Surface Seawater Temperature as A Potential Cause of
Delayed Arrival of the Japanese Eel
Anguilla japonica
Elvers on the Coast of Taiwan
Wann-Nian Tzeng(Received, March 08, 2006; Revised, April 23, 2006; Accepted, April 25, 2006)
ABSTRACT
The Japanese eel Anguilla japonica is one of the most important food fish in Taiwan. For cultivation, elvers of the eel are caught from the wild population in estuaries during their upstream migration. The relationship between recruitment timing of elvers and surface seawater temperature was analyzed based on the daily catch data of elvers in an estuary of northeastern Taiwan since 1980. It was found that most elvers recruited to the estuary during the period from December to January, with a rhythmic activity that follows the lunar cycle. However, the peak recruitment period in the years of 1986-87, 1990-91 and 1995-96 occurred in March, delayed approximately two months. This delay coincided with low temperature years. The low surface seawater temperature, in winter, in the continental shelf area of Taiwan may influence the recruitment period of the elvers in estuaries. Key words: Japanese eel, Elver, Catch, Water temperature, Otolith.
INTRODUCTION
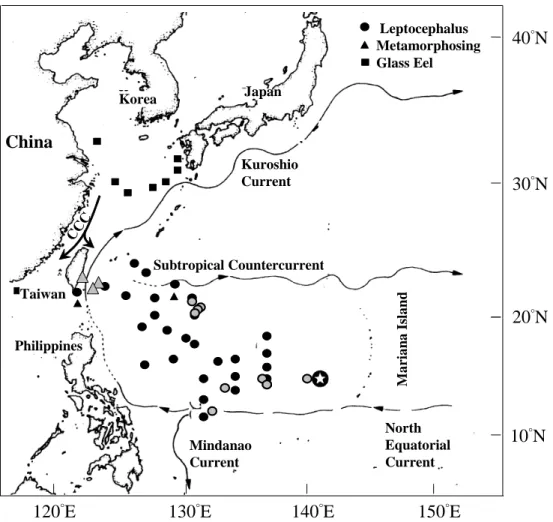
The Japanese eel, Anguilla japonica Temminck and Schlegel, is a catadromous fish. It spawns in the middle Pacific Ocean west of the Mariana Islands, 14-16°N 134- 143°E (Tsukamoto, 1992), during June and July (Tsukamoto, 1990; Tzeng, 1990; Tzeng and Tsai, 1994). Its leaf-like larva (leptocephalus) drifts on the North Equatorial Current and Kuroshio Current and metamorphoses to a glass eel in the continental shelf areas of Taiwan, Mainland China, Korea and Japan. Glass eels become pigmented elvers at entry of the estuaries. Elvers are harvested for cultivation or restocking at estuaries in winter during their upstream migration (Tzeng, 1983, 1985, 1986). Eel migration from the oceanic spawning grounds to the estuaries takes 4 to 5 months (Cheng and Tzeng, 1996). The eel lives in rivers for 5 to 12 years (Tzeng et al., 2000a). During late autumn when eels become
premature, they migrate downstream to the ocean to spawn and die (Tesch, 1977).
The catch of elvers was influenced by many environmental factors in estuaries (such as temperature, salinity, turbidity, pH, stream- water odor, and tidal cycle) as well as by moon phase and rainfall. These factors may act alone or in combination to influence the migration of elvers (Sloane, 1984; Tzeng, 1985; Sorensen and Bianchini, 1986; Tosi et al., 1990; Chen Lee et al., 1994). Previous research is primarily associated with the upstream behavior of elvers in estuaries. No documents have referred to the environ- mental factors in the offshore area which may influence the timing of peak recruitment of elvers at estuaries. For successfully harvesting elvers for aquaculture, it is necessary to predict the time of their estuarine arrival.
The transportation of eel larvae from the spawning ground to estuaries involves two development stages, which live in two different current systems. The two stages Institute of Fisheries Science, College of Life Science, National Taiwan University, Taipei, Taiwan 106, ROC. E-mail: [email protected]
transportation processes are shown in Figure 1. The lepotocephalus stage is transported by the warm North Equatorial Current and the Kuroshio Current from spawning ground to continental shelf. Glass eel stage is transported by the cold coastal current from coastal waters to the estuaries (Tzeng, 1996; Cheng and Tzeng, 1996). The fluctuation of these currents may influence the transporta- tion of eel larvae, and the fluctuation will be reflective of the seawater temperature.
