ORIGINAL RESEARCH
A systematic review of microRNA expression profiling
studies in human gastric cancer
Sirjana Shrestha1,a, Sheng-Da Hsu2,3,a, Wei-Yun Huang1, Hsi-Yuan Huang2,3, WenLiang Chen1,
Shun-Long Weng1,4,5,6& Hsien-Da Huang1,2,3,7
1Department of Biological Science and Technology, National Chiao Tung University, Hsinchu 300, Taiwan 2Institute of Bioinformatics and Systems Biology, National Chiao Tung University, Hsinchu 300, Taiwan 3Center for Bioinformatics Research, National Chiao Tung University, Hsinchu 300, Taiwan
4Department of Obstetrics and Gynecology, Hsinchu Mackay Memorial Hospital, Hsinchu 300, Taiwan 5Mackay Medicine, Nursing and Management College, Taipei 112, Taiwan
6Department of Medicine, Mackay Medical College, New Taipei City 252, Taiwan
7Department of Biomedical Science and Environmental Biology, Kaohsiung Medical University, Kaohsiung 807, Taiwan
Keywords
Biomarker, gastric cancer, microRNA, miRNA target genes
Correspondence
Hsien-Da Huang, Department of Biological Science and Technology, Institute of Bioinformatics and Systems Biology, Center for Bioinformatics Research, National Chiao Tung University, Hsinchu 300, Taiwan, ROC & Department of Biomedical Science and Environmental Biology, Kaohsiung Medical University, Kaohsiung 807, Taiwan, ROC. Tel: +886-3-5712121 (x) 56952; Fax: +886-3-5729288; E-mail: [email protected] Funding Information
This manuscript was supported by the National Science Council of the Republic of China [Contracts NSC 101-2311-B-009-003-MY3 and NSC 102-2911-I-009-101] and the Veterans General Hospitals and University System of Taiwan (VGHUST) [Grant VGHUST103-G5-1-2].
Received: 30 November 2013; Revised: 14 February 2014; Accepted: 13 March 2014 Cancer Medicine 2014; 3(4): 878–888 doi: 10.1002/cam4.246
a
These authors contributed equally to this work.
Abstract
Gastric cancer (GC) is the second leading cause of global cancer mortality. Most GC patients are diagnosed with advanced-stage disease and show extre-mely poor prognosis. All of the GC research has a common interest to search for the specific and sensitive biomarkers for early diagnosis of GC. Number of microRNAs play important role in GC. We carried out a system-atic review of published miRNA profiling studies that compared the miRNA expression profiles between GC tissues and paired noncancerous gastric tis-sue. A vote-counting strategy was followed with the collection of information like total number of studies reporting differential expression of miRNA, total number of tissue samples used in the studies, direction of differential expres-sion and fold change. A total of 352 differentially expressed microRNAs were reported in the 14 microRNA expression profiling studies that compared GC tissues with normal tissues with 120 microRNAs reported at least in two studies. In the group of consistently reported microRNAs, miR-21 was reported upregulated in 10 studies followed by 25, 92, and miR-223 upregulated in eight studies. MiR-375 and miR-148a were found down-regulated in six and five studies, respectively, followed by miR-638 in four studies. MiR-107 and miR-103 were reported in nine and eight studies, respectively, but their expression were inconsistent. From this study, the most consistently reported upregulated microRNA was found to be miR-21. This systematic review study of human GC microRNA expression profiling studies would provide information on microRNAs with potential role as the biomar-kers in gastric cancer.
Cancer Medicine
Introduction
Gastric cancer (GC) is the second leading cause of can-cer-related death worldwide. About one million new cases per year of GC were estimated to have occurred [1]. GC is diagnosed at advanced stage accompanied by extensive invasion and lymphatic metastasis [2]. Thus, early detec-tion of GC is a key measure to reduce the mortality and improve the prognosis of gastric cancer. Recent studies on diagnostic techniques and peri-operative management have increased the early detection of GC and decreased the mortality [3, 4]. Therefore, it is very important to increase the sensitivity and specificity of diagnostic markers and/or methods for the treatment and prevention of GC [5].
MicroRNAs are a class of ~22 nucleotides noncoding RNAs that regulate the expression of target genes by interacting with complementary sites in the 30 untrans-lated region of the target mRNAs [6]. Studies have shown that miRNAs have been involved in the regula-tion of different biological processes, including apoptosis, proliferation, metabolism, cellular differentiation, embryogenesis, gene regulation [7, 8]. Recently microR-NA expression profiling has been extensively used in order to screen the expression of the large number of microRNAs through extensive sample collections. Several microRNA expression profiling studies in cell lines, tis-sue samples, serum have revealed the number of mi-croRNAs as the biomarkers in GC [9–11]. These studies provide hundreds of differentially expressed miRNAs, and finally only a small number of them may have clini-cal use and act as diagnostic and prognostic biomarkers. Different profiling studies show inconsistency in the identified differentially expressed miRNAs. A logical method to identify the most consistently reported differ-entially expressed miRNAs is to search for the intersec-tions of miRNAs identified in multiple independent studies. Griffith [12] and Chan [13] vote-counting strat-egy have cope with the challenges to collect and com-bine the results of those miRNAs expression profiling studies with different profiling platform, different meth-ods to ascertain differential expression like normalization and significance level.
We carried out a systematic review from the published microRNAs expression profiling studies that compared the miRNA expression profiles in GC tissues with those in noncancerous gastric tissues. A vote-counting strategy was used which considered the total number of studies that reported differential expression, total number of tis-sue samples used in studies and the average fold change. We also presented the consistently reported differential miRNAs and the rank of the differentially expressed upregulated and downregulated miRNAs.
Materials and Methods
Literature survey
In order to find all relevant literatures, we used PubMed search engine for the GC microRNA expression profiling studies between the periods February 2006 and December 2012 [14–27]. Search term “miR and gastric cancer” was used. Papers with miRNA expression profiling studies that used tissue samples obtained from surgically resected GC tumors and corresponding noncancerous gastric tissues were selected.
