緒 言
近年來,我國荔枝、龍眼、番荔枝、枇杷、梅、 梨及一些重要經濟果樹經常發生黃化、萎凋、落葉、 生長衰退,甚而急速立枯死亡之情形。依據植物病害 名彙記載( 2 6 ),危害經濟果樹與木本觀賞植物根部之真菌 性病害不下十餘種,其中以褐根病菌 Phellinus noxius (Corner) G. H. Cunningham 為引起目前果樹立枯之最主 要 病 因( 6 , 8 , 9 , 1 0 , 11 , 1 4 , 1 5 , 1 7 , 1 8 ), 其 他 重 要 者 有 疫 病 菌 (P h y t o p h t h o r a s p p . )( 3,4,13,18,19,25)、白紋羽病菌 (R o s e l l i n i a n e c a t r i x P r i l l i l e u x )( 1 8 , 2 2 )、靈芝類 (G a n o d e r m a spp.) ( 5 , 7 , 1 8 , 3 3 )、 斑孔木層孔菌 (Phellinus punctatus ( F r.) Pilat.)( 1 8 )、 K retzchmaria clavus ( F r.) Sacc.(7) 及 Nectria haematococca Berk. et Br.(27) 等。本文將筆者等自 1987 年至 2004 年間 之田間病害調查、病菌分離結果及田間分布情形做一 綜合報告,以明瞭造成台灣木本園藝作物立枯死亡之 真菌病原種類。材料與方法
立枯病害之調查與病菌分離 採集外觀出現立枯死亡與生長衰弱之果樹與觀賞 植物之根系或腐朽之組織,先將表皮剝離,再將腐朽 之罹病木材組織切成 5 × 2 × l mm 之小塊,經 0 . 5 % NaClO 表面消毒 0.5-1 min,放置於含馬鈴薯葡萄糖培 養基 (potato dextrose agar, PDA) 之直徑 9 cm 培養皿 內,每皿四塊組織。每公升 PDA 培養基中含 200g 之 馬鈴薯連皮煎汁液、20g 葡萄糖 (Sigma Co.) 及 15% 洋 菜 (惠昇)。培養基滅菌後並添加 ampicillin (10 ppm) 及 streptomycin sulfate (100 ppm) (鏈黴素),以抑制細菌等 雜菌生長( 6 , 1 0 )。分離用培養皿放置於 2 4℃ 下,等菌絲長 出後,切取菌絲尖端,經純粹培養後於 2 4℃ 下保存於 無菌水中。此外,將病組織密封於塑膠袋內,置於 2 4-台灣果樹及木本觀賞植物立枯型真菌性病害
之調查與分布
安寶貞
1蔡志濃
1 , 3張東柱
2王姻婷
1 1 台中縣 行政院農業委員會農業試驗所 植物病理組 2 台北市 行政院農業委員會林業試驗所 3 聯絡作者,電子郵件信箱: [email protected]. t w;傳真機: 04-23338162 接受日期 : 民國 94 年 7 月 21 日摘 要
安寶貞、蔡志濃、張東柱、王姻婷 . 2005. 台灣果樹及觀賞植物立枯型病害之調查與分布 . 植病 會刊 14: 203-210. 依據 1987 至 2004 年之田間調查與病菌分離結果,與台灣果樹、庭園路樹及觀賞植物立枯 死亡有關的根部病原真菌有十數種,依病菌分離比率,依序為褐根病菌 (Phellinus noxius)、疫 病菌 (P h y t o p h t h o r a s p p .﹐包括 P. parasitica, P. palmivora, P. citro p h t h o r a, P. cinnamomi 及其他)、 靈芝類 (G a n o d e r m a spp., 包括 G. australe, G. tro p i c u m, G. lucidum 及其他)、白紋羽菌 (R o s e l l i n i a n e c a t r i x)、K retzchmaria clavus、斑孔木層孔菌 (Phellinus punctatus)、小孔硬孔菌 (R i g i d o p o ru s m i c ro p o ru s) 及白絹病菌 (S c l e rotium ro l f s i i)。其他另有七種擔子菌 (屬無摺菌目 A p h y l l o p h o r a l e s ) 與五種炭化菌 (屬 Xylariaceae) 亦被分離檢出,但尚未鑑定至種。這些病原菌中,除疫病菌與白 絹病菌外,其餘均為木材腐朽菌 (wood decay fungi),而褐根病菌之檢出率,約佔木材腐朽菌之 50% 左右。