ContentslistsavailableatScienceDirect
Journal
of
Ethnopharmacology
j o u r n al hom ep a g e :w w w . e l s e v i e r . c o m / l o c a t e / j e t h p h a r m
Annonacin
induces
cell
cycle-dependent
growth
arrest
and
apoptosis
in
estrogen
receptor-
␣-related
pathways
in
MCF-7
cells
Yu-Min
Ko
a,
Tung-Ying
Wu
b,
Yang-Chang
Wu
b,
Fang-Rong
Chang
b,
Jinn-Yuh
Guh
c,d,∗,
Lea-Yea
Chuang
e,∗aGraduateInstituteofMedicine,CollegeofMedicine,KaohsiungMedicalUniversity,Kaohsiung,Taiwan,ROC bGraduateInstituteofNaturalProducts,CollegeofPharmacy,KaohsiungMedicalUniversity,Kaohsiung,Taiwan,ROC
cDepartmentofInternalMedicine,FacultyofMedicine,CollegeofMedicine,KaohsiungMedicalUniversity,Kaohsiung,Taiwan,ROC
dDepartmentofInternalMedicine,KaohsiungMedicalUniversityHospital,KaohsiungMedicalUniversity,100Zihyou1stRoad,Kaohsiung,Taiwan,ROC eDepartmentofBiochemistry,CollegeofMedicine,KaohsiungMedicalUniversity,100Zihyou1stRoad,Kaohsiung,Taiwan,ROC
a
r
t
i
c
l
e
i
n
f
o
Articlehistory:
Received16February2011 Receivedinrevisedform11July2011 Accepted28July2011 Available online xxx Keywords: Annonacin Tamoxifen Estrogenreceptor-␣ Breastcancer Cellcycle
a
b
s
t
r
a
c
t
Ethnopharmacologicalrelevance:Tamoxifenresistanceiscommoninestrogenreceptor-␣(ER␣)-positive breastcancers.Pawpawandsoursopareanticancerannonaceousplantsincomplementarymedicine. Thus,westudiedtheeffectsofannonacin,anannonaceousacetogenin,inbreastcancercells.
MaterialsandMethods:CellgrowthandER␣-relatedpathwayswerestudied.Theeffectsofannonacin weretestedinMCF-7xenograftsinnudemice.
Results: In ER␣-positive MCF-7 cells, annonacin (half-effective dose ED50=0.31M) and
4-hydroxytamoxifen(ED50=1.13M)decreasedcellsurvivalwhereasannonacin(0.5-1M)increasedcell
deathat48h.Annonacinand4-hydroxytamoxifenwereadditiveininhibitingcellsurvival.Annonacin (0.1M) induced G0/G1 growtharrest whileincreasingp21WAF1 andp27kip1 anddecreasing cyclin
D1protein expression. Annonacin (0.1M) decreased cyclin D1protein expressionmore than 4-hydroxytamoxifen (1M). Annonacin (0.1M) increased apoptosis while decreasingBcl-2 protein expression.Thecombinationofannonacin(0.1M)and4-hydroxytamoxifen(1M)decreasedBcl-2 proteinexpressionandER␣transcriptionalactivitymorethanannonacin(0.1M)didalone.Annonacin, butnot4-hydroxytamoxifen,decreasedER␣ proteinexpression.Moreover,annonacindecreased phos-phorylationofERK1/2,JNKandSTAT3.Innudemice,annonacindecreasedMCF-7xenografttumorsizeat 7–22days.Moreover,annonacindecreasedER␣,cyclinD1andBcl-2proteinexpressioninthexenograft at22days.
Conclusions:AnnonacininducedgrowtharrestandapoptosisinER␣-relatedpathwaysinMCF-7cells. Annonacinand4-hydroxytamoxifenwereadditiveininhibitingcellsurvivalandER␣transcriptional activity.Moreover,annonacinattenuatedMCF-7xenografttumorgrowthwhileinhibitingER␣,cyclinD1 andBcl-2proteinexpressionsinnudemice.
© 2011 Elsevier Ireland Ltd. All rights reserved.
1. Introduction
Thepathogenesisofbreastcancerincludesestrogenand estro-genreceptor-␣(ER␣)-relatedpathways(OsborneandSchiff,2011) whereby nuclear ER␣ activates target genes via the estrogen-responseelements(ERE)(OsborneandSchiff,2011).Additionally,
Abbreviations: ER␣, estrogen receptor-␣;ED50, 50% effective dose; ERK,
extracellularsignal-regulatedkinase;JNK,c-JunN-terminalkinase;STAT,signal transducersandactivatorsoftranscription;GAPDH,Glyceraldehyde3-phosphate dehydrogenase;cdk,cyclin-dependentkinase;MTT, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-tetrazoliumbromide;PBS,phosphate-bufferedsaline.
∗ Correspondingauthors.Tel.:+88673121101x7353;fax:+88673218309. E-mailaddresses:[email protected](J.-Y.Guh),[email protected]
(L.-Y.Chuang).
nonnuclearER␣rapidlyactivatesgrowthfactordownstream sig-nalssuchasextracellularsignal-regulatedkinases(ERK1/2),c-Jun N-terminalkinase(JNK)andsignaltransducersandactivatorsof transcription3(STAT3)(MusgroveandSutherland,2009;Osborne
andSchiff,2011).Interestingly,STATintegrates cytoplasmicand
nuclearestrogenactions(Foxetal.,2009).
Selectiveestrogenreceptormodulators(SERM,suchas tamox-ifen)orselectiveestrogenreceptordown-regulators(SERD,suchas fulvestrant)areeffectivetreatmentsforER␣-positivebreast can-cerscommonlylimitedbyresistance(OsborneandSchiff,2011). EndocrineresistancemaybeinducedbythelossofER␣,increased nonnuclearER␣orgrowthfactorreceptorsignaling,deranged sig-naltransducers (suchasERK1/ERK1, JNKand STAT3),cellcycle regulators (such as cyclin D1 and the cyclin-dependent kinase inhibitorsp21WAF1 andp27kip1)orapoptosisregulators(suchas
0378-8741/$–seefrontmatter © 2011 Elsevier Ireland Ltd. All rights reserved.