Study selection criteria
Selected study should meet the criteria: (1) they were miRNA expression profiling studies in GC patients, (2) they used GC and neighboring noncancerous gastric tis-sues for comparison, (3) use of microarray methods, (4) had information on cut off criteria of differentially expressed miRNAs, and (5) had reported total number of samples used in study. Thus, due to this inclusion criteria miRNA profiling studies using different cell lines, serum samples of GC patients, plasma samples, or using different miRNA technologies (RNA-seq and qPCR) and review articles were excluded.
Data collection
From each selected paper, following information were retrieved: author, journal and year of publication, location of study, period of sample collection (if available), selec-tion and characteristics of recruited GC patient, total samples used for the study, platform used for miRNA expression profiling, cut-off criteria of statistically differ-entially expressed miRNAs, total number of differdiffer-entially expressed miRNAs, list of upregulated and downregulated miRNAs and their fold change (if available).
Ranking
Each of the published miRNA expression profiling studies that compare miRNA expression between the GC and neighboring noncancerous gastric tissues provided a list of differentially expressed miRNAs. Then for systematic review the vote-counting strategy based method of ranking potential molecular markers as given by Griffith [12] and Chan [13] was used. MiRNAs were ranked on the basis of criteria as: (1) number of the studies that consistently reported the miRNA as differentially expressed, (2) the consistent direction of change of differentially expressed microRNA, and (3) total number of samples for compari-son in agreement.
Identify the experimentally validated microRNA target genes
To explore the target genes of miRNAs related to GC miRTarBase (mirtarbase.mbc.nctu.edu.tw), was used which is the recently established database with largest col-lection of more than 34,000 manually curated miRNA-target interactions, all of which are experimentally validated by luciferase reporter assay, western blot, or microarray experiments with overexpression or knock-down of miRNAs [28].
Enrichment analysis
Enrichment analyses for gene ontology (GO) [29] terms and Kyoto encyclopedia of genes and genomes (KEGG) [30] pathways were carried out with Database for Anno-tation, Visualization, and Integrated Discovery (DAVID) [31]. DAVID is a web-accessible program that integrates functional genomics annotations with intuitive graphical summaries. List of gene or protein identifiers are rapidly annotated and summarized according to shared categori-cal data for Gene Ontology, protein domain, and biochemical pathway membership. For this, we listed all the target genes of downregulated microRNAs. The top 20 GO terms and KEGG pathways showing association with target genes were listed with GO terms, KEGG path-way, number of genes in the term, number of genes in the pathways, andP-value.
Results
Included independent studies
In total, 214 studies were recorded using PubMed. According to the inclusion and exclusion criteria, only 14 independent studies were included in our analysis. The system work flow that was used in our analysis was as shown in Figure 1. The information of the selected stud-ies was tabulated in Table 1 in the descending order of their year of publication. These studies show the different platforms as well as various statistical and biocomputa-tional analyses had been used for the microRNA expres-sion profiling. The number of differentially expressed microRNAs ranges from 9 to 326. Two studies Carvalho [14], Oh [18] provided the top five and top 40 differen-tially expressed microRNAs, respectively, one study Tsu-kamoto [21] provided the differentially expressed microRNAs with the raw signal over 500 and one study Yao [23] provided the differentially expressed microRNAs above twofold changes. Nine of the 14 eligible studies provided fold change (FC) information of differentially expressed miRNAs.
Differentially expressed microRNAs
A total of 352 differentially expressed miRNAs were reported from 14 microRNA expression profiling studies that compared GC tissue with corresponding
Table 1. Characteristics of miRNA datasets in human gastric cancer. Dataset Year Region Tumor type No. of
samples (cancer/ normal)
Platform (manufacturer) Total miRNA Differentially expressed miRNAs Criteria Up Down Total Carvalho [14] 2012 Netherlands GCI 47 (37/10) miRNAChip_human_v2 (National DNA-Microarray Facility, University of Aveiro, Portugal) 703 P < 0.05, FDR <+ 0.05 5 1 5 1 70 Kim [15] 2011 Korea GCI, GCD 124 (90/34) LMT miRNA microarray (Agilent technologies) 1667 P < 0.005 62 63 125 Li [16] 2011 China GCI 12 (6/6) miRCURY Array LNA microRNA Chip (v.14.0) (Exiqon) 904 P < 0.01 FC > 2 40 36 76 Li [17] 2011 China NR 20 (10/10) TaqMan Human miRNA Array v1.0 (Applied Biosystems) 365 P < 0.05 16 6 2 2 Oh [18] 2011 Singapore GCI, GCD 80 (40/40) Agilent Human miRNA Microarrays (V2, Agilent) 723 FDR < 0.01 40 1 40 1 146 Tchernitsa [19] 2010 Germany GCI 12 (6/6) NCode TM MultiSpecies miRNA Microarray V1 (Invitrogen) 373 Significance of class comparison = 0.05 20 2 2 2 Ding [20] 2010 China GCI, GCD 12 (6/6) l Paraflo microfluidic chip (LC Sciences) NR P < 0.05 8 7 15 Tsukamoto [21] 2010 Japan GCI, GCD 27 (22/5) G4470A Human MiRNA Microarray (Agilent technologies) 470 P < 0.05 33 2 6 2 102 Ueda [22] 2010 Japan GCI, GCD 353 (184/169) microRNA microarray chip (OSU_CCC version 3.0, ArrayExpress) 326 P < 0.01 22 13 35 Yao [23] 2009 China NR 6 (3/3) miRCURY LNA microarray Array(v.11.0) (Exiqon) 847 FC > 25 9 3 46 3 326 Luo [24] 2009 China NR 27 (24/3) NR 328 P < 0.05 7 1 9 2 6 Liu [25] 2009 China NR 8 (4/4) microRNA Microarray (Packard Biochip Technologies ScanArray Express microarray) 243 P < 0.05 4 5 9 Petrocca [26] 2008 Italy GCI 40 (20/20) second generation miRNA microarray chips (V2)(Amersham BioScience Codelink) 250 Significance analysis of microarray (SAM) 14 5 1 9 Volinia [27] 2006 USA NR 41 (20/21) miRNA microarray (Amersham BioScience Codelink) 190 FDR = 0.06 22 6 2 8 1Only top most miRNAs selected. 2miRNAs above with raw signal above 500 selected [21] . 3miRNAs over twofold change provided (GCI, gastric cancer intestinal; GCD, gastric cancer diffuse, NR, not reported).
ous gastric tissue, with 120 microRNAs reported in at least two studies.
