Gastric microbiota and predicted gene functions are altered after subtotal
gastrectomy in patients with gastric cancer
Ching-Hung Tseng1-3, Jaw-Town Lin4,5, Hsiu J. Ho6, Zi-Lun Lai6, Chang-Bi Wang7, Sen-Lin Tang1,2*, Chun-Ying Wu6-11*
1Bioinformatics Program, Taiwan International Graduate Program, Academia Sinica, Taipei 11529, Taiwan; 2Biodiversity Research Center, Academia Sinica, Taipei 11529, Taiwan;
3Institute of Biomedical Informatics, National Yang-Ming University, Taipei 11221, Taiwan; 4School of Medicine, Fu Jen Catholic University, New Taipei City 24205, Taiwan;
5Institute of Population Health Sciences, National Health Research Institutes, Miaoli 35053, Taiwan; 6Division of Gastroenterology, Taichung Veterans General Hospital, Taichung 40705, Taiwan; 7Department of Public Health, China Medical University, Taichung 40402, Taiwan
8Graduate Institute of Clinical Medical Sciences, China Medical University, Taichung 40402, Taiwan; 9Faculty of Medicine and Graduate Institute of Clinical Medicine, National Yang-Ming University, Taipei
11221, Taiwan;
10National Institute of Cancer Research, National Health Research Institutes, Miaoli 35053, Taiwan;
11Department of Life Sciences and Rong Hsing Research Center for Translational Medicine, National
Chung-Hsing University, Taichung 40227, Taiwan
*Correspondence:
Chun-Ying Wu, MD, PhD, MPH, Professor
Division of Gastroenterology, Taichung Veterans General Hospital No. 1650, Section 4, Taiwan Boulevard, Taichung 40705, Taiwan
Tel: +886-4-23592525 #3304; Fax: +886-4-23741331; E-mail: [email protected] Sen-Lin Tang, PhD, Associate Research Fellow
Biodiversity Research Center, Academia Sinica
No. 128, Section 2, Academia Road, Taipei 11529, Taiwan
Tel: +886-2-27892814; Fax: +886-2-27890844; E-mail: [email protected] CYW and SLT contributed equally to this work as corresponding authors.
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36
Abstract
Subtotal gastrectomy (i.e., partial removal of the stomach), a surgical treatment for early-stage distal gastric cancer, is usually accompanied by highly selective vagotomy and Billroth II reconstruction, leading to dramatic changes in the gastric environment. Based on
accumulating evidence of a strong link between human gut microbiota and host health, a 2-year follow-up study was conducted to characterize the effects of subtotal gastrectomy. Gastric microbiota and predicted gene functions (16S rRNA gene sequencing) were analyzed before and after surgery. The results demonstrated that gastric microbiota is significantly more diverse after surgery. Ralstonia and Helicobacter were the top two genera of
discriminant abundance in the cancerous stomach before surgery, while Streptococcus and
Prevotella were the two most abundant genera after tumor excision. Furthermore,
N-nitrosation genes were prevalent before surgery, whereas bile salt hydrolase, NO and N2O reductase were prevalent afterward. To our knowledge, this is the first report to document changes in gastric microbiota before and after surgical treatment of stomach cancer. 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58
Introduction
Advances in sequencing technologies and analytical methods have enabled characterization of the human gut microbiota. As part of the human gut, the stomach is inhabited by a wide variety of bacteria, despite the long-held notion of it being a hostile environment for microbial colonization1,2. In healthy individuals, several genera other than
Helicobacter, including Streptococcus, Prevotella, Veillonella, Rothia, and Neisseria, are
abundant in the stomach, based on cloning3,4 and pyrosequencing5,6. Regarding effects of diseases, the gastric microbiota shifts towards decreasing diversity with progression of gastritis, intestinal metaplasia, and gastric cancer7. However, the gastric microbiota of elderly
stomach cancer patients does not significantly differ from that of dyspeptic controls8.
Although chronic infection with Helicobacter pylori causes serious gastric complications, there are no significant associations between microbial phylotypes and H. pylori status of the stomach3. In that regard, H. pylori status only explains 28% of the variance in gastric
microbiota, whereas 44% is explained by host factors9. Understanding of human gastric
microbiota is in its infancy and is complicated by changes over time in persons with
complicated gastric syndromes (which require long-term follow-up and invasive sampling). Subtotal gastrectomy (i.e., partial removal of the stomach) is a surgical treatment for distal gastric cancer, a multifactorial disease causing numerous cancer-related deaths around the world10. Patients receiving subtotal gastrectomy for gastric cancer are often subjected to
other surgical procedures, which alter the gastric environment. For example, highly selective vagotomy affects gastric secretion of gastric acid11; cholecystectomy elevates gastric pH
value12; Billroth II reconstruction reduces pancreatic polypeptide secretion13. Following
subtotal gastrectomy, there are several common side effects, including marginal ulcers, bile reflux, and stump cancer. Bile reflux after subtotal gastrectomy has been associated with the presence of Streptococcus and Veillonella in gastric aspirates14 and Escherichia, Klebsiella,
59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 74 75 76 77 78 79 80 81 82 83
and Clostridium in the intestine15. Although gastric microbiota is altered after subtotal
gastrectomy14, changes in diversity have not been well characterized. Despite characterization
of gastric microbiota by culture-independent approaches, changes following subtotal gastrectomy in patients with gastric cancer are not completely understood. Therefore, there are many knowledge gaps, leading to a number of questions. For example, are different anatomic sites inhabited by different microbes? What is the compositional variation in gastric microbiota after subtotal gastrectomy? What is the biodiversity pattern before and after subtotal gastrectomy? Do metabolic functions embedded in gastric microbiota correspond to changes caused by subtotal gastrectomy? In this study, we aimed to address these questions by deep sequencing of microbial 16S ribosomal RNA (rRNA) genes in gastric tissues.