Recently we found that estuarine arrival of elvers was delayed in particular years when surface seawater temperature was lower than other years. This study attempts
to clarify the cause and effect of the delayed arrival of elvers in estuaries of Taiwan.
MATERIALS AND METHODS The daily catches of Japanese eel elvers used in this study was collected in the coastal waters off Shuang-Chi River in the north- eastern Taiwan from 1980 to 1998 (sampling sites refer to Figure 1 of Tzeng (1985)). The elvers are caught daily by a hand trawling net on the coast through out the fishing season (Tzeng, 1985). The catch data collected on the coast are more suitable than those collected in the river mouth and inner river to
China
Korea Taiwan Philippines Japan Kuroshio Current Subtropical Countercurrent Mindanao Current Mar ian a Is la n d North Equatorial Current40°N
30°N
20°N
10°N
120°E
130°E
140°E
150°E
Leptocephalus Metamorphosing Glass Eel CC C
China
Korea Taiwan Philippines Japan Kuroshio Current Subtropical Countercurrent Mindanao Current Mar ian a Is la n d North Equatorial Current40°N
30°N
20°N
10°N
120°E
130°E
140°E
150°E
Leptocephalus Metamorphosing Glass Eel
China
Korea Taiwan Philippines Japan Kuroshio Current Subtropical Countercurrent Mindanao Current Mar ian a Is la n d North Equatorial Current40°N
30°N
20°N
10°N
120°E
130°E
140°E
150°E
Leptocephalus Metamorphosing Glass Eel
CC C
Fig. 1. Oceanic current and dispersal of leptocephali (circle), metamorphosing leptocephali (triangle), and marine glass eels (square) of the Japanese eel Anguilla japonica (CCC: China Coastal Current; Star: the presumed spawning ground; Solid symbols: data from Tsukamoto and Umezawa 1990, Tsukamoto 1992; Gray symbols: data from Liao et al. 1999 and unpublished) (modified from Tzeng, 2002).
study whether or not the arrival of elvers is delayed because the elvers collected on the coast are newly recruited (Tzeng, 1985). The catch data were reported from approximately 70 fishermen covering approximately 80% of all the total effort in the studied area.
Eels in the coastal waters of Taiwan include four species, i.e.; Anguilla japonica, A. marmorata, A. bicolor pacifica and A. celebesensis (Tzeng, 1982; Tzeng and Tabeta, 1983). The fishermen classify these elvers into white- and black-type according to the pigmentation on the tail (Tzeng, 1983). The white-type which belongs to the elvers of A. japonica is sold for eel culture, whereas the black-type which includes the elvers of A. marmorata, A. bicolor pacifica and A. celebesensis has not been cultured in Taiwan. Only the catch data of the white-type were used in this study.
To understand the relationship between delayed arrival of elvers and seawater tempera- ture, annual mean surface seawater tempera- ture in the offshore of the elver’s sampling areas, 121-124°E 25-26°N, was calculated from the data of the Integrated Global Ocean Service System Products Bulletin (http://ingrid. Idgo.columbia.edu/SOURCES/.IGOSS/nmc/)
In addition, the hatching date and growth rate of elvers were compared between a delayed year (1996) and a non-delayed year (1998) by examining daily growth increments in otoliths. The procedure of otolith prepara- tion for daily growth increment examination and age determination is the same as previous studies (Tzeng, 1990; Tzeng and Tsai, 1994; Cheng and Tzeng, 1996).
