中國醫藥大學機構典藏 China Medical University Repository, Taiwan:Item 310903500/41373
49
0
0
全文
(2) 致謝 在碩士的這兩年,從剛進入蘇振良老師實驗室到現在可以在這裡寫下致謝, 其中的心理感受與一切的酸甜苦辣皆盡在不言中,從生疏到熟悉、從嗷嗷待哺到 振翅飛翔,一步一步的成長,皆伴隨著無數的血汗與喜怒哀樂,如今可以寫出這 篇論文,要感謝的人實在太多,要讓我從一無所知到寫出論文,過程想必十分辛 苦也必定麻煩到許許多多的人。一路的跌跌撞撞,可以說是從一堆不符合預期或 甚至整個過程就失敗的實驗數據中堆砌而成的論文,自己心中有股感動與珍惜。 那這兩年的過程中,謝謝蘇振良老師不厭其煩得對我的英文或是實驗不斷的 教導,也提供了很多豐富的資源讓我可以學習到五花八門的實驗,讓自己的眼界 寬擴了許多。那雖然因為打籃球輸掉了我的半年,但在這裡很感謝老師還是讓我 偷偷的落跑,不過我也學習到了十賭九窮阿!!再一次的感謝老師的教導與照顧, 也十分感謝蕭宏昇老師、夏興國老師、黃志興老師、黃志揚老師與陳健尉老師在 我的碩士生涯中對我的教導。癌生所的謝嘉玲老師、宋賢穎老師、李龍緣老師、 黃偉謙老師與余永倫老師的指導。謝謝陳雅惠學姊當初把我從 cloning 的地獄深 淵拉出來。謝謝實驗室各位的陪伴。謝謝邱勤學姐幫我解惑或是常在重要時刻提 出很受用的意見。謝謝薏雯學姐在實驗上細心的教導、討論甚至是幫助,以及製 造實驗室歡笑氣氛。謝謝筱晴幫忙處理很多平常的瑣事,真是辛苦你了。最後也 感謝尚志、志駿、竺英、百昇、志成、瓜瓜陪我走過碩士生涯,有了你們,讓我 在實驗室一點也不孤單。謝謝我的同學們,怡蕙、筠琦、大翔、嘉紘,我們都擁 有共同走過這段路的記憶,耐人尋味。謝謝所辦的秘書小姐,霆琍學姊真的幫我 們處理很多大大小小的事情,真的很感謝妳!最後,感謝思穎的陪伴,不管悲傷 或是歡笑,都不厭其煩的讓我分享這些心情。謝謝我的爸爸和媽媽讓我可以無後 顧之憂的走過碩士生涯,你們的栽培與奉獻讓我不知如何表達我的感謝。走道這 裡,真的由衷的感謝幫助過我的每一位,我的成長有著你們,我不再徬徨無助, 最後我再說一次,謝謝你們!!!. I.
(3) 目錄 致謝 -------------------------------------------------------------------------------- I 目錄 -------------------------------------------------------------------------------- II 中文摘要 -------------------------------------------------------------------------- IV 英文摘要 -------------------------------------------------------------------------- V. 1.. 2.. 前言 (Introduction) ------------------------------------------------------ (1) 1.1. Breast cancer ------------------------------------------------------- (1). 1.2. E1A Gene Therapy ----------------------------------------------- (1). 1.3. Glucose-regulated protein 78 ---------------------------------- (2). 1.4. Ubiquitin-proteasome system (UPS) ------------------------ (3). 1.5. Hypothesis ---------------------------------------------------------- (4). 研究方法 (Materials and Methods)---------------------------------- (5) 2.1. Cell line -------------------------------------------------------------- (5). 2.2. RNA Isolation and RT-PCR ----------------------------------- (5). 2.3. Transwell migration and invasion assays ------------------(6). 2.4. MTT assay ---------------------------------------------------------- (7). 2.5. Cell tracing assay ------------------------------------------------- (7). 2.6. Western blotting -------------------------------------------------- (7). 2.7. Immunoprecipitation -------------------------------------------- (8). 2.8. Statistical analysis ------------------------------------------------ (8). II.
(4) 3.. 研究結果 (Results) ------------------------------------------------------- (10) 3.1. GRP78 Protein Is Critical for E1A–mediated inhibition of cell mobility -------------------------------------------------------- (10). 3.2. E1A Enhances Ub-dependent Proteolysis of GRP78 -- (11). 3.3. Gp78 Is Required for E1A-mediated Degradation of GRP78 and Suppression Cell Mobility -------------------- (13). 4.. 討論 (Discussion) --------------------------------------------------------- (15). 5.. 參考文獻 (References) -------------------------------------------------- (18). 6.. 圖片 (Figure) ---------------------------------------------------------------(28). III.
(5) 中文摘要 腺病毒蛋白 type 5 E1A (E1A)在抗腫瘤的基因治療上具有相當的潛 力,並已在進行乳癌、頭頸癌和卵巢癌的臨床試驗測試。之前的研究指出 E1A 基因治療確實可以顯著降低腫瘤的發生,並且促使癌細胞進行細胞凋 亡和抑制癌細胞的移動性。然而,E1A 抗腫瘤方面的分子機制尚未全然了 解。我們研究發現 E1A 的表現會顯著得抑制葡萄糖調節蛋白 78 (GRP78), 且此蛋白是一個致癌基因,已廣泛被報導在腫瘤的生長過程中扮演一個重 要的角色。我們觀察到當葡萄糖調節蛋白 78 過度表現時,可以抵消 E1A 所產生的抑制細胞遷移和侵襲能力。另外,為了了解 E1A 如何調控葡萄糖 調節蛋白 78,我們發現到 E1A 會影響到葡萄糖調節蛋白 78 的蛋白層面, 但不會影響核醣核酸的層面。所以我們在 E1A 表現的細胞處理蛋白酶體抑 制劑 MG132,發現可以恢復 E1A 所抑制的葡萄糖調節蛋白 78 的表現量,這 表示了了泛素─蛋白酶體系統可能參予在 E1A 導致葡萄糖調節蛋白 78 降 解的機轉中。因此,這些結果顯示了 E1A 可以促進葡萄糖調節蛋白 78 的 泛素化導致此蛋白降解,進而抑制癌細胞的遷移和侵襲能力。. IV.
(6) Abstract The adenovirus type 5 E1A (E1A) plays a potential role in anti-cancer gene therapy and has been tested in clinical trails. Previous reports suggest that expression of E1A significantly reduces tumorigenesis, promotes cell death, and inhibits cancer cell mobility. However, the molecular mechanisms involved in E1A-mediated anti-cancer activity are not yet completely clear. Here, we show that expression of E1A significantly suppresses the glucose-regulated protein 78 (GRP78), an oncoprotein which has been widely reported to play critical roles in tumor progression. We found that E1A-mediated suppression of cell mobility and invasion ability was overturned by ectopic expression of GRP78. To verify how E1A regulates GRP78, we observed that E1A affects GRP78 protein level but not mRNA level. Treatment with proteasome inhibitor, MG132, recovered GRP78 expression, suggesting that ubiquitin-proteasome system may involve in the E1A-mediated degradation of GRP78. Taken together, these results indicated that E1A facilitates the ubiquitination and degradation of GRP78 and subsequent suppression of cell mobility.. V.
(7) 1.. 前言 (Introduction). 1.1 Breast cancer The three most common cancers are cancers of the lung and bronchus, breast, and colon and rectum, accounting for 50% of estimated new cancer cases in women in 2010. Breast cancer is the most diagnosed cancer and remains the second leading cause of cancer deaths in women in 2010 (1). Metastasis at distant sites is the main cause of cancer death including the cause of 90% death from solid tumors (2). Metastases can remain latent for many years following primary tumor removal. Present therapy focuses on varying combinations of surgery, chemotherapy, and radiation treatment (3). Despite healthcare improvements, metastatic disease remains poorly responsive to conventional therapy and a new modality of treatment is needed.. 1.2 E1A Gene Therapy The adenovirus type 5 E1A (E1A) is the first viral gene expressed after viral infection of host cells and is a well-known transcription factor and is later classified as an immortalization oncogene (4-5). The immortalization of E1A may help ras or E1B oncogene to transform primary embryo cells, but E1A can not exhibit transforming phenotypes alone (6-9). However, many studies show that E1A associates with multiple anti-cancer activities. E1A gene was first reported to reduce the metastatic potential of ras-transformed rat embryo fibroblast cells in 1988 (10). It has been reported that expression of E1A in cancer cells reduce tumorigenic potential, increase contact inhibition, and promote apoptosis in vivo (11-13). Previous studies show that E1A does not bind to the DNA directly, but interacts with transcription factors, including the. 1.