2 8℃ 下,1-2 天後,病菌會在病組織上長出濃厚之菌 絲,可用於分辨褐根病菌、白紋羽病菌、靈芝類等。 對於非腐朽性 (non-wood decay) 之罹病組織,則將新鮮 罹病組織切成同樣大小之小塊組織,經消毒後,直接 放置於不添加抗生素之 PDA 平板上,分離可疑病原 菌。 立枯型真菌性病害之鑑定 分離得到之真菌依其形態與子實體特性,至少鑑 定至目為止( 2 , 2 1 , 2 4 )。其中非木材腐朽菌方面,依據分離菌 株特性,歸納成兩類,即 (1) 會產生胞囊與胞囊間接發 芽時,游走子在胞囊內直接分化的特性歸屬於疫病菌 (P h y t o p h t h o r a d e B a r y );(2) 菌絲白色如絹絲,並會形成 球形菌核者歸屬於白絹病菌。在木材腐朽菌方面則歸 納成五類,即 (1) 罹病木材有褐色線紋者 (brown lines or brown pockets) 歸屬褐根病菌或相關種( 2 0 , 2 9 );(2) 罹病 木材長出靈芝子實體,而菌絲有扣子體 ( c l a m p c o n n e c t i o n ),且會在太空包培養基( 8 )上長出靈芝子實體 者歸屬於靈芝( 3 1 );(3) 罹病木材出現黑色線紋且分離之 菌絲先端膨大者歸屬於白紋羽菌( 2 2 , 3 0 );(4) 罹病木材出 現黑色線紋,在太空包培養基上長出灰黑色柱狀菌絲 組 織 , 但 菌 絲 先 端 不 會 膨 大 者 歸 於 炭 化 菌 科 (Xylariaceae) 其他屬( 2 );(5) 而菌絲有扣子體形成,且會 在太空包培養基長出子實體,子實體之腹面具多孔者 歸屬擔子菌中之無摺菌目 ( A p h y l l o p h o r a l e s )( 2 4 )。
結 果
自 1987 年 8 月至 2004 年 12 月,調查我國木本園 藝作物 (包括果樹、路樹及庭院觀賞植物 ) 立枯死亡的 原因,共計採集樣本 234 次,作物 72 種,包括果樹 3 4 種:柑橘 (椪柑、桶柑、柳橙、茂谷柑、文旦、葡萄 柚、四季橘 )、金柑、百香果、酪梨、龍眼、荔枝、 桃、梅、櫻花、梨、枇杷、柿、豆柿、番荔枝 (釋 迦)、鳳梨釋迦、山刺番荔枝、楊桃、葡萄、蓮霧、番 石榴、蘋婆、掌葉蘋婆、馬拉巴栗、澳洲胡桃、波羅 蜜、破布子、愛玉子、洛神葵,及木本觀賞植物 3 8 種:月橘、朱槿、杜鵑、西洋杜鵑、金露花、矮仙 丹、含笑花、變葉木、羊蹄甲、洋紫荊、艷紫荊、阿 勃勒、黃金風鈴木、紫花風鈴木、大花紫薇、美人 樹、莿桐、櫸木、盾柱木、臘腸樹、瓊崖海棠、巴西 胡椒木、欖仁、小葉欖仁、垂榕、竹葉榕、樟樹、木 麻黃、相思樹、白千層、茄冬、桉樹、銀樺、菩提 樹、肯氏南洋杉、蘇鐵、美葉蘇鐵、大王椰子。分離 結果如圖一與表一所示,病原真菌出現與分離頻率由 高向低依序為褐根病菌 (P. noxius ( 11 2次) 與 P. puntatus ( 3次) )、疫病菌 (P h y t o p h t h o r a s p p . ) ( 5 9次)、靈芝屬 (G a n o d e r m a spp.) (36次)、白紋羽菌 (R. necatrix) ( 1 5 次)、(K. clavus) ( 6次)、小孔硬孔菌 (R. micro p o ro u s) ( 3 次),白絹菌 (S. ro l f s i i) ( 2次) 及 P e re n n i p o r i a s p . ( 1次)。 其他另有七種擔子菌 (屬無摺菌目 Aphyllophorales) 與 五種炭化菌 (屬 Xylariaceae) 亦被分離檢出,但尚未鑑 定至種。這些與立枯病害有關之真菌中,疫病菌與白 絹病菌為非木材腐朽菌,雖然會引起植物生長衰落與 植株死亡,但不會造成罹病根系與木材腐朽;其餘褐 根病菌、白紋羽菌、靈芝、炭化菌均屬木材腐朽真菌 (wood-decay fungi),罹病植株的木材最後均會白腐 (white rotted),甚而嚴重崩潰 ( c o l l a p s e )。 圖一、台灣地區木本果樹與觀賞植物由木材腐朽菌引 起之立枯型病害的調查與分布Fig. 1.Distribution of wood-decay fungi causing fruit and ornamental tree decline in Taiwan (1987-2004)
