行政院國家科學委員會專題研究計畫 成果報告
水稻根部低溫耐性分子機制之探討﹝3/3﹞
計畫類別: 個別型計畫 計畫編號: NSC92-2313-B-002-006- 執行期間: 92 年 08 月 01 日至 93 年 07 月 31 日 執行單位: 國立臺灣大學農藝學系暨研究所 計畫主持人: 張孟基 計畫參與人員: 陳維平 報告類型: 完整報告 處理方式: 本計畫可公開查詢中 華 民 國 93 年 11 月 2 日
行政院國家科學委員會補助專題研究計畫成果報告
※※※※※※※※※※※※※※※※※※※※※※※※※※
※ ※
※ 水稻根部低溫耐性分子機制之探 ※
※ ※
※※※※※※※※※※※※※※※※※※※※※※※※※※
計畫類別:▓個別型計畫 □整合型計畫
計畫編號:NSC 91-2313-B-002-259-
執行期間:
92 年 08 月 01 日至 93 年 07 月 31 日
計畫主持人:張孟基助理教授 共同主持人:盧虎生教授
計畫參與人員:二年級碩士生陳維平
本成果報告包括以下應繳交之附件:
□赴國外出差或研習心得報告一份
□赴大陸地區出差或研習心得報告一份
□出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份
執行單位:國立台灣大學農藝學系
中 華 民 國 93 年 10 月 28 日
行政院國家科學委員會專題研究計畫成果報告
水稻根部低溫耐性分子機制之探討
Molecular study of the cold-tolerance mechanism in the root of
rice (Oryza sativa L.)
計畫編號:NSC 92-2313-B-002-006-
執行期限:92 年 08 月 01 日至 93 年 07 月 31 日
主持人:張孟基助理教授 執行機構及單位名稱國立台灣大學農藝學系
共同主持人:盧虎生教授 執行機構及單位名稱國立台灣大學農藝學系
計畫參與人員:陳維平 執行機構及單位名稱國立台灣大學農藝學系
一、中文摘要 水稻經常 因遭受 非生 物性之生 理逆 境,如低溫而導致嚴重減產。因此極其重 要的是去瞭解水稻在低溫逆境下之耐冷機 制,尤其是其內部所有基因或蛋白質之表 現,找出與低溫反應相關之基因或蛋白 質。本計畫前二年已利用低溫下不同耐性 之水稻根部為材料,以 cDNA-AFLP 及蛋 白質雙向電泳等方法找出諸多在低溫下表 現之基因與蛋白質。本年之計畫持別針對 其中 3 個基因 ADF, porin-like protein 及 β-glucosidase 等,釣取其 cDNA 全長、決 定該基因於水稻基因體之考貝數及該基因 於不同非生物逆境,尤其是低温逆境之表 現。 結果發現 ADF 屬於水稻多基因家族, 共 有 9 個 isoforms. 對 不 同 生 物 性 逆 境 ADF-2 及 ADF-3 基因具差異性表現。另外 則發現porin-like protein 基因為一具陰離子 通道特性之膜孔類似蛋白質。在三個水稻 品種TNG67、IR36 和 PANGONIA 中可能 都僅有一個拷貝數。在台農67 號根部中經 過低溫處理後該基因之表現會立即被抑 制,而在24 小時後恢復。該基因表現直接 會受到高溫抑制。 此外本試驗以 β-glucosidase 之基因專 一性引子進行該基因全長之釣取,意外因 引子之相似性,選殖到 amine oxidase 基 因。經過南方墨點分析,發現此基因在水 稻基因體上可能僅有一個拷貝數。在北方 墨點分析中發現,此基因可受低溫處理所 誘導在一小時後大量表現,二小時後表現 降低,在 RT-PCR 中也可觀察到相似之結 果。另外在不同逆境處理後,在地上部和 根部表現之情形也有所不同。此基因參與 多元胺之代謝路徑,將多元胺降解後,會 造成過氧化氫和胺的累積。其與水稻低溫 耐受性之關係仍待闡明。
關鍵詞:水稻,根,低溫耐性. AbstractAbiotic stresses, such as low temperature frequently result in significant reduction of the potential yield of rice. Therefore, in order to understand the cold tolerance mechanism in rice, it is essential to evaluate and determine the genes/ or proteins expression profile in rice under chilling stress, and identify key transcripts or proteins that are associated with this process. In past two years, we have identified several differentially expressed genes or proteins in response to low temperature by cDNA-AFLP and 2D gel methods. In this study, we especially focused on three genes, including ADf, porin-like protein and β-glucosidase to determine their corresponding gene copy numbers and expression patterns under various abiotic stress.
belong to a multiple gene families that consisted of 9 genes. RT-PCR analysis with 3’-specific primer pairs showed that ADF-2 and ADF-3 genes respond differentially to various abiotic stresses. In addition, the porin – like protein gene displays the property which similar to the anion channel of membrane protein. After southern blotting analysis, we found it’s a single copy gene in rice cultivars – TNG67, TN1 and Pangonia. In TNG67 root, the expression of this gene shut down when low temperature started after one hour, but it expressed again after 24 hours. Under differential abiotic stress treatment, it shows the porin-like protein gene expression repressed under heat treatment in both tissues.
In order to get the full – length cDNA of beta – glucosidase gene, unexpectedly, due to the misannealing of primer pairs, we isolate another gene, which is amine oxidase. The southern blot analysis showed that there is only a single copy gene in the rice genome. After low temperature treatment, the expression of this gene was induced approximately two folds higher than the control. But after two hours, the expression reduced again. The gene may play an important role in polyamine degradation, that causes the H2O2 and NH3 accumulation.