ARTICLE IN PRESS
GModelJEP-6930; No.ofPages8
2 Y.-M.Koetal./JournalofEthnopharmacologyxxx (2011) xxx–xxx
theanti-apoptoticBcl-2)(MusgroveandSutherland,2009;Osborne
andSchiff,2011).
Pawpaw(Asiminatriloba)andsoursop(graviola,Annona muri-cata)areannonaceousplantsusedasanticancerfolktherapiesin (North,CentralandSouth)AmericaandSoutheastAsiaandhave beenstudiedinafewobservationalclinicalstudies(Cassileth,2008;
Coothankandaswamyetal.,2010;Liawetal.,2010;McLaughlin,
2008). Annonaceous acetogenins are cytotoxic to multidrug-resistantMCF-7cells(Oberliesetal.,1997).Annonacin(C35H64O7),
anannonaceous acetogenincontaining a mono-tetrahydrofuran ringwithtwoflankinghydroxyls,alsoinhibitsgrowthinMCF-7 cells(Yuanetal.,2003).However,themolecularmechanismsare notunderstood.
Thus, we studied the growth-inhibitory mechanisms of annonacinin terms of ER␣-related pathways(p-ERK1/2,p-JNK, p-STAT3,cyclinD1,Bcl-2, p21WAF1 and p27kip1)in MCF-7cells.
Moreover,theeffectsofannonacinonMCF-7xenograftsinnude micewerealsoinvestigated.
2. Materialsandmethods 2.1. Cellcultureandreagents
ER␣-positiveMCF-7cellsandER␣-negativeMDA-MB-231cells werepurchased onFebruary 18,2009fromBioresource Collec-tion and Research Center (Hsinchu,Taiwan), where cells were authenticatedbyDNAfingerprintsofshorttandemrepeatprofiling. Cellswereculturedin DMEM/F-12(1:1)mediumsupplemented with1%penicillin/streptomycinand10%fetalbovineserum(FBS, Gibco,GrandIsland,NY,USA)inahumidified5%CO2 incubator
at37◦C. Cellswerefastedfor24hbeforeaddingfreshmedium containing10%FBS andvarious concentrationsof annonacinor 4-hydroxytamoxifen.
Cyclin D1, cyclin-dependent kinase 4 (cdk4), cyclin E, p21WAF1,p27kip1,Bcl-2,Glyceraldehyde3-phosphate
dehydroge-nase(GAPDH),ER␣,STAT3andJNKantibodieswerepurchasedfrom SantaCruzBiotechnologyInc.(SantaCruzCo.,CA).␣-tubulin anti-bodywaspurchasedfromLabVisionCorporation(Fremont,CA). Serine118phosphorylatedER␣(pSer118ER␣),p-STAT3,ERK1/2, p-ERK1/2,JNKandp-JNKantibodieswerepurchasedfromCell Sig-nalingTechnology(Danvers,MA).Wehad isolated,purified and characterizedannonacinfromtheleavesofFormosanA.muricata
(Liawet al.,2002)(supplementaryMethods andsupplementary
Table1),whichhasa differentmolecularstructurefromthatof
tamoxifen(supplementaryFig.1).4-hydroxytamoxifen,anactive metaboliteoftamoxifenandotherchemicalswerepurchasedfrom Sigma–AldrichChemicalCompany(St.Louis,MO).Annonacinwas dissolvedindimethylsulfoxide(DMSO,0.1%finalconcentration). Becauseannonacinhasnotbeen usedinhumans clinically,the effectiveconcentrationwaschosenbasedonourpreviousinvitro study(Yuanetal.,2003).17-estradioland4-hydroxytamoxifen weredissolvedinethanol(0.1%finalconcentration).
2.2. Measurementofcellsurvivalandcelldeath
Cell survival was performedin quadruplicatesimilar toour previousstudy (Chou et al., 2008). Briefly, viable cell numbers weremeasuredbythe 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-tetrazolium bromide (MTT) assay. MTT was purchased from Sigma–AldrichChemicalCompany(St.Louis,MO).Briefly,MCF-7 cells(6500cells/well)orMDA-MB-231cells(3500cells/well)were platedandincubatedin96-wellplates.MTT(1mg/ml)wasadded intoeachwell.Productionofinsolubleformazanbyviablecellswas measuredat540nm.
Dose–response curves for various combinations of 4-hydroxytamoxifen and annonacin and the half-effective dose (ED50) for cell survival at 48h were analyzed by the
four-parameter log-logistic model (Sørensen et al., 2007).The dose rangesusedwere:0%4-hydroxytamoxifen(i.e.annonacin,0.01, 0.05,0.1, 0.5and1M),50%4-hydroxytamoxifen (0.1,0.5,1,5 and10M),91%4-hydroxytamoxifen (0.1,0.5, 1,5and10M) and100%4-hydroxytamoxifen(0.1,0.5,1,5and10M), respec-tively. The concentration addition model and the isobologram method(Sørensen etal.,2007)wasusedtoassessthesynergy, additivityor antagonismof thecombinationsof annonacinand 4-hydroxytamoxifen.
Cell death was assessed by lactate dehydogenase release throughusingtheCytoTox96nonradioactivecytotoxicityassay kitaccordingtothemanufacturer’sinstructions(PromegaCorp., Madison,WI).
2.3. Cellcycleanalysis
Cellcycleanalysiswasperformedsimilartoourpreviousstudy
(Chuangetal.,2006).Briefly,cellsweretrypsinizedandsuspended
in cold phosphate-buffered saline (PBS). Suspended cells were washedtwicewithcoldPBSandfixedwith70%ice-coldethanol andplacedat−20◦C overnight.Cellswerethencentrifugedand
resuspendedwithnuclearstainingbuffer(0.1%TritonX-100inPBS, 200g/mlRNase,and100g/mlpropidiumiodide)for30minat 37◦C.Foreachsample,atleast1×104eventswererecorded.Cell
cycleprofileswereobtainedwithaFACScanflowcytometer(Becton DickinsonCo.,SanJose,CA)anddatawereanalyzedwithWinCycle software(PhoenixFlowSystemsInc.,SanDiego,CA).