Gastric microbiota in gastric cancer patients (at various anatomic sites and before and after subtotal gastrectomy) was characterized by 16S rRNA gene sequencing. Within the 2-year timeframe of this follow-up study, 24 gastric biopsies were collected from 6 patients subjected to subtotal gastrectomy. Variations in gastric microbiota and metabolic capabilities before and after tumor excision (subtotal gastrectomy) were determined.
Results
Statistical summaries of sequencing results
To characterize stomach bacterial microbiota and potential variations associated with subtotal gastrectomy, we collected tumor (abbreviated as T in figures) and non-tumor (N) tissues before surgery, as well as gastric stump (S) and high body (B) tissues after surgery, from 6 gastric cancer patients. A total of 4.6 million pair-end reads were generated, of which 3.2 million reads passed quality filtering and were non-chimeric. To determine bacterial community diversity and composition, reads were aligned to the Greengenes database and non-bacterial sequences removed. On average, 85% of reads in a sample were retained. In total, 2.7 million reads (on average, 113±42 thousand reads per sample) were used for 84 85 86 87 88 89 90 91 92 93 94 95 96 97 98 99 100 101 102 103 104 105 106 107 108
subsequent analyses.
Bacterial diversity
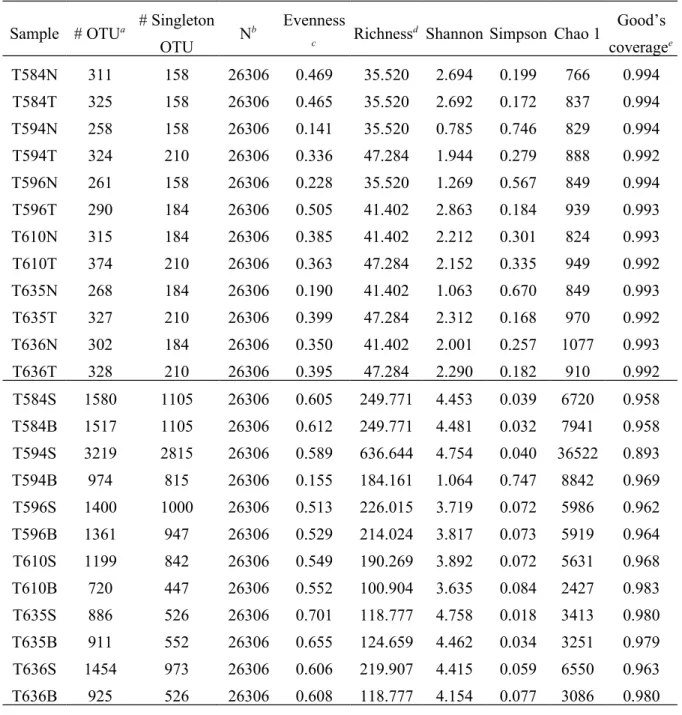
Based on 16S rRNA gene sequencing data, bacterial richness (number of operational taxonomic units (OTUs), richness value, and Chao 1 index) increased after surgery (Table 1). Similarly, there was greater diversity of bacterial communities after surgery (Shannon index) than before surgery (P<0.01, one-tailed Student’s t-test; Table 1). However, within patients, there were no differences (P>0.05, two-tailed paired t-test of Shannon indices) between tumor and non-tumor tissues (before surgery) or between gastric stump and high body tissues (after surgery). Before-surgery communities were characterized by rarefaction curves approaching an asymptote, whereas after-surgery communities were characterized by curves with steeper slopes (Figure S1), indicating the potential for greater diversity to be discovered with more sequencing efforts. Good’s coverage estimates sampling completeness by calculating the probability that a randomly selected read from a sample has been sequenced. At 97% sequence similarity level, Good’s coverage values for all sampled bacterial communities ranged from 0.964 to 0.996 (except T594S, with 0.893 Good’s coverage) when estimated using all reads (Table S1).