(8) TATA-binding protein, p300 (14-15), several TATA-binding protein-associated factors. (16-17),and. others transcription. factor. such as c-Jun. (18).. E1A-mediated tumor suppression may results from the inhibition of HER-2/neu protein expression through transcriptional repression at the HER-2/neu promoter (11). In addition to E1A-mediated tumor suppressive activity in HER-2/neu-overexpression tumors, other studies indicate that E1A associated with tumor-suppression activities independent of HER-2/neu. A series of studies have shown that E1A inhibits oncogenic signaling pathways including downregulation of HER-2, Akt, IKK and NF-B (19-21) as well as activates tumor suppressing pathways such as enhanced expression of p53, p21, PP2A phosphatase and E-cadherin (22-24). In addition, the E1A has been shown to repress expression of matrix metalloprotease and inhibit metastasis (25-26). Furthermore, E1A has been tested in multiple clinical trails including breast, ovarian and head and neck cancers (27-30). Thus, different from ovarian or head and neck cancer in which regional treatment may be sufficient to improve therapeutic efficacy, breast cancer requires systemic gene delivery system to be effective for therapeutic efficacy. The development of gene therapy to treat breast cancer has been hindered by need for systemic delivery.. 1.3 Glucose-regulated protein 78 The glucose-regulated proteins (GRPs) were first described as protein synthesis induction when cells were deprived of glucose (31). GRP78, also referred to HSPA5, is a major endoplasmic reticulum (ER) chaperone, belonging to member of the heat shock protein 70 (HSP70) family (32). GRP78 is involved in many cellular processes that promote proper protein folding and prevent aggregation of newly synthesized proteins. GRP78 would maintain ER 2.
(9) calcium homeostasis and serve as a critical role under ER stress and is widely used as a maker for ER stress since its discovery in the 1980s (33-34). GRP78 is highly induced through intrinsic factors such as altered glucose metabolism of cancer cells or extrinsic factors such as glucose deprivation, hypoxia, and acidosis in the microenvironment of solid tumor (35). Overexpression of GRP78 was widely reported in many tumors, including lung (36), breast (37-38), stomach (39), prostate (40), colon (41), and liver (42) cancer cells. In the previous studies, GRP78 has been shown to promote cancer metastasis through activation of FAK in hepatocellular carcinoma (43). In addition, knockdown of GRP78 also has been reported to inhibit the activation of Akt in prostate cancer cells (44). GRP78 shows oncogenic activity through promoting tumor proliferation, survival, metastasis, and drug resistance (45-47).. 1.4 Ubiquitin-proteasome system (UPS) To maintain the cellular functionality and viability, the UPS virtually involve in any cellular process that depends on the degradation, regulation or life-span determination of damaged, modified, misfolded proteins or proteins that have become ‘unnecessary’. Most of the 26S proteasome substrates have to be polyubiquitinated protein resulting degradation natively folded intact protein in an ATP-dependent fashion (48). The UPS contains a very complex system, involving in four different kinds of enzymes (E1-E4). First, the step of substrate poly-ubiquitination is the ATP dependent activation of ubiquitin (Ub) by E1 enzyme. The activated ubiquitin is transferred to a lysine-residue of E2 ubiquitin conjugating enzyme, and both E2 and doomed substrate are bound by an E3 enzyme resulting in catalyzing the ubiquitin transfer from the E2 to substrate. The E3 enzymes specifically binds to substrate, containing thousands 3.
(10) of Ub-ligases, each one only specific for a limited number of substrate proteins (49). Multiple cycles of these reactions and the E4 which is ubiquitin-chain elongation factor leads to poly-ubiquitination of the substrate which is recognized and degraded by the 26S proteasome (50). Previous studies show the important biological role of UPS can be involved in pathophysiological processes causing the development of various diseases including cancers (51-53).. 1.5 Hypothesis GRP78 involved in tumorigenesis and cancer progression and provided a therapeutic target and mediator of cancer specific therapy. (54-57). We identified the molecular mechanisms of E1A-mediated inhibition of cancer metastasis. GRP78 was shown to be downregulated by E1A and required for E1A-mediated tumor suppression activities. In current study showed that GRP78 promotes cell mobility and E1A-mediated ubiquitination of GRP78 may contribute to E1A mediated invasion suppression activity.. 4.
(11) 2. Materials and Methods 2.1 Cell line MDA-MB-231, MDA-MB-468, HS578T and HBL100 cells were obtained from the American Type Culture Collection (ATCC) and grown in Dulbecco’s modified Eagle’s medium (DMEM) / F12 supplemented with 10% fetal bovine serum, 100 units/mL penicillin, and 100 mg/mL streptomycin at 37℃ in a humidified atmosphere of 95% air and 5%CO2. The human breast cancer cell line MDA-MB-231 and its E1A/vector-stable transfectants have been described previously (58). The transfectants were grown under the same conditions as the controls,. except. that. G418. was. added. to. the. culture. medium.. MDA-MB-231/E1A cells were transfected with GRP78-expressing vector using Lipofectamine LTX (Invitrogen). 24 hours after transfection, cells were trypsinized and replated in DMEM/F12 with 10% fetal bovine serum and blasticidin. (5. g/mL;. invitrogen).. Blasticidin-resistant. clones. (231/E1A-GRP78) were selected. 2.2 RNA Isolation and RT-PCR Total RNA was extracted from cultured cells using the TRIzol Reagent (Invitrogen). An aliquot of 1-5 g of total RNA was subjected to RT-PCR using reverse transcriptase kit (Invitrogen). Equal amount of cDNA was used to perform PCR. The sequences of GRP78 and gp78 along with internal reference GAPDH primers are as follows: GRP78 forward, 5’-ATGAAAGAAACCGCTGAGGC-3’; reverse, 5’-CCAGGTCAAACACCAGGATG-3’; gp78. forward, 5’-ATGAAAGAAACCGCTGAGGC-3’; 5.
(12) reverse, 5’-CCAGGTCAAACACCAGGATG-3’; GAPDH forward, 5’-ACCACAGTCCATGCCATCAC-3’; reverse, 5’-ATGTCGTTGTCCCACCACCT-3’. After initial denaturation at 95℃ for 5 minutes, cycling parameters were as follows : GRP78 ─ denaturation (95℃, 30 second), annealing (55℃, 30 second), and extension (72℃, 30 second); the reaction included amplification 30 cycles; gp78 ─ denaturation (95℃, 30 second), annealing (52℃, 30 second), and extension (72℃, 30 second); the reaction included amplification 30 cycles; GAPDH ─ denaturation (94℃, 30 second), annealing (55℃, 30 second), and extension (72℃, 30 second); the reaction included amplification 25 cycles.. 2.3 Transwell migration and invasion assays For transwell migration assays, 1×105 or 5×104 cells were plated in the top chamber onto the non-coated membrane (24-well insert; pore size, 8 μm; Corning Costar, Corning, NY). For invasion assay, 1×105 or 5×104 cells were plated in the top chamber onto the Matrigel-coated membrane. Each well was coated freshly with Matrigel (30 μg; BD Bioscience) before the invasion assay. In both assays, cells were plated in medium without serum, and medium supplemented with serum was used as a chemoattractant in the lower chamber. The cells were incubated for 24 hours (migration assay) or 48 hours (invasion assay) and cells that did not migrate or invade through the pores were removed by a cotton swab. Cells on the lower surface of the membrane were fixed with methanol and stained with crystal violet. The number of cells migrating through or invading the membrane were counted under a light microscope (40X, there random fields per well). 6.