2.4. Immunoblotting
Immunoblottingwasperformedsimilartoourpreviousstudy (Guh et al., 2003). Briefly, a 30g sample of cell lysates was electrophoresedon10% sodiumdodecyl sulfate-polyacrylamide gels, transferred to polyvinylidene difluoride membranes. After blocking,blotswereincubatedwithantibodyinblockingsolution overnight(phospho-antibodies)orfor2h(otherantibodies) fol-lowedby5minwashtwiceinPBScontaining0.1%Tween20and thenincubatedwithhorseradishperoxidase-conjugatedsecondary antibodies(SantaCruzBiotechnologiesInc.,SantaCruz,CA)for1h. Enhancedchemiluminescencereagentswereemployedtodepict proteinbandsonthemembrane.
2.5. Measurementofapoptosis
Apoptosis was measured by flow cytometry by using the VybrantTM Apoptosis Assay Kit #2 (Alexa Fluor® 488 annexin
V/Propidium Iodide kit #2) supplied by Molecular Probes Inc. (Eugene,OR)accordingtothemanufacturer’sinstructions.Thiskit detectstheexternalizationofphosphatidylserineinapoptoticcells usingthegreen-fluorescentAlexaFluor®488annexinandthe
red-fluorescentpropidiumiodidenucleicacidstain.Propidiumiodide stainsnecroticcells withredfluorescence.Aftertreatmentwith bothprobes,apoptoticcellsshowgreen fluorescence,deadcells showredandgreenfluorescence,andlivecellsshowlittleorno fluorescence.
2.6. Transienttransfectionandluciferaseassay
Transienttransfectionwasperformedsimilartoourprevious study(Chouetal.,2008).Briefly,MCF-7cellswereplatedinto 6-wellplatesatdensityof1.2×105cells/wellinDMEM/F12medium
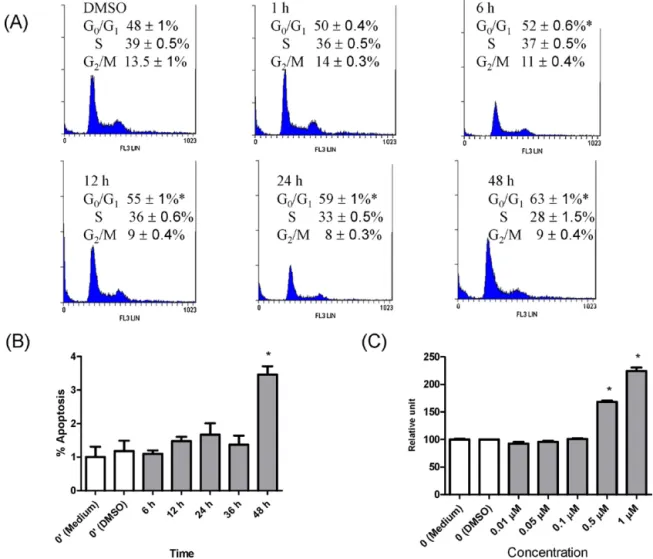
Fig.1.Time-dependenteffectsofannonacinoncellcycledistributionandcelldeathinMCF-7cells.MCF-7cells(2.5×105cells/dish)wereplatedandincubatedin6cm
dishes.Cellswerefastedfor24hbeforeaddingfreshmedium(10%FBS)containing0.1%DMSOor0.1Mannonacin.Cellcyclewasmeasuredbyflowcytometry.Apoptosis wasmeasuredbyflowcytometrybyusingtheVybrantTMApoptosisAssayKit#2(AlexaFluor®488annexinV/PropidiumIodidekit#2).(A)Time-dependent(1–48h)effects
ofannonacin(0.1M)oncellcycledistribution.(B)Time-dependent(6–48h)effectsofannonacin(0.1M)onapoptosis.(C)Dose-dependenteffectsofannonacinoncell death.MCF-7cells(6500cells/well)wereplatedandincubatedin96-wellplates.Cellswerefastedfor24hbeforeaddingfreshmedium(10%FBS,openbars)containing 0.1%DMSO(openbars)orvariousconcentrationsofannonacin(graybars).Celldeathwasmeasuredbylactatedehydrogenaserelease.Theresultswereexpressedasthe mean±SEMofthreeindependentexperiments.*P<0.05versusDMSO.
ER␣transcriptionalactivityreporterplasmidcontaining3X ERE-TATAluc(AddgeneInc.,Cambridge,MA)withLipofectAMINE2000 (LifeTechnology,Gaithersburg,MD)andluciferaseactivitieswere assayedbyintegratingthetotallightemissionover10sbyusing theDynatechML1000luminometer.
2.7. Invivotumorxenograftstudy
Female nudemice (4weeks old; BALB/cA-nu [nu/nu])were purchased fromthe NationalLaboratory AnimalCenter (Taipei, Taiwan)andwerehousedinspecificpathogen-freeconditionsfor 2weeks.Afterwards,MCF-7cells(5×106cellsin200LPBS)were
injectedsubcutaneouslyintotheflanks,andtumorswereallowed todevelopfor30days.Thenelevenmicewererandomlydivided intotwogroups.Annonacin-treatedmice(N=6)were intraperi-toneallyinjecteddailywithannonacin(50mg/kg/day)in200L of25% polyethyleneglycol.Controlmice(N=5)were intraperi-toneally injected daily with200L of 25% polyethyleneglycol. Tumorvolumewasmeasuredusingcalipersat0,3,7,11,15,19 and22days.Tumorvolumewasestimatedbythefollowing for-mula:tumorvolume(mm3)=L×W×W/2(L:length;W:width).