The coreelation between amine oxidase gene expression and rice cold tolerance would require further study.
Keywords: Rice, Root, Cold-tolerance, Actin depolymerization factor (ADF), porin-like protein, beta-glucosidase, amine oxidase.
二、緣由與目的 水稻(Oryza Sativa. L)為亞洲及台灣多數人 賴以維生的重要作物. 但在早春的幼苗期 經常遭受寒害而導致產量受損。因此對於 作物如何獲致低溫耐性之分子機制的瞭 解,一直是植物生理學、作物育種學上所 關心的共同研究課題。其研究結果亦可提 供未來對低溫具耐性之農作物在分子育種 上的考量。要達到此目的,極其重要的是 去研究植物在低溫逆境下其內部所有基因 之表現,找出與低溫反應相關之基因,並 檢測該基因如何在生理功能上參與低溫耐 性之調控. 本研究先前已利用基因差異顯 示法、PCR-selected cDNA substraction 及 cDNA-AFLP 及 蛋 白 質 體 學 (proteomics) 2D-gel 之分析,分別從對低溫具耐性(台農 67 號, TNG67) 及對低溫敏感(台中在來一 號, TN1) 之水稻根部,篩選出諸多低温反 應相関之基因及蛋白質。但此些基因在水 稻低温耐性生理機制上究竟扮演何種角色 極待闡明。是以在前述既有之基礎下,本 計劃擬進一步探討水稻根部中與低溫耐性 有 關 之 ADF, porin-like protein 及 beta-glucosidase 基因在低溫下之反應。 三、結果與討論 低溫反應相關基因在分子層次上之研究分析 1. ADF 本研究先前發現外加 ABA 於 TN1 時,可 誘導 ADF 蛋白質之表現。其中 ADF-1 之 表現於處理ABA 1 小時後即開始增加,而 ADF-2 則要至 3 小時後量才開始有變化, 但不論如何,兩者之蛋白質表現量皆隨 ABA 之處理時間而增加。此蛋白質的功能 為將F-actin 的(-)端之 actin 去除,這個動作 可使細胞骨架(cytoskelton)重組,在低溫逆 境下可使植物之細胞膜維持流動性,使細 胞仍能維持正常之生理功能。經搜尋水稻 (KOME、 TIGR、BGI, both for indica and japonica) 結果發現共有 9 個 ADF 之相似基 因。我們的屬 ADF-3.经由 DNA start 之軟 體程式建立其基因演化樹如圖 1a.並由設 計 3’ 端 之 gene-specific 引 子 進 行 RT-PCR 偵測 ADF-2 及 ADF-3 基因於不同逆 境之表現並不相同。 2. porin-like protein: 此差異性片段全長117bp,經資料庫比對後
為一個porin-like protein。對照 KOME 水稻 基因體資料庫,本試驗設計此基因之專一
性引子,進行RT-PCR,得到一個全長為
1018bp 之產物(附圖 1)。將之送交定序,
確認無誤後,繼續採用此段cDNA 作為探
針,進行南方和北方墨點分析及RT-PCR。
實驗結果顯示Porin-like protein gene 之南
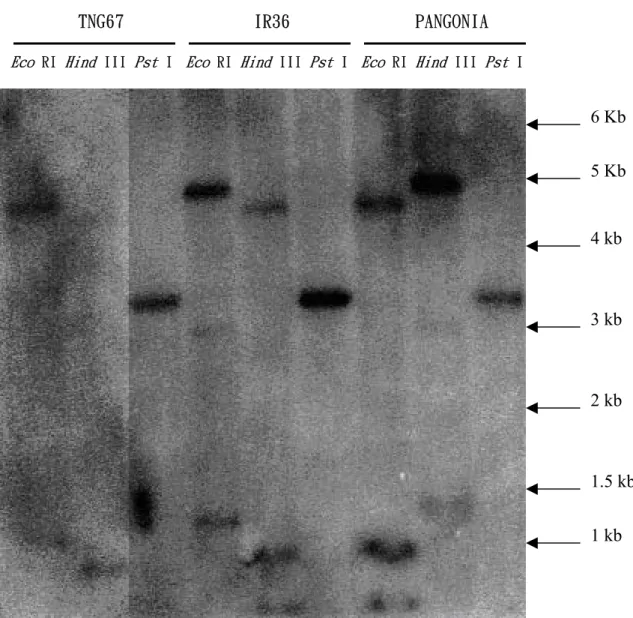
方墨點分析(圖3),在梗稻台農67 號中,
經過限制酵素 Eco RI 切割的處理出現了三 條條帶,經 Hind III 切割後的處理有兩條條 帶,位於上方之條帶並不明顯,而限制酵 素 Pst I 所切割的處理則只有一條條帶。在
秈稻 IR36 所觀察到的結果是經限制酵素
Eco RI 和 Hind III 切割的處理均出現了三
條條帶,位置和台農 67 號得到的結果類
似,而限制酵素 Pst I 所切割的處理則只有
一條條帶。大粒種PANGONIA 得到的結果
是條帶數目和秈稻 IR36 所得到的結果相
同,但是在經限制酵素 Eco RI 和 Hind III
切割的處理後的條帶位置明顯的與台農 67
號和IR36 不同,可能是品種上基因之多型
性(polymorphism)所致。經比對 cDNA 全長上之 Eco RI,Hind III 及 Pst I 切點推 測該基因在水稻基因體中可能僅有一個拷 貝。 爲了證明上述選殖到之低溫反應相關 性基因與水稻耐冷性之關係,本試驗利用 經過低溫不同時間點處理之水稻根部 total RNA 進行北方墨點分析或 RT-PCR,以證 明其和低溫耐性之關聯。本研究最早選用 一段Porin-like protein gene 1018bp 之 PCR 產物(其核苷酸序列全長及引子對應位置