(13) 2.4 MTT assay The MTT assay are colorimetric assays for detecting the viable cells resulting from determining the activity of enzymes that reduce MTT (3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium. bromide,. a. yellow. tetrazole) to purple formazan in living cells. 1×104 cells were seeded into 96-well plates with 200ul of medium and each cell line was set up in 3 replicate wells. After incubation for 24 or 48 hours, the MTT reagent was added with final concentration of 1 ug/ul to each well and incubated for 3 hours. Removing medium, DMSO was added to dissolve the insoluble purple formazan product into a purple solution. The absorbance of this solution was measured at 575 nm by amiceoplate reader (BioTek).. 2.5 Cell tracing assay Cells were seeded onto 6 cm dish and grown in DMEM/ F12 supplemented with 10% fetal bovine serum. After 8 hours, cells set down a fluorescence microscope (Axioplan 2, Zeiss) equipped with a charge-couple device camera (Axiocam, Zeiss) and incubate at 37℃ in humidified 5% CO2. Cells were imaged every 15 min over a 18.5-h period and were quantified using image J.. 2.6 Western blotting Extracts were prepared in lysis buffer (50 mM Tris-HCl, 150mM NaCl, 1% Nonidet P-40, 0.5% Na-Deoxycholate, 2mM EDTA) containing protease inhibitor cocktail. Protein were heated in 4X sample buffer at 100℃ for 10 minutes and then loaded into each well for SDS-polyacrylamide gel 7.
(14) electrophoresis (SDS-PAGE). The separated protein was transferred to a PVDF membrane (Millipore) by all-wet method (BIO-RAD) under 88V for two hours. After blocking, blots were incubated with specific primary antibodies, and after washing and incubating with secondary antibodies (Santa Cruz, CA, USA), immunoreactive. proteins. were. visualized. using. an. enhanced. chemiluminescence detection system (Perkielmer). Followed by various primary antibody; GRP78 (Santa Cruz), gp78 (Santa Cruz), E1A (BD Bioscience), -actin (Sigma-Aldrich), -tubulin (Sigma-Aldrich). All primary antibodies were diluted to a recognition level (1:1000).. 2.7 Immunoprecipitation Cells were washed with ice-cold PBS and lysed in lysis buffer (50 mM Tris-HCl, 150mM NaCl, 1% Nonidet P-40, 0.5% Na-Deoxycholate, 2mM EDTA) containing protease inhibitor cocktail (PIC) and sodium orthovanadate (Na3VO4). Cell extracts were centrifugated at 13,000 rpm for 30 min. Lysates were incubated with protein A/G Sepharose beads (Santa Cruz, CA, USA) and then rotated for two hour at 4℃ (pre-clearing). Pre-clearing lysates were centrifugated at 3,000 rpm for 5 min. The supernatant was incubated with various antibodies and then rotated for three hour at 4℃. Next, lysates containing antibody were incubated with protein A/G Sepharose beads at 4℃ overnight. Immune complexes were washed three times with lysis buffer. Precipitates were resolved by SDS-PAGE, and the separated proteins were analyzed by western blot.. 2.8 Statistical analysis Data are analyzed as the mean ± SD. one-tailed Student’s t test was used to 8.
(15) analyze the difference between the means of the different groups. Differences with a p value of less than 0.05 were considered statistically significant.. 9.
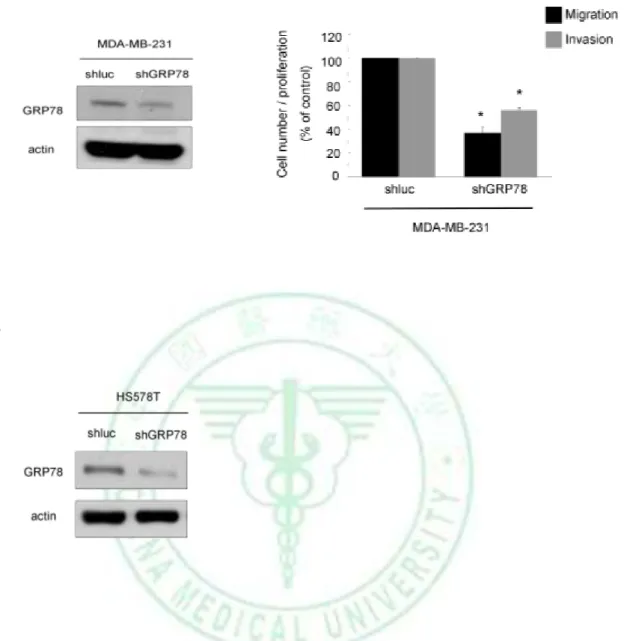
(16) 3. Results 3.1 GRP78 Protein Is Critical for E1A–mediated inhibition of cell mobility To investigate whether E1A affects the cancer metastatic activity, we transfected control vector or E1A expression vector into MDA-MB-231, HS578T and HBL100 cells. As expected, ectopic expression of E1A significantly reduce cell mobility and invasion activity in the vitro model (Fig. 1). To investigate the downstream target related to E1A-mediated anti-metastatic activity, we analyzed the differential expression of proteins in MDA-MB-231 breast cancer cells stably transfected with control vector (231/V) and. E1A. expression. vector. (231/E1A). using. two-dimensional. gel. electrophoresis assay. We found 3 candidate proteins were increased and 9 candidate proteins were decreased in 231/E1A cells compared with 231/V cells (Table. 1). Expression of GRP78 protein relates to E1A-mediated anti-metastasis activity and GRP78, one of the downregulated protein in 231/E1A cells, make us interesting to understand which mechanism may involoved. Recent researches demonstrated that overexpression of GRP78 confers antiapoptosis and chemoresistant to promote tumor survival and has also been implicated in proliferation and cell motility in different types of tumors (55, 59). In the previous report, GRP78 is a key survival factor of cancer cells, and reduction of GRP78 expression inhibits tumor formation, growth, and suppresses cancer cell metastasis in xenografts models (45-46, 60). In breast cancer patient, GRP78 expression is significantly higher in primary tumor compared with that in benign tissues (37). Because of the importance of GRP78 in cancer progression, we further determined whether GRP78 involves. 10.