Thexenograftsoftheannonacin-treatedandthecontrolmicewere harvested and fixedin 4% formaldehyde,embeddedin paraffin
forimmunohistochemistryat22days.Allanimalprocedureswere approvedanddoneinaccordancewiththenationalguidelinesand theguidelinesbytheKaohsiungMedicalUniversityAnimal Exper-imentCommittee.
2.8. Immunohistochemistry
Paraffin-embeddedtumortissueswerecutto4msectionsfor immunohistochemistry.Thesectionsweretreatedwithmicrowave at100◦Cfor30min,andblockednonspecificresponse.Thesections wereincubatedat4◦C overnightwithprimaryantibodies(ER␣, cyclinD1andBcl-2).Afterwashingtwice10minwithPBS contain-ing0.2%Tween20,thesectionswereincubatedwithbiotinylated secondaryantibodiesfor1h.AfterwashingtwicewithPBS con-tain0.2%Tween20for10min,sectionswerestainedbyUniversal DAB+kit/HRP(Dako Corp., Carpinteria, CA) and counterstained withhematoxylin.
2.9. Statisticalanalysis
Theresultswereexpressedasthemean±standard errorsof themean.UnpairedStudent’st-tests wereusedforthe compar-isonbetweentwogroups.P<0.05wasconsideredasstatistically
ARTICLE IN PRESS
GModelJEP-6930; No.ofPages8
4 Y.-M.Koetal./JournalofEthnopharmacologyxxx (2011) xxx–xxx
Fig.2.EffectsofannonacinoncellcycleregulatoryproteinsinMCF-7cells. MCF-7cells(2.5×105cells/dish)wereplatedandincubatedin6cmdishes.Cellswere
fastedfor24hbeforeaddingfreshmedium(10%FBS)containing0.1%DMSOor 0.1Mannonacin.Cellcycleregulatoryproteins(cyclinD1,cdk4,cyclinE,p21WAF1
andp27kip1)weremeasuredbyimmunoblotting.(A)Time-dependenteffectsof
annonacin(0.1M)onp21WAF1andp27kip1proteinexpression.(B)Time-dependent
effectsofannonacin(0.1M)oncyclinD1,cdk4andcyclinEproteinexpression.This figureisrepresentativeofthreeindependentexperiments.
significant. Concentration addition model and the isobologram method (Sørensen et al., 2007) were assessed by the drc (dose–response curves) package in the R statistical program
(Knezevicetal.,2009).
3. Results
3.1. Effectsofannonacinoncellcycledistributionandcelldeath inMCF-7cells
Annonacin(0.1M)time-dependently(6–48h)arrestedcells intheG0/G1phaseofthecellcycle(Fig.1A)whileincreasing
apo-ptosisat48h(Fig.1B).Additionally,annonacindose-dependently (0.5–1M)increasedcelldeathat48h(Fig.1C).Annonacinalso dose-dependently (0.05–1M) decreased cell survival at 48h
(supplementary Fig. 2A). Moreover, annonacin (0.1M)
time-dependently(24–48h)decreasedcellsurvival(supplementaryFig. 2B).However,annonacin(0.1M)didnot affectcellsurvival at 6–48hinMDA-MB-231cells(supplementaryFig.2C).
3.2. Time-dependenteffectsofannonacinoncellcycleregulatory proteinsinMCF-7cells
Annonacin (0.1M) time-dependently (30min to 12h) increased p21WAF1 but decreased p21WAF1 protein expression
at 24–48h (Fig. 2A). In contrast, annonacin (0.1M) time-dependently increased p27kip1 protein expression at 1–6h
(Fig. 2A). Additionally, annonacin (0.1M) time-dependently (1–48h) decreased cyclin D1 and time-dependently (12–48h) decreased cdk4 protein expression, but not that of cyclin E (Fig.2B).
3.3. Effectsofannonacinand4-hydroxytamoxifenoncellsurvival andcyclinD1orBcl-2proteinexpressioninMCF-7cells
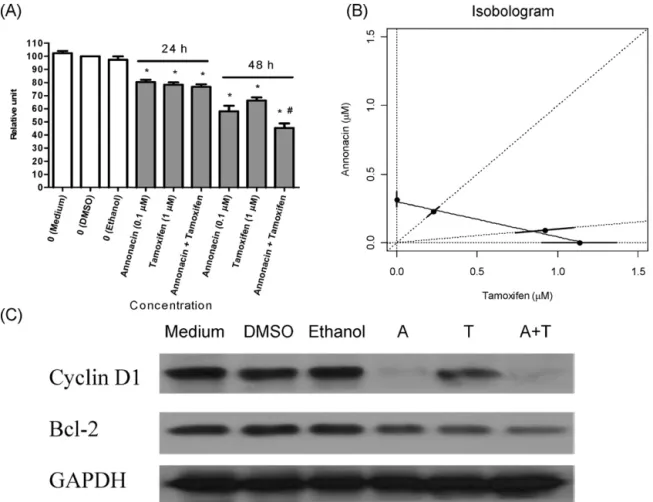
Annonacin(0.1M),4-hydroxytamoxifen(1M)anda com-binationofannonacin(0.1M)plus4-hydroxytamoxifen(1M) decreasedcellsurvivaltoasimilardegreeat24h(Fig.3A).However, thecombinationofannonacin(0.1M)plus4-hydroxytamoxifen
(1M)decreasedcellsurvivalmorethanannonacin(0.1M)did aloneat48h(Fig.3A).
The estimated half-effective doses from dose–response curves at 48h were: 0% 4-hydroxytamoxifen (i.e. annonacin, ED50=0.31M),50% 4-hydroxytamoxifen (ED50=0.45M),91%
4-hydroxytamoxifen(ED50=1.01M),100%4-hydroxytamoxifen
(ED50=1.13M), respectively. Isobologram analysis showed
that the ED50 of the various combinations (50% and 91%
4-hydroxytamoxifen)coincided withthe estimatedconcentration additionisobole(Fig.3B).Inotherwords,thecombinationeffects of annonacinand 4-hydroxytamoxifenwere additiveinsteadof synergistic(ED50 ofthevarious combinationsshiftedtoleft)or
antagonistic(ED50ofthevariouscombinationsshiftedtoright).