Bacterial community structure
Bacterial abundance (i.e., read count) was normalized by corresponding copy number of 16S rRNA genes (prior to community compositional analysis) to reduce potential bias of abundance estimation due to copy number variation16. Before surgery, stomach microbiota in
tumor and non-tumor tissues was dominated by the phyla Proteobacteria (80% in N and 67% in T) and Actinobacteria (15% in N and 24% in T), followed by Firmicutes and Bacteroidetes (<4% in both N and T; Figure 1A). However, after surgery, Firmicutes and Bacteroidetes dramatically increased in both gastric stump and high body tissues (32 and 20% respectively, on average), while Proteobacteria and Actinobacteria decreased (Figure 1A).
At the class level, there was obvious personal variation in stomach microbiota. For 109 110 111 112 113 114 115 116 117 118 119 120 121 122 123 124 125 126 127 128 129 130 131 132 133 134
example, in 3 before-surgery samples Epsilonproteobacteria was dominant (>95%), whereas 5 samples harbored Actinobacteria (at least 25%; Figure 1B). After surgery, Bacilli (of
Firmicutes) and Bacteroidia (of Bacteroidetes) significantly increased in the stomach of most patients, representing (on average) 25 and 19% of the microbiota, respectively.
Principal component analysis (PCA) with surgery status as instrumental variable revealed significant differences in bacterial genera abundance before and after surgery (P=0.0001, Monte-Carlo simulation; Figure 1C). Furthermore, from the results of LEfSe analysis, there were a total of 63 known genera with differential abundance before and after surgery. Among 19 genera known to be more abundant before surgery, Ralstonia and
Helicobacter were the two with the largest linear discriminant analysis (LDA) effect size
(Figure 1D), whereas Streptococcus and Prevotella represented the top two genera (of 43) after surgery.
Group differences (i.e., before and after surgery) in bacterial community structure were also observed using the Bray-Curtis measure of beta diversity17. The within-group distance
was significantly lower than the between-group distance when samples were divided into before- and after-surgery groups (P<0.001; Figure 2A). However, comparisons between tumor and non-tumor tissues before surgery (Figure 2B) and between gastric stump and high body tissues after surgery (Figure 2C) showed insignificant or only marginally significant differences in distances. Interestingly, community dissimilarity within patients was lower than that between patients (regardless of surgery status), although only after-surgery communities demonstrated a significant difference (Figure 2D).
Bacterial gene functions
Based on the community structure derived from 16S rRNA gene sequencing data, predicted gene functions of each sample were inferred from referenced bacterial genomes18.
Enrichment analysis was performed (two-group comparison19) to identify metabolic functions
135 136 137 138 139 140 141 142 143 144 145 146 147 148 149 150 151 152 153 154 155 156 157 158 159
of differential abundances that were significantly enriched (Benjamini-Hochberg adjusted
P<0.05) in bacterial microbiota before and after surgery. In agreement with the dramatically
changed community composition, there were 1879 and 2394 Clusters of Orthologous Groups (COGs) enriched in samples before (Table S2) and after surgery (Table S3), respectively. Given that there were so many genes of differential abundance, and based on the
understanding of metabolic diversity of gut microbiota20,21, functional genes related to gastric disease and carcinogenesis were prioritized for further investigation.
Metabolic enzymes involved in denitrification (e.g., nitrate and nitrite reductases) were more abundant in the gastric microbiota before surgery (Figure 3A). Furthermore, genes related to nitrosation (conversion of organic compounds to nitroso derivatives) were also more abundant before surgery, including the gene families of phenol degradation (Figure 3A; COG4313) and cytochrome (COG3474 and COG3258). From pathway-centric comparisons, biosynthesis of various vitamins, including biotin (vitamin H), riboflavin (vitamin B12), pyridoxal phosphate (vitamin B6), cobalamin (vitamin B12), thiamine (vitamin B1), and menaquinone (vitamin K), was more prevalent in the gastric microbiota before surgery than after surgery (Table S4). Conversely, after-surgery gastric microbiota encoded more bile salt hydrolase (Figure 3B; COG3049 and K01442) and genes related to reduction of nitric oxide (NO) and nitrous oxide (N2O).
Discussion
This is the first follow-up study in which deep sequencing is applied to investigate the stomach microbiota in gastric cancer patients before and after subtotal gastrectomy. This study is also the first to predict gene functions associated with variations in gastric
microbiota. There was an obvious shift in community composition of the gastric microbiota after surgery. Decreases in Proteobacteria and Actinobacteria and increases in Firmicutes and Bacteroidetes defined the after-surgery microbiota shift (at phylum level). Ralstonia and 160 161 162 163 164 165 166 167 168 169 170 171 172 173 174 175 176 177 178 179 180 181 182 183 184
Helicobacter were the top two genera of discriminant abundance in the stomach of patients
with gastric cancer, while Streptococcus and Prevotella were the top two genera after surgery. In contrast, there was relatively low divergence of gastric microbiota among various sites in the stomach. Corresponding to the community shift, the gastric microbiota also exhibited differential predicted gene functions. For example, denitrification and nitrosation genes were prevalent in patient stomachs before surgery, whereas bile salt hydrolase and NO and N2O reductases were prevalent after surgery.