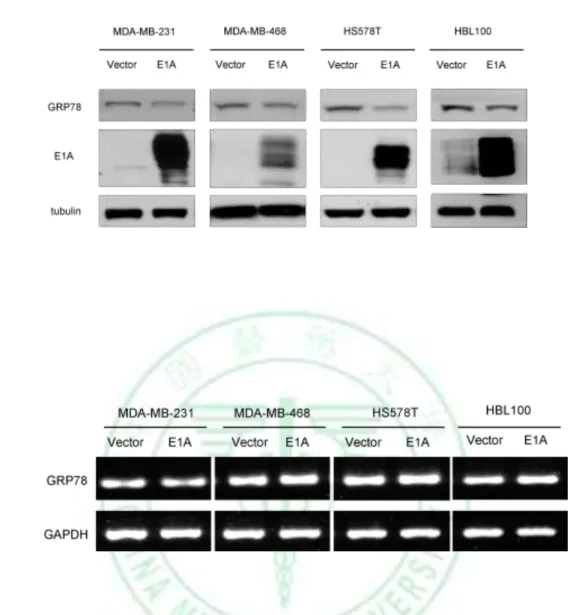
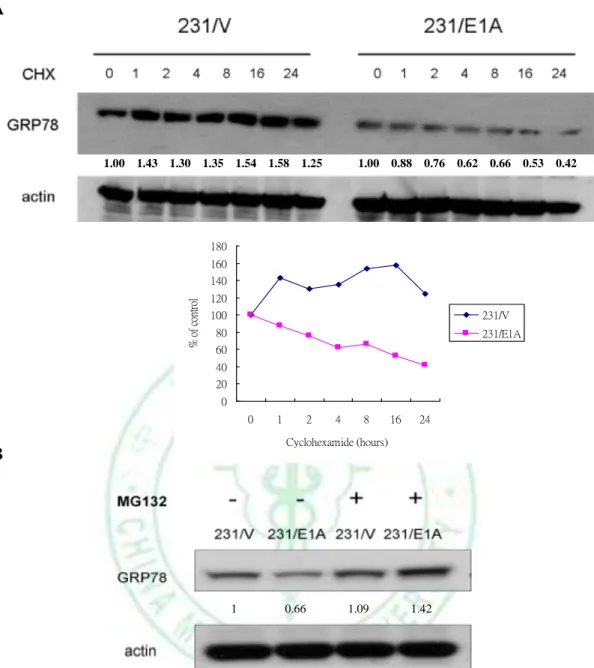
(17) in E1A-mediated tumor suppression activity. We examined the effects of E1A on GRP78 expression in various types of breast cancer cells and found that E1A suppresses the expression of GRP78 in protein levels but not mRNA level in E1A-transfected cells (Fig. 2). To define the role of GRP78 in breast cancer cells, we found that knockdown of GRP78 expression using GRP78 specific shRNA (shGRP78) significantly reduces migration and invasion ability compared with control shRNA in MDA-MB-231 and HS578T cells (Fig. 3). To investigate the effects of GRP78 on E1A-mediated suppression of cell mobility, we stably transfected GRP78 or control vector (pcDNA6) in 231/E1A cells (Fig. 4A). Transfection of 231/E1A cells with GRP78 showed markedly promoted cell migration and invasion by transwell assay and cell tracing assays (Fig. 4B, 4C). However, cancer metastasis involves multiple steps including detachment of the metastatic cells from neighboring cells, the acquisition of motility and invasion to other tissue. Among these steps, reorganization of the actin cytoskeleton is an important for cell mobility. To investigate the effect of GRP78 on actin stress fiber formation, we examined whether GRP78 participates in actin remodeling to enhance metastasis. These data suggest that GRP78 is critical for E1A-mediated inhibition of cell mobility in breast cancer cells.. 3.2 E1A Enhances Ub-dependent Proteolysis of GRP78 As shown in figure 2, E1A suppresses GRP78 in the protein level but not mRNA level. To determine the stability of GRP78 protein in response to E1A in MDA-MB-231, we treated with cycloheximide (CHX) for the indicated times in 231/V and 231/E1A cells. We found that the degradation rate of GRP78 protein is faster in 231/E1A cells than in 231/V cells (Fig. 5A). In 11.
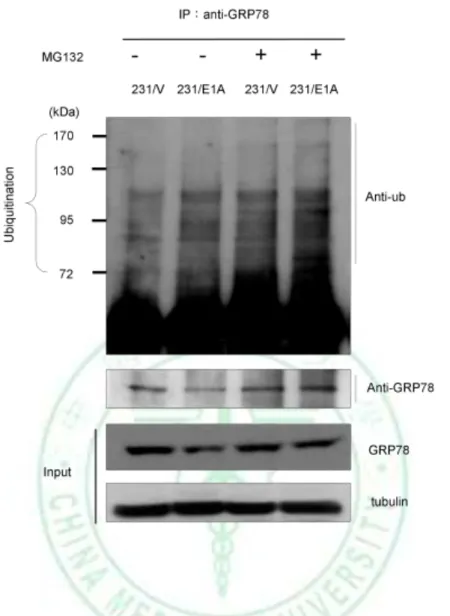
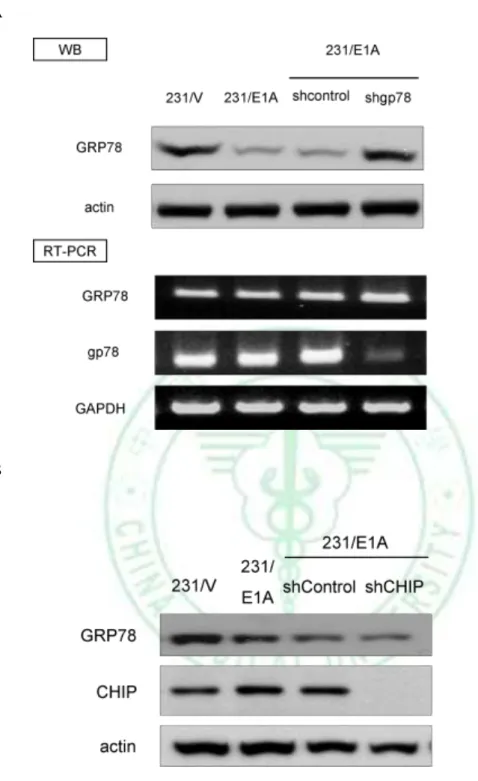
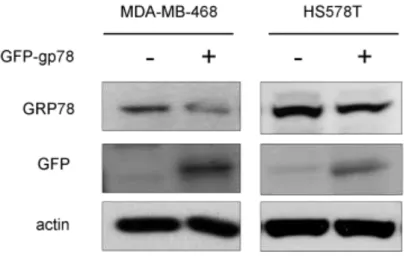
(18) attempt to determine degradation of GRP78 protein in response to E1A, we analyzed expression of endogenous GRP78 protein in presence with proteasome inhibitor such as MG132. Our results showed that E1A-mediated GRP78 downregulation was rescued by MG132 (Fig. 5B), suggesting that E1A-mediated GRP78 degradation via ubiquitin proteasome process. To confirm that the ubiquitination of GRP78 is enhanced by E1A, we detected the level. of. GRP78. ubiquitination. by. immunoblotting. (IB). after. immunoprecipitation (IP) of GRP78 or ubiquitin in231/V and 231/E1A cells. After IP with anti-GRP78 antibody and following by IB with anti-Ub, the ubiquitination of GRP78 was increased in the MG132-treated 231/E1A cells (Fig. 6). We showed that ubiquitin-dependent proteolysis of GRP78 is required for E1A-mediated GRP78 degradation, but which E3 ubiquitin ligase involves in this degradation process is unkown. Because of the ER localization of GRP78, we searched a series of ER-related E3 ubiquitin ligases such as gp78, Parkin, CHIP, Cul5 (61-63). Previous report showed that knockdown of gp78 increased the level of GRP78, and induced cell death in HEK 293 cells (64). Here, we found that knockdown of gp78 could revert protein expression of GRP78 but not mRNA level in 231/E1A cells (Fig. 7A). To further examine whether the expression of GRP78 was also suppressed by the other E3 ubiquitin ligases, we used shRNA against the expression of CHIP , which play roles in the degradation of protein in ER and reduces chaperone efficiency (65-68). We found that knockdown of CHIP had slight effect on E1A-mediated GRP78 degradation (Fig. 7B). To further investigate the effects of gp78 on GRP78 E3 ubiquitin ligase, we transfected with gp78 or control vector (pcDNA3.1) in MDA-MB-468 and HS578T, and found that gp78 suppresses the expression of GRP78 (Fig. 8). Above data indicated that gp78 involves in 12.
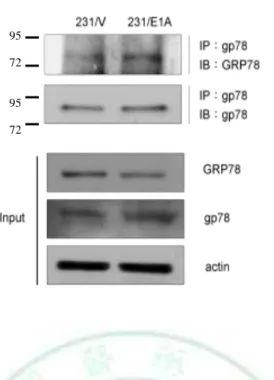
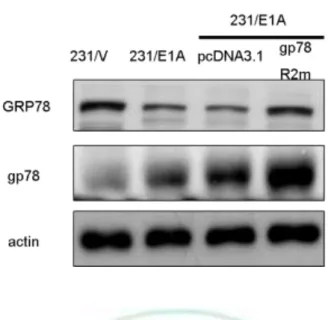
(19) E1A-mediated suppression of cancer cell migration and invasion activity through degradation of GRP78 expression.. 3.3 Gp78 Is Required for E1A-mediated Degradation of GRP78 and Suppression Cell Mobility To further define the relationship between gp78 and GRP78, coimmunoprecipitation assays demonstrated that gp78 physically interacted with GRP78 (Fig. 9), indicting that gp78 might serve as a GRP78 E3 ubiquitin ligase for GRP78. To further the E3 ligase activity of gp78 is required for e1A-induced GRP78 degradation, a double mutation of gp78 RING finger (gp78R2m) which loss of E3 ligase activity is used (69-71). Transfected with gp78R2m into 231/E1A inhibits E1A-induced degradation of GRP78 was inhibited (Fig.10). As gp78 is a RING finger-domain-containing E3 ligase that regulates ubiquitin-dependent degradation of its substrates, we further transiently transfected with either wild-type gp78 or gp78/R2m along with haemagglutinin (HA)-tagged ubiquitin. The level of GRP78 ubiquitination detected by immunoblotting after immunoprecipitation of GRP78 shows that ubiquitination of GRP78 was enhanced by wild-type but not gp78/R2m (Fig. 11). Because gp78 functions as an E3 ligase for GRP78 protein, we attempted to determine whether knockdown of gp78 affect cell mobility. Our data showed that knockdown of gp78 in rescue the expression of GRP78, resulting in increasing cell migration and invasion in 231/E1A cells (Fig. 12). Furthermore, knockdown of GRP78 re-suppressed cell migration and invasion in gp78-silenced 231/E1A cells (Fig. 12), suggesting that gp78 is required for E1A-mediated inhibition of cell migration and invasion through degradation of GRP78 expression. Take together, these results suggest that gp78-mediated 13.
