and the amino acids involved (Asp137, Leu140,
Lys141, Leu143, Met145, His146, Ile147, and Arg148). Although some charged residues of the exosite are involved, their interaction with vitamin D3 is mainly hydrophobic with the
charged groups of LG sticking out of the pocket. The data suggest that the contact is via a hydrophobic interaction. There is no evidence that the hydroxyl group of vitamin D3
interacts with η1 N of Arg148 as the distance (4.9 Å) is greater than that of hydrogen bonding (bond length less than 3.13 Å) (Figure 8B).
Furthermore, LG is primarily oriented as a β structure (50%); the only α-helix region of LG consisting of three turns is located at the COOH-terminus between residues 130 and 141.
residues (Asp130, Glu131, Glu134, Asp137, Lys138, and Lys141) clustered at one face with hydrophobic or noncharged residues (Ala132, Leu133, Phe136, Ala139, and Leu140) at another face without an exception (Figure 9A). A remarkable feature is that the exosite is comprised of an amphipathic α-helix providing hydrophobic residues (Phe136, Ala139, and Leu140) at one side and a β-strand providing a hydrophobic Ile147 and a backbone His146 at the other side (Figure 9B). These two sides are linked by a loop containing hydrophobic residues Ala142, Leu143, and Pro144. Thus, the binding pocket provides a strong hydrophobic force to stabilize vitamin D3 binding. The stereo view of such an interaction is
also drawn in Figure 9B, depicting the binding on the surface of LG. The carbons of vitamin D3 (n = 27 in total) close to the surface are C5, C8, C10, C13, C14, C15, C16, C17, C18, C19,
C22, C24, C25, and C26 (n = 14). They are oriented toward the surface consistent with the analysis of Figure 5.
CD spectrum analysis of heated LG
In general, the CD spectrum at 222 nm is used for the calculation of the α-helical content of a given protein. Because the α-helix region of LG is located in the second binding site and heating LG retains ~40% of the maximal vitamin D3 binding to the whole LG molecule,
we monitored whether there were spectral changes of LG at 222 nm of LG upon heating at 50, 60, 70, 80, 90, and 100 °C. Figure 10 reveals that there were no significant changes in spectra at 222 nm. As expected, the β-configuration was disordered at temperatures above
70 °C, consistent with our previous observation.4 Thus, it temptingly offers support that the proposed exosite is somewhat thermally stable.
Discussion
During the past 40 years, LG has been extensively studied for its biochemical properties, and an abundance of literature exists about its physicochemical nature.9 Although the exact physiological functions of LG are not fully explored, one of its roles is to transport hydrophobic molecules, such as retinol, fatty acids, and vitamin D.49,50 The active form of vitamin D is 1α, 25(OH)2 vitamin D3, which maintains calcium homeostasis and plays
important roles on the immune system and prevents the growth and differentiation of cancer cells. Recent studies indicate that increased plasma vitamin D3 concentrations are associated
with decreased incidence of breast, ovarian, prostate and colorectal cancers,51 and osteoporotic fractures.52 The concentration of vitamin D3 in plasma is about 80 nM. A
double-blind placebo controlled study conducted in Europe indicated this level to be significantly reduced over the winter season (about 37%) due to the lack of exposure to sunlight.53 However, drinking vitamin D3 fortified milk (312 nM) significantly compensated
the seasonal loss of vitamin D by greater than 50%. For this reason, it has been recommended that milk enriched with vitamin D3 be provided in high-latitude European
complex in this study. Structurally, the only difference of vitamin D3 from D2 is the latter
being a double bond between the carbon positions 22 and 23 (Figure 1). Despite the minor difference, their binding characteristics to LG are similar.31 One additional goal in the present study is to test the possibility of binding vitamin D3 to thermally denatured milk,
which is often produced in the processing of milk.