After gastrectomy, the stomach was dominated by only four phyla (Proteobacteria, Firmicutes, Bacteroidetes, and Actinobacteria), which are in abundance in the gastric microbiota of healthy individuals3,5. The relative abundance of these phyla covaries with H.
pylori status9. Composition of the gastric microbiota gradually changes along with
progression of gastric diseases (from gastritis to intestinal metaplasia and ultimately gastric cancer7,22). This study extended observations corresponding to surgical removal of gastric cancer tissue, resulting in increases in Firmicutes and Bacteroidetes and decreases in Actinobacteria and Proteobacteria.
In the present study, gastric cancer tissue and neighboring normal tissue demonstrated similar microbiota. After subtotal gastrectomy, the within-patient microbiota revealed even greater similarity between gastric stump and high body tissues, similar to the findings of a previous report in which there was little difference in gastric microbiota between antrum and body biopsy specimens4. Based on cultures of biopsy specimens, 62% of gastric microbiota
exist in both the antrum and body of the stomach23, which is as expected as these two sites are
distinct niches for microbial colonization, given their differential ability to secrete gastric acid24. Based on the present results and those of previous studies, we inferred that the gastric
environment as a whole is more critical than the individual anatomical sites in determining microbial composition. 185 186 187 188 189 190 191 192 193 194 195 196 197 198 199 200 201 202 203 204 205 206 207 208 209
Gastric acidity is a barrier to microbial overgrowth25,26. However, the secretion of gastric acid is reduced after vagotomy11 (i.e., cutting branches of the vagus nerve), resulting in a
mildly acidic stomach that allows for more bacterial colonization25 (mostly ingested).
Similarly, acid-reducing drugs reportedly increase bacterial colonization in stomach27. This
phenomenon correlated with our results that the richness and diversity (Shannon index) of gastric microbiota increased in after-surgery samples. Another potential side effect of
vagotomy is cobalamin deficiency, due to decreased gastric intrinsic factor28. Consistent with
this prediction, our pathway-centric comparison identified decreased abundance of genes for biosynthesis of cobalamin (and other vitamins) after surgery. Therefore, growth of bacteria lacking cobalamin synthesis genes might be the microbial response following vagotomy that contributes to cobalamin deficiency, although it has been proposed that gastrointestinal bacteria in humans are competitors for cobalamin rather than contributors29.
In addition to vitamin biosynthesis, other predicted metabolic functions of differential abundances were analyzed before and after surgery. For example, nitrate and nitrite reductase, phenol degradation gene, and cytochrome (all of which are related to bacteria-mediated N-nitrosation) were differentially abundant before surgery. Phenols (from protein fermentation) have been reportedly to interact with nitrite (converted from nitrate in food and saliva by nitrate reductase, which is enriched before surgery) under acidic conditions, such as in the stomach, to generate mutagenic p-nitrosophenol30, of which a large amount can catalyze N-nitrosation31. Therefore, when dimethylamine is ingested along with a high-protein diet, the nitrosophenol (from the interaction of phenols and nitrite) may catalyze and even enhance N-nitrosation of dimethylamine to form carcinogenic nitrosamine. By nitrite reductase, nitrite is converted to NO. The produced NO may bind to the cytochrome as a ferrous nitrosyl, a potential nitrosating agent21. Besides that, NO may also interact with dimethylamine under anaerobic conditions to form nitrosamine in the presence of metal ions, such as copper32. It is 210 211 212 213 214 215 216 217 218 219 220 221 222 223 224 225 226 227 228 229 230 231 232 233 234
well known that N-nitroso compounds are causative factors in carcinogenesis. Therefore, enriched genes functionally associated with N-nitrosation before surgery were in agreement with the findings of the cancerous stomach in this study. Since cytochrome commonly presents in respiratory chains and nitrate reductase is used by many Proteobacteria for anaerobic respiration, the differential abundance of these genes was likely a reflection of the high level of Proteobacteria before surgery. As for nitrite reductase, COG1251 (nirB) and COG2146 (nirD) are functionally associated with nitrate assimilation in various bacteria33,
suggesting that these two genes are not involved in gastric NO production.
After surgery, the gastric microbiota demonstrated increases in bile salt hydrolase (COG3049 and K01442), NO reductase (COG3256 and COG3901), and N2O reductase (COG4263). After Billroth II reconstruction and cholecystectomy (i.e., gallbladder removal), the continuous flow of bile salts through the stomach (bile reflux) physically changed the gastric environment, making the stomach a niche for microbes capable of degrading bile salts. This plausibly explains the enrichment of bile salt hydrolase in gastric microbiota after subtotal gastrectomy. Furthermore, NO can be produced enzymatically by activated
leukocytes and bacteria34 and its bactericidal effect has been suggested to protect the stomach
from pathogenic colonization35. However, bacterial NO reductase participates in the defense
against NO toxicity36, implying that the gastric microbiota has a higher capability of NO
detoxification after surgery than before surgery. Abundant N2O reductase after surgery corresponds to the reported increase in N2O concentration after partial gastrectomy37, although the microbial effect of N2O in stomach is not fully understood.