Asshown in supplementary Fig.3A,annonacin (0.01–2M) evenly decreased cyclin D1 and Bcl-2 protein expression at 48h.Additionally,annonacin(0.1M)time-dependently(1–48h) decreased Bcl-2 protein expression (supplementary Fig. 3B). Annonacin (0.1M) decreased cyclin D1 protein expression more than 4-hydroxytamoxifen (1M)didat 48h(Fig. 3C). In contrast, annonacin (0.1M) and 4-hydroxytamoxifen (1M) decreased Bcl-2 protein expression to a similar degree at 48h (Fig.3C).Moreover,thecombinationofannonacin(0.1M)plus 4-hydroxytamoxifen (1M)decreased Bcl-2 proteinexpression morethanannonacin(0.1M)didaloneat48h(Fig.3C).
3.4. Effectsofannonacinand4-hydroxytamoxifenonER˛ transcriptionalactivity,ER˛proteinexpressionand phosphorylationinMCF-7cells
As shown in Fig. 4A, annonacin (0.1M) and 4-hydroxytamoxifen(1M)decreasedER␣transcriptionalactivity to the same degree at 24–48h. Moreover, the combination of annonacin(0.1M) plus4-hydroxytamoxifen (1M) decreased ER␣transcriptional activity more than annonacin (0.1M) did aloneat24–48h(Fig.4A).
AsshowninsupplementaryFig.4A,17-estradiolincreasedER␣ transcriptionalactivity whereas annonacin(0.1M) attenuated ER␣transcriptionalactivity at12–48h. Additionally, annonacin (0.1M)decreasedER␣proteinexpressionat24handdecreased ER␣proteinserine118phosphorylationat12h(supplementary Fig. 4B). Annonacin dose-dependently (0.01–2M) decreased ER␣ protein expression at 48h (Fig. 4B). In contrast, 4-hydroxytamoxifen(1M)didnotaffectER␣proteinexpressionat 48h(Fig.4C).Finally,thecombinationofannonacin(0.1M)plus 4-hydroxytamoxifen(1M)decreasedER␣proteinexpressionto asimilardegreeasannonacin(0.1M)didaloneat48h(Fig.4C).
3.5. Time-dependenteffectsofannonacinonphosphorylationof ERK1/2,JNKandSTAT3proteininMCF-7cells
Annonacin (0.1M) time-dependently (1–48h) decreased ERK1/2proteinphosphorylation(Fig.5A).Annonacin(0.1M)also time-dependently(1–48h) decreased JNK protein phosphoryla-tion(Fig.5B).Additionally,annonacin(0.1M)time-dependently (30minto48h)decreasedSTAT3proteinphosphorylation(Fig.5C).
3.6. EffectsofannonacinonMCF-7xenografttumorsizeand expressionofER˛,cyclinD1andBcl-2proteinsinnudemice
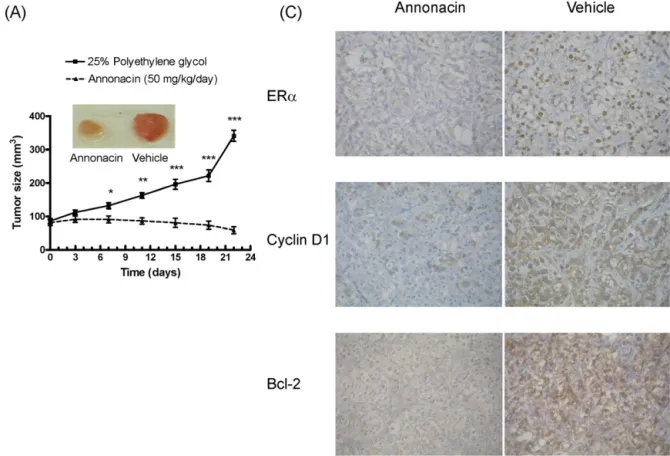
Toconfirmthegrowth-inhibitoryeffectsofannonacininvivo, MCF-7cellsweregraftedintotheflanksofnudemiceasxenografts. AsshowninFig.6A,annonacindecreasedtumorsizeat7–22days. AsshowninFig.6B,annonacinattenuatedtheexpressionofER␣, cyclinD1andBcl-2proteininthenudemiceat22days.
Fig.3.Combinationeffectsofannonacinand4-hydroxytamoxifenoncellsurvivalandcyclinD1andBcl-2proteinexpressioninMCF-7cells.MCF-7cells(6500cells/well) wereplatedandincubatedin96-wellplates.Cellswerefastedfor24hbeforeaddingfreshmedium(10%FBS)containing0.1%DMSOor0.1%ethanol.Cellsurvivalwas measuredbytheMTTassay.(A)Effectsofannonacin(A,0.1M),4-hydroxytamoxifen(T,1M)ortheircombination(A+T)oncellsurvivalat24h(lanes4–6)or48h(lanes 7–9).*P<0.05versuscontrol.#P<0.05versusannonacinalone.(B)Isobologramanalysisofvariouscombinations(50%4-hydroxytamoxifenand91%4-hydroxytamoxifen)
ofannonacinand4-hydroxytamoxifenat48h.Dosecombinations(dottedlines)achievingED50(dots)withstandarderrorstogetherwiththeestimatedisobolesareshown.
Theestimatedconcentrationadditionisoboleisshownasthesolidlineconnectingthe100%and0%4-hydroxytamoxifenED50dots.MCF-7cells(2.5× 105cells/dish)were
platedandincubatedin6cmdishes.ExpressionofcyclinD1andBcl-2proteinwasmeasuredbyimmunoblottingandnormalizedtothatofGAPDH.(C)Effectsofannonacin (A,0.1M),4-hydroxytamoxifen(T,1M)ortheircombination(A+T)oncyclinD1andBcl-2proteinexpressionat48h.Thisfigureisrepresentativeofthreeindependent experiments.