RESULTS 1. Daily changes of elver catch
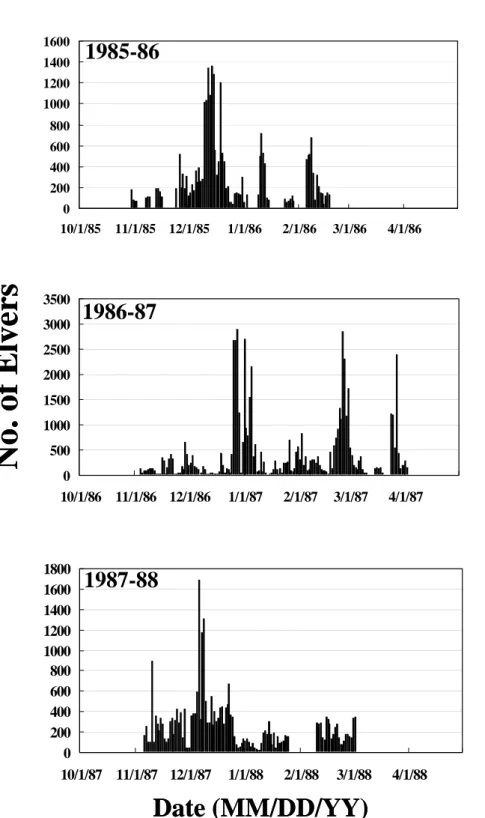
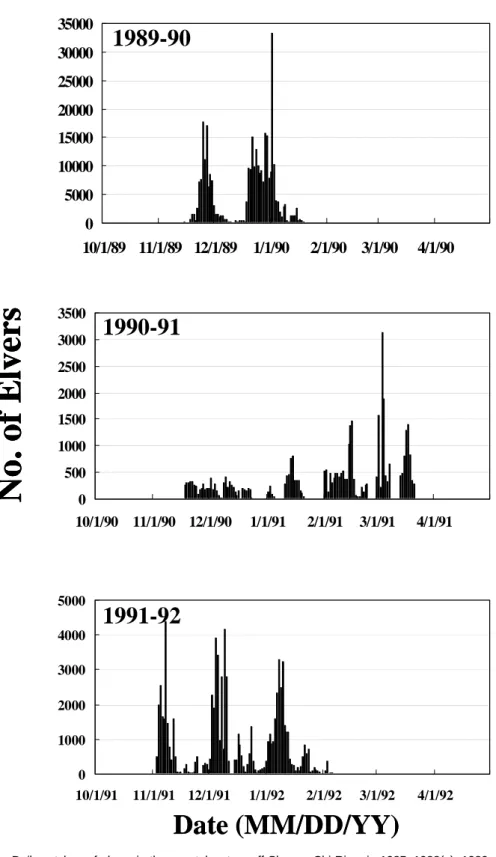
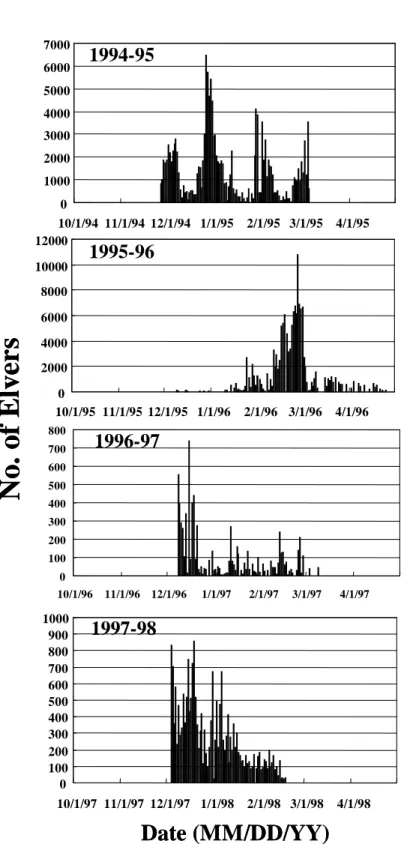
Daily catch of the Japanese eel elvers in the coastal waters off Shuang-Chi River in the years of 1985-88, 1989-1992 and 1994- 1997 was selected for analysis because surface sea water temperature decreased abnormally in these years (Figures 2a, b, and c). It was found that maximal daily catches of elvers were delayed in the years of 1986-
87, 1990-91, and 1995-96. In normal years, the maximal daily catch of elvers occurred around December and January, but around March in the delayed years. This difference is approximately 2 months.
On the other hand, regardless of regular or delayed years, the catch of elvers reached a peak with an interval approximately one month. This was due to the rhythmic activity of elvers in the coastal waters that follows the lunar cycle of new moon. This phenomenon was similar to the previous study (Tzeng, 1985).