In conclusion, subtotal gastrectomy alters gastric microbiota in terms of diversity, community composition, and predicted gene functions. These changes in the microbial community of the stomach are closely associated with the altered gastric environment after subtotal gastrectomy. 235 236 237 238 239 240 241 242 243 244 245 246 247 248 249 250 251 252 253 254 255 256 257 258 259
Methods
Study subjects and gastric tissue specimen collection
Gastric tissue specimens were collected from early-stage gastric cancer patients before and after curative subtotal gastrectomy at Taichung Veterans General Hospital. Patients with previous malignancies or who had received chemotherapy, radiation therapy or prior gastric surgery were excluded. In addition, patients who had received proton pump inhibitors, H2 receptor antagonists, antibiotics, or probiotics within 1 month of tissue collection were excluded.
Gastric cancerous tissues and neighboring non-tumor tissues were collected before surgery. All patients underwent subtotal gastrectomy, which included excision of 40–50% stomach, vagotomy, cholecystectomy, and Billroth II reconstruction. Bile flow was through the stomach after surgery due to the method of Billroth II reconstruction. Approximately 2 years after subtotal gastrectomy, gastric tissue specimens were collected from the gastric stump (1 cm away from the anastomosis site) and high body of the lesser curvature of the stomach. Comprehensive oral explanations were given and signed informed consent was obtained from all study subjects. The experiments were carried out in accordance with the protocols approved by the Institute Review Board of Taichung Veterans General Hospital.
Bacterial genomic DNA extraction
Bacterial genomic DNA was extracted with the Qiagen DNA Mini Kit (Qiagene, MD, USA). Briefly, tissue samples (~20 mg) yielded 15–20 μg genomic DNA for direct use in polymerase chain reaction (PCR) assays and 16S rRNA gene sequencing. Each tissue sample was homogenized by adding lysozyme (100 mg/mL, Sigma-Aldrich, St. Louis, MO, USA) to lysis buffer to promote lysis of Gram-positive bacteria, thereby enhancing total DNA yields. Amount and quality of isolated genomic DNA were determined with NanoDrop ND-1000 (Thermo Scientific, Wilmington, DE, USA). Genomic DNA was stored at –80°C prior to 16S rRNA sequencing. 260 261 262 263 264 265 266 267 268 269 270 271 272 273 274 275 276 277 278 279 280 281 282 283 284 285
16S rRNA sequencing and analysis
The hypervariable region V1–V3 of bacterial 16S rRNA genes was amplified by PCR using bar-coded universal primers 27F (F, forward primer;
AGAGTTTGATCMTGGCTCAG-3′) and 534R (R, reverse primer;
5′-GTATTACCGCGGCKGCTG-3′). Library construction and sequencing of amplicon DNA samples were conducted with Genomics BioScience (Taipei, Taiwan). A pair-end library (insert size of 490 bp for each sample) was constructed in accordance with the manufacturer’s instructions of MiSeq Reagent Kit v3 (Illumina, Wilmington, DE, USA) and high-throughput sequencing was performed on the Illumina MiSeq 2000 platform (Illumina).
On a per-sample basis, pair-end reads were merged using USEARCH (v7.0.1090)38, with
minimum overlap of read pair set at eight base pairs (bp). Merged reads were quality-filtered with Mothur (v1.34.3)39 to remove reads shorter than 450 bp or longer than 550 bp, as well as
reads with minimum average quality score <27. In addition, reads containing an ambiguous base or homopolymer exceeding 8 bp were excluded. Chimera detection was performed using USEARCH (reference mode and 3% minimum divergence).
Quality-filtered and non-chimeric reads were analyzed (UPARSE40 pipeline) to generate
OTUs per sample (at 97% identity level). The OTU representative sequences were searched against the Greengenes 13_5 database using USEARCH global alignment to identify the corresponding taxonomy of the best hit. Any OTU without a hit or with only a weak hit, i.e. the function “(% sequence identity + % alignment coverage)/2” was <93, was excluded from further analysis. The abundance of each taxon was counted and corrected with PICRUSt18, in
which the pipeline divided the read count of each taxon by the corresponding 16S rRNA gene copy number. Diversity indices (e.g., Shannon, Simpson, Chao 1, and Good’s coverage) were estimated with Mothur.
Statistical analysis of bacterial community
286 287 288 289 290 291 292 293 294 295 296 297 298 299 300 301 302 303 304 305 306 307 308 309 310
unless otherwise specified. Gene copy number-corrected abundance of genera was total-sum scaled per sample. A pseudocount of 0.0001 was added to the relative abundance (in
percentage) before logarithmic transformation41. PCA was performed on log-transformed data
using the R package ade442 to analyze genera abundance before and after surgery.