Consistently and inconsistently reported microRNAs
Among 352 microRNAs, 120 miRNAs (34.18%) were reported in at least two studies. Among 120 differen-tially expressed miRNAs, 69 miRNAs (57.5%) were with consistent direction of expression of which 41 were reported to be upregulated (Table 2) and 28 downregu-lated (Table 3). Fifty-one miRNAs (42.5%) were
reported to be with inconsistent direction of expression (Table S1).
In the group of consistently reported upregulated miRNAs, miR-21 was reported in 10 studies followed by three miRNAs, miR-25, miR-92 and miR-223 in eight studies. In the consistently reported downregulated miR-NAs, miR-375 was reported in six studies followed by two miRNAs, miR-148a, and miR-30d in five studies. However, in the group of inconsistently reported miRNAs, miR-107 was reported in nine studies with upregulation in eight studies and downregulation in one study followed by miR-103 in eight studies with seven
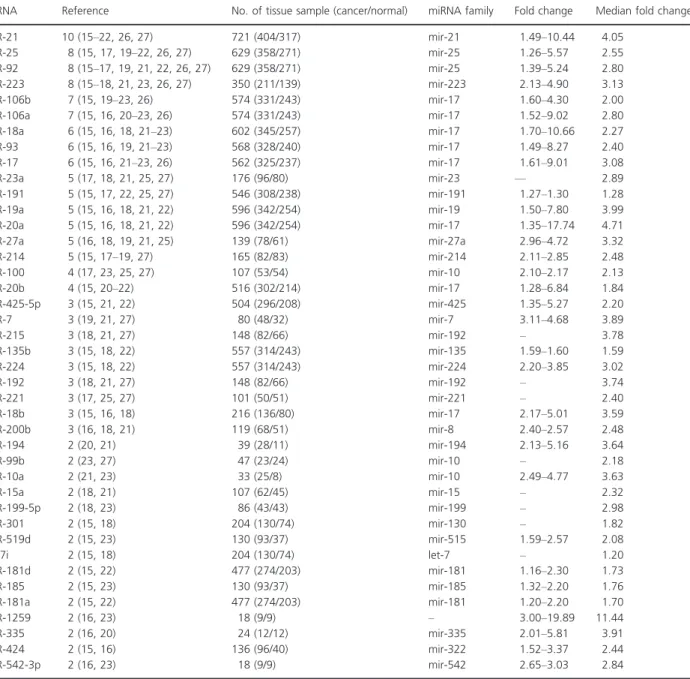
Table 2. Consistently reported upregulated miRNAs in profiling studies (gastric cancer tissue vs. normal).
miRNA Reference No. of tissue sample (cancer/normal) miRNA family Fold change Median fold change
miR-21 10 (15–22, 26, 27) 721 (404/317) mir-21 1.49–10.44 4.05 miR-25 8 (15, 17, 19–22, 26, 27) 629 (358/271) mir-25 1.26–5.57 2.55 miR-92 8 (15–17, 19, 21, 22, 26, 27) 629 (358/271) mir-25 1.39–5.24 2.80 miR-223 8 (15–18, 21, 23, 26, 27) 350 (211/139) mir-223 2.13–4.90 3.13 miR-106b 7 (15, 19–23, 26) 574 (331/243) mir-17 1.60–4.30 2.00 miR-106a 7 (15, 16, 20–23, 26) 574 (331/243) mir-17 1.52–9.02 2.80 miR-18a 6 (15, 16, 18, 21–23) 602 (345/257) mir-17 1.70–10.66 2.27 miR-93 6 (15, 16, 19, 21–23) 568 (328/240) mir-17 1.49–8.27 2.40 miR-17 6 (15, 16, 21–23, 26) 562 (325/237) mir-17 1.61–9.01 3.08 miR-23a 5 (17, 18, 21, 25, 27) 176 (96/80) mir-23 — 2.89 miR-191 5 (15, 17, 22, 25, 27) 546 (308/238) mir-191 1.27–1.30 1.28 miR-19a 5 (15, 16, 18, 21, 22) 596 (342/254) mir-19 1.50–7.80 3.99 miR-20a 5 (15, 16, 18, 21, 22) 596 (342/254) mir-17 1.35–17.74 4.71 miR-27a 5 (16, 18, 19, 21, 25) 139 (78/61) mir-27a 2.96–4.72 3.32 miR-214 5 (15, 17–19, 27) 165 (82/83) mir-214 2.11–2.85 2.48 miR-100 4 (17, 23, 25, 27) 107 (53/54) mir-10 2.10–2.17 2.13 miR-20b 4 (15, 20–22) 516 (302/214) mir-17 1.28–6.84 1.84 miR-425-5p 3 (15, 21, 22) 504 (296/208) mir-425 1.35–5.27 2.20 miR-7 3 (19, 21, 27) 80 (48/32) mir-7 3.11–4.68 3.89 miR-215 3 (18, 21, 27) 148 (82/66) mir-192 – 3.78 miR-135b 3 (15, 18, 22) 557 (314/243) mir-135 1.59–1.60 1.59 miR-224 3 (15, 18, 22) 557 (314/243) mir-224 2.20–3.85 3.02 miR-192 3 (18, 21, 27) 148 (82/66) mir-192 – 3.74 miR-221 3 (17, 25, 27) 101 (50/51) mir-221 – 2.40 miR-18b 3 (15, 16, 18) 216 (136/80) mir-17 2.17–5.01 3.59 miR-200b 3 (16, 18, 21) 119 (68/51) mir-8 2.40–2.57 2.48 miR-194 2 (20, 21) 39 (28/11) mir-194 2.13–5.16 3.64 miR-99b 2 (23, 27) 47 (23/24) mir-10 – 2.18 miR-10a 2 (21, 23) 33 (25/8) mir-10 2.49–4.77 