4. Discussionandconclusions
This study adds mechanistic insights to our previous find-ingthat annonacin inhibitsgrowthin MCF-7cells (Yuanet al., 2003).Wefoundthatannonacininducedcelldeathonly athigh doses(≥0.5M).Incontrast,low-dose (0.1M)annonacin,like tamoxifen and fulvestrant (Cariou et al., 2000; Musgrove and
Sutherland,2009), inducedcell-cycle-dependent(G0/G1)growth
arrestconcomitantlywiththeinductionofp21WAF1andp27kipand
theinhibitionofcyclinD1proteinexpression.
Cell cycle (G0/G1 transition) inhibitors p21WAF1 and p27kip
areimplicatedinthepathogenesisorprognosisofbreastcancer
(Caldonetal.,2006).Forexample,thelossofp21WAF1orp27kip
maymediatetamoxifenresistanceinbreastcancer(Musgroveand
Sutherland,2009).Conversely,transfectionofp21WAF1orp27kip
genesinhibitsMCF-7cellgrowth(Jiangetal.,2009).Surprisingly, annonacininhibitedbothp21WAF1andcyclinD1proteinexpression
at24–48h. However,inhibitionof p21WAF1alone cannot
atten-uate cyclinD1 inhibition-inducedgrowtharrest incancer cells
(MasamhaandBenbrook,2009).
OurfindingthatannonacininhibitedcyclinD1protein expres-sioniscompatiblewiththenotionthatcyclinD1overexpressionis associatedwithpoorprognosisandtamoxifenresistanceinER ␣-positivebreastcancer(Buttetal.,2008;MusgroveandSutherland, 2009).Interestingly, cyclinD1 overexpression induces whereas
cyclinD1knockoutinhibitsbreastcancerinmice(Buttetal.,2008). Similarly,cyclinD1overexpressionincreaseswhereas cyclinD1 inhibitiondecreasesproliferationinMCF-7cells(Grilloetal.,2006). Annonacin, liketamoxifen and fulvestrant(Lamet al.,2009;
Musgrove and Sutherland, 2009), also inducedapoptosis while
inhibiting Bcl-2 protein expression. Note that cellsurvival, cell death and cell cycle pathways are interconnected in cancers
(Maddika et al., 2007). For example, apoptosis is a cell-cycle
checkpointduringcellinjuryandBcl-2isananti-apoptotic pro-tein which also inhibits the G1/S checkpoint of the cell cycle
(Maddikaetal.,2007;MusgroveandSutherland,2009).
Transfec-tionoftheBcl-2geneinhibitswhereasinhibitionoftheBcl-2gene enhanceschemosensitivityofbreastcancercells(Emietal.,2005). Moreover,Bcl-2proteinexpressionisassociatedwithtamoxifen resistancein breast cancerpatients(Musgroveand Sutherland,
2009).
The observation that annonacin decreased cell survival in ER␣-positive MCF-7 cells, but not ER␣-negative MDA-MB-231 cells,suggeststhatER␣is requiredfortheeffects ofannonacin. Interestingly,annonacin(ED50=0.31M)wasmorepotentthan
4-hydroxytamoxfien (ED50=1.13M), while annonacin and
4-hydroxytamoxifen were additive, in inhibiting cell survival. Similarly,annonacinwasmorepotentthan4-hydroxytamoxifen, andannonacinand4-hydroxytamoxifenwereadditive,in inhibit-ingER␣transcriptionalactivity.
ARTICLE IN PRESS
GModelJEP-6930; No.ofPages8
6 Y.-M.Koetal./JournalofEthnopharmacologyxxx (2011) xxx–xxx
Fig.4.Effectsofannonacinand4-hydroxytamoxifenonER␣ transcriptionalactivityandER␣ proteinexpressioninMCF-7cells.MCF-7cells(1.2× 105cells/well)wereplated
andincubatedin6-wellplates.Cellswerefastedfor24hbeforeaddingfreshmedium(M,10%FBS)containing0.1%DMSO(D)or0.1%ethanol.(A)Effectsofannonacin (0.1M),4-hydroxytamoxifen(1M)ortheircombinationonER␣transcriptionalactivityat24hor48h.Cellsweretransfectedwith3XERE-TATAlucwithLipofectAMINE 2000andluciferaseactivitieswereassayedbyintegratingthetotallightemissionover10sbyusingtheDynatechML1000luminometer.*P<0.05versuscontrol.#P<0.05
versusannonacinalone.MCF-7cells(2.5×105cells/dish)wereplatedandincubatedin6cmdishes.Cellswerefastedfor24hbeforeaddingfreshmediumcontaining0.1%
DMSO,0.1%ethanol,annonacinor4-hydroxytamoxifen.ExpressionofER␣ proteinwasmeasuredbyimmunoblotting.(B)Dose-dependenteffectsofannonacin(A, 0.01-2M)onER␣proteinexpressionat48h.ExpressionofER␣wasnormalizedtothatof␣-tubulin.(C)Effectsofannonacin(A,0.1M),4-hydroxytamoxifen(T,1M)ortheir combination(A+T)onER␣proteinexpressionat48h.ExpressionofER␣wasnormalizedtothatofGAPDH.BothfiguresBandCarerepresentativeofthreeindependent experiments.
Fig.5. Time-dependenteffectsofannonacinonphosphorylationofERK1/2,JNKandSTAT3proteininMCF-7cells.MCF-7cells(2.5× 105cells/dish)wereplatedandincubated
in6cmdishes.Cellswerefastedfor24hbeforeaddingfreshmedium(M,10%FBS)containing0.1%DMSO(D)orannonacinfor30minto48h.PhosphorylationofERK1/2, JNKandSTAT3proteinwasmeasuredbyimmunoblottingandnormalizedtothatofERK1/2,JNKandSTAT3.(A)Time-dependenteffectsofannnonacin(0.1M)onp-ERK1/2 proteinphosphorylation.(B)Time-dependenteffectsofannnonacin(0.1M)onp-JNKproteinphosphorylation.(C)Time-dependenteffectsofannnonacin(0.1M)on p-STAT3proteinphosphorylation.Thisfigureisrepresentativeofthreeindependentexperiments.