實驗期間,疫病菌自多種罹患立枯型果樹之腐敗 根系與莖基部分離得到,包括 Phytophthora parasitica Dastur 分離自柑橘 (含文旦)、枇杷、百香果、番石榴、 洛神葵、朱槿及變葉木;P. cinnamomi Rands 分離自根 系 腐 敗 而 出現 立枯 衰 弱 的 酪梨 、 杜 鵑 及蘇 鐵; P. palmivora Butler (Butler) 亦自罹病之柑橘與酪梨根系 獲得;P. citro p h t h o r a Leonian 可引起金柑與西洋杜鵑的 快速死亡。柑橘被疫病菌侵染樹幹後,一般會引起流 膠,罹病組織褐變,病菌主要侵染樹幹表皮與形成 層;罹病植株外觀出現枝葉黃化、生長衰弱等慢性立 枯的現象,嚴重時亦偶會急速萎凋死亡。酪梨一般根 部被感染而嚴重腐敗,植株出現衰敗 (decline) 現象, 但颱風豪雨過後亦會出現急速萎凋,植株於短時間內 ( 1 - 3月) 大量死亡的的情形。枇杷、百香果及落神葵則 為主根與主幹被疫病菌侵染,植株出現基腐病 ( b a s a l stem rot)、頸腐 (neck rot)、領腐 (collar rot)、裾腐 ( f o o t
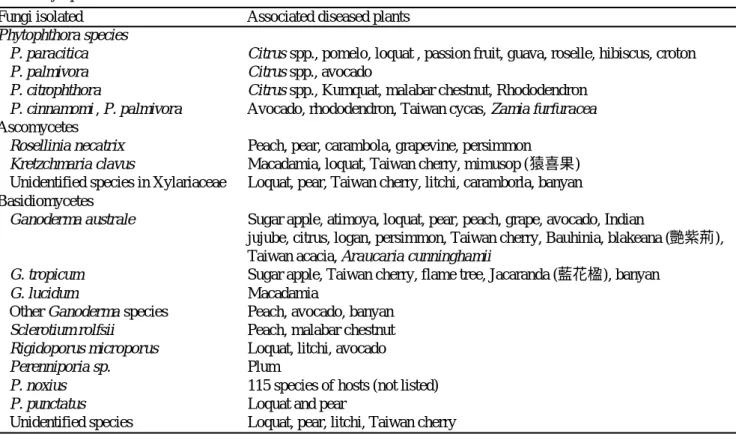
rot) 等病徵,罹病植株樹幹基部褐變,組織崩潰,植株 快速萎凋死亡,在潮溼的環境下病組織上會長出白色 的黴狀物,為其菌絲。 白絹病菌 S. ro l f s i i Sacc. 則兩次自枯萎之馬拉巴栗 (南投仁愛 ) 與桃樹 (台中霧峰 ) 根部與莖基部分離得 到,罹病部位之表面亦長出大量之白色密緻菌絲與褐 色圓球型菌核 (尚未發表與完成接種試驗 ),甚易鑑定 病菌。 在引起立枯型病害之木材腐朽菌方面,主要分離 到的為擔子菌與子囊菌,其中以擔子菌中之褐根病菌 (P. noxius) 之檢出率最高,它危害 21 種果樹與 46 種觀 賞植物,是台灣木本植物立枯死亡的主因,主要分佈 在海拔 1000 公尺以下之地區,台灣各地與金門地區均 有蹤跡,其寄主範圍( 6 , 8 , 1 0 , 11 )與苗木對其之抗病性檢定 ( 1 2 ) 等業於多種期刊發表,近兩年 (2003-2004) 發現之新紀 錄寄主有七種,包括酪梨 (Persea americana)、柳橙 廣東檸檬 (C i t rus limon )、小葉欖仁 (Te r m i n a l i a m a n t a l y )、 盾 柱 木 (P e l t o p h o rum inerme )、 莿 桐 (E rythrina variegata)、竹葉榕 (Ficus stenopylla) 及巴西 胡椒木 (Schinus tere b i n t h i f o l i n u s) 等。而斑孔木層孔菌 (P. punctatus) 則自枇杷 (台中新社) 與梨 (嘉義竹崎) 分 離得到 (未發表,已完成接種試驗 )。褐根病與斑孔木 層孔菌引起的病徵稍異,褐根病之褐色菌絲束在腐朽 木材上形成之網紋較大,而後者較小與密緻,兩者都 會造成木材 (心材與邊材) 完全腐朽。 靈芝亦為引起立枯型病害的重要病原菌之一,它 自罹病之澳洲胡桃、番荔枝、枇杷、山櫻花、葡萄、 印度棗、梨、酪梨、柑橘、龍眼、桃、鳳凰木、藍花 楹、榕樹、鳳凰木、艷紫荊、相思樹、南洋杉分離得 到,部份菌株對寄主植物的病原性檢定業已完成,包 表一、1987 至 2004 年台灣地區木本果樹與觀賞植物立枯型真菌病害的之分離情形
Table 1. Isolation of fungi from stem and/or root systems of fruit trees and ornamental woody plants associated with decline symptoms from 1987 to 2004 in Ta i w a n
Fungi isolated Associated diseased plants Phytophthora species
P. paracitica C i t rus spp., pomelo, loquat , passion fruit, guava, roselle, hibiscus, croton
P. palmivora C i t ru s spp., avocado
P. citro p h t h o r a C i t ru s spp., Kumquat, malabar chestnut, Rhododendron P. cinnamomi , P. palmivora Avocado, rhododendron, Taiwan cycas, Zamia furfuracea A s c o m y c e t e s