3.63 miR-15a 2 (18, 21) 107 (62/45) mir-15 – 2.32 miR-199-5p 2 (18, 23) 86 (43/43) mir-199 – 2.98 miR-301 2 (15, 18) 204 (130/74) mir-130 – 1.82 miR-519d 2 (15, 23) 130 (93/37) mir-515 1.59–2.57 2.08 let-7i 2 (15, 18) 204 (130/74) let-7 – 1.20 miR-181d 2 (15, 22) 477 (274/203) mir-181 1.16–2.30 1.73 miR-185 2 (15, 23) 130 (93/37) mir-185 1.32–2.20 1.76 miR-181a 2 (15, 22) 477 (274/203) mir-181 1.20–2.20 1.70 miR-1259 2 (16, 23) 18 (9/9) – 3.00–19.89 11.44 miR-335 2 (16, 20) 24 (12/12) mir-335 2.01–5.81 3.91 miR-424 2 (15, 16) 136 (96/40) mir-322 1.52–3.37 2.44 miR-542-3p 2 (16, 23) 18 (9/9) mir-542 2.65–3.03 2.84
and one study showing up- and downregulation, respectively.
Target genes of differentially expressed microRNAs
After we identified the total microRNAs that were down-regulated, updown-regulated, and both down and upregulated in gastric cancer, we used the miRTarBase to search all of the target genes for the set of miRNAs. We identified 362 and 489 nonredundant target genes for the set of miRNAs downregulated and upregulated, respectively, in GC (Tables S2 and S3). MicroRNAs can act as both tumor suppressor and oncogenes. A total of 701 nonredundant target genes were identified for the set of miRNAs that were both upregulated and downregulated in GC (Table S4). Downregulated microRNAs play important role as tumor suppressors, so we focused on the set of downreg-ulated microRNAs target genes.
Enrichment analysis result
We used Database for Annotation, Visualization, and Inte-grated Discovery (DAVID) to find the molecular networks
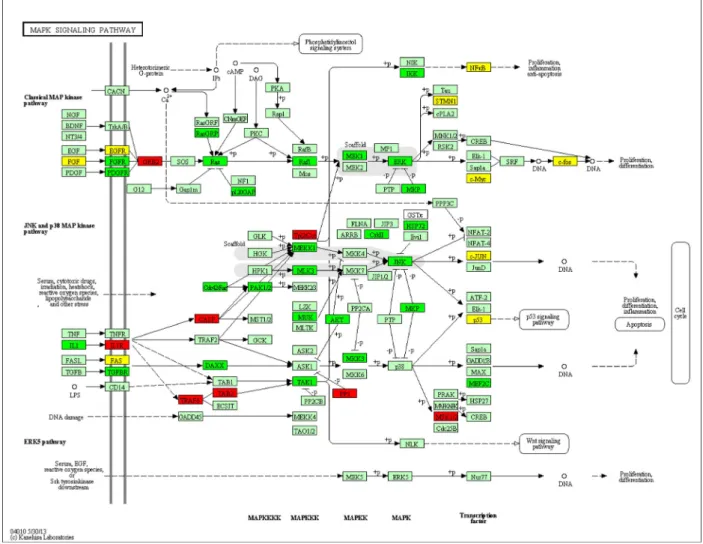
of downregulated microRNAs target genes. The top 20 GO terms and KEGG pathways showing significant association with target genes were listed with GO terms, KEGG path-way, number of genes in the terms, number of genes in the pathways, andP-value (Tables S5 and S6). The significant GO terms were related to regulation of cell proliferation, regulation of apoptosis, regulation of cell cycle (Table S5 and Fig. S1), and KEGG pathways were related to various types of cancer, focal adhesion, adherens junction, signal-ing pathways such as MAPK, toll-like receptor and p53 (Table S6 and Fig. S2). One of the signaling pathways, MAPK signaling pathway, plays important role in gastric cancer. Figure 2 showed that genes like TRAF6, GRB2, and IL1R involved in MAPK signaling pathway were targeted by downregulated miRNAs marked in red boxes. Green boxes represent the genes targeted by upregulated miRNAs and yellow boxes indicate the genes targeted by both up-regulated and downup-regulated miRNAs. We used KEGG API to produce color-coded pathways shown in Figure 2.
Discussion
As different microRNA expression profiling studies use dif-ferent platforms and difdif-ferent processing methods, there is
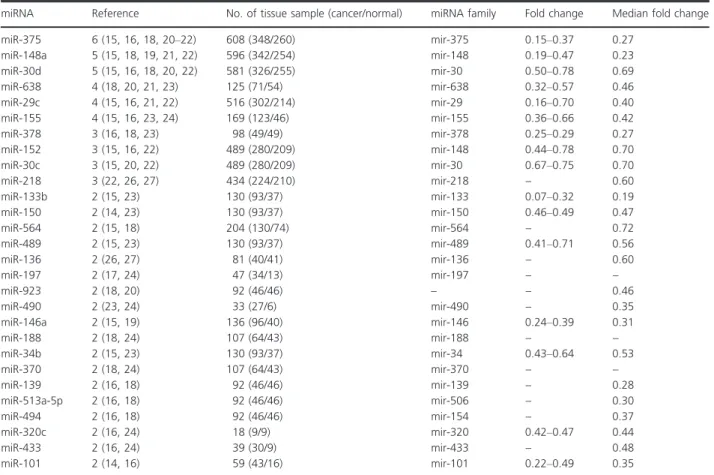
Table 3. Consistently reported downregulated miRNAs in profiling studies (gastric cancer tissue vs. normal).