(20) GRP78 degradation is critical for E1A’s anti-metastatic activity.. 14.
(21) 4.. Discussion In current study, we showed that E1A suppresses cancer cell mobility and. decreases expression of GRP78 protein in breast cancer cells. The previous data have been reported that GRP78 may enhance the activation of FAK which is correlation with mediating key signal transduction that regulation of actin remodeling (43, 72). We identified that GRP78 was degraded by E1A through the ubiquitin-proteasome system. Ubiquitin is covalently attached to target proteins through the action of three enzymes known as E1, E2, and E3. The ultimate outcome of this post-translational modification depends on the nature of the ubiquitin linkage and the extent of polyubiquitination. In most cases, ubiquitination results in degradation of the target proteins in the 26S proteasome. We show that the half-life of GRP78 protein expression was shorter in231/E1A cells compared with 231/V cells. GRP78 was rescued by treatment with MG132. Previous study showed that the GRP78 protein expression was increased in cells exposed to CHX (73), our data demonstrated that CHX indeed evaluate GRP78 expression in 231/V cells but not in 231/E1A cells, indicating that expression of E1A also attenuates GRP78 inducer. To investigate which E3 ubiquitin ligase involved in E1A-mediated degradation of GRP78, we silenced the expression of several ER-related E3 ubiquitin ligases by specific shRNA and found that knockdown of gp78 cause enhanced of GRP78 expression. However, we found that ectopic expression of E1A increased the association of gp78 with GRP78, following enhanced polyubiquitination and reduced GRP78 protein levels. Previous study indicated that both gp78 and CHIP E3 ligase effectively regulate hepatic CYP3A content (74). In our system, knockdown of CHIP expression had no effect on E1A-mediated GRP78 degradation, suggesting that gp78 serves as GRP78 15.
(22) specific E3 ligase. The domain of GRP78 for gp78 recognition and binding is unknown and some yet-to-be-identified protein candidate may be involved in the regulation of the gp78-GRP78 interaction. Based on our study, E1A-mediated gp78 ligase activity plays a role in the anti-cancer property through regulation of GRP78. The microRNAs (miRNA) are an extensive class of small noncoding RNAs (18-25 nucleotides) with important roles in the regulation of gene expression. Mostly, miRNA bind to the 3’ untranslated region (UTR) of message RNAs and negatively regulate the gene expression by inhibiting their translation. In this case, it has not been reported miRNA regulate the expression of GRP78. We predicted miRNAs binding to 3’UTR of GRP78 mRNA. However, we did not find miRNAs associated with regulation of GRP78 mRNA. In addition to metastasis, E1A gene therapy has been reported to induce sensitization to multiple anticancer drugs (75-77). Recent study shows that GRP78 has antiapoptosis activity and associates with drug resistance such as etoposide and Temozolomide (78). Therefore, we investigated whether E1A-mediating anti-cancer drug sensitivity through regulation of GRP78 expression. GRP78 was not involved in E1A-mediated sensitization of taxol, suggesting that E1A-mediated taxol sensitivity is requiring for the other mechanisms. In the other hand, GRP78 also plays important role in the maintenance of cancer stem cells population (79). We hope that E1A-mediated downregulation of GRP78 might be a potential therapeutic target for cancer cells resulting in eliminating cancer stem cells. In summary, we found that downregulation of GRP78 is critical for E1A-mediated inhibition metastasis. According to our finding, we provide a 16.
(23) model in which E1A represses GRP78 though induction of gp78-GRP78 interaction, which inhibits the breast cancer cells migration and invasion (Fig. 13).. 17.
(24) 5.. Reference. 1.. Jemal A, Siegel R, Xu J, Ward E. Cancer statistics, 2010. CA Cancer J. Clin. 2010 Sep-Oct;60(5):277-300. 2.. Nguyen DX, Massague J. Genetic determinants of cancer metastasis. Nat. Rev Genet. 2007 May;8(5):341-52. 3. Chalasani P, Downey L, Stopeck AT. Caring for the breast cancer survivor: a guide for primary care physicians. Am J Med. 2010 Jun;123(6):489-95. 4.. Nevins JR. Adenovirus E1A-dependent trans-activation of transcription.. Semin Cancer Biol. 1990 Feb;1(1):59-68. 5.. Nevins JR, Raychaudhuri P, Yee AS, Rooney RJ, Kovesdi I, Reichel R.. Transactivation by the adenovirus E1A gene. Biochem Cell Biol. 1988 Jun;66(6):578-83. 6.. Byrd PJ, Grand RJ, Gallimore PH. Differential transformation of primary. human embryo retinal cells by adenovirus E1 regions and combinations of E1A + ras. Oncogene. 1988 May;2(5):477-84. 7.. Ruley HE. Adenovirus early region 1A enables viral and cellular. transforming genes to transform primary cells in culture. Nature. 1983 Aug 18-24;304(5927):602-6. 8.. Moran E, Zerler B, Harrison TM, Mathews MB. Identification of separate. domains in the adenovirus E1A gene for immortalization activity and the activation of virus early genes. Mol Cell Biol. 1986 Oct;6(10):3470-80. 9.. Shenk T, Flint J. Transcriptional and transforming activities of the. adenovirus E1A proteins. Adv Cancer Res. 1991 Jan;57:47-85. 10. Shih JY, Tsai MF, Chang TH, Chang YL, Yuan A, Yu CJ, et al. Transcription repressor slug promotes carcinoma invasion and predicts outcome of patients with lung adenocarcinoma. Clin Cancer Res. 2005 Nov 18.
(25) 15;11(22):8070-8. 11. Frisch SM, Mymryk JS. Adenovirus-5 E1A: paradox and paradigm. Nat Rev Mol Cell Biol. 2002 Jun;3(6):441-52. 12. Yu DH, Scorsone K, Hung MC. Adenovirus type 5 E1A gene products act as transformation suppressors of the neu oncogene. Mol Cell Biol. 1991 Mar;11(3):1745-50. 13. Ueno NT, Yu D, Hung MC. E1A: tumor suppressor or oncogene? Preclinical and clinical investigations of E1A gene therapy. Breast Cancer. 2001 Jan;8(4):285-93. 14. Song CZ, Loewenstein PM, Toth K, Green M. Transcription factor TFIID is a direct functional target of the adenovirus E1A transcription-repression domain. Proc Natl Acad Sci U S A. 1995 Oct 24;92(22):10330-3. 15. Chen H, Hung MC. Involvement of co-activator p300 in the transcriptional regulation of the HER-2/neu gene. J Biol Chem. 1997 Mar 7;272(10):6101-4. 16. Geisberg JV, Chen JL, Ricciardi RP. Subregions of the adenovirus E1A transactivation domain target multiple components of the TFIID complex. Mol Cell Biol. 1995 Nov;15(11):6283-90. 17. Mazzarelli JM, Atkins GB, Geisberg JV, Ricciardi RP. The viral oncoproteins Ad5 E1A, HPV16 E7 and SV40 TAg bind a common region of the TBP-associated factor-110. Oncogene. 1995 Nov 2;11(9):1859-64. 18. Maguire K, Shi XP, Horikoshi N, Rappaport J, Rosenberg M, Weinmann R. Interactions between adenovirus E1A and members of the AP-1 family of cellular transcription factors. Oncogene. 1991 Aug;6(8):1417-22. 19. Yu D, Wolf JK, Scanlon M, Price JE, Hung MC. Enhanced c-erbB-2/neu expression in human ovarian cancer cells correlates with more severe malignancy that can be suppressed by E1A. Cancer Res. 1993 Feb 19.