Rosellinia necatrix Peach, pear, carambola, grapevine, persimmon K retzchmaria clavus Macadamia, loquat, Taiwan cherry, mimusop (猿喜果) Unidentified species in Xylariaceae Loquat, pear, Taiwan cherry, litchi, caramborla, banyan B a s i d i o m y c e t e s
Ganoderma australe Sugar apple, atimoya, loquat, pear, peach, grape, avocado, Indian
jujube, citrus, logan, persimmon, Taiwan cherry, Bauhinia, blakeana (艷紫荊), Taiwan acacia, Araucaria cunninghamii
G. tro p i c u m Sugar apple, Taiwan cherry, flame tree, Jacaranda (藍花楹), banyan
G. lucidum Macadamia
Other Ganoderma s p e c i e s Peach, avocado, banyan S c l e rotium ro l f s i i Peach, malabar chestnut R i g i d o p o rus micro p o ru s Loquat, litchi, avocado
P e renniporia sp. P l u m
P. noxius 115 species of hosts (not listed)
P. punctatus Loquat and pear
括澳洲胡桃、枇杷、梨、番荔枝、山櫻花等,由於為 台灣新紀錄,將另發表。在雨季,一般被靈芝侵染的 罹病植株的樹幹基部與根系均會長出子實體,較易鑑 定至科的分類地位,分離最多的靈芝為南方靈芝 (Ganoderma australe ( F r,) Pat.)(19 次,分別自山櫻花、 柑橘、枇杷、番荔枝、鳳梨釋迦、桃、葡萄、酪梨、 印度棗、艷紫荊、相思樹、南洋杉分離得到 )、其次為 熱帶靈芝 (G. tro p i c u m (Jungh) Bres.)(7 次,分別分離自 番荔枝、山櫻花、鳳凰木、藍花楹、榕樹 )、再其次為 靈芝 (G. lucidum ( C u r t . : F r. ) Karst)(3 次,澳洲胡桃與鳳 梨釋迦),尚有數種未能鑑定者。寄主植物最常見的為 番荔枝、山櫻花、鳳凰木及相思樹等;罹病植株在出 現生長衰弱現象與靈芝子實體後,約 2-3 年後才會枯 死,甚而有些樹種僅出現生育緩慢情形,並不會枯 死;罹病植株會枯死者包括澳洲胡桃、番荔枝、枇 杷、山櫻花、葡萄、印度棗、梨、酪梨、鳳凰木、藍 花 楹 及 相 思 樹 。 尚 有 一 些 屬 於 無 摺 菌 目 (Aphyllophorales) 之擔子菌,從枇杷、荔枝、梨、桃、 李、山櫻花、酪梨分離得到,其菌絲大部分具有扣子 體 (clamp connection) 形成,但子實體為非靈芝之多孔 菌,且多數均已完成病原性檢定。其中完成初步鑑定 者包括 R i g i d o p o rus micro p o ru s (Klotzsch) Imazeki 分離 自枇杷、荔枝及酪梨的腐敗根系,其菌絲不具扣子 體;而 P e re n n i p o r i a sp. 分離自李樹根部 (將另文發 表)。 在 木 材 腐 朽 菌 部 份 , 子 囊 菌 中 以 炭 化 菌 科 (Xylariaceae) 分離到的比率最高。其中,白紋羽病菌 R . n e c a t r i x 自七種罹病作物分離得到,為腐朽菌中分離頻 率第三位,包括梨、桃、楊桃、葡萄、柿及火龍珠, 而罹病寄主多發現在台灣之中北部地區,且為海拔 1000 公尺以上 (圖一)。此外,K. clavus 自澳洲胡桃、 荔枝、山櫻花、猿喜果等四種植物之罹病根系組織檢 出。這些病菌,除火龍珠、圓喜果分離之菌株外,均 已完成病原性測定,其中危害澳洲胡桃已經發表( 7 ),其 餘尚未正式發表。此外,尚有一些炭化菌科中之真菌 自澳洲胡桃、枇杷、山櫻花、桃、番荔枝、梨、酪梨 等罹病植物分得,其中危害山櫻花、桃、番荔枝、梨 的菌株已完成病原性檢定,但尚未完成病原菌鑑定, 將另文發表。
討 論