miRNA Reference No. of tissue sample (cancer/normal) miRNA family Fold change Median fold change
miR-375 6 (15, 16, 18, 20–22) 608 (348/260) mir-375 0.15–0.37 0.27 miR-148a 5 (15, 18, 19, 21, 22) 596 (342/254) mir-148 0.19–0.47 0.23 miR-30d 5 (15, 16, 18, 20, 22) 581 (326/255) mir-30 0.50–0.78 0.69 miR-638 4 (18, 20, 21, 23) 125 (71/54) mir-638 0.32–0.57 0.46 miR-29c 4 (15, 16, 21, 22) 516 (302/214) mir-29 0.16–0.70 0.40 miR-155 4 (15, 16, 23, 24) 169 (123/46) mir-155 0.36–0.66 0.42 miR-378 3 (16, 18, 23) 98 (49/49) mir-378 0.25–0.29 0.27 miR-152 3 (15, 16, 22) 489 (280/209) mir-148 0.44–0.78 0.70 miR-30c 3 (15, 20, 22) 489 (280/209) mir-30 0.67–0.75 0.70 miR-218 3 (22, 26, 27) 434 (224/210) mir-218 – 0.60 miR-133b 2 (15, 23) 130 (93/37) mir-133 0.07–0.32 0.19 miR-150 2 (14, 23) 130 (93/37) mir-150 0.46–0.49 0.47 miR-564 2 (15, 18) 204 (130/74) mir-564 – 0.72 miR-489 2 (15, 23) 130 (93/37) mir-489 0.41–0.71 0.56 miR-136 2 (26, 27) 81 (40/41) mir-136 – 0.60 miR-197 2 (17, 24) 47 (34/13) mir-197 – – miR-923 2 (18, 20) 92 (46/46) – – 0.46 miR-490 2 (23, 24) 33 (27/6) mir-490 – 0.35 miR-146a 2 (15, 19) 136 (96/40) mir-146 0.24–0.39 0.31 miR-188 2 (18, 24) 107 (64/43) mir-188 – – miR-34b 2 (15, 23) 130 (93/37) mir-34 0.43–0.64 0.53 miR-370 2 (18, 24) 107 (64/43) mir-370 – – miR-139 2 (16, 18) 92 (46/46) mir-139 – 0.28 miR-513a-5p 2 (16, 18) 92 (46/46) mir-506 – 0.30 miR-494 2 (16, 18) 92 (46/46) mir-154 – 0.37 miR-320c 2 (16, 24) 18 (9/9) mir-320 0.42–0.47 0.44 miR-433 2 (16, 24) 39 (30/9) mir-433 – 0.48 miR-101 2 (14, 16) 59 (43/16) mir-101 0.22–0.49 0.35
the lack of agreement among these studies. Vote counting is the most intuitive and simplest method for combining results from different studies to discover the patterns among those results. Although vote-counting method is statistically inefficient, but it is suitable for dealing with the issue that only the list of differentially expressed genes is available. Here, a possible solution for this problem is to determine the overlap among many studies using different platforms and observe which miRNAs are consistently reported as differentially expressed ones [12, 13].
This type of systematic review had been done previ-ously to determine differentially expressed genes in thy-roid [12] and colorectal cancer [13] at gene level. Ma [32] carried out the systematic review in exploring candi-date miRNA biomarkers in human colorectal cancer. Guan [33] carried out the meta-analysis of human lung cancer microRNA expression profiling studies comparing cancer tissues with normal tissues. The meta-analysis approach was suitable and effective solution for
identifica-tion of statistically significant miRNA meta-signature by combining several miRNA expression studies [34]. How-ever, to our knowledge, no such systematic review of miRNA profiling studies has been performed in gastric cancer.
The present systematic review identified one most con-sistently reported upregulated miRNA, miRNA-21, which was consistently reported to be differentially expressed in 10 studies followed by miR-25, miR-92, and miR-223 in eight studies. MiR-375 was reported to be consistently downregulated in six studies followed by miR-148a and miR-30d in five studies.
The most consistently reported differentially expressed miRNA of upregulation in our systematic review was miR-21 which had an oncogenic activity. The upregulation of miR-21 significantly promoted cell proliferation and revealed a higher proportion of cells at S phase and knock-down of miR-21 expression resulted in cell-cycle arrest at G2/M phase and inhibited cell proliferation [35]. Lee [36]
showed higher expression of miR-21 in breast cancer tis-sues than in normal tistis-sues and the high miR-21 expression was associated with mastectomy, larger tumor size, higher stage, higher grade, estrogen receptor (ER) negative, human epidermal growth factor receptor 2 (HER2) positive. From our study after miR-21, the most consistently reported upregulated microRNAs in eight studies were miR-25, miR-92, and miR-223. The up-regulation of miR-25 was significantly correlated with the status of lymph node metastasis and TNM (Tumor, Node, and Metastasis) stage and overexpression markedly promoted migration and invasion of esophageal squamous cell carcinoma [37]. Previous miRNA expression analysis had shown high expression of miR-25 in colorectal carcinoma [38]. miR-92a was implicated in human hepatocellular carci-noma development and miR-92a in human blood had the potential to be a noninvasive molecular marker for the diag-nosis of human hepatocellular carcinoma [39]. Expression profiling studies had shown that miR-92 is overexpressed in oral carcinoma [40]. The miR-223 expression was signifi-cantly higher in cancerous tissues than in the corresponding normal tissues in esophageal squamous cell carcinoma and high miR-223 expression demonstrated a significantly poorer prognosis than those with low expression [41].