如附圖 1 所示)作為探針,開始進行北方
墨點分析,但是其結果並不成功,因此改
用 RT-PCR 來證明其與低溫處理時間點,
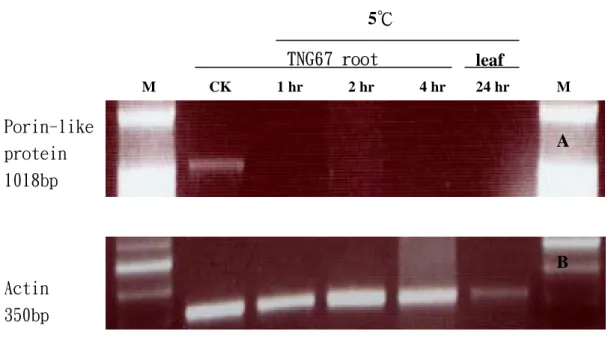
不 同 逆 境 處 理 和 組 織 專 一 性 等 關 係 。 Porin-like protein gene 在 RT-PCR 所得之結 果顯示開始低溫處理一小時後此基因之表 現隨即消失,在地上部低溫處理24 小時時 也出現相同情況(圖4,圖 5)。 3. β-glucosidase gene 此基因(其核苷酸序列之全長及引子 對應位置如附圖 2 所示),在剛開始進行 3’& 5’RACE,RT-PCR,北方和南方墨點 分析時,設計了一組位於 ORF 內之 gene specific 引子,但是在完成所有分析後,比 對定序結果才發現因為設計之引子在進行 RT-PCR 時做出了錯誤的基因片段,該基 因是putative amine oxidase。原先預期之產
物為 150bps,結果卻發現了一個為 250bp
之 cDNA 片段,經定序後發現在嘗試過許
多不同引子組合及PCR 條件後,此基因目
前無法得到正確之PCR 產物。
4. Amine Oxidase gene
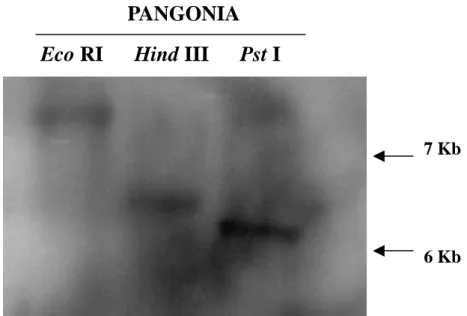
原先用來做β-glucosidase gene 之專一 性引子,序列和此基因不完全吻合,其中 SAR5’ 引 子 序 列 為 5’GGTGACGTGGCTCAAGAACC 3’,對應 在 amine oxidase 上 之 位 置 為 5’ CCCTGAACCGCTCAAGAACC 3’,其中 有6 個核苷酸序列不同,以劃線部份表示; SAR3’ 引 子 序 列 為 5’GTTCCTCCTGCCCACGCTGC,對應在 amine oxidase 上 之 位 置 為 5’GAAGATCCTGCCCAACGCTGC 3’ , 其中有 5 個核苷酸序列不同,以劃線部份 表示。雖然序列不吻合,但是每次進行PCR 反應即使調整黏合溫度至 57℃,均能做出 此一段約250bp 之產物(附圖 3),因此誤 判將之當作 β-glucosidase 基因,並使用這 段 250bp 之 cDNA 作為探針,開始進行了 南方和北方墨點分析,並使用該引子進行 RT-PCR 等差異性表現分析。 用梗稻台農 67 號,秈稻 IR36 與大粒 種 PANGONIA 等 三 種 不 同 的 水 稻 之 genomic DNA 為材料,每種 genomic DNA 再分別用 Eco RI,Hind III 和 Pst I 三種不 同之限制酵素做切割後,進行南方墨點分 析法,觀察amino oxidase gene 之表現(圖
6),結果只有大粒種水稻PANGONIA 有結 果,三個 lane 都只有出現一條明顯的條 帶,梗稻台農 67 號和秈稻 IR36 則完全沒 有訊號出現,就此推測該基因在水稻基因 體上僅為單一拷貝數。amine oxidase 之北 方墨點分析結果(圖7)則發現,該基因之 表現量與處理時間有關,5℃處理第一個小 時後,可明顯看出其表現量有增加,但是 第二個小時之後其表現量即隨之降低,其 表現量很明顯的較對照組和低溫處理一小 時來的低,第四小時也出現相同的情況。 此外以不同時間低溫處理之 RT-PCR 結果 (圖8)與北方墨點分析之結果互相驗證, 其結果和北方墨點分析結果類似,在經過 5℃低溫處理兩個小時後,其表現量及就隨 之降低,低溫處理四小時時似乎已看不到 該基因有表現。 由前人研究顯示,不同的非生物性逆 境之基因表現往往有cross-talk 之訊息傳遞 與基因調控之關係,如:Shinozaki 等人在 2003 發表,以水稻為模式植物,利用 DNA microarray 及 RNA-gel blot 證實在所發現之 73 個受到低溫,乾旱,高鹽及 ABA 之基
因彼此調控遠高於低溫與 ABA 或者是低
溫與高溫之cross talk。為更深入明瞭低溫
反應相關性基因之生理功能,除了探討該 些基因於低溫下之表現,亦需就其在水稻
不同組織之專一性表達及與其他非生物性 逆 境 間 做 一 研 究 。 是 以 我 們 採 取 了 RT-PCR,並於 TNG67 根(root)及地上部 (shoot)分別用 mannitol (滲透壓)、NaCl (高鹽)、ABA、CuCl2(重金屬)、4℃(低