Between-group inertia percentages were tested (Monte-Carlo test with 10000 permutations) to determine the P-values of PCA results. To identify organismal features differentiating communities of stomach bacteria before and after surgery, LEfSe43 was applied with α=0.05
(Kruskal-Wallace and Wilcoxon tests) and effect size threshold of 2 on linear discriminant analysis (LDA) through the web site, http://huttenhower.sph.harvard.edu/galaxy. Community structure similarities within and between groups were assessed using the Bray-Curtis
distance17 via the R package vegan44, based on relative abundance of bacterial genera. Prediction and analysis of gene functions of bacterial microbiota
Metabolic profiles of bacterial communities were predicted with PICRUSt, which forecasts abundance of genes of metabolic function based on the 16S copy number-corrected OTU composition. Functional genes were categorized (by PICRUSt) into COGs and KEGG Orthology (KO) gene families. To identify gene functions that differentiated bacterial communities before and after surgery, COG gene abundance was subjected to enrichment analysis of two-group comparison, using the R package ShotgunFunctionalizeR19. This
analysis normalized gene abundances using a generalized linear model with Poisson canonical logarithmic link function and determined differential significance (P-value) via a binomial method with a Benjamini-Hochberg false discovery rate correction to adjust q-values for multiple testing.
References
1 Wu, W. M., Yang, Y. S. & Peng, L. H. Microbiota in the stomach: new insights. J Dig
Dis 15, 54-61 (2014).
2 Yang, I., Nell, S. & Suerbaum, S. Survival in hostile territory: the microbiota of the 312 313 314 315 316 317 318 319 320 321 322 323 324 325 326 327 328 329 330 331 332 333 334 335 336 337
stomach. FEMS Microbiol Rev 37, 736-761 (2013).
3 Bik, E. M. et al. Molecular analysis of the bacterial microbiota in the human stomach.
Proc Natl Acad Sci U S A 103, 732-737 (2006).
4 Li, X. X. et al. Bacterial microbiota profiling in gastritis without Helicobacter pylori infection or non-steroidal anti-inflammatory drug use. PLoS One 4, e7985 (2009). 5 Andersson, A. F. et al. Comparative analysis of human gut microbiota by barcoded
pyrosequencing. PLoS One 3, e2836 (2008).
6 Delgado, S., Cabrera-Rubio, R., Mira, A., Suarez, A. & Mayo, B. Microbiological survey of the human gastric ecosystem using culturing and pyrosequencing methods.
Microb Ecol 65, 763-772 (2013).
7 Aviles-Jimenez, F., Vazquez-Jimenez, F., Medrano-Guzman, R., Mantilla, A. & Torres, J. Stomach microbiota composition varies between patients with non-atrophic gastritis and patients with intestinal type of gastric cancer. Sci Rep 4, 4202 (2014). 8 Dicksved, J. et al. Molecular characterization of the stomach microbiota in patients
with gastric cancer and in controls. J Med Microbiol 58, 509-516 (2009).
9 Maldonado-Contreras, A. et al. Structure of the human gastric bacterial community in relation to Helicobacter pylori status. ISME J 5, 574-579 (2011).
10 Polk, D. B. & Peek, R. M., Jr. Helicobacter pylori: gastric cancer and beyond. Nat Rev
Cancer 10, 403-414 (2010).
11 Jepson, K. & Johnston, D. Effect of vagotomy on human gastric acid secretion stimulated by gastrin pentapeptide and by histalog. Gastroenterology 55, 665-669 (1968).
12 Brown, T. H., Walton, G., Cheadle, W. G. & Larson, G. M. The alkaline shift in gastric pH after cholecystectomy. Am J Surg 157, 58-65 (1989).
13 Rieu, P. N., Jansen, J. B., Hopman, W. P., Joosten, H. J. & Lamers, C. B. Effect of partial gastrectomy with Billroth II or Roux-en-Y anastomosis on postprandial and cholecystokinin-stimulated gallbladder contraction and secretion of cholecystokinin and pancreatic polypeptide. Dig Dis Sci 35, 1066-1072 (1990).
14 Poxon, V. A., Morris, D. L., Youngs, D. J., Albutt, E. C. & Keighley, M. R. Exposure to bile acids and bacteria over 24 hours following partial gastrectomy, vagotomy, and pyloroplasty. World J Surg 10, 981-989 (1986).
15 Domellof, L., Reddy, B. S. & Weisburger, J. H. Microflora and deconjugation of bile acids in alkaline reflux after partial gastrectomy. Am J Surg 140, 291-295 (1980). 16 Kembel, S. W., Wu, M., Eisen, J. A. & Green, J. L. Incorporating 16S gene copy
number information improves estimates of microbial diversity and abundance. PLoS
Comput Biol 8, e1002743 (2012).
17 Bray, J. R. & Curtis, J. T. An ordination of the upland forest communities of southern 338 339 340 341 342 343 344 345 346 347 348 349 350 351 352 353 354 355 356 357 358 359 360 361 362 363 364 365 366 367 368 369 370 371 372 373 374
18 Langille, M. G. et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat Biotechnol 31, 814-821 (2013).
19 Kristiansson, E., Hugenholtz, P. & Dalevi, D. ShotgunFunctionalizeR: an R-package for functional comparison of metagenomes. Bioinformatics 25, 2737-2738 (2009). 20 Blaut, M. & Clavel, T. Metabolic diversity of the intestinal microbiota: implications
for health and disease. J Nutr 137, 751S-755S (2007).