Fig.6.EffectsofannonacinonMCF-7xenografttumorsizeandexpressionofER␣,cyclinD1andBcl-2proteininthenudemice.MCF-7cells(5×106cellsin200LPBS)were
graftedintotheflanksofthenudemiceasxenografts,andtumorswereallowedtodevelopfor30days.Afterwards,annonacin(50mg/kg/day)orvehicle(25%polyethylene glycol)wasadministeredintraperitoneallyfor21days.(A)Tumorvolumewasmeasuredevery3–4daysusingcalipersat0,3,7,11,15,19and22days(inset:tumorat22 days).(B)Tumorwasexcisedat22daysandimmunohistochemistrywasperformedfortheexpressionofER␣,cyclinD1andBcl-2protein.Theresultswereexpressedasthe mean±SEMof5miceineachgroup.*P<0.05,**P<0.01,***P<0.001versusvehicle.
ThemodeofactionofannonacincanbestudiedbyER␣binding assayandER␣transcriptionalactivationassay(Dangetal.,2011). Forexample,SERMandSERDusuallyactbyinhibitingER␣ ligand-binding.WedidnotstudytheER␣-bindingabilityofannonacin. However,wefoundthatannonacin,likeSERD,alsodecreasesER␣ proteinexpression.Thus,annonacin,likeSERD,maybeusefulin tamoxifenresistance(OsborneandSchiff,2011).Wealsofoundthat annonacin decreased ERE-dependent ER␣ transcriptional activ-ity.Note thatEREisindispensableforER␣-inducedphenotypes in breast cancer cells (Nott et al., 2009). Moreover,annonacin decreasedER␣serine118phosphorylation,whichisrequiredfor someER␣-inducedeffects(Duplessisetal.,2011).Incontrast,both tamoxifenandfulvestrantincreaseER␣serine118
phosphoryla-tion(Lipfertetal.,2006;Maggi,2011).However,therearemany
otherpossiblemodesofactionwherebyannonacincaninhibitER ␣-inducedeffects(Shapiroetal.,2011).
Annonacin,unliketamoxifen(Ishiietal.,2008;Lametal.,2009;
Musgrove and Sutherland, 2009), inactivated ERK1/2, JNK and
STAT3inthisstudy.Interestingly,inhibitionofERK1/2attenuates tamoxifenresistance(Ghayadetal.,2010)andinhibitionofJNK orSTAT3inducesapoptosisinbreastcancercells(Kunigaletal.,
2009;Mingo-Sionetal.,2004).Insummary,annonacindiffersfrom
tamoxifenintermsofER␣proteinabundanceand phosphoryla-tion,p-ERK1/2,p-JNKandp-STAT3.Theseobservationsmaypartly accountforthefindingthatannonacinandtamoxifenwereadditive ininhibitingMCF-7cellgrowth.
Theinvitroeffectsofannonacinwerecorroboratedbyour find-ingsthatannonacinattenuatedtumorsizeandtheexpressionof ER␣,cyclinD1andBcl-2proteininMCF-7cell-graftednudemice. Inthisregard,cyclinD1isoverexpressedin50%ofbreastcancers
(Caldonetal.,2006).Breastepithelialcell-specificoverexpression
ofcyclinD1inducesbreastcancerinmice,whilecyclinD1-nullmice areresistanttooncogene-inducedbreastcancer(Buttetal.,2008). Moreover,inhibitionofBcl-2incombinationwithchemotherapy waseffectiveinsomebreastcancerpatientsinaclinicaltrial(Fato
etal.,2008).
Inconclusion,annonacininducedcell-cycle-dependentgrowth arrest andinduced apoptosisinER␣-related pathways(ERK1/2, JNK,STAT3,cyclinD1,Bcl-2,p21WAF1andp27kip1)inMCF-7cells.
Annonacinand 4-hydroxytamoxifen wereadditive ininhibiting growth and ER␣ transcriptional activity. Moreover, annonacin attenuatedMCF-7xenografttumorgrowthwhileinhibitingER␣, cyclinD1andBcl-2proteinexpressioninnudemice.
Acknowledgement
ThisworkwassupportedbytheNationalScienceCouncilof Taiwan(NSC-94-2321-B-037-006)toLea-YeaChuang.
AppendixA. Supplementarydata
Supplementarydataassociatedwiththisarticlecanbefound,in theonlineversion,atdoi:10.1016/j.jep.2011.07.056.
References
Butt,A.,etal.,2008.Cellcyclemachinery:linkswithgenesisandtreatmentofbreast cancer.AdvancesinExperimentalMedicineandBiology630,189–205. Caldon,E.,etal.,2006.Cellcyclecontrolinbreastcancercells.JournalofCellular
Biochemistry97,261–274.
Cariou, S., et al., 2000. Down-regulation of p21WAF1/CIP1 or p27Kip1
ARTICLE IN PRESS
GModelJEP-6930; No.ofPages8
8 Y.-M.Koetal./JournalofEthnopharmacologyxxx (2011) xxx–xxx
ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica 97,9042–9046.
Cassileth,B.,2008.Complementarytherapies,herbs,andotherOTCagents.Oncology (WillistonPark)22,1202.
Chou,W.-W.,etal.,2008.Arecoline-inducedgrowtharrestandp21WAF1expression
aredependentonp53inrathepatocytes.Toxicology243,1–10.
Chuang,L.-Y.,etal.,2006.Roleofnitricoxideinhighglucose-inducedmitogenic responseinrenalfibroblasts.MolecularEndocrinology20,2548–2558. Coothankandaswamy,V.,etal.,2010.Thealternativemedicinepawpawandits
ace-togeninconstituentssuppresstumorangiogenesisviatheHIF-1/VEGFpathway. JournalofNaturalProducts73,956–961.