2. Delayed arrival of elvers and surface seawater temperature
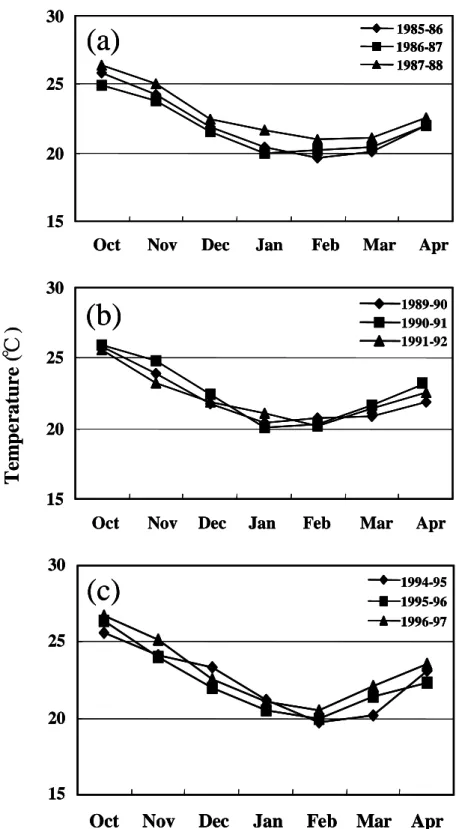
Monthly change of surface seawater temperature off northeastern Taiwan (121- 124°E 25-26°N) for the above mentioned years were shown in Figure 3. In comparison with the seasonal change of the elver catch (Figure 2), it was found that elvers recruited to the estuary in the low surface seawater temperature winter. Furthermore, it was found that the years of delayed arrival always occurred in the low water temperature years, e.g. in 1986, 1990 and 1995 (Figure 4). This indicates that the delayed arrival of elvers was related to surface seawater temperature. 3. Comparison of hatching dates and
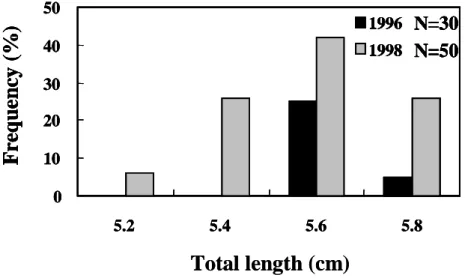
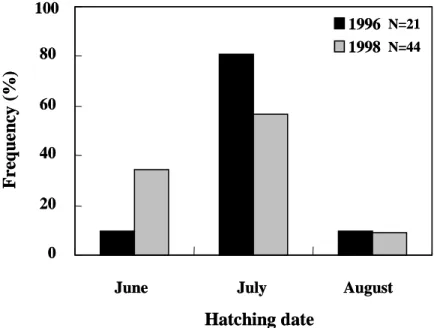
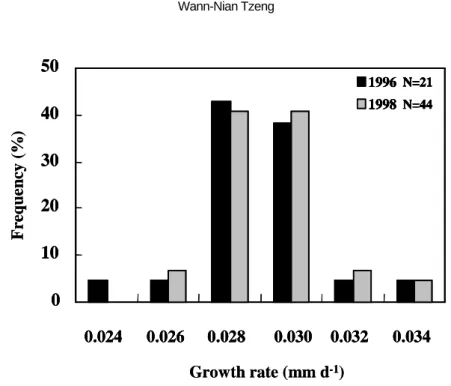
growth rate of elvers between years Of elvers which arrived at estuaries in January, daily ages, hatching dates, and growth rates were all similar between 1996, a delayed arrival year, and 1998, a non-delayed arrival year (Figures 5-8). This indicates that elvers that hatched at the same time will arrive at the estuary simultaneously.
DISCUSSION AND CONCLUSION The phenomenon of delayed arrival of elvers on the coast of Taiwan was found also in Japan (Yanagizawa, 1999; personal communication). The causes of the dely may stem from both biotic and abiotic factors. Biotic factors may involve the changes of spawning location and time. Abiotic factors
No. of E
lvers
0 200 400 600 800 1000 1200 1400 1600 10/1/85 11/1/85 12/1/85 1/1/86 2/1/86 3/1/86 4/1/86 0 500 1000 1500 2000 2500 3000 3500 10/1/86 11/1/86 12/1/86 1/1/87 2/1/87 3/1/87 4/1/87 0 200 400 600 800 1000 1200 1400 1600 1800 10/1/87 11/1/87 12/1/87 1/1/88 2/1/88 3/1/88 4/1/881985-86
1986-87
1987-88
Date (MM/DD/YY)
No. of E
lvers
0 200 400 600 800 1000 1200 1400 1600 10/1/85 11/1/85 12/1/85 1/1/86 2/1/86 3/1/86 4/1/86 0 500 1000 1500 2000 2500 3000 3500 10/1/86 11/1/86 12/1/86 1/1/87 2/1/87 3/1/87 4/1/87 0 200 400 600 800 1000 1200 1400 1600 1800 10/1/87 11/1/87 12/1/87 1/1/88 2/1/88 3/1/88 4/1/881985-86
1986-87
1987-88
Date (MM/DD/YY)
Fig. 2a. Daily catches of elvers in the coastal waters off Shuang-Chi River in 1985~1988(a), 1989~1992(b), and 1994~1998(c).