溫)、45℃(高溫)等條件處理,以期明瞭
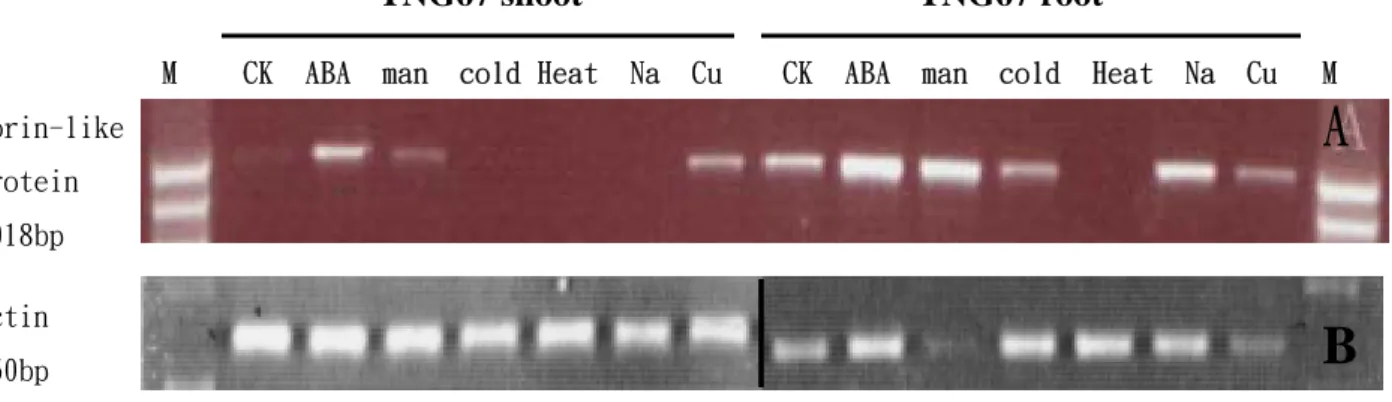
此些基因於低溫或其他逆境之作用。 結 果 發 現 在 水 稻 TNG67 之地 上部 中,porin-like protein 會被 ABA,mannitol
和 CuCl2等 3 種滲透壓逆境所誘導。但在 低溫及高溫處理後均被完全抑制。在根部 的表現和地上部有些許差異,水稻根部經 過 ABA 和 mannitol 等 兩 種 處 理 後 , porin-like protein 會被大量誘導。而經過低 溫處理後,porin-like protein 並無太大變 化。此外在高溫處理下則和地上部一樣, 此基因表現完全被抑制。經NaCl 處理後此 基因表現量增加,和地上部之表現不同。 而在 CuCl2 處理後,表現量會降低,但是 在地上部中,此基因表現情況則被完全抑 制(圖5)。 Amine oxidase 基因經不同逆境處理後 之表現,在水稻地上部中amine oxidase 基 因會被ABA,mannitol 及低溫逆境誘導增 加其表現量。尤其是在後兩種處理,表現 量大幅度提高。但是經高溫,NaCl 和 CuCl2 處理後,此基因之表現則完全被抑制。在 水稻根部中,ABA,mannitol 和低溫處理 都會誘導此基因表現量之增加,尤其以低 溫處理使其表現量增加幅度最大。經NaCl 和CuCl2處理後,此基因表現量持平,沒 有變化。而高溫處理後,此基因表現則被 抑制(圖9)。 四、計畫成果自評 本計畫之最終目標乃在闡明水稻根部低溫 耐性之分子機制,尤其是發現其中進行調 控 之 特 殊 基 因 或 蛋 白 質(specific master regulatory transcripts or proteins) 。目前本 研究室已初步的找到一些與低溫表現相關 的基因或蛋白質。對其中許多未知生理功 能但於低溫或 ABA 處理下表現有差異之 蛋白質,我們已利用 RT-PCR 之方式得到 與其相對應之cDNA clones,進一步探討此 些蛋白質、基因之表現與低溫耐性之關 係。此與原計畫先前提出之研究內容與進 度相符。在受低溫影響之基因表現上,我 們 已 開 始 以 基 因 轉 殖 之 方 式 (RNA interference 及 overexpression)來進行基因 功能之分析。其研究結果應可增進吾人對 水稻低温耐性之瞭解並提供未來對低溫具 耐性之農作物在分子育種上的考量。 五、參攷文獻
1. Boyer, J. S. (1982) Plant productivity and environment. Science 218, 443-448. 2. Chen, S. L., Lur, H. S., Yu, S. M., Chen,
H. H., and Chu, C. (2000) Identification of novel genes related to chilling tolerance in rice (Oryza sativa L.) seedlings by differential display. Journal of A. A. C. 6, 614-633.
3. Chu, C. and Lee, T. M. (1992) Regulation of chilling tolerance in rice seedlings by plant hormones. Korean. J. Crop. Sci. 37, 288-298.
4. Karine, G., Claudette, J., Steven, P. C. G., Magda, P., Hans, D., Joel, V., and Dominique., J. (2001) Proteomic analysis of Arabidopsis seed germination and priming. Plant Physiology 126, 835-848. 5. Lang, V., Heino, P., Palva, E.T. (1989)
Low temperature acclimation and treatment with exogenous abscisic acid induce common polypeptides: in
Arabidopsis thaliana (L.) Heynh. Theor
Appl genet 77, 729-734.