21 Hillman, B. Role of gut bacteria in human toxicology and pharmacology. 1st edn, (CRC Press, 2004).
22 Eun, C. S. et al. Differences in gastric mucosal microbiota profiling in patients with chronic gastritis, intestinal metaplasia, and gastric cancer using pyrosequencing methods. Helicobacter 19, 407-416 (2014).
23 Khosravi, Y. et al. Culturable bacterial microbiota of the stomach of Helicobacter pylori positive and negative gastric disease patients. Scientific World J 2014, 610421 (2014).
24 Sachs, G., Weeks, D. L., Melchers, K. & Scott, D. R. The gastric biology of Helicobacter pylori. Annu Rev Physiol 65, 349-369 (2003).
25 Giannell.Ra, Zamcheck, N. & Broitman, S. A. Gastric-acid barrier to ingested microorganisms in man: studies in-vivo and in-vitro. Gut 13, 251-256 (1972). 26 Martinsen, T. C., Bergh, K. & Waldum, H. L. Gastric juice: a barrier against
infectious diseases. Basic Clin Pharmacol Toxicol 96, 94-102 (2005).
27 Mowat, C. et al. Omeprazole, Helicobacter pylori status, and alterations in the
intragastric milieu facilitating bacterial N-nitrosation. Gastroenterology 119, 339-347 (2000).
28 Streeter, A. M., Duraiappah, B., Boyle, R., O'Neill, B. J. & Pheils, M. T. Malabsorption of vitamin B12 after vagotomy. Am J Surg 128, 340-343 (1974). 29 Degnan, P. H., Taga, M. E. & Goodman, A. L. Vitamin B12 as a modulator of gut
microbial ecology. Cell Metab 20, 769-778 (2014).
30 Kikugawa, K. & Kato, T. Formation of a mutagenic diazoquinone by interaction of phenol with nitrite. Food Chem Toxicol 26, 209-214 (1988).
31 Davies, R. & McWeeny, D. Catalytic effect of nitrosophenols on N-nitrosamine formation. Nature 266, 657-658 (1977).
32 Brackman, W. & Smit, P. J. Studies in homogeneous catalysis: Kinetics and
mechanism of the copper-catalysed reaction of nitric oxide and diethylamine (I). Recl
Trav Chim Pays-Bas 84, 357-371 (1965).
33 Lin, J. T. & Stewart, V. Nitrate assimilation by bacteria. Adv Microb Physiol 39, 1-30 (1998).
34 Patel, B. A. & Crane, B. R. When it comes to antibiotics, bacteria show some NO-how. J Mol Cell Biol 2, 234-236 (2010).
376 377 378 379 380 381 382 383 384 385 386 387 388 389 390 391 392 393 394 395 396 397 398 399 400 401 402 403 404 405 406 407 408 409 410 411 412 413
35 Benjamin, N. et al. Stomach NO synthesis. Nature 368, 502 (1994).
36 Bowman, L. A., McLean, S., Poole, R. K. & Fukuto, J. M. The diversity of microbial responses to nitric oxide and agents of nitrosative stress close cousins but not identical twins. Adv Microb Physiol 59, 135-219 (2011).
37 Mitsui, T. & Kondo, T. Increased breath nitrous oxide after ingesting nitrate in patients with atrophic gastritis and partial gastrectomy. Clin Chim Acta 345, 129-133 (2004).
38 Edgar, R. C. Search and clustering orders of magnitude faster than BLAST.
Bioinformatics 26, 2460-2461 (2010).
39 Schloss, P. D. et al. Introducing mothur: open-source, platform-independent,
community-supported software for describing and comparing microbial communities.
Appl Environ Microbiol 75, 7537-7541 (2009).
40 Edgar, R. C. UPARSE: highly accurate OTU sequences from microbial amplicon reads. Nat Methods 10, 996-998 (2013).
41 Costea, P. I., Zeller, G., Sunagawa, S. & Bork, P. A fair comparison. Nat Methods 11, 359 (2014).
42 Dray, S. & Dufour, A. B. The ade4 package: Implementing the duality diagram for ecologists. J Stat Softw 22, 1-20 (2007).
43 Segata, N. et al. Metagenomic biomarker discovery and explanation. Genome Biol 12, R60 (2011).
44 Dixon, P. VEGAN, a package of R functions for community ecology. J Veg Sci 14, 927-930 (2003). 414 415 416 417 418 419 420 421 422 423 424 425 426 427 428 429 430 431 432 433 434 435 436
Acknowledgements
This work was supported in part by the Ministry of Science and Technology (NSC-103-2314-B-030-004-MY2) and the Biodiversity Research Center of Academia Sinica, Taiwan.