(26) 15;53(4):891-8. 20. Yu D, Suen TC, Yan DH, Chang LS, Hung MC. Transcriptional repression of the neu protooncogene by the adenovirus 5 E1A gene products. Proc Natl Acad Sci U S A. 1990 Jun;87(12):4499-503. 21. Shao R, Tsai EM, Wei K, von Lindern R, Chen YH, Makino K, et al. E1A inhibition of radiation-induced NF-kappaB activity through suppression of IKK activity and IkappaB degradation, independent of Akt activation. Cancer Res. 2001 Oct 15;61(20):7413-6. 22. de Stanchina E, McCurrach ME, Zindy F, Shieh SY, Ferbeyre G, Samuelson AV, et al. E1A signaling to p53 involves the p19(ARF) tumor suppressor. Genes Dev. 1998 Aug 1;12(15):2434-42. 23. Timofeev OV, Pospelov VA. [The p21(WAF1) cyclin-kinase inhibitor: in vivo interaction with E1A adenoviral Ad2 and Ad12 oncoproteins]. Tsitologiia. 2003 May;45(11):1109-18. 24. Grooteclaes ML, Frisch SM. Evidence for a function of CtBP in epithelial gene regulation and anoikis. Oncogene. 2000 Aug 3;19(33):3823-8. 25. Bernhard EJ, Hagner B, Wong C, Lubenski I, Muschel RJ. The effect of E1A transfection on MMP-9 expression and metastatic potential. Int J Cancer. 1995 Mar 3;60(5):718-24. 26. Santoro M, Battaglia C, Zhang L, Carlomagno F, Martelli ML, Salvatore D, et al. Cloning of the rat tissue inhibitor of metalloproteinases type 2 (TIMP-2) gene: analysis of its expression in normal and transformed thyroid cells. Exp Cell Res. 1994 Aug;213(2):398-403. 27. Hortobagyi GN, Hung MC, Lopez-Berestein G. A Phase I multicenter study of E1A gene therapy for patients with metastatic breast cancer and epithelial ovarian cancer that overexpresses HER-2/neu or epithelial ovarian 20.
(27) cancer. Hum Gene Ther. 1998 Aug 10;9(12):1775-98. 28. Hortobagyi GN, Ueno NT, Xia W, Zhang S, Wolf JK, Putnam JB, et al. Cationic liposome-mediated E1A gene transfer to human breast and ovarian cancer cells and its biologic effects: a phase I clinical trial. J Clin Oncol. 2001 Jul 15;19(14):3422-33. 29. Villaret D, Glisson B, Kenady D, Hanna E, Carey M, Gleich L, et al. A multicenter phase II study of tgDCC-E1A for the intratumoral treatment of patients with recurrent head and neck squamous cell carcinoma. Head Neck. 2002 Jul;24(7):661-9. 30. Madhusudan S, Tamir A, Bates N, Flanagan E, Gore ME, Barton DP, et al. A multicenter Phase I gene therapy clinical trial involving intraperitoneal administration of E1A-lipid complex in patients with recurrent epithelial ovarian cancer overexpressing HER-2/neu oncogene. Clin Cancer Res. 2004 May 1;10(9):2986-96. 31. Lee AS. The glucose-regulated proteins: stress induction and clinical applications. Trends Biochem Sci. 2001 Aug;26(8):504-10. 32. Ni M, Lee AS. ER chaperones in mammalian development and human diseases. FEBS Lett. 2007 Jul 31;581(19):3641-51. 33. Lievremont JP, Rizzuto R, Hendershot L, Meldolesi J. BiP, a major chaperone protein of the endoplasmic reticulum lumen, plays a direct and important role in the storage of the rapidly exchanging pool of Ca2+. J Biol Chem. 1997 Dec 5;272(49):30873-9. 34. Quinones QJ, de Ridder GG, Pizzo SV. GRP78: a chaperone with diverse roles. beyond. the. endoplasmic. reticulum.. Histol. Histopathol.. 2008. Nov;23(11):1409-16. 35. Lee AS. GRP78 induction in cancer: therapeutic and prognostic 21.
(28) implications. Cancer Res. 2007 Apr 15;67(8):3496-9. 36. Wang Q, He Z, Zhang J, Wang Y, Wang T, Tong S, et al. Overexpression of endoplasmic reticulum molecular chaperone GRP94 and GRP78 in human lung cancer tissues and its significance. Cancer Detect Prev. 2005 Nov;29(6):544-51. 37. Fernandez PM, Tabbara SO, Jacobs LK, Manning FC, Tsangaris TN, Schwartz AM, et al. Overexpression of the glucose-regulated stress gene GRP78 in malignant but not benign human breast lesions. Breast Cancer Res Treat. 2000 Jan;59(1):15-26. 38. Gazit G, Lu J, Lee AS. De-regulation of GRP stress protein expression in human. breast. cancer. cell. lines.. Breast. Cancer. Res. Treat.. 1999. Mar;54(2):135-46. 39. Zhang J, Jiang Y, Jia Z, Li Q, Gong W, Wang L, et al. Association of elevated GRP78 expression with increased lymph node metastasis and poor prognosis in patients with gastric cancer. Clin Exp Metastasis. 2006 Dec;23(7-8):401-10. 40. Pootrakul L, Datar RH, Shi SR, Cai J, Hawes D, Groshen SG, et al. Expression of stress response protein Grp78 is associated with the development of castration-resistant prostate cancer. Clin Cancer Res. 2006 Oct 15;12(20 Pt 1):5987-93. 41. Xing X, Lai M, Wang Y, Xu E, Huang Q. Overexpression of glucose-regulated protein 78 in colon cancer. Clin Chim Acta. 2006 Feb;364(1-2):308-15. 42. Shuda M, Kondoh N, Imazeki N, Tanaka K, Okada T, Mori K, et al. Activation of the ATF6, XBP1 and grp78 genes in human hepatocellular carcinoma:. a. possible. involvement 22. of. the. ER. stress. pathway in.
(29) hepatocarcinogenesis. J Hepatol. 2003 May;38(5):605-14. 43. Su R, Li Z, Li H, Song H, Bao C, Wei J, et al. Grp78 promotes the invasion of hepatocellular carcinoma. BMC Cancer. 2010 Jan;10:20. 44. Fu Y, Wey S, Wang M, Ye R, Liao CP, Roy-Burman P, et al. Pten null prostate tumorigenesis and AKT activation are blocked by targeted knockout of ER chaperone GRP78/BiP in prostate epithelium. Proc Natl Acad Sci U S A. 2008 Dec 9;105(49):19444-9. 45. Fu Y, Lee AS. Glucose regulated proteins in cancer progression, drug resistance and immunotherapy. Cancer Biol Ther. 2006 Jul;5(7):741-4. 46. Jamora C, Dennert G, Lee AS. Inhibition of tumor progression by suppression of stress protein GRP78/BiP induction in fibrosarcoma B/C10ME. Proc Natl Acad Sci U S A. 1996 Jul 23;93(15):7690-4. 47. Misra UK, Deedwania R, Pizzo SV. Activation and cross-talk between Akt, NF-kappaB, and unfolded protein response signaling in 1-LN prostate cancer cells consequent to ligation of cell surface-associated GRP78. J Biol Chem. 2006 May 12;281(19):13694-707. 48. Finley D. Recognition and processing of ubiquitin-protein conjugates by the proteasome. Annu Rev Biochem. 2009 Jun;78:477-513. 49. Crews CM. Feeding the machine: mechanisms of proteasome-catalyzed degradation of ubiquitinated proteins. Curr Opin Chem Biol. 2003 Oct;7(5):534-9. 50. Jung T, Catalgol B, Grune T. The proteasomal system. Mol Aspects Med. 2009 Aug;30(4):191-296. 51. Sorokin AV, Kim ER, Ovchinnikov LP. Proteasome system of protein degradation and processing. Biochemistry (Mosc). 2009 Dec;74(13):1411-42. 52. Bouska A, Lushnikova T, Plaza S, Eischen CM. Mdm2 promotes genetic 23.