The most consistently reported differentially expressed microRNA of downregulation in our systematic review is miR-375 which has an antioncogenic activity. The overex-pressed miR-375 supoverex-pressed cell proliferation, blocked G1-to S cell-cycle transition, and inhibited cell migration and invasion in human cervical SiHa and CaSki cells, sug-gesting that miR-375 is a candidate tumor suppressor [42]. In this systematic review study, we found miR-148a and miR-30d were consistently reported downregulated in five studies. Liffers [43] found that miR-148a exhibited a significant fourfold downregulation in pancreatic ductal adenocarcinoma as opposed to normal pancreatic ductal cells and observed that stable lentiviral mediated overex-pression of miR-148a in pancreatic cancer cell line, inhib-ited tumor cell growth and colony formation. Zhao [44] observed a decrease in miR-30d expression in the islets of diabetic db/db mice, in which MAP4K4 expression level was elevated and overexpression of miR-30d protected b-cells against TNF-a suppression on both insulin tran-scription and insulin secretion.
The MAPK pathway is an important pathway in caus-ing the gastric cancer. From our study, we found one important gene, TNF receptor-associated factor 6 (TRAF6), involved in MAPK signaling pathway was tar-geted by downregulated microRNA. Comprehensive analysis using cDNA microarray showed that RUNX3 upregulated 17 apoptosis-related genes including TRAF6 in MKN-1 cells [45]. A positive correlation between TRAF6 and ubiquitin expression was found suggesting
TRAF6 may upregulate ubiquitin activity in cancer cachexia [46]. Inhibition of TRAF6 in human lung can-cer cells suppressed NF-jB activation, anchorage-inde-pendent growth and tumor formation [47]. The genes such as Ras, Raf, MEK, MAPK, ERK that were targeted by upregulated microRNAs in our analysis had signifi-cant role in the pathway. MAPK pathway had been often found to be deregulated in cancers and consists of sev-eral kinases (Ras, Raf, MEK) that were activated by phosphorylation upon ligand binding to a membrane receptor, ending up in the activation of several proteins involved in processes of cell invasion, apoptosis, tran-scription, survival, and drug resistance [48]. Ras/MAPK activation was found to be associated with cell prolifera-tion and gastric carcinomas [49]. ERK1/2 the final effec-tors of this pathway, were also found to be activated in gastric cancers [50].
In our study, 51 microRNAs among 352 microRNAs were reported to be inconsistent expressed in at least two independent studies. An elegant way to identify inconsis-tent studies is to correlate all studies with each other (study–study inconsistent miRNA matrix). Table S7 shows that study 15, 23, 18, and 16 have the higher inconsistency with other studies; especially reference 15 reported 13 microRNAs inconsistent with reference 18. The study number is the same as the reference number shown in the main manuscript. Previous studies have shown that some differentially expressed miRNAs are reported in inconsistent direction among various studies when using microRNA arrays for a particular disease such as a cancer [34, 51]. The other three possible reasons for these observed inconsistencies are: the lack of sufficient relevant clinical data, the heterogeneous tissue samples, and the poor study design.
In conclusion, from our systematic review study we identified that miR-21 is the most consistently reported upregulated microRNA and miR-375 is the most consis-tently reported downregulated microRNA in gastric can-cer. The findings of our present study may have important clinical significance. The results of this system-atic review of GC miRNA expression profiling studies provide some clues of the potential biomarkers in gastric cancer. The most consistently reported differentially expressed microRNAs may be used as efficient biomarkers and therapeutic targets. Our study found several promis-ing miRNAs that had been consistently reported. How-ever, still more investigations are needed for the clinical studies focusing on these miRNAs in order to understand the potential roles of these microRNAs in gastric cancer.
Conflict of Interest
References
1. Ferlay, J., H. R. Shin, F. Bray, D. Forman, C. Mathers, and D. M. Parkin. 2010. Estimates of worldwide burden of cancer in 2008: GLOBOCAN 2008. Int. J. Cancer 127:2893–2917. 2. Coburn, N. G. 2009. Lymph nodes and gastric cancer.
J. Surg. Oncol. 99:199–206.
3. Konishi, H., D. Ichikawa, S. Komatsu, A. Shiozaki, M. Tsujiura, H. Takeshita, et al. 2012. Detection of gastric cancer-associated microRNAs on microRNA microarray comparing pre- and post-operative plasma. Br. J. Cancer 106:740–747.
4. Li, B. S., Y. L. Zhao, G. Guo, W. Li, E. D. Zhu, X. Luo, et al. 2012. Plasma microRNAs, miR-223, miR-21 and miR-218, as novel potential biomarkers for gastric cancer detection. PLoS One 7:e41629.
5. Jiang, Z., J. Guo, B. Xiao, Y. Miao, R. Huang, D. Li, et al. 2010. Increased expression of miR-421 in human gastric carcinoma and its clinical association. J. Gastroenterol. 45:17–23.
6. Bartel, D. P. 2004. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell 116:281–297.
7. Esquela-Kerscher, A., and F. J. Slack. 2006. Oncomirs— microRNAs with a role in cancer. Nat. Rev. Cancer 6: 259–269.
8. Alvarez-Garcia, I., and E. A. Miska. 2005. MicroRNA functions in animal development and human disease. Development 132:4653–4662.
9. Zhu, W., D. Zhu, S. Lu, T. Wang, J. Wang, B. Jiang, et al. 2012. miR-497 modulates multidrug resistance of human cancer cell lines by targeting BCL2. Med. Oncol. 29:384–391.
10. Guo, J., Y. Miao, B. Xiao, R. Huan, Z. Jiang, D. Meng, et al. 2009. Differential expression of microRNA species in human gastric cancer versus non-tumorous tissues. J. Gastroenterol. Hepatol. 24:652–657.