Author Contributions
CHT, JTL, and CYW conceived the study design; ZLL and CYW collected tissue samples; ZLL performed molecular experiments; CHT, HJH, and CBW conducted bioinformatics analyses; CHT, ZLL, SLT, and CYW authored the first draft. All authors contributed to data interpretation and approved the final manuscript.
Additional Information
Supplementary information accompanies this paper at http://www.nature.com/srep Accession codes: Illumina pair-end sequencing reads of 24 samples were deposited in the
NCBI Sequence Read Archive (accession number SRP057951).
Competing financial interests: The authors declare no competing financial interests.
Figure Legends
Figure 1. Bacterial community composition of human stomach, before and after surgery. (a) Average relative abundance of phyla across all samples, divided by tissue type and surgery
status. (b) Relative abundance of classes across all samples. (c) Principal component analysis of bacterial genera abundance, with before and after surgery serving as instrumental variable.
(d) Top-10 known genera with the highest LDA effect sizes reported by LEfSe in the stomach
bacterial community, before and after surgery. 1This Prevotella genus is affiliated with Prevotellaceae. 2This Prevotella genus is affiliated with Paraprevotellaceae, a recommended family (based on the Greengenes database).
Figure 2. Bacterial community structure dissimilarity summary. Dissimilarity was
measured using Bray-Curtis distance of beta diversity (genus level) and group difference was 437 438 439 440 441 442 443 444 445 446 447 448 449 450 451 452 453 454 455 456 457 458 459 460
determined (two-tailed Wilcoxon rank-sum test, with significance and sample size N noted on the figure). (a) Comparison of dissimilarity values among communities before and after surgery. (b) Comparison of dissimilarity values among communities in tumor and non-tumor tissues before surgery. (c) Comparison of dissimilarity values among communities in gastric stump and high body tissues after surgery. (d) Comparison of dissimilarity values among communities within and between patients, before and after surgery. Abbreviations: w/n, within; b/t, between; pt., patient.
Figure 3. Abundance of metabolic gene families differentially enriched in the gastric microbiota before and after surgery. Significance was defined as adjusted P<0.05. (a)
Metabolic functions enriched in the before-surgery microbiota. (b) Metabolic functions enriched in the after-surgery microbiota. Horizontal lines indicate group means (based on normalized relative frequencies).
461 462 463 464 465 466 467 468 469 470 471 472
Tables
Table 1. Bacterial community diversity indices based on 16S rRNA gene libraries.
Sample # OTUa # Singleton
OTU N
b Evenness
c Richness
d Shannon Simpson Chao 1 Good’s
coveragee T584N 311 158 26306 0.469 35.520 2.694 0.199 766 0.994 T584T 325 158 26306 0.465 35.520 2.692 0.172 837 0.994 T594N 258 158 26306 0.141 35.520 0.785 0.746 829 0.994 T594T 324 210 26306 0.336 47.284 1.944 0.279 888 0.992 T596N 261 158 26306 0.228 35.520 1.269 0.567 849 0.994 T596T 290 184 26306 0.505 41.402 2.863 0.184 939 0.993 T610N 315 184 26306 0.385 41.402 2.212 0.301 824 0.993 T610T 374 210 26306 0.363 47.284 2.152 0.335 949 0.992 T635N 268 184 26306 0.190 41.402 1.063 0.670 849 0.993 T635T 327 210 26306 0.399 47.284 2.312 0.168 970 0.992 T636N 302 184 26306 0.350 41.402 2.001 0.257 1077 0.993 T636T 328 210 26306 0.395 47.284 2.290 0.182 910 0.992 T584S 1580 1105 26306 0.605 249.771 4.453 0.039 6720 0.958 T584B 1517 1105 26306 0.612 249.771 4.481 0.032 7941 0.958 T594S 3219 2815 26306 0.589 636.644 4.754 0.040 36522 0.893 T594B 974 815 26306 0.155 184.161 1.064 0.747 8842 0.969 T596S 1400 1000 26306 0.513 226.015 3.719 0.072 5986 0.962 T596B 1361 947 26306 0.529 214.024 3.817 0.073 5919 0.964 T610S 1199 842 26306 0.549 190.269 3.892 0.072 5631 0.968 T610B 720 447 26306 0.552 100.904 3.635 0.084 2427 0.983 T635S 886 526 26306 0.701 118.777 4.758 0.018 3413 0.980 T635B 911 552 26306 0.655 124.659 4.462 0.034 3251 0.979 T636S 1454 973 26306 0.606 219.907 4.415 0.059 6550 0.963 T636B 925 526 26306 0.608 118.777 4.154 0.077 3086 0.980
aOTUs were defined at the 97% sequence identity level, using hypervariable regions V1–V3 of 16S
rRNA gene sequences.
bThe read number was rarefied to the minimum sample size by resampling with 1000 iterations. Data
derived from all reads per sample are available (Supplementary Table S1).
cEvenness was defined as Shannon/ln(# OTU).
dRichness was defined as (# singleton OTU-1)/log
10N. The maximum value was (N-1)/log10N.
eGood’s coverage was defined as 1-(# singleton OTU)/N.
473 474 475 476 477 478 479 480 481 482