依據文獻記載( 2 ),引起木本植物出現立枯病徵的病 原真菌有很多種,以是否會造成罹病木材腐敗 ( d e c a y & collapse),可區分為木材腐朽菌 (wood decay & rotting fungi) 與其他真菌。木材腐朽菌主要為擔子菌與 子囊菌,擔子菌中以無摺菌目 ( A p h y l l o p h o r a l e s ) (包括 Ganoderma, Phellinus 等) 與傘菌目 ( A g a r i c a l e s ) (以 Armillaria 為主 ) 最為重要;而子囊菌中則以炭化菌 (xylariacious fungi) 為主,如白紋羽菌、 K re t z c h m a r i a spp. 及 X y l a r i a spp. 等。在非木材腐朽菌中,則包括疫 病菌、鐮孢菌、Ceratocystis spp. 等。 在台灣,依據植物病害名彙記載( 2 6 )與本次試驗之調 查結果,顯示與果樹與木本觀賞植物立枯死亡有關的 真菌約有十數種,其中以褐根病菌 (Phellinus noxius)、 疫病菌 (P h y t o p h t h o r a s p p . )、白紋羽病菌 (R o s e l l i n i a n e c a t r i x) 及靈芝類 (G a n o d e r m a spp.) 最為普遍與重要。 本試驗分離之相關真菌大部分為木材腐朽菌,包括褐 根病菌、白紋羽菌、靈芝、炭化菌及多種擔子菌 (無摺 菌目 ) 等,均會造成寄主植物罹病組織之腐朽崩潰 (decay & collapse);而經常分到的另一真菌疫病菌則非 屬木材腐朽病菌,罹病寄主植物經常快速萎凋死亡, 但被感染之木材不會出現腐朽的現象。這些分離菌種 與國外危害樹木的病菌不盡相似,褐根病菌在熱帶與 亞熱帶較為普遍( 3 1 ),靈芝則依種分布於熱帶、亞熱帶與 溫帶地區( 3 1 );白紋羽菌分布於溫帶地區,在歐美、日本 均十分普遍( 3 0 );疫病菌之分布則為世界性( 2 3 )。然而,在 溫帶地區危害果樹與森林最嚴重的病菌 (A r m i l l a r i a s p p . )(2) 在本試驗中並未被檢出。 褐根病菌、靈芝、白紋羽病菌、及 K. clavus 等木 材腐朽菌,其引起植株之立枯病之外觀病徵很相似, 不易區別,但造成之根部病徵則有明顯差異,褐根病 菌危害會有褐色菌絲塊及褐色網紋狀之菌絲束,感染 靈芝之植株在降雨季節於樹幹基部會長出靈芝之子實 體,而其炭化菌引起之腐敗木材會出現黑色線紋,因 此 本 文 設計 一 簡 單 鑑別 這些 根 部 病 菌之 檢索表 ( k e y s ) (表四)。以目前台灣病害發生情形,此後三者皆 不如褐根病普遍與嚴重,目前 P. noxius 已成為我國果 樹與樹木根部的最重要病原菌之一。P. noxius 之寄主範 圍與分佈十分廣泛,分佈於澳洲、紐西蘭、非洲、中 南美洲、北太平洋及亞洲等熱帶與亞熱帶地區( 1 , 2 0 , 2 1 , 2 8 ), 寄主作物超過 200 種以上,而且,不同寄主分得之褐 根病病原菌可以互相感染其他寄主,尚未發現菌株間 有明顯寄主分化之現象,但是菌株間之毒力 ( v i r u l e n c e ) 略有強弱差異,尤其菌株對其分離之寄主之毒力似乎 較強( 2 8 )。在其他研究報告方面,發現土壤浸水為一經濟 且有效的病害防治方法,淹水一個月可殺死土壤中存 活於木材之 P. noxius( 1 6 ),但是有很多褐根病發生在山坡 地,無法實施浸水防治,因此,病害還是以藥劑防治 為主,且已在實驗室篩選出多種有效藥劑 ( 3 2 )。因 P. n o x i u s在台灣不易產生子實體( 6 ),藉由空氣傳播擔孢表二、台灣地區木本果樹與觀賞植物立枯型真菌性病害之調查與比率
Table 2. Survey of fruit and ornamental tree decline caused by fungi in Taiwan (1987-2004) P a t h o g e n No. of diseased samples F r e q u e n c y ( % ) R e m a r k s
P h y t o p h t h o r a spp. 5 9 2 3 . 0 Slow & quick decline, with stem gummosis in c i t r u s
Phellinus noxius 11 2 43.8 (57.4) 1 Below sea level 1000 m.