With respect to the ligand binding of LG, many research groups28,31,32 have shown that the stoichiometry for binding retinol or palmitate to LG is 1:1, and most experimental evidence points to the calyx of LG as the binding site for retinol and palmitate.29 However, there remains a debate about the stoichiometry for vitamin D3 binding being 1 or 2. Wang et
al.31,34 proposed that LG has another binding site for vitamin D in addition to the central calyx,
but doubt has been raised based on crystallographic analysis.9,29 In fact, the presence of a secondary site for ligand binding has been described and proposed for some time,16 but the identity of a LG-ligand complex by an X-ray crystal structure has not been elucidated.9
In the present work, using the method of extrinsic fluorescence emission and fluorescence enhancement and quenching established previously,4,31,32 we show the maximal binding ratios of retinol or palmitate with LG to be 1:1, whereas it was 2:1 for that of vitamin D3 (Figure 2). The latter result is consistent to that reported by Wang et al.31,32 It is worth
mentioning that using the intrinsic fluorescence of Trp can give results that indicate significantly tighter ligand binding than other methods, especially equilibrium dialysis. This
effect is particularly noticeable when the Trp fluorescence decreases with ligand addition.9,35 A recent review9 suggests that a surface low-affinity binding site together with a central high-affinity binding site (calyx) would appear to satisfy most of the reported experimental observations. In brief, the diverse reports of more than a single binding site may be dependent on the method used.
In an attempt to resolve the controversy about an additional binding site for vitamin D3,
we used several additional approaches involving structural change to study the interaction between vitamin D3 and LG. First, it has been established that the EF loop acts as a gate
over the calyx.9,18,29,54 At a low pH, the loop is in a “closed” position, and the ligand binding into the calyx is inhibited. On the contrary, at a high pH above the Tanford transition, the loop is “open” allowing ligands to penetrate into the calyx.4,54,55 We explored the binding ability between LG and vitamin D3 at various pH. Similar to that of retinol and palmitate, the
present study shows that there is a notable transition of vitamin D3 binding to the calyx of LG
occurring between pH 6.0 and 8.0 (Figure 3). Most interestingly, at pH between 2 and 6 we show vitamin D3 interacting with LG (with about 35% of maximal binding ability), but not
retinol and palmitate (Figure 3C). Because the EF loop is “closed” below the Tanford transition, such binding suggests the presence of another binding site for vitamin D3.
tract.56,57 Notably, the binding of vitamin D3 or palmitate to LG was decreased to some
extent at pH 9-10 (Figure 3). One of the possible explanations is that the positively charged groups of lysine residues inside calyx are neutralized at pH above 8 resulting in a weakening of the interaction with the carboxyl group of palmitate.4
Second, we have shown that the β-strand D of the calyx is directly involved in the thermal denaturation.4 The conformational changes of LG were rapid, extensive, and irreversible upon heating over 70-80 °C.5 As a result, it completely diminishes the binding of palmitate and retinol. To test the hypothesis that there is a putative second binding site for vitamin D3 located independently from the calyx, we thermally denatured the calyx
(between 50 and 100 °C) and then conducted the binding for retinol, palmitate, or vitamin D3
over time (Figure 4). It is of interest that only vitamin D3 was able to bind to heated LG (at
100 °C for 16 min) with a stoichiometry of almost 1:1, instead of 2:1 (Figure 4D). Our data suggests that the second binding site for vitamin D3 is heat stable to some extent.
Third, analysis of the binding shows that the binding affinity between native LG and vitamin D3 is relatively high within a nM range, about 10 times greater than palmitate and
retinol (Table 1). The result is almost the same as that of vitamin D2 reported by Wang et
al.,31 but somewhat higher than the affinity reported for vitamin D3.31 The reason
contributing to such discrepancy remains elusive. One possibility may be due to the 5 μM concentration (pH 8.0) of LG that we employed, while Wang et al. used a 20 μM
concentration (pH 7.0) for a typical emission spectrum.34 The other possibility may be due to the pH; the fluorescence of Trp may be affected by ionization of neighboring prototropic groups or by conformation changes due to the dimerization of the protein.2,34 Nevertheless, the calculated affinity for vitamin D3 (Kdapp= 45.67 ± 3.12 nM) to putative second or
thermally stable binding site is about 10 times lower than that to native LG (calyx plus second site) (Table 1). Thus, it seems to be consistent with the hypothesis proposed by Kontopidis
et al.9 that the central site (calyx) is a main binding site possessing a high affinity for most of
the hydrophobic ligands, whereas the affinity for the secondary site is low. Ultimately, the secondary binding may depend on the nature of the ligands, such as their size, structure, and hydrophobicity. It would be of interest to further investigate other ligand bindings to heated LG, although the calculation of binding affinity of heated LG is somewhat complicated owing to the formation of large LG polymers as mentioned (see Results).