11. Song, M. Y., K. F. Pan, H. J. Su, L. Zhang, J. L. Ma, J. Y. Li, et al. 2012. Identification of serum microRNAs as novel non-invasive biomarkers for early detection of gastric cancer. PLoS One 7:e33608.
12. Griffith, O. L., A. Melck, S. J. Jones, and S. M. Wiseman. 2006. Meta-analysis and meta-review of thyroid cancer gene expression profiling studies identifies important diagnostic biomarkers. J. Clin. Oncol. 24:5043–5051. 13. Chan, S. K., O. L. Griffith, I. T. Tai, and S. J. Jones. 2008.
Meta-analysis of colorectal cancer gene expression profiling studies identifies consistently reported candidate
biomarkers. Cancer Epidemiol. Biomarkers Prev. 17: 543–552.
14. Carvalho, J., N. C. van Grieken, P. M. Pereira, S. Sousa, M. Tijssen, T. E. Buffart, et al. 2012. Lack of
microRNA-101 causes E-cadherin functional deregulation through EZH2 up-regulation in intestinal gastric cancer. J. Pathol. 228:31–44.
15. Kim, C. H., H. K. Kim, R. L. Rettig, J. Kim, E. T. Lee, O. Aprelikova, et al. 2011. miRNA signature associated with outcome of gastric cancer patients following
chemotherapy. BMC Med. Genomics 4:79.
16. Li, X., F. Luo, Q. Li, M. Xu, D. Feng, G. Zhang, et al. 2011. Identification of new aberrantly expressed miRNAs in intestinal-type gastric cancer and its clinical significance. Oncol. Rep. 26:1431–1439.
17. Li, X., Y. Zhang, H. Zhang, X. Liu, T. Gong, M. Li, et al. 2011. miRNA-223 promotes gastric cancer invasion and metastasis by targeting tumor suppressor EPB41L3. Mol. Cancer Res. 9:824–833.
18. Oh, H. K., A. L. Tan, K. Das, C. H. Ooi, N. T. Deng, I. B. Tan, et al. 2011. Genomic loss of miR-486 regulates tumor progression and the OLFM4 antiapoptotic factor in gastric cancer. Clin. Cancer Res. 17:2657–2667.
19. Tchernitsa, O., A. Kasajima, R. Schafer, R. J. Kuban, U. Ungethum, B. Gyorffy, et al. 2010. Systematic evaluation of the miRNA-ome and its downstream effects on mRNA expression identifies gastric cancer progression. J. Pathol. 222:310–319.
20. Ding, L., Y. Xu, W. Zhang, Y. Deng, M. Si, Y. Du, et al. 2010. MiR-375 frequently downregulated in gastric cancer inhibits cell proliferation by targeting JAK2. Cell Res. 20:784–793.
21. Tsukamoto, Y., C. Nakada, T. Noguchi, M. Tanigawa, L. T. Nguyen, T. Uchida, et al. 2010. MicroRNA-375 is downregulated in gastric carcinomas and regulates cell survival by targeting PDK1 and 14-3-3zeta. Cancer Res. 70:2339–2349.
22. Ueda, T., S. Volinia, H. Okumura, M. Shimizu, C. Taccioli, S. Rossi, et al. 2010. Relation between microRNA expression and progression and prognosis of gastric cancer: a microRNA expression analysis. Lancet Oncol. 11:136–146.
23. Yao, Y., A. L. Suo, Z. F. Li, L. Y. Liu, T. Tian, L. Ni, et al. 2009. MicroRNA profiling of human gastric cancer. Mol. Med. Report 2:963–970.
24. Luo, H., H. Zhang, Z. Zhang, X. Zhang, B. Ning, J. Guo, et al. 2009. Down-regulated miR-9 and miR-433 in human gastric carcinoma. J. Exp. Clin. Cancer Res. 28:82. 25. Liu, T., H. Tang, Y. Lang, M. Liu, and X. Li. 2009.
MicroRNA-27a functions as an oncogene in gastric adenocarcinoma by targeting prohibitin. Cancer Lett. 273:233–242.
26. Petrocca, F., R. Visone, M. R. Onelli, M. H. Shah, M. S. Nicoloso, I. de Martino, et al. 2008. E2F1-regulated microRNAs impair TGFbeta-dependent cell-cycle arrest and apoptosis in gastric cancer. Cancer Cell 13: 272–286.
27. Volinia, S., G. A. Calin, C. G. Liu, S. Ambs, A. Cimmino, F. Petrocca, et al. 2006. A microRNA expression signature of human solid tumors defines cancer gene targets. Proc. Natl Acad. Sci. USA 103:2257–2261.
28. Hsu, S. D., F. M. Lin, W. Y. Wu, C. Liang, W. C. Huang, W. L. Chan, et al. 2011. miRTarBase: a database curates experimentally validated microRNA-target interactions. Nucleic Acids Res. 39:D163–D169.
29. Harris, M. A., J. Clark, A. Ireland, J. Lomax, M. Ashburner, R. Foulger, et al. 2004. The gene ontology (GO) database and informatics resource. Nucleic Acids Res. 32:D258–D261.
30. Kanehisa, M., S. Goto, S. Kawashima, Y. Okuno, and M. Hattori. 2004. The KEGG resource for deciphering the genome. Nucleic Acids Res. 32:D277–D280.
31. Dennis, G. Jr., B. T. Sherman, D. A. Hosack, J. Yang, W. Gao, H. C. Lane, et al. 2003. DAVID: Database for Annotation, Visualization, and Integrated Discovery. Genome Biol. 4:P3.
32. Ma, Y., P. Zhang, J. Yang, Z. Liu, Z. Yang, and H. Qin. 2012. Candidate microRNA biomarkers in human colorectal cancer: systematic review profiling studies and experimental validation. Int. J. Cancer 130:2077–2087. 33. Guan, P., Z. Yin, X. Li, W. Wu, and B. Zhou. 2012.
Meta-analysis of human lung cancer microRNA expression profiling studies comparing cancer tissues with normal tissues. J. Exp. Clin. Cancer Res. 31:54.