Slow & quick decline or sudden wilt
P. punctatus 3 1 . 2 ( 1 . 5 ) Below sea level 1000 m.
Slow & quick decline or sudden wilt G a n o d e r m a s p p . 3 6 1 4 . 0 ( 1 8 . 5 ) Slow decline
R i g i d o p o rus micro p o ru s 3 1 . 2 ( 1 . 5 ) Quick decline
S c l e rotium ro l f s i i 2 0 . 8 Quick wilt
Other basidiomycete fungi 1 2 4.7 (6.2) Slow or quick decline
Rosellinia necatrix 1 5 5.8 (7.7) At high land, Slow & quick decline
K retzchmaria clavus 5 2.0 (2.6) Slow decline
Other xylariaceae fungi 9 3 . 5 ( 4 . 6 ) Slow decline mostly
To t a l 256 (195)
1 Figures in parenthesis indicated that only wood decay fungi were counted.
表三、褐根病菌與白紋羽病菌特性與引起病害之比較
Table 3.Comparision of the pathogen characteristics and diseases caused by Phellinus noxius and Rosellinia necatrix I t e m Phellinus noxius Rosellinia necatrix
C l a s s i f i c a t i o n B a s i d i o m y c e t e s A s c o m y c e t e s Brown root rot fungus Black root rot fungus
C o l o n y White initially, turn brown or dark White initially, turn gray and black after mature brown after mature
Growth temperature Ranges: 8-36℃, optimum 24-33℃ Ranges :8-30℃, optimum 20-28℃ Mycelium Clamp connection absent, with arthrospores Swollen at the hyphal base
and trichocyst hyphae
Symptoms on diseased 1.brown mycelial mats sticking on diseased 1.white thick mycelia turning gray slightly root with soil particles
t i s s u e s 2.brown mycelial match and pocket 2.iregullar black line (caused by interaction of host and pathogen)
Type of wood rot White rot White rot
D i s t r i b u t i o n Island wide, mainly at central and southern . Cool places (Central and northern Ta i w a n ) Taiwan under sea level 1000 cm
表四、台灣果樹與木本觀賞植物真菌性立枯型病害之分類檢索
Table 4. A key for identification of fungi associated with decline of woody fruit and ornamental trees in Ta i w a n
1. Decay and decomposed of infected tissues (2) 1. Not decomposed (6)
2.Gray to black lines on diseased tissues, with thick white to gray mycelial mats (xylariaceae fungi) (3) 2. Irregular or pocketed brown lines on diseased tissues, with brown mycelial mats (P h e l l i n u s spp.) (4) 2. Mycelial mats with other than described above (other basidiomycetes) (5)
3. Mycelia growing very slow, linear grow less than 2 mm per day……… K retzchmaria clavus
3. Mycelia growing much faster, swollen on hyphal base……… Rosellinia necatrix
3. Mycelia growing much faster, without swollen on hyphal base……… Other xylariaceae fungi
4. Linear mycelia growth > 5 mm per day, diseased wood with brown pocket. > 1cm in diam
……… Phellinus noxius
4. Linear mycelia growth < 5 mm per day, with small brown pocket < 1cm in diam………… Phellinus punctatus
5.Fruiting bodies of Ganoderma formed in natural or on sow dust media………G a n o d e r m a s p p .
5.Fruiting bodies of other types with caps, mycelia with clamp connection or not………other basidiomycet fungi
5.No fruiting bodies, mycelia with clamp connection ……… other basidiomycete fungi
6. Silky white mycelial mats and round sclerotia formed around diseased tissues………S c l e rotium ro l f s i i
6. White mycelial mats and sporangia of P h y t o p h t h o r a formed under moist condition…………P h y t o p h t h o r a s p p .