Finally, to confirm our hypothesis that there exists a second vitamin D3 binding site
remote from the calyx, we used a synchrotron radiation X-ray to determine the crystal structure of the LG-vitamin D3 complex. Our crystal of the complex is defined in trigonal
(lattice Z) space group P3221. This final model reveals that one vitamin D3 molecule binds
to the calyx (central internal binding-site) of LG and the other binds to the surface of LG between the α-helix and β-strand I (external binding site; Figure 5).
In the electron density map at 2.4 Å resolution, vitamin D3 is well fitted into the bulk of
electron density around the calyx or the exosite (Figure 6). In central calyx binding mode, the aliphatic tail of vitamin D3 clearly inserts into the binding cavity where the 3-OH group of
vitamin D3 binds externally. In an early report using vitamin D2,9 the end that inserted into
the calyx was not conclusively identified because the electron density was not enough to cover the entire ligand. It is not clear whether vitamin D3 is superior to vitamin D2 in
binding to LG. The other difference is that in our study the 3-OH group of vitamin D3 forms a
hydrogen bond with the carbonyl of Lys60 (Figure 7B) instead of Lys69 as proposed using vitamin D2.9 Again, the electron density was not strong enough for the outer extremity of
vitamin D2 making the exact conclusion difficult. Another explanation is that there might be
a significant difference in the orientation between vitamin D2 and D3, although the difference
in chemical structure is subtle. Regardless, the vitamin D3-LG binding mode is quite similar
to that of retinol-LG interaction over the calyx.29 Pro38, Leu39, Val41, Ile71, Ala86, Phe105, and Met107 are all involved in providing hydrophobic interactions with the displayed distance of less than 3.8 Å to the carbon backbone of vitamin D3 (Figure 7B).
In exosite binding mode, the vitamin D3 molecule attaches at a pocket between the
C-terminal α-helix and β-strand I (Figure 5). We specifically demonstrated that the exosite of LG provides a hydrophobic force to stabilize vitamin D3 (Figure 8B) and concluded that
no strong evidence to show that the 3-OH group of vitamin D3 is capable of interacting with
LG (Figure 8B). A stereo view shown in Figure 9B reveals that part of the vitamin D3
molecule is exposed toward the surface, consistent to that depicted in Figure 5. Although this second binding is located near the surface of LG, our data suggest that the binding affinity of vitamin D3 to exosite is reasonably high and almost equivalent to that of retinol or
palmitate to calyx (Table 1). Apparently, vitamin D3 interacts mostly with those
hydrophobic amino acids within residues 136-149 with distances less than 3.8 Å (Figure 8B). The linking-loop residues 142-145 (Ala-Leu-Pro-Met) between the α-helix and β-strand I are all hydrophobic. With such an orientation, a hydrophobic pocket is constructed in facilitating the binding for vitamin D3. It is worth mentioning that this exosite is very
similar to the surface hydrophobic site (mentioned and discussed above) that has been described by Monaco et al.16 and proposed by Wang et al.31
It is of interest that the α-helix involved in the exosite is typically amphipathic. This amphipathic region could be heat resistant as suggested from our binding experiment for vitamin D3 and heated LG. A similar situation is seen in a typically amphipathic
apolipoprotein A-I; its conformation and lipid binding properties are completely maintained upon heating over 100 °C.58 It might be worthwhile to study the crystal structure of the heated-LG-vitamin D3 complex to finally prove its heat resistance. Unfortunately, we are