(30) instability and transformation independent of p53. Mol Cell Biol. 2008 Aug;28(15):4862-74. 53. Huang LE, Gu J, Schau M, Bunn HF. Regulation of hypoxia-inducible factor 1alpha is mediated by an O2-dependent degradation domain via the ubiquitin-proteasome pathway. Proc Natl Acad Sci U S A. 1998 Jul 7;95(14):7987-92. 54. Kim Y, Lillo AM, Steiniger SC, Liu Y, Ballatore C, Anichini A, et al. Targeting heat shock proteins on cancer cells: selection, characterization, and cell-penetrating properties of a peptidic GRP78 ligand. Biochemistry. 2006 Aug 8;45(31):9434-44. 55. Pyrko P, Schonthal AH, Hofman FM, Chen TC, Lee AS. The unfolded protein response regulator GRP78/BiP as a novel target for increasing chemosensitivity. in. malignant. gliomas.. Cancer. Res.. 2007. Oct. 15;67(20):9809-16. 56. Martin S, Hill DS, Paton JC, Paton AW, Birch-Machin MA, Lovat PE, et al. Targeting GRP78 to enhance melanoma cell death. Pigment Cell Melanoma Res. 2010 Oct;23(5):675-82. 57. Dong D, Ni M, Li J, Xiong S, Ye W, Virrey JJ, et al. Critical role of the stress chaperone GRP78/BiP in tumor proliferation, survival, and tumor angiogenesis in transgene-induced mammary tumor development. Cancer Res. 2008 Jan 15;68(2):498-505. 58. Meric F, Liao Y, Lee WP, Pollock RE, Hung MC. Adenovirus 5 early region 1A does not induce expression of the ewing sarcoma fusion product EWS-FLI1 in breast and ovarian cancer cell lines. Clin Cancer Res. 2000 Oct;6(10):3832-6. 59. Lee E, Nichols P, Groshen S, Spicer D, Lee AS. GRP78 as potential 24.
(31) predictor for breast cancer response to adjuvant taxane therapy. Int J Cancer. 2011 Feb 1;128(3):726-31. 60. Wang M, Wey S, Zhang Y, Ye R, Lee AS. Role of the unfolded protein response regulator GRP78/BiP in development, cancer, and neurological disorders. Antioxid Redox Signal. 2009 Sep;11(9):2307-16. 61. Chen C, Seth AK, Aplin AE. Genetic and expression aberrations of E3 ubiquitin ligases in human breast cancer. Mol Cancer Res. 2006 Oct;4(10):695-707. 62. Cyr DM, Hohfeld J, Patterson C. Protein quality control: U-box-containing E3 ubiquitin ligases join the fold. Trends Biochem Sci. 2002 Jul;27(7):368-75. 63. Soucy TA, Smith PG, Rolfe M. Targeting NEDD8-activated cullin-RING ligases for the treatment of cancer. Clin Cancer Res. 2009 Jun 15;15(12):3912-6. 64. Ying Z, Wang H, Fan H, Zhu X, Zhou J, Fei E, et al. Gp78, an ER associated E3, promotes SOD1 and ataxin-3 degradation. Hum Mol Genet. 2009 Nov 15;18(22):4268-81. 65. Ballinger CA, Connell P, Wu Y, Hu Z, Thompson LJ, Yin LY, et al. Identification of CHIP, a novel tetratricopeptide repeat-containing protein that interacts with heat shock proteins and negatively regulates chaperone functions. Mol Cell Biol. 1999 Jun;19(6):4535-45. 66. Murata S, Minami Y, Minami M, Chiba T, Tanaka K. CHIP is a chaperone-dependent E3 ligase that ubiquitylates unfolded protein. EMBO Rep. 2001 Dec;2(12):1133-8. 67. Connell P, Ballinger CA, Jiang J, Wu Y, Thompson LJ, Hohfeld J, et al. The co-chaperone CHIP regulates protein triage decisions mediated by heat-shock proteins. Nat Cell Biol. 2001 Jan;3(1):93-6. 25.
(32) 68. McDonough H, Patterson C. CHIP: a link between the chaperone and proteasome systems. Cell Stress Chaperones. 2004 Apr;8(4):303-8. 69. Fang S, Ferrone M, Yang C, Jensen JP, Tiwari S, Weissman AM. The tumor autocrine motility factor receptor, gp78, is a ubiquitin protein ligase implicated in degradation from the endoplasmic reticulum. Proc Natl Acad Sci U S A. 2001 Dec 4;98(25):14422-7. 70. Ballar P, Shen Y, Yang H, Fang S. The role of a novel p97/valosin-containing protein-interacting motif of gp78 in endoplasmic reticulum-associated. degradation.. J. Biol. Chem.. 2006. Nov. 17;281(46):35359-68. 71. Chen B, Mariano J, Tsai YC, Chan AH, Cohen M, Weissman AM. The activity of a human endoplasmic reticulum-associated degradation E3, gp78, requires its Cue domain, RING finger, and an E2-binding site. Proc Natl Acad Sci U S A. 2006 Jan 10;103(2):341-6. 72. Gerthoffer WT, Gunst SJ. Invited review: focal adhesion and small heat shock proteins in the regulation of actin remodeling and contractility in smooth muscle. J Appl Physiol. 2001 Aug;91(2):963-72. 73. Faria. G,. Cardoso. CR,. Larson. RE,. Silva. JS,. Rossi. MA.. Chlorhexidine-induced apoptosis or necrosis in L929 fibroblasts: A role for endoplasmic. reticulum. stress.. Toxicol. Appl. Pharmacol.. 2009. Jan. 15;234(2):256-65. 74. Kim SM, Acharya P, Engel JC, Correia MA. Liver cytochrome P450 3A ubiquitination in vivo by gp78/autocrine motility factor receptor and C terminus of Hsp70-interacting protein (CHIP) E3 ubiquitin ligases: physiological and pharmacological relevance. J Biol Chem. 2010 Nov 12;285(46):35866-77. 26.
(33) 75. Ueno. NT,. Yu. D,. Hung. MC.. Chemosensitization. of. HER-2/neu-overexpressing human breast cancer cells to paclitaxel (Taxol) by adenovirus type 5 E1A. Oncogene. 1997 Aug 18;15(8):953-60. 76. Liao Y, Zou YY, Xia WY, Hung MC. Enhanced paclitaxel cytotoxicity and prolonged animal survival rate by a nonviral-mediated systemic delivery of E1A gene in orthotopic xenograft human breast cancer. Cancer Gene Ther. 2004 Sep;11(9):594-602. 77. Sakakibara A, Tsukuda M, Kondo N, Ishiguro Y, Kimura M, Fujita K, et al. Examination of the optimal condition on the in vitro sensitivity to telomelysin in head and neck cancer cell lines. Auris Nasus Larynx. 2011 Oct;38(5):589-99. 78. Virrey JJ, Dong D, Stiles C, Patterson JB, Pen L, Ni M, et al. Stress chaperone GRP78/BiP confers chemoresistance to tumor-associated endothelial cells. Mol Cancer Res. 2008 Aug;6(8):1268-75. 79. Wu MJ, Jan CI, Tsay YG, Yu YH, Huang CY, Lin SC, et al. Elimination of head and neck cancer initiating cells through targeting glucose regulated protein78 signaling. Mol Cancer. 2010 Oct;9:283.. 27.
(34) 6. Figure A. B. Figure 1. E1A mediated inhibition of cell migration and invasion in malignant breast cancer cells. A, different breast cancer cells were transfected with E1A. 48h after transfection, cancer cell migration and invasion were confirmed by transwell migration and matrigel invasion assay. B, quantitative analysis of cell numbers was shown. Columns, migration or invasion activity compared with that of vector control; bars, SD. Each assay was done in three independent experiments. *, P < 0.05, between breast cancer cells transfected with vector or E1A. 28.