子之風險降低,但必須注意種苗帶菌及土壤帶菌與 否。褐根病之防治要領,預防重於治療,一旦植株出 現病徵,根部已有 60-70 % 以上受害,不容易根治病 害。因此,發展偵測技術相當重要,若能在病徵尚未 出現之前提早偵測出病原,必能達到早期防治的效 果;而在病害防治方面,冀望開發更有效之病害防治 策略與方法。
謝 辭
本文部份經費承蒙行政院農業委員會與動植物防 疫檢疫局計畫支持,及柯文雄教授修正英文,謹此致 謝。引用文獻 (LITERATURE CITED)
1 . Abe, Y., Kobayashi, T., Onuki, M., Hattori, T., andTsurumachi, M. 1995. Brown root rot of trees caused by Phellinus noxius in windbreaks on Ishigaki Island, Japan – Incidence of disease, pathogen and artificial inoculation. Ann. Phytopathol. Soc. Jpn. 61:425-433. 2 . Alexopoulos, C. J., Mims, C. W., and Blackwell,
M.1996. Introductory Mycology. 4t hed. John Wiley & Sons, Inc. New York. 869 pp.
3 . Ann, P. J. 1984. Species, mating types and pathogenicity of P h y t o p h t h o r a distributed in citrus orchards in Taiwan. Trans. Br. Mycol. Soc. 82:631-6 3 4 .
4 . Ann, P. J. 1995. Soil-borne fungal diseases of citrus. TARI special publication 51: 209-219. (in Chinese with English abstract)
5 . Ann, P. J., Chang, T. C., Tsai, J. N., and Wang, I. T. 2000. Decline of fruit trees associated with root rot caused by G a n o d e r m a species in Taiwan. Plant Pathol. Bull. 9:178. (English abstract)
6 . Ann, P. J., Chang, T. T., and Ko, W. H. 2002. P h e l l i n u s n o x i u s brown root rot of fruit and ornamental trees in Taiwan. Plant Dis. 86: 820-826.
7 . Ann, P. J., and Ko, W. H. 1988. Root rot of macadamia caused by Ganoderma lucidum and K re t z c h m a r i a c l a v u s in Taiwan. J. Agric. Res. China. 37:424-429. 8 . Ann P. J., and Ko, W. H.1992. Longan decline:
association with brown root rot caused by P h e l l i n u s n o x i u s . Plant Pathol.Bull.1:19-25.
9 . Ann, P. J., and Ko, W. H. 1994. Studies on ecology of brown root rot of fruit trees caused by Phellinus noxius and disease control. Plant Pathol. Bull. 3:69. (Abstract) 1 0 . Ann P. J., Lee, H. L., and Huang, T. C.1999. Brown root rot of ten fruit trees caused by Phellinus noxius i n Taiwan. Plant Dis.83:746-750.
11 . Ann, P. J., Lee, H. L., and Tsai, J. N. 1999. Survey of brown root disease of fruit and ornamental trees caused by Phellinus noxius in Taiwan. Plant Pathol. Bull. 8:51-60. (in Chinese with English abstract) 1 2 . Ann, P. J., Tsai, J. N., Wang, I. T., and Hsien M. L.
1999. Responses of fruit trees and ornamental plants to brown root rot disease by artificial inoculation with Phellinus noxius . Plant Pathol. Bull. 8:61-66. (in Chinese with English abstract)
1 3 . Ann, P. J., Wu, I. F., and Lin, C. Y. 1997. Studies on etiology of quick decline and death of Jingkang in Ilan areas and the disease control. Plant Pathol. Bull.6:198. (English abstract).
1 4 . Chang, T. T. 1992. Decline of some forest trees associated with brown root rot caused by P h e l l i n u s n o x i u s. Plant Pathol. Bull.1:90-95.
1 5 . Chang, T. T. 1995. Decline of nine tree species associated with brown rot root caused by P h e l l i n u s n o x i u s in Taiwan. Plant Dis. 79:962-965.
1 6 . Chang, T. T. 1996. Survival of Phellinus noxius in soil and in the roots of dead host plants. Phytopathology 8 6 : 2 7 2 - 2 7 6 .
1 7 . Chang, T. T., and Yang, W. W. 1998. Phellinus noxius in Taiwan: distribution, host plants and the pH and texture of the rhizosphere soils of infected hosts. Mycol. Res. 102:1085-1088.
1 8 . Chang, T. T., Hsieh, H. J., Chang, R. J., and Fu, C. S. 1999. Common Tree Diseases in Taiwan. Ta i w a n Forestry Research Institute. Taipei. 202 pp.
1 9 . Chern, L. L., Ann, P. J., Young, H. R. 1998. Foot and root rot of loquat in Taiwan caused by P h y t o p h t h o r a . Plant Dis 82:651-656.
2 0 . C o r n e r, E. J. H. 1932. The identification of the brown root fungus. Gard. Bull. Straits Settl. 5:317-350. 2 1 . Cunningham, G. H. 1965. Polyporaceae of New
Zealand. N. Z. Dept. Sci. Indust. Res. Bull. 164:221-2 164:221-2 164:221-2 .