Dang,Z.,etal.,2011.Comparisonofchemical-inducedtranscriptionalactivationof fishandhumanestrogenreceptors:regulatoryimplications.ToxicologyLetters 201,152–175.
Duplessis,T.T.,etal.,2011.Phosphorylationofestrogenreceptor␣atserine118 directsrecruitmentofpromotercomplexesandgene-specifictranscription. Endocrinology152,2517–2526.
Emi,M.,etal.,2005.TargetedtherapyagainstBcl-2-relatedproteinsinbreastcancer cells.BreastCancerResearch7,R940–R952.
Fato,R.,etal.,2008. Generationof reactiveoxygen speciesby mitochondrial complexI:implicationsinneurodegeneration.NeurochemicalResearch33, 2487–2501.
Fox,E.,etal.,2009.Novelactionsofestrogentopromoteproliferation:integration ofcytoplasmicandnuclearpathways.Steroids74,622–627.
Ghayad,S.,etal.,2010.EndocrineresistanceassociatedwithactivatedErbB sys-teminbreastcancercellsisreversedbyinhibitingMAPKorPI3K/Aktsignaling pathways.InternationalJournalofCancer126,545–562.
Grillo,M.,etal.,2006.ValidationofcyclinD1/CDK4asananticancerdrugtarget inMCF-7breastcancercells:effectofregulatedoverexpressionofcyclinD1 andsiRNA-mediatedinhibitionofendogenouscyclinD1andCDK4expression. BreastCancerResearchandTreatment95,185–194.
Guh,J.-Y.,etal.,2003.Beta-hydroxybutyrate–inducedgrowthinhibitionand col-lagenproductioninHK-2cellsaredependentonTGF-betaandSmad3.Kidney International64,2041–2051.
Ishii,Y.,etal.,2008.TamoxifenstimulatesthegrowthofcyclinD1-overexpressing breastcancercellsbypromotingtheactivationofsignaltransducerandactivator oftranscription3.CancerResearch68,852–860.
Jiang,D.-D.,etal.,2009.[EffectsoftransfectedpIRES-p21waf1-p27kip1geneonthe
duplicationofcentrosomesandtheproliferationofbreastcancercellline MCF-7].ZhonghuaYiXueZaZhi89,3295–3298.
Knezevic,S.Z.,etal.,2009.UtilizingRsoftwarepackagefordose–responsestudies: theconceptanddataanalysis.WeedTechnology21,840–848.
Kunigal,S.,etal.,2009.Stat3-siRNAinducesFas-mediatedapoptosisinvitroand invivoinbreastcancer.InternationalJournalofOncology34,1209–1220. Lam,L.,etal.,2009.TamoxifenandICI182,780increaseBcl-2levelsandinhibit
growthofbreastcarcinomacellsbymodulatingPI3K/AKT,ERKandIGF-1R pathwaysindependentofERalpha.BreastCancerResearchandTreatment118, 605–621.
Liaw,C.-C.,etal.,2002.Newcytotoxicmonotetrahydrofuranannonaceous aceto-geninsfromAnnonamuricata.JournalofNaturalProducts65,470–475. Liaw,C.C.,etal.,2010.HistoricperspectivesonAnnonaceousacetogeninsfromthe
chemicalbenchtopreclinicaltrials.PlantaMedica76,1390–1404.
Lipfert,L.,etal.,2006.Antagonist-induced,activationfunction-2-independent estro-genreceptoralphaphosphorylation.MolecularEndocrinology20,516–533. Maddika,S.,etal.,2007.Cellsurvival,celldeathandcellcyclepathwaysare
inter-connected:implicationsforcancertherapy.DrugResistanceUpdates10,13–29. Maggi,A.,2011.Ligandedandunligandedactivationofestrogenreceptorand hor-monereplacementtherapies.BiochimicaetBiophysicaActa1812,1054–1060. Masamha,C.P.,Benbrook,D.M.,2009.CyclinD1degradationissufficienttoinduce G1cellcyclearrestdespiteconstitutiveexpressionofcyclinE2inovariancancer cells.CancerResearch69,6565–6572.
McLaughlin,J.L.,2008.Pawpawandcancer:annonaceousacetogeninsfrom discov-erytocommercialproducts.JournalofNaturalProducts71,1311–1321. Mingo-Sion,A.,etal.,2004.InhibitionofJNKreducesG2/Mtransitindependent
ofp53,leadingtoendoreduplication,decreasedproliferation,andapoptosisin breastcancercells.Oncogene23,596–604.
Musgrove,E.,Sutherland,R.,2009.Biologicaldeterminantsofendocrineresistance inbreastcancer.NatureReviews.Cancer9,631–643.
Nott,S.L.,etal.,2009.Genomicresponsesfromtheestrogen-responsive element-dependentsignalingpathwaymediatedbyestrogenreceptoralphaarerequired toelicitcellularalterations.JournalofBiologicalChemistry284,15277–15288. Oberlies,N.H.,etal.,1997.Structure−activityrelationshipsofdiverseannonaceous acetogeninsagainstmultidrug resistanthuman mammaryadenocarcinoma (MCF-7/Adr)cells.JournalofMedicinalChemistry40,2102–2106.
Osborne,C.K.,Schiff,R.,2011.Mechanismsofendocrineresistanceinbreastcancer. AnnualReviewofMedicine62,233–247.
Sørensen,H.,etal.,2007.Anisobole-basedstatisticalmodelandtestfor syner-gism/antagonisminbinarymixturetoxicityexperiments.Environmentaland EcologicalStatistics14,383–397.
Shapiro,D.J,etal.,2011.Smallmoleculeinhibitorsasprobesforestrogenand andro-genreceptoraction.JournalofBiologicalChemistry286,4043–4048. Yuan,S.-S.,etal.,2003.Annonacin,amono-tetrahydrofuranacetogenin,arrests
can-cercellsattheG1phaseandcausescytotoxicityinaBax-andcaspase-3-related pathway.LifeSciences72,2853–2861.