N
o.
of
E
lve
rs
0 5000 10000 15000 20000 25000 30000 35000 10/1/89 11/1/89 12/1/89 1/1/90 2/1/90 3/1/90 4/1/90 0 500 1000 1500 2000 2500 3000 3500 10/1/90 11/1/90 12/1/90 1/1/91 2/1/91 3/1/91 4/1/91 0 1000 2000 3000 4000 5000 10/1/91 11/1/91 12/1/91 1/1/92 2/1/92 3/1/92 4/1/921989-90
1990-91
1991-92
Date (MM/DD/YY)
N
o.
of
E
lve
rs
0 5000 10000 15000 20000 25000 30000 35000 10/1/89 11/1/89 12/1/89 1/1/90 2/1/90 3/1/90 4/1/90 0 500 1000 1500 2000 2500 3000 3500 10/1/90 11/1/90 12/1/90 1/1/91 2/1/91 3/1/91 4/1/91 0 1000 2000 3000 4000 5000 10/1/91 11/1/91 12/1/91 1/1/92 2/1/92 3/1/92 4/1/921989-90
1990-91
1991-92
N
o.
of
E
lve
rs
0 5000 10000 15000 20000 25000 30000 35000 10/1/89 11/1/89 12/1/89 1/1/90 2/1/90 3/1/90 4/1/90 0 500 1000 1500 2000 2500 3000 3500 10/1/90 11/1/90 12/1/90 1/1/91 2/1/91 3/1/91 4/1/91 0 1000 2000 3000 4000 5000 10/1/91 11/1/91 12/1/91 1/1/92 2/1/92 3/1/92 4/1/921989-90
1990-91
1991-92
0 5000 10000 15000 20000 25000 30000 35000 10/1/89 11/1/89 12/1/89 1/1/90 2/1/90 3/1/90 4/1/90 0 500 1000 1500 2000 2500 3000 3500 10/1/90 11/1/90 12/1/90 1/1/91 2/1/91 3/1/91 4/1/91 0 1000 2000 3000 4000 5000 10/1/91 11/1/91 12/1/91 1/1/92 2/1/92 3/1/92 4/1/921989-90
1990-91
1991-92
Date (MM/DD/YY)
Fig. 2b. Daily catches of elvers in the coastal waters off Shuang-Chi River in 1985~1988(a), 1989~1992(b), and 1994~1998(c).
0 100 200 300 400 500 600 700 800 900 1000 10/1/97 11/1/97 12/1/97 1/1/98 2/1/98 3/1/98 4/1/98 0 100 200 300 400 500 600 700 800 10/1/96 11/1/96 12/1/96 1/1/97 2/1/97 3/1/97 4/1/97 0 2000 4000 6000 8000 10000 12000 10/1/95 11/1/95 12/1/95 1/1/96 2/1/96 3/1/96 4/1/96
N
o
.
o
f
E
lv
er
s
1995-96
1996-97
1997-98
Date (MM/DD/YY)
0 1000 2000 3000 4000 5000 6000 7000 10/1/94 11/1/94 12/1/94 1/1/95 2/1/95 3/1/95 4/1/951994-95
0 100 200 300 400 500 600 700 800 900 1000 10/1/97 11/1/97 12/1/97 1/1/98 2/1/98 3/1/98 4/1/98 0 100 200 300 400 500 600 700 800 10/1/96 11/1/96 12/1/96 1/1/97 2/1/97 3/1/97 4/1/97 0 2000 4000 6000 8000 10000 12000 10/1/95 11/1/95 12/1/95 1/1/96 2/1/96 3/1/96 4/1/96N
o
.
o
f
E
lv
er
s
1995-96
1996-97
1997-98
Date (MM/DD/YY)
0 1000 2000 3000 4000 5000 6000 7000 10/1/94 11/1/94 12/1/94 1/1/95 2/1/95 3/1/95 4/1/951994-95
Fig. 2c. Daily catches of elvers in the coastal waters off Shuang-Chi River in 1985~1988(a), 1989~1992(b), and 1994~1998(c).