2 2 . D u an , C. H. , T sa i , W . H ., a nd T u , C. C . 1 990 . Dissemination of white root rot disease of loquate and its control. Jour. Agric. Res. China 39:47-54. (in Chinese, with English abstract)
2 3 . Erwin, D., and Ribeiro, O. 1996. P h y t o p h t h o r a Diseases Worldwide. APS press. Minnesota. 562 pp. 2 4 . F l o r a n c e , E . R . , a n d S h a w , C . G . 1 9 8 8 . S u r f a c e
morphology of basidiospores from decay fungi that are common in Pacific Northwest forest U.S.A.. Northwest S.C.I.5:547-550.
2 5 . Ho, H. H., Ann, P. J., and Chang, H. S. 1995. The Genus P h y t o p h t h o r a in Taiwan. Acad. Sin. Mon. Ser. 15. Taipei, Taiwan, ROC. 86 pp.
2 6 . Hsu, S. T. et al e d i t o r. 2002. List of plant diseases in Taiwan. Taiwan Phytopathological Society. Ta i c h u n g .
386 pp. (in Chinese).
2 7 . Lin, Y. S., & Chang, H. L. 1982. Root rot and collar rot of passion fruit. Plant Prot. Bull. 24:275. (Chinese a b s t r a c t )
2 8 . Nandrid, D., Nicole, M., and Geiger, J. P. 1987. Variation in virulence among R i g i d o p o rus lignosus a n d Phellinus noxius isolates from West Africa. Eur. J. For. P a t h o l . 1 7 : 2 7 1 . 2 8 1 .
2 9 . P e g l e r, D. N., and Waterston, J. M. 1968. P h e l l i n u s n o x i u s. No. 195 in: Descriptions of Pathogenic Fungi and Bacteria. Commonw. Mycol. Inst., Kew, England. 3 0 . S i n c l a i r, W. A., Lyon, H. H., and Johson, W. T. 1987.
Diseases of trees and shrubs. Cor. Uniu. Press. Ithaca, N. Y.
3 1 . Sivanesan, A., and Holliday, P. 1972. R o s e l l i n i a n e c a t r i x. No. 352 in: Descriptions of Pathogenic Fungi and Bacteria. Commonw. Mycol. Inst., Kew, England. 3 2 . Tsai. J. N., Ann, P. J., and Hsieh, W. H. 2005.
Evaluation of fungicides for suppression of three major wood-decay fungi Phellinus noxius, R o s e l l i n i a n e c a t r i x and Ganoderma australe in Taiwan. Plant Pathol. Bull. 14 (printed)(in Chinese with English a b s t r a c t )
3 3 . Wang, Z. N. 1990. Three species of G a n o d e r m a pathogenic to macadamia tree and their control. R e p . Taiwan Sugar Res. Inst. 129:1-10.
A B S T R A C T
Ann, P. J. 1, Tsai. J. N. 1 , 3, Chang, T. T.2, and Wang, I. T. 12005. Distribution of decline of fruit trees and ornamental woody plants in Taiwan. Plant Pathol. Bull.14:203-210. (1 Plant Pathology Division, Agricultural Research Institute, Wufeng, Taichung, Taiwan; 2 Plant Protection Division, Forestry Research Institute, Taipei; 3 corresponding author, E-mail: [email protected]; Fax No.: +886-4 - 2 3 3 3 8 1 6 2 )
Species of important economic woody fruit trees and ornamental trees with decline symptoms were investigated from 1987 through 2004 in Taiwan. Based on the survey on the fields and pathogen isolation, more than ten species of root- and wood-infecting fungi were founded to be associated with the decline of these ornamental woody plants. According to the isolation frequency in descending o r d e r, there are Phellinus noxius , P h y t o p h t h o r a spp.(including P. parasitica , P. palmivora , P. citro p h t h o r a, P. cinnamomi and others) , G a n o d e r m a spp.(including G. australe, G. tro p i c u m, G. lucidum and others), Rosellinia necatrix , K retzchmaria clavus , P. punctatus, R i g i d o p o ru s m i c ro p o ru s and S c l e rotium ro l f s i i. Besides, a total of 12 species of unidentified fungi including 7 species in Aphllophorales and 5 species in Xylariaceae were also isolated. Most of these root-infected fungi belonged to wood decay fungi in Ascomycetes or Basidiomycetes, except P h y t o p h t h o r a spp. and S. ro l f s i i. Among them, P. noxius, which was detected from more than 50% wood decay samples, is the most destructive and widely distributed.