6. Lee, T. M., Lur, H. S. and Chu, C. (1995) Role of abscisic acid in chilling-tolerant rice (Oryza sativa L.).seedlings. I. Endogenous abscisic acid levels. Plant Cell Environ 16, 481-490.
7. Link, A.J. (1999) Direct analysis of protein complexes using mass spectrometry. Nature Biotechnology 17, 676-682.
8. Ruedi, A., Matthias, M. (2003) Mass spectrometry-based proteomics. Nature 422, 198-207.
9. Sung, D-Y., Kaplan, F., Lee, K-J., and Guy, C. L. (2003) Acquired tolerance to temperature extremes. Trends in Plant Sciernce 8(4), 179-187.
10.Thomashow, M. F. (1999) Plant cold acclimation: freezing tolerance genes and regulatory mechanisms. Annu. Rev. Plant.
Physiol. 50:571-599.
11.Wijk, K. J. (2001) Challenges and prospects of plant proteomics. Plant Physiology126, 501-508.
12. Wandrey, M., Trevaskis, B., Brewin, N., and Udvardi, M. K.(2004) Molecular and cell biology of a family of voltage-dependent anion channel porins in Lotus japonicus. Plant Physiol. 134: 1-12.
13. Robbani, M.A., Maruyama, K., Abe, H., Khan, M.A., Katsura, K., Ito, Y., Yoshiwara, K., Seki, M., Shinozaki, K., and Yamaguchi – Shinozaki, K. (2003) Monitoring expression profile of rice genes under cold, drought, and high – salinity stresses and abscicic acid application using cDNA microarray and RNA gel – blot analyses. Plant Physiol. 133:1755-1767.
14. Pokorna J, Schwarzerova K, Zelenkova S, Petrasek J, Janotova I, Capkova V, Opatrny Z (2004) Sites of actin filament initiation and reorganization in cold-treated tobacco cells. Plant, Cell and Environment 27, 641-653
15. Smertenko AP, Jiang CJ, Simmons NJ, Weeds AG, Davies DR, Hussey PJ (1998) Ser 6 in the maize actin-depolymerizing factor, ZmADF3, is phosphorylated by a calcium-stimulated protein kinase and is essential for the control of functional activity. The Plant Journal 14, 187-193
7 (a) (b) ADF1 pI/Mw: 5.56 / 15936.25 ADF2 pI/Mw: 8.40 / 16479.97 ADF3 pI/Mw: 4.90 / 16176.00 ADF4 pI/Mw: 5.11 / 18057.70 ADF5 pI/Mw: 5.72 / 15946.10 ADF6 pI/Mw: 5.65 / 16794.01 ADF7 pI/Mw: 9.46 / 15576.81 ADF8 pI/Mw: 5.94 / 17444.65
Fig.1. (a) Phylogenetic analysis of known ADF genes in rice genomes and (b) their corresponding pI and molecular weight of encoded proteins
7
ADF-2 CK ABA man cold Heat Na Cu CK ABA man cold Heat Na Cu
ADF-3
Actin-1
Fig2. ADF-2, ADF3 及 actin-1 基因在水稻 TN1 及 TNG67 幼苗根部及地上部分別 於不同逆境處理下之 RT-PCR 結果。
其逆境處理分別如下:CK 為對照組無任何處理,ABA 為 10μM ABA 處理 24 小時,man 為 0.5 M mannitol 處理 24 小時,cold 為 5℃低溫處理 24 小時,Heat 為 45℃高溫處理 24 小時,Na 為 200 mM NaCl 處理 24 小時, Cu 為 0.25 M CuCl2處理 24 小時。其中 actin 為 internal control。
7
圖 3. 以南方墨點分析法決定 porin - like protein gene 在水稻中之基因拷貝數。 品種名稱及各種酵素名稱標示如上,其中每個 well loading 10μg 水稻 genomic DNA,並以 1%之洋菜膠體及 TBE buffer 進行 DNA 電泳之分離。雜合所使用之探 針係以 random primer labeling 法標定,放射性核種為α-P32
dCTP。探針使用 porin-like protein 全長之 cDNA 片段,共 1018 bp,其序列及限制酵素切位如 附圖 1。
4 kb
2 kb
1.5 kb TNG67 IR36 PANGONIA
Eco RI Hind III Pst I Eco RI Hind III Pst I Eco RI Hind III Pst I
6 Kb
5 Kb
3 kb
7
圖 4. Porin - like protein gene 於水稻 TNG67 低溫處理下之 RT-PCR 結果。 (A)其基因專一性引子核苷酸序列請參照附表 3。M 為 100bp DNA ladder