1985-86 1986-87 1987-88
(a)
15
20
25
30
Oct Nov Dec Jan Feb
Mar
Apr
1989-90 1990-91 1991-92
(b)
15
20
25
30
Oct Nov Dec Jan Feb
Mar
Apr
℃
Tem
p
e
rat
ure (
)
15
20
25
30
Oct Nov Dec Jan Feb
Mar
Apr
1994-95 1996-97 1995-96
(c)
1985-86 1986-87 1987-88(a)
15
20
25
30
Oct Nov Dec Jan Feb
Mar
Apr
1985-86 1986-87 1987-88 1985-86 1985-86 1986-87 1986-87 1987-88 1987-88
(a)
15
20
25
30
Oct Nov Dec Jan Feb
Mar
Apr
1989-90 1990-91 1991-92
(b)
15
20
25
30
Oct Nov Dec Jan Feb
Mar
Apr
1989-90 1990-91 1991-92
(b)
15
20
25
30
Oct Nov Dec Jan Feb
Mar
Apr
℃
Tem
p
e
rat
ure (
)
15
20
25
30
Oct Nov Dec Jan Feb
Mar
Apr
1994-95 1996-97 1995-96
(c)
15
20
25
30
Oct Nov Dec Jan Feb
Mar
Apr
1994-95 1996-97 1995-96 1994-95 1996-97 1995-96
(c)
Fig. 3. Mean monthly surface seawater temperature in the offshore area of northeastern Taiwan in 1985~ 1988(a), 1989~1992(b) and 1994~1997 (c).
may involve advection of the current that transported the leptocephalus from the spawning ground to the continental shelf, and those that transported glass eel from coastal waters to the estuary.
The Japanese eel spawns in the waters west of Mariana Islands (Tsukamoto, 1992) and its larva (leptocephalus) drifts on the North Equatorial Current to the west and then turns northwards to connect to the Kuroshio Current by Ekman transport (Kimura et al., 1994). A recent study indicated that Ekman transport might change from northward to
southward during El Niño years (Kimura et al., 1999). Meanwhile, the salinity front where the eel spawns may also shift southward during El Niño. These changes in abiotic factors may influence the movement of leptocephalus from the spawning ground to the continental shelf. Both the North Equatorial and Kuroshio Current are warm and highly saline. The decrease of surface seawater temperature in the waters off northeastern Taiwan may be a result of the weakening of the current that transported the leptocephalus and, thus, lead to the delayed arrival of leptocephalus off the
℃
Te
m
p
era
ture ( )
Year
24.0
24.5
25.0
25.5
1 982 1983 1984 1985 1986 1987 1988 1989 1990 1991 1992 1993 1994 1995 1 996 19 9 7 1 998℃
Te
m
p
era
ture ( )
Year
24.0
24.5
25.0
25.5
1 982 1983 1984 1985 1986 1987 1988 1989 1990 1991 1992 1993 1994 1995 1 996 19 9 7 1 998Fig. 4. Mean annual surface seawater temperature in the offshore area of northeastern Taiwan, 1982~1998. Arrows indicate the delayed arrival years of elvers.
F
re
q
ue
nc
y
(
%
)
0 10 20 30 40 50 5.2 5.4 5.6 5.8 1996 1998N=30
N=50
Total length (cm)
F
re
q
ue
nc
y
(
%
)
0 10 20 30 40 50 5.2 5.4 5.6 5.8 1996 1998N=30
N=50
1996 1998N=30
N=50
F
re
q
ue
nc
y
(
%
)
0 10 20 30 40 50 5.2 5.4 5.6 5.8 1996 1998N=30
N=50
Total length (cm)
F
re
q
ue
nc
y
(
%
)
0 10 20 30 40 50 5.2 5.4 5.6 5.8 1996 1998N=30
N=50
1996 1998N=30
N=50
continental shelf of Taiwan.