(35) A. B. Figure 2. E1A suppresses the expression of GRP78 in protein levels but not mRNA level in E1A-transfected cells. The different breast cancer cells were transfected with E1A. 48h after transfection, the total lysates of these cells were prepared. A, E1A and GRP78 protein expression were confirmed by western blotting. -tubulin was used as the internal protein loading control. The E1A protein is identified as a doublet at ~35-46 kDa. B, The expression of GRP78 mRNA was determined by RT-PCR. GAPDH served as an internal control. The PCR product size of GRP78 is ~300 b.p.. 29.
(36) A. B. Figure 3. Knockdown of GRP78 expression significantly reduces migration and invasion ability. Left, (A) MDA-MB-231 and (B) HS578T transfected with shluc or shGRP78 were analyzed by western blotting. -actin was used as the internal protein loading control. Transwell migration assay and matrigel invasion assay of (A) MDA-MB-231 and (B) HS578T transfected with shluc or shGRP78. Columns, migration or invasion activity compared with that of MDA-MB-231 and HS578T transfected with shluc. *, P < 0.05, between MDA-MB-231 and HS578T transfected with shluc.. 30.
(37) A. B. C. 31.
(38) Figure 4.. GRP78 may contribute to E1A-mediated invasion suppression activity.. A, 231/E1A cells stably transfected with GRP78 or control vector were analyzed by western blotting. -actin was used as the internal protein loading control. B, In 231/V, 231/E1A and 231/E1A stably transfected with control vector and GRP78 were analyzed for transwell migration assay and matrigel invasion assay. Columns, migration or invasion activity compared with that of 231/V; bars, SD. Each assay was done in three independent experiments.. *, P < 0.05, between 231/V and. 231/E1A cells. #, P < 0.05, between 231/E1A cells stably transfected with vector control and GRP78. C, Cell mobility were confirmed by cell tracing assays in 231/V, 231/E1A and 231/E1A stably transfected with control vector and GRP78; bar, SD. n=20. 32.
(39) A. 1.00 1.43 1.30 1.35 1.54 1.58 1.25. 1.00 0.88 0.76. 0.62 0.66 0.53. 0.42. 180 160. % of control. 140 120 100. 231/V. 80. 231/E1A. 60 40 20 0 0. 1. 2. 4. 8. 16. 24. Cyclohexamide (hours). B. 1. 0.66. 1.09. 1.42. Figure 5. E1A enhances proteolysis of GRP78 and its effect was rescued by MG132. A, Determination of protein stability of GRP78 in 231/Vand 231/E1A cells. The 231/V and 231/E1A cells were treated with 50 M cycloheximide (CHX) for the indicated times and GRP78 expression were analyzed by western blotting. Folds of protein expression are shown below the lanes and .quantified (bottom). B, Cells were treated with or without the proteasome inhibitor MG132 (5 M) for 12 hours, and GRP78 expression were analyzed by western blotting Folds of protein expression are shown below the lanes and the expression levels of each protein compared with 231/V. 33.
(40) Figure 6. The ubiquitin-dependent proteolysis of GRP78 is required for E1A-mediated GRP78 degradation. 231 cells stably expressed E1A were treated with either DMSO (control) or the proteasome inhibitor MG132 (5 M) for 12 hours. The total lysates of these cells were prepared and Immunoprecipitation (IP) was performed as described above and followed by Immunoblotting (IB) with an anti-Ub Ab. The input IB data were performed with Abs against GRP78 and -tubulin as described.. 34.
(41) A. B. Figure 7. Expression of GRP78 was rescued by knockdown of E3 ligase, gp78 but not CHIP in protein level. A, 231/E1A transfected shRNA of gp78 were analyzed for GRP78 expression by western blotting or RT-PCR. B, 231/E1A transfected shRNA of CHIP were analyzed for GRP78 expression by western blotting,. 35.
(42) Figure 8. gp78 suppresses the expression of GRP78. The different breast cancer cells were transfected with gp78. 48h after transfection, GFP and GRP78 expression were confirmed by western blotting.. 36.
(43) M. W 95 72 95 72. Figure 9. gp78 physically interacted with GRP78. 231/V and 231/E1A cells were treated with the proteasome inhibitor MG132 (5 M) for 12 hours. The total lysates of these cells were prepared and immunoprecipitated with anti-gp78 antibody and followed by IB with an anti-GRP78 antibody. The input IB data were performed with Abs against GRP78, gp78, and actin as described.. 37.
(44) Figure 10. E1A-mediated downregulation of GRP78 is required for gp78’s E3 ubiquitin ligase activity. 231/E1A transfected with vector control (pcDNA3.1) or RING finger mutated gp78 (gp78R2m) were analyzed by western blotting.. 38.
(45) Figure 12. Gp78 regulates ubiquitination of GRP78 protein. Wild-type or mutant of gp78 were expressed in 293T cells along with GRP78 and HA-ubiquitin (ub). 48 hr post transfection, cells were treated with MG132 (5M) for 12 hr and levels of GRP78 ubiquitination were evaluated by immunoprecipitation of GRP78 using anti-GRP78 antibody followd by anti-HA immunoblotting. 39.
(46) A. B. 40.
(47) Figure 12. Knockdown of GRP78 re-suppressed cell migration and invasion in 231/E1A with knockdown of gp78. A, 231/E1A transfected with shcontrol (231/E1A-shcontrol) or shgp78 (231/E1A-shgp78) and 231/E1A-shgp78 transfected with shluc or shGRP78 were analyzed by western blotting. B, Transwell migration assay and matrigel invasion assay of these cells. Columns, migration or invasion activity compared with that of 231/V; bars, SD. Each assay was done in two independent experiments. *, P < 0.05.. 41.
(48) Figure 13. E1A represses GRP78 through induction of gp78-GRP78 interaction, which inhibits the breast cancer cells migration and invasion. 42.
(49) The protein were upregulated in 231/E1A cells compared with 231/V cells prot site 24. prot describe Lonp1 Lon protease homolog. 588. Desmoplakin. 692. Prohibitin. The protein were downregulated in 231/E1A cells compared with 231/V cells. prot site. prot describe. 82. Hspa5 78 kDa glucose-regulated protein. 284. Lmna Isoform C of Lamin-A/C. 486. Alpha-enolase. 523. Vim Vimentin. 588. Cops4 COP9 signalosome complex subunit 4. 621 724. Isocitrate dehydrogenase [NAD] subunit alpha, mitochondria Anxa2 Annexin A2. 730. Pyruvate dehydrogenase E1 component subunit beta. 954. Prdx4 Peroxiredoxin-4. Table 1. Differential expressions of protein in 231/E1A cells versus 231/V cells were analyzed by Two-dimensional gel electrophoresis.. 43.
(50)
數據


+7
相關文件
Milk and cream, in powder, granule or other solid form, of a fat content, by weight, exceeding 1.5%, not containing added sugar or other sweetening matter.
Center for Faculty Development, Department of Medical Education, Taichung Veterans General Hospital 1 Department of Nursing, Hungkuang University, Taichung, Taiwan 2. Section
Center for Faculty Development, Department of Medical Education, Taichung Veterans General Hospital 2 School of Medicine, National Yang-Ming University, Taipei, Taiwan 3.
We present numerical results for Algorithm Elastic-Inexact applied to a set of 18 MPECs, showing the outcomes for each problem and analyzing whether exact complementarity,
Center for Faculty Development, Department of medical education, Taichung Veterans General hospital 1 School of Medicine, National Yang-Ming University, Taipei, Taiwan 2.
臺大機構典藏NTUR (National Taiwan University 二 Repository, http://ntur.lib.ntu.edu.tw) 經驗與協助推 動臺灣學術機構典藏TAIR (Taiwan Academic Institutional Repository,
† Institute of Applied Mathematical Sciences, NCTS, National Taiwan University, Taipei 106, Taiwan.. It is also important to note that we obtain an inequality with exactly the
Department of Physics, National Chung Hsing University, Taichung, Taiwan National Changhua University of Education, Changhua, Taiwan. We investigate how the surface acoustic wave