marker。處理時間及部位如上標示。(B)為 internal control,採用水稻 之 actin 基因引子,其中每一 well 之樣品 loading 之順序及材料處理和圖 A 相同。EtBr 之分析均採用 1.2%之洋菜膠體及 TAE buffer 進行電泳。 1018bp A 350bp B A B Porin-like protein Actin TNG67 root M CK 1 hr 2 hr 4 hr leaf 24 hr M 5℃
7
圖 5. Porin-like protein 基因在水稻 TNG67 幼苗根部及地上部分別於不同逆 境處理下之 RT-PCR 結果。
(A)所使用之 porin-like protein 基因專一性引子(如附表 3 所示)。M 為 100bp DNA ladder marker。材料為水稻台農 67 上地上部(shoot)和
根部(root),其逆境處理分別如下:CK 為對照組無任何處理,ABA 為 10
μM ABA 處理 24 小時,man 為 0.5 M mannitol 處理 24 小時,cold 為 5 ℃低溫處理 24 小時,Heat 為 45℃高溫處理 24 小時,Na 為 200 mM NaCl 處理 24 小時,Cu 為 0.25 M CuCl2處理 24 小時。(B)為 internal control, 除了使用之 primers 為水稻 actin gene 外,其樣本 loading 順序及材料 處理如圖 A。EtBr 之分析均使用 1.2﹪洋菜膠體及 TAE buffer 進行 DNA 電泳。
B
Porin-like protein 1018bp Actin 350bp TNG67 shoot TNG67 rootM CK ABA man cold Heat Na Cu CK ABA man cold Heat Na Cu M
A
7
圖 6. 以南方墨點法分析 amine oxidase 基因在大粒種水稻 PANGONIA 中之基因 拷貝數。
品種名稱及各種酵素名稱標示如上,其中每個 well loading 10μg 水稻 genomic DNA,使用 1%之洋菜膠體及 TBE buffer 進行 DNA 之電泳分離。探針係以 random primer labeling 法標定,放射性核種為α-P32
dCTP。探針使用 amine oxidase 部分 cDNA 片段,共 250 bp,如附圖 3。
PANGONIA
Eco RI Hind III Pst I
7 Kb
7
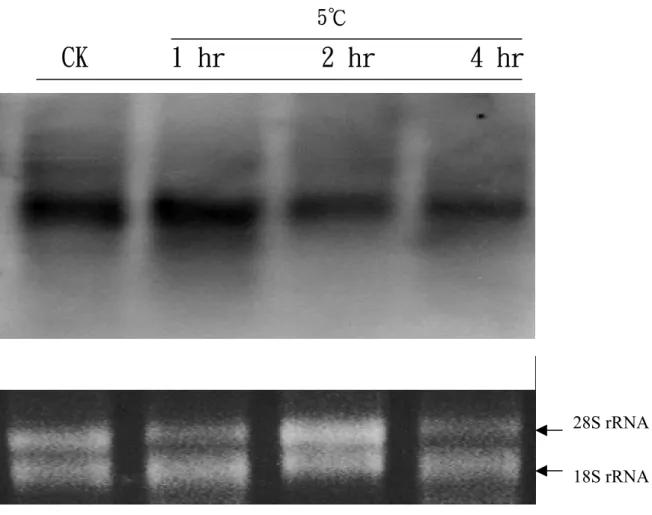
圖 7. Amine oxidase 基因在水稻台農 67 號三葉齡幼苗根部中,經過 5℃低溫 處理後不同時間之表現情形。
Total RNA 係以 1.2%甲醛變性膠體電泳進行分離,其中每個 well 各 loading 20μg total RNA。探針是採用 random primer labeling 方式製 作,標定核種為α-P32 dCTP。其中探針係以 RT-PCR 所得 amine oxidase 之部分 cDNA 片段 250 bp(如附圖 3 所示)。 28S rRNA 18S rRNA
CK 1 hr 2 hr 4 hr
5℃
7
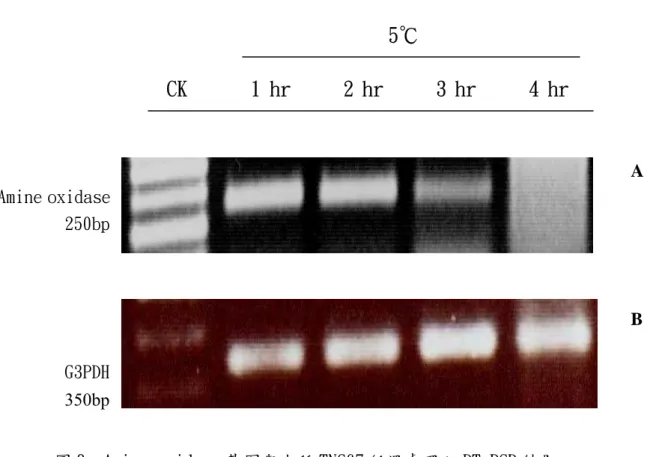
圖 8. Amine oxidase 基因與水稻 TNG67 低溫處理之 RT-PCR 結果。
(A)所使用之 amine oxidase 基因專一性引子核苷酸序列如附表 3。M 為 100bp DNA ladder marker。低溫處理時間如圖上所標示。(B)為 internal control,選用水稻 G3PDH gene primers,其 well 之樣品 loading 順序 與處理和圖 A 相同。EtBr 之分析均使用 1.4%之洋菜膠和 TAE buffer 進行 DNA 電泳。
CK 1 hr 2 hr 3 hr 4 hr
250bp 350bp A B Amine oxidase G3PDH5℃
7
圖 9. Amine oxidase 基因在水稻 TNG67 幼苗根部及地上部分別於不同逆境處理 下之 RT-PCR 結果。
(A)所使用之 Amine oxidase 基因專一性引子如附表 3 所示。M 為 100bp DNA ladder marker。材料為水稻台農 67 上地上部(shoot)和根部(root), 其逆境處理分別如下:CK 為對照組無任何處理,ABA 為 10μM ABA 處理 24 小時,man 為 0.5 M mannitol 處理 24 小時,cold 為 5℃低溫處理 24 小時,Heat 為 45℃高溫處理 24 小時,Na 為 200 mM NaCl 處理 24 小時, Cu 為 0.25 M CuCl2處理 24 小時。(B)為 internal control,除了使用 之 primers 為水稻 actin gene 外,其樣本 loading 順序及材料處理如圖 A。EtBr 之分析均使用 1.2﹪洋菜膠體及 TAE buffer 進行 DNA 電泳。
B
Amine oxidase 250bp Actin 350bp TNG67 shoot TNG67 rootM CK ABA man cold Heat Na Cu CK ABA man cold Heat Na Cu M
7
附圖 1. II-43 Porin-like protein gene 之 cDNA 全長核苷酸序列及專一性引 子相對位置及限制酵素切位(此全長得自 KOME 資料庫,材料為水稻品 種日本晴) g a t t t c t c g t g g c g c c c c a c c a t t c c c t t c t a c t a g t g g c g g a a c c t c t c t g a c c t c t c c t t t c t c t c t c c c t c c t c t t c g g c g g c c g c c t c c c c c c t c c c c c c c t t c c c c a a g c c g c c g ccgccgccATGGCCATGAAGGGACCCGGCCTCTTCTCCGACATTGGCAAGAGGGCCAAGG ATCTGCTCACCAAGGACTACACCTATGACCAGAAGCTGACCGTCTCCACCGTCAGCTCCT CCGGAGTGGGCCTCACTTCCACAGCTGTGAAGAAAGGGGGGCTTTATACTCTTGATGTCA GCTCAGTTTACAAGTACAAGAGTACTCTCGTCGATGTCAAAGTGGACACAGAATCTAATA TCTCTACTACTTTGACTGTGTTTGATGTCCTTCCATCCACAAAGCTTGTGACAAGTGTCA