34. Vosa, U., T. Vooder, R. Kolde, J. Vilo, A. Metspalu, and T. Annilo. 2013. Meta-analysis of microRNA expression in lung cancer. Int. J. Cancer 132:2884–2893.
35. Zhong, Z., Z. Dong, L. Yang, and Z. Gong. 2012. miR-21 induces cell cycle at S phase and modulates cell
proliferation by down-regulating hMSH2 in lung cancer. J. Cancer Res. Clin. Oncol. 138:1781–1788.
36. Lee, J. A., H. Y. Lee, E. S. Lee, I. Kim, and J. W. Bae. 2011. Prognostic implications of microRNA-21
overexpression in invasive ductal carcinomas of the breast. J. Breast Cancer 14:269–275.
37. Xu, X., Z. Chen, X. Zhao, J. Wang, D. Ding, Z. Wang, et al. 2012. MicroRNA-25 promotes cell migration and invasion in esophageal squamous cell carcinoma. Biochem. Biophys. Res. Commun. 421:640–645.
38. Nishida, N., M. Nagahara, T. Sato, K. Mimori, T. Sudo, F. Tanaka, et al. 2012. Microarray analysis of colorectal cancer stromal tissue reveals upregulation of two oncogenic miRNA clusters. Clin. Cancer Res. 18: 3054–3070.
39. Shigoka, M., A. Tsuchida, T. Matsudo, Y. Nagakawa, H. Saito, Y. Suzuki, et al. 2010. Deregulation of miR-92a expression is implicated in hepatocellular carcinoma development. Pathol. Int. 60:351–357.
40. Scapoli, L., A. Palmieri, L. Lo Muzio L, F. Pezzetti, C. Rubini, A. Girardi, et al. 2010. MicroRNA expression profiling of oral carcinoma identifies new markers of tumor progression. Int. J. Immunopathol. Pharmacol. 23: 1229–1234.
41. Kurashige, J., M. Watanabe, M. Iwatsuki, K. Kinoshita, S. Saito, Y. Hiyoshi, et al. 2012. Overexpression of
microRNA-223 regulates the ubiquitin ligase FBXW7 in oesophageal squamous cell carcinoma. Br. J. Cancer 106:182–188.
42. Wang, F., Y. Li, J. Zhou, J. Xu, C. Peng, F. Ye, et al. 2011. miR-375 is down-regulated in squamous cervical cancer and inhibits cell migration and invasion via targeting transcription factor SP1. Am. J. Pathol. 179:2580–2588. 43. Liffers, S. T., J. B. Munding, M. Vogt, J. D. Kuhlmann, B.
Verdoodt, S. Nambiar, et al. 2011. MicroRNA-148a is down-regulated in human pancreatic ductal
adenocarcinomas and regulates cell survival by targeting CDC25B. Lab. Invest. 91:1472–1479.
44. Zhao, X., R. Mohan, S. Ozcan, and X. Tang. 2012. MicroRNA-30d induces insulin transcription factor MafA and insulin production by targeting mitogen-activated protein 4 kinase 4 (MAP4K4) in pancreatic beta-cells. J. Biol. Chem. 287:31155–31164.
45. Nagahama, Y., M. Ishimaru, M. Osaki, T. Inoue, A. Maeda, C. Nakada, et al. 2008. Apoptotic pathway induced by transduction of RUNX3 in the human gastric
carcinoma cell line MKN-1. Cancer Sci. 99:23–30. 46. Sun, Y. S., Z. Y. Ye, Z. Y. Qian, X. D. Xu, and J. F. Hu.
2012. Expression of TRAF6 and ubiquitin mRNA in skeletal muscle of gastric cancer patients. J. Exp. Clin. Cancer Res. 31:81.
47. Starczynowski, D. T., W. W. Lockwood, S. Delehouzee, R. Chari, J. Wegrzyn, M. Fuller, et al. 2011. TRAF6 is an amplified oncogene bridging the RAS and NF-kappaB pathways in human lung cancer. J. Clin. Invest. 121:4095–4105.
48. Kim, E. K., and E. J. Choi. 2010. Pathological roles of MAPK signaling pathways in human diseases. Biochim. Biophys. Acta 1802:396–405.
49. Regalo, G., C. Resende, X. Wen, B. Gomes, C. Duraes, R. Seruca, et al. 2010. C/EBP alpha expression is associated with homeostasis of the gastric epithelium and with gastric carcinogenesis. Lab. Invest. 90:1132–1139.
50. Liang, B., S. Wang, X. G. Zhu, Y. X. Yu, Z. R. Cui, and Y. Z. Yu. 2005. Increased expression of mitogen-activated protein kinase and its upstream regulating signal in human gastric cancer. World J. Gastroenterol. 11:623–628. 51. Gong, X., R. Wu, H. Wang, X. Guo, D. Wang, Y. Gu,
et al. 2011. Evaluating the consistency of differential expression of microRNA detected in human cancers. Mol. Cancer Ther. 10:752–760.
Supporting Information
Additional Supporting Information may be found in the online version of this article:
Figure S1. The top 20 significant GO terms for downreg-ulated miRNAs target genes.
downregulated miRNAs target genes.
Table S1. Inconsistently reported miRNAs in profiling studies (gastric cancer tissue vs. normal).
Table S2. Experimentally validated target genes for mi-croRNAs downregulated in gastric cancer.
Table S3. Experimentally validated target genes for mi-croRNAs upregulated in gastric cancer.
Table S4. Experimentally validated target genes for
mi-croRNAs up- and downregulated in gastric cancer. Table S5. Gene ontology terms of target genes for down-regulated miRNAs in gastric cancer.
Table S6. KEGG pathways of target genes for downregu-lated miRNAs in gastric cancer.
Table S7. The number of microRNAs mentioned in two references with inconsistent direction (study–study incon-sistent miRNA matrix).