On the other hand, glass eels were transported by the cold China Coastal Current from the continental shelf to the coast of Taiwan (Tzeng, 1996). The lower mean surface seawater temperature may indicate that the duration of the cold China
Coastal Current in the waters off Taiwan was longer due to the weakening of the Kuroshio, allowing thus the fishing season of elvers to be extended. A previous study indicated that the date of peak catch matched the date of lowest water temperature (Tzeng, 1985). Accordingly, the delayed arrival of elvers on
Freq
u
en
cy
(
%
)
0 10 20 30 40 50 60 70 170 180 190 200 210Age in days
Freq
u
en
cy
(
%
)
0 10 20 30 40 50 60 70 170 180 190 200 210 1996 1998 N=7 N=10Freq
u
en
cy
(
%
)
0 10 20 30 40 50 60 70 170 180 190 200 210Age in days
Freq
u
en
cy
(
%
)
0 10 20 30 40 50 60 70 170 180 190 200 210 1996 1998 N=7 N=10 1996 1998 N=7 N=10Fig. 6. Frequency distribution of daily ages of elvers at estuarine arrival in January of 1996 and 1998.
Hatching date
F
req
ue
ncy
(
%
)
0 20 40 60 80 100June July August
N=21 N=44 1996 1998
Hatching date
F
req
ue
ncy
(
%
)
0 20 40 60 80 100June July August
N=21 N=44 1996 1998 N=21 N=44 1996 1998
the coast of Taiwan was related to the change of current.
A previous study indicated that the early- hatched and fast-growing larvae recruited to the estuary earlier than those that were late- hatched and slow-growing (Tzeng, 1990; Cheng and Tzeng, 1996). To understand if the spawning season and growth rate of eel larvae influence the delayed arrival of elvers, the hatching date distribution and growth rate of the elvers were compared. However, they are similar between delayed (e.g. 1996) and non-delayed arrival years (e.g. 1998). In addition, the age and size of leptocephalus investigated in August of 1995, between their spawning ground and the Philippines, increased from west to east. This indicates that the leptocephalus could drift with the eastward currents, which opposes the normal westward transportation of larvae (Kajihara, 1988; Ozawa et al., 1992; Liao et al., 1996). These results indicate that abiotic factors are the principal cause of delayed arrival of elvers on the coast of Taiwan.
ACKNOWLEDGEMENTS
This study was financially supported by the National Science Council of the Republic
of China (Project No. NSC 89-2611-B002- 04). The authors are grateful to Mr. K. N. Shen for preparing the manuscript, and Miss Jennifer M. Martin and the anonymous reviewer for the helpful comments on the previous manuscript.
REFERENCES
Chen Lee, Y. L., H. Y. Chen and W. N. Tzeng (1994). Reappraisal of the importance of rainfall in affecting catches of Anguilla japonica elvers in Taiwan. Aust. J. Mar. Freshw. Res.
45: 185-190.
Cheng, P. W. and W. N. Tzeng (1996). Timing of metamorphosis and estuarine arrival across the dispersal range of the Japanese eel Anguilla japonica. Mar. Ecol. Prog. Ser. 131: 87-96.
Kajihara, T. (1988). Distribution of Anguilla japonica leptocephali in western Pacific during September 1986. Nippon Suisan Gakkaishi
54: 929-933.
Kimura, S., K. Tsukamoto and T. Sugimoto (1994). A model for the larval migration of the Japanese eel: roles of the trade winds and salinity front. Mar. Biol. 119: 185-190.
Liao I C., C. L. Kuo, W. N. Tzeng, S. T. Hwang, C. L. Wu, C. H. Wang and Y. T. Wang (1996). Growth rate (mm d-1) Fr eq u e n cy ( % )
0
10
20
30
40
50
0.024 0.026 0.028 0.030 0.032 0.034 N=44 N=21 1996 1998 N=44 N=21 1996 1998 Growth rate (mm d-1) Fr eq u e n cy ( % )0
10
20
30
40
50
0.024 0.026 0.028 0.030 0.032 0.0340
10
20
30
40
50
0.024 0.026 0.028 0.030 0.032 0.034 N=44 N=21 1996 1998 N=44 N=21 1996 1998 N=44 N=21 1996 1998 N=44 N=21 1996 1998The first time of leptocephali of Japanese eel Anguilla japonica collected by Taiwanese researchers. J. Taiwan Fish. Res. 4(2): 107- 116.