AGCTGCCTGACTACAATTCTGGAAAGGTGGAGATGCAATACTTCCATGAGAATGCAAGTT TTGCCACTGCTGTTGGCATGAAGCCATCTCCTGTGGTTGAATTTTCTGGAACAGCTGGTG CTCAAGGACTTGCCTTTGGTGCAGAAGCTGGGTTTGACACTGCTACAGGAAAATTTACCA AGTACAGTGCTGCGATAGGTGTAACCAAGCCAGATTATCATGCTGCAATCGTTCTGGCGG ACAAAGGTGACACCGTTAAAGTGTCAGGTGTATATCACCTTGATGATAAGCAGAAATCCT CTGTTGTTGCTGAATTAACCCGCAGGCTCTCAACAAATGAGAACACTCTCACTGTTGGTG GCCTGTACAAGGTTGACCCCGAGACAGCTGTGAAGGCAAGGCTCAACAACACTGGAAAGC TTGCTGCTCTTCTCCAGCATGAGGTTAAACCCAAGTCGGTTTTGACAATCTCTGGTGAAT TCGACACCAAGGCCTTGGACAGACCCCCCAAGTTTGGTCTAGCGCTTGCACTCAGGCCCT G At a a c a t c g t c t g t g g t t g a t g g c t g t t a a a t t t c g a a t t c t t t g g t t g t t t t g c a a a g c a a t g g c c a a t a a t t t g t t c c g c a g a c c t g t t a c t t c a a t c t t t t a a a g t t t t g c t g c t c t g t g g g c a g g a g c c a a t g g g t g t t t t a c a t g a t t t t a t c c g c t a c t t c t g t t g a t g a a a g c a t c t a g t c a t t g t t a g a t g t a t g c t g t t t t c c t c t c t t t c t t a g c t t a a a g c t c a c t t t c t a t g t a c t a c t g a g g t g c c a t g c c a c c a g t a g t c t a t a t t g a t a t t c c t t g c a c a a g g t acgtg PL-F PL-R Eco RI Eco RI Hind III Hind III
7
附圖 2. II-22 β-glucosidase gene 之 cDNA 全長核苷酸序列及專一性引子相 對位置(此全長得自 KOME 資料庫,材料為水稻品種日本晴) a t c a g t c c g t g c a g t t t g g a g c c c a c a c a c c a c g a t c t c c c g t a c c t t c t c t c t g c g a gA TGGGGAGGATAAAGAGTAGTAGTGGGCGATGCAGCACGGCGAGGCTGGAGGCGGTCGCCG TCCTCGTCGTCGTCTTCGGCGTCGCGTCGTCGTCGCTGCGAGGATGCATTGCGCAGCAGA GCGGAGGAGGGCTAACCAGGGGCAGCTTCCCCGAGGGGTTCGTCTTCGGCACCGCCTCCG CCGCGTACCAGTACGAGGGAGCTGTGAAGGAGGACGGGAGAGGGCAGACCATCTGGGACA CGTTCGCGCACACCTTTGGAAAGATCACCGACTTCAGCAATGCTGATGTTGCAGTTGATC AGTACCACCGTTTCGAGGAGGATATACAGCTCATGGCAGACATGGGGATGGATGCGTATC GGTTCTCGATAGCATGGTCAAGAATCTACCCAAATGGTGTTGGTCAAGTCAATCAAGCTG GTATCGACCACTACAACAAGCTGATCGATGCACTTCTAGCAAAAGGAATTCAGCCATATG TGACACTCTACCACTGGGACCTCCCCCAGGCCCTTGAAGACAAGTACAAGGGCTGGCTTG ACAGGCAGATAGTGGACGATTTCGCGGCGTACGCGGAGACGTGCTTCAGGGAGTTCGGGG ACAGGGTGAAGCACTGGATCACGCTCAACGAGCCGCACACGGTGGCCATCCAGGGCTACG ACGCAGGGCTCCAGGCCCCCGGCCGCTGCTCCGTGCTGCTCCACCTCTACTGCAAGGCCG GCAACTCCGGCACCGAGCCCTACGTCGTCGCCCACCACTTCATCCTCGCCCACGCCGCCG CCGCCAGCATCTACAGGACAAAATACAAGGCGACGCAGAACGGGCAGCTTGGGATAGCGT TCGACGTGATGTGGTTCGAGCCGATGTCCAACACCACGATCGACATCGAGGCGGCCAAGA GAGCGCAGGAGTTTCAGCTAGGATGGTTTGCTGATCCGTTCTTCTTCGGCGACTACCCGG CGACGATGAGGGCGAGGGTGGGGGAGAGGCTGCCGAGGTTCACGGCGGATGAGGCCGCCG TCGTCAAGGGGGCGCTGGATTTCGTCGGCATAAACCACTACACCACCTACTACACGAGGC ACAACAACACCAACATCATCGGGACATTGCTCAACAACACCTTGGCAGACACCGGCACCG TCAGCCTCCCATTCAAGAATGGGAAGCCAATTGGAGATAGGGCAAATTCGATATGGCTGT ACATTGTGCCCCGAGGGATGAGGAGCCTGATGAACTATGTCAAGGAAAGGTACAACAGCC CACCAGTGTACATCACTGAAAACGGGATGGATGACAGCAACAACCCGTTCATTTCCATCA AGGACGCCCTCAAGGACAGCAAGAGGATCAAATACCACAATGACTACCTCACCAATCTGG SRGF
7 CTGCTTCCATCAAGGAGGACGGGTGCGACGTACGTGGGTACTTCGCGTGGTCTCTGCTGG ACAACTGGGAGTGGGCGGCCGGATACTCCTCGAGATTCGGGCTCTACTTCGTGGACTACA AGGATAACCTCAAGAGATACCCCAAGAACTCGGTGCAGTGGTTCAAGGCCCTCCTGAAGA C CT G Aa c t c c a g c t g a g a a g t a g t g c t a c a a t a g t t t g g g c g c c a t t t c c a t a c a t c c g a t a g a t t t a t t g g t c t g c a c a t a t a c g a a t a t a c g a t a c c c t g c a t a t c t c g g a a a t g t t t t g t g g t t t t t g t t t t g t t t g t g a t a c g a t a g a a t g t a t c a c t g a a t c g t c t g g t c t g t t a SRGR SRG5N SAG3’ SAG5’
7
附圖 3. Amine oxidase 基因之全長核苷酸序列及β-glucosidase 基因專一性 引子相對應位置(此全長得自 KOME 資料庫,材料為水稻品種日本晴) g g a a g a a a c t a a g g a a g a c a t t t c t a t t g c g a a g g c c a t t g c a a t t g t tA T GG A G A G A A A T C C A C A C T T G A G G C A A G A A G G G A T T G C T C A T G A T G T T C T T C A G T G G T A T T T G T G C C G C A T G G A A G G T T G G T T T G C T A C T G A T G C G G A C G C A A T C T C T C T T C A G G G T T G G G A C C A G G A G G T A C T G C T T C C T G G T G G C C A T G G T C T C A T G G T T C G T G G A T A T C G T C C A G T A A T A A A T A C T C T G G C A A A A G G C C T A G A T A T A C G C C T T G G C C A C A G G G T T G T T G A G A T T G T T C G C C A C A G G A A T A G G G T G G A G G T T A C T G T A A G C A G T G G T A A A A C A T T T G T T G C G G A T G C T G C A G T C A T T G C T G T C C C C T T A G G T G T T C T T A A A G C A A A C A C C A T T A A A T T T G A A C C T A G G C T G C C A G A G T G G A A G G A G G A A G C A A T A A G A G A A C T C T C A G T T G G A G T C G A G A A C A A A A T A A T T C T C C A C T T C A G C G A G G T T T T C T G G C C T A A T G T G G A G T T C C T T G G A G T A G T T T C A T C C A C C A C A T A T G G C T G C A G C T A T T T C C T C A A C C T C C A C A A G G C T A C T G G C C A T C C T G T T C T A G T T T A C A T G C C T G C G G G T C G G C T T G C A T G T G A C A T T G A A A A G C T G T C A G A T G A G G C T G C T G C C C A A T T T G C C T T C T C T C A G C T G A A G A A G A T C C T G C C C A A C G C T G C C G A G C C G A T A C A T T A C C T A G T G T C A C A T T G G G G C T C A G A C G A G A A T A C A C T T G G A T C C T A C A C C T T C G A T G G G G T G G G C A A A C C T C G T G A C C T G T A T G A G A A G C T G C G G A T C C C A G T C G A T A A C C T A T T C T T T G C T G G A G A A G C C A C C A G T G T C C A G T A C A C T G G C A C G G T G C A T G G C G C C T T C T C C A C T G G T C T G A T G G C A G C T G A G G A G T G C C G A A T G A G G G T T C T T G A G C G G T T C A G G G A G C T G G A C A T G C T G G A G A T G T G C C A C C C T G C A A T G G G G G A G C A G A C C G C C A C T G T C T C A G T C C C A C T G C T C A T C T C C C G G C T GT G Aa c a g a c c a g g a g c t g c a c a a a a a t t a t a g t c t t c t a a g g c c a t t t g g c a c t t g g c a *粗體加底線字母代表此基因序列和 SAG5’,SAG3’引子不同者

![TraditionalMLCalgorithmsmainlytacklethebatchMLCproblem,wheretheinputdataarepresentedinabatch[24,28].Nevertheless,inmanyMLCapplicationssuchase-mailcategorization[22],multi-labelexamplesarriveasastream.Onlineanalysisistherefore dimensionreducermotivatedbyma](data:image/gif;base64,R0lGODlhAQABAIAAAP///wAAACH5BAEAAAAALAAAAAABAAEAAAICRAEAOw==)