Ozawa, T., F. Kakizoe, O. Tabeta, T. Maeda and Y. Yuwaki (1992). Larval growth and drift of the Japanese eel Anguilla japonica estimated from leptocephali collection. Nippon Suisan Gakkaishi 58: 15-23.
Sloane,R. D. (1984). Upstream migration by young pigmented freshwater eels (Anguilla australis australis Richardson) in Tasmania. Aust. J. Mar. Freshw. Res. 35: 61-73.
Sorensen, P. W. and M. L. Bianchini (1986). Environmental correlates of the freshwater migration of elvers of American eel in a Rhode Island brook. Trans. Am. Fish. Soc.
115: 258-268.
Tesch, F. W. (1977). The eel: biology and manage- ment of anguillid eels. Translated from German by J. Greenwood. Chapman and Hall/John Wiley & Sons, New York, USA, pp 434. Tosi, L., A. Spampanato, C. Sola and P. Tongiorgi
(1990). Relation of water odour, salinity and temperature to ascent of glass-eels, Anguilla anguilla (L.): a laboratory study. J. Fish Biol. 36: 327-340.
Tsukamoto, K. and A. Umezawa (1990). Early life history and oceanic migration of the eel Anguilla japonica. La Mer 28: 188-198. Tsukamoto, K. (1992). Discovery of the spawning
area for Japanese eel. Nature 356: 789-791. Tzeng, W. N. (1982). Newly record of the elver,
Anguilla celebesensis Kaup, from Taiwan. Bio- Science 19: 57-66. (in Chinese with English abstract)
Tzeng, W. N. (1983). Species identification and commercial catch of the anguillid elvers from Taiwan. China Fisheries Monthly 366: 16-23. (in Chinese with English abstract)
Tzeng, W. N. (1985). Immigration timing and activity rhythms of the eel, Anguilla japonica, elvers in the estuary of northern Taiwan, with emphasis on environment influences. Bull. Jpn. Soc. Fish. Oceanogr. 47/48: 11-28. Tzeng, W. N. (1986). Resources and ecology of
the Japanese eel Anguilla japonica elvers in the coastal waters of Taiwan. China
Fisheries Monthly 404: 19-24. (in Chinese) Tzeng, W. N. (1990). Relationship between growth
rate and age at recruitment of Anguilla japonica elvers in a Taiwan estuary as inferred from otolith growth increments. Mar. Biol. 107: 75- 81.
Tzeng, W. N. (1996). Short- and long-term fluctua- tions in catches of elvers of the Japanese eel Anguilla japonica in Taiwan. In: D. A. Hancok, D. C. Smith, A. Grand and J. P. Beumer (editors) Developing and sustaining world fisheries resources: the state of science and management. 2nd world Fisheries Congress Proceedings, CSIRO publishing, Collingwood, VIC 3006, Australia. 85-89.
Tzeng, W. N. (2002). The process of onshore migration of Japanese eel Anguilla japonica as revealed by otolith microstructure. In: K. Aid, K. Tsukamoto and K. Yamauchi (editors) Advances in Eel Biology. Springer-verlag Tokyo, Japan. 181-190.
Tzeng, W. N. and O. Tabeta (1983). First record of the short-finned eel Anguilla bicolor pacifica from Taiwan. Nippon Suisan Gakkaishi 49: 27-32.
Tzeng, W. N. and Y. C. Tsai. (1994). Changes in otolith microchemistry of the Japanese eel, Anguilla japonica, during its migration from the ocean to the rivers of Taiwan. J. Fish Biol. 45: 671-684.
Tzeng, W. N., K. P. Severin and H. Wickström (1997). Use of otolith microchemistry to inves- tigate the environmental history of European eel Anguilla anguilla. Mar. Ecol. Prog. Ser.
149: 73-81.
Tzeng, W. N., K. P. Severin, H. Wickström and C. H. Wang (1999). Strontium bands in relation to age marks in otolith of European eel Anguilla anguilla. Zool. Stud. 38(4): 452-457.
Tzeng, W. N., H. R. Lin, C. H. Wang and S. N. Xu (2000a). Differences in size and growth rates of male and female migrating Japanese eels in Pearl River, China. J. Fish Biol. 57(5): 1245-1253.
Tzeng, W. N., C. H. Wang, H. Wickström and M. Reizenstein (2000b). Occurrence of the semi- catadromous European eel Anguilla anguilla (L.) in Baltic Sea. Mar. Biol. 137: 93-98.