國立臺中教育大學環境教育研究所
碩士論文
經水解及醱酵之稻殼廢水再產能
的可行性研究
指導教授:林明瑞 博士
研 究 生:林一傑 撰
中 華 民 國 九 十 九 年 六 月
謝 誌
在研究所修業期間,首先要感謝恩師林明瑞教授對於學生在專業知識 及待人處事上殷切的指導,使學生受益匪淺,謹致上最誠摯的敬意及謝 意。論文口試期間,承蒙鄭幸雄、林秋裕、賴俊吉教授悉心指導,並提供 學生許多寶貴的意見,使本論文內容更佳完善,在此獻上最誠摯的感謝。 回想起過去艱辛做實驗的日子,幸虧後來有學弟冠忠、宜曄陪伴著我 一起同甘共苦,使沉重的工作量得以減輕,論文也順利完成。也感謝林素 華老師、劉惠元老師、劉思岑老師、品詩學姐、雅鳳學姐、倩毓學姐以及 筱苓學姐在課業及生活上給我的指導與鼓勵;也感謝我的同窗好友程帆一 直給我鼓勵及研究所同學品瑢、哲賓、鴻斌、令嫻、瑜君、綱玲、賀翔還 有許多我未提及的朋友同學及學弟妹在學業上的相互扶持,使得研究生活 得以充實,此溫暖情誼將長存我心。 最後以此論文獻給我最親愛的父母多年來的栽培及鼓勵,因為有你們 的全力支持與愛護,讓兒子得以順利完成學業。也感謝譽瑋、哥哥及弟弟 不斷的給我鼓勵及支持,在此對你們獻上最深的敬意。 林一傑 謹致 中華民國九十九年七月I
摘要
台灣每年約產生十萬公噸的稻殼廢棄物,可用於產能,但是經謝孟宏 (2010)的研究顯示,稻穀廢水經醱酵產氫後仍含有相當程度的COD及揮發 酸,很適合再進行後續產能。本研究利用經水解醱酵產氫後之稻殼廢水作 為基質,以了解這類廢水適合再產能的反應及最佳的操作條件為何。 首先,本研究先進行厭氧光合產氫與甲烷化反應槽的比較試驗,光合 產氫是以完全混合式與柱塞流式反應槽為試驗槽體,而甲烷化反應是以流 體化床反應槽為試驗槽體。由試驗結果發現,甲烷化流體化床反應槽有最 佳的產能率,為535cal / g-CODin,所以後續實驗以甲烷化流體化床為試驗 槽體,變化不同的操作條件。 在不同HRT之甲烷產能試驗中,就單位體積甲烷產率而言,當HRT操 作在6hrs時,有最佳之甲烷產率39.4mmole- CH4 / L.day;就單位基質甲烷 產率而言,以HRT為24 hr時,有最佳之甲烷產率,可達1.03 mmole- CH4 /g- CODin。在不同pH之甲烷產能試驗中,當pH為 7.0時,有最佳的甲烷產量,為0.92 mmole- CH4 /g- CODin。在不同進流COD濃度之甲烷產能試驗中,就
單位體積甲烷產量而言,以COD濃度為30000 mg/L時,有最佳之甲烷產率 51.1 mmole- CH4 / L.day;就單位基質甲烷產率而言,以COD濃度操作在 10000 mg/L時,有最佳之甲烷產率,可達1.00 mmole- CH4 /g- CODin。在不 同溫度之甲烷產能試驗中,以反應槽溫度為35℃時,有最高之甲烷產率, 為1.03 mmole- CH4 /g- CODin。在不同迴流流量之甲烷產能試驗中,以迴流 流量操作在1000 ml/min時,有最高之甲烷產率,為1.06 mmole- CH4 /g- CODin。 經醱酵產氫後之稻殼廢水的甲烷產能總迴歸式分別為:
(1)每克進流COD甲烷產率(mmole- CH4 / g-CODin)=有機負荷(kg-COD /
II
(2)單位體積反應槽甲烷產率(mmole- CH4 / L˙day )=有機負荷(kg-COD /
m3.day)1.049˙0.999│1000-迴流流量│˙0.836│溫度 - 35│˙0.771│pH-7.0│ 本研究之菌相大部分是以平頭長桿菌甲烷菌菌群為主。而在掃瞄式電 子顯微鏡菌相觀察照片比較下,發現迴流速率較快下甲烷菌生長得較細 小,推估是因為迴流速率較快,使得菌體間摩擦力較大,黏附的菌體容易 脫落,因此導致菌體長得較為細小。 由基質利用率P值可發現,各個試驗的P值大都介於1.00左右,代表各 個試驗的反應槽在平衡後最大甲烷產率並無太大差別,代表各個試驗最大 產能均能充分利用基質而達到最大產能。而在KS值方面,以不同COD濃度 之甲烷產能試驗最小,代表試驗過程中變換不同的COD濃度較快達到最高 基質轉換率;而不同溫度之甲烷產能試驗最大,代表試驗過程中變換不同 的溫度需經過較長時間才能達到最高基質轉換率,基質利用效率易受溫度 變化的影響。 曾智鉉(2007)的研究中指出,以二相式連續流反應槽(前段活性碳棉 攪拌式反應槽+後段流體化反應槽)進行污泥與酒糟複合基質的產能試 驗,後段甲烷化流體化床最佳的單位體積甲烷產率為37.6 mmole- CH4 / L˙
day、最佳單位基質甲烷產率為0.895mmole-CH4 / g-CODin與最佳COD去除
率為30.3%,而本研究最佳的單位體積甲烷產率為43.7 mmole- CH4 / L˙ day
、最佳單位基質甲烷產率1.06mmole-CH4 / g-CODin為與最佳COD去除率為
35.0%,代表本試驗之產甲烷與去除COD效果算是不錯的,且產能後之廢 水通常都經由處理過後而直接排出,如果能回收再加以繼續產能,更可增 加其經濟效益,所以利用經水解醱酵產氫後之稻殼廢水再產甲烷的可行性 高。 關鍵字:稻殼、厭氧醱酵產氫後廢水之處理、厭氧光合產氫、甲烷化反應、 流體化床反應槽
III
The reenergization of Rice Husk Wastewater after
Fermentative Hydrogenesis
Abstract
The wastes of rice husk in Taiwan amount to one hundred million tons per year, which can be used for energy production. However, according to Hsieh(2010), rice husk wastewater contains a high strength of COD and volatile acids even after fermentative hydrogenesis, thus is suitable for further production. The rice husk wastewater after fermentative hydrogenesis was used as substrate in the study, in order to explore whether the wastewater was
switable for re-energization and what was optimal operating conditions for further energy production.
First, a re-energization comparison between the anaerobic photosynthetic hydrogen producing and methane producing reactors was conducted in the study. Both the complete mixing reactor and plug flow reactor was used as photosynthetic hydrogenic reactors . The fluidized bed reactor was used as the experimental reactor for methanogenic reactor. The results were found that methanogenic fluidized bed reactor has the optimal energy production of 535 cal / g-CODin. Thus, the methanogenic fluidized bed reactor was used as the experimental reactor to test various operating conditions in subsequent experiments.
In methane production tests of various HRTs, for the daily methane production of per unit volume reactor, when HRT was 6hrs, the optimal methane production was got, 39.4mmole- CH4 / L.day. In term of methane production of per gram influent COD, when HRT was 24hrs, the optimal
IV
methane production could be reached, 1.03 mmole- CH4 /g- CODin. In
methane production tests of various pH, when pH was 7.0, the optimal methane production was got, 0.92 mmole- CH4 /g- CODin. In methane product tests of various influent COD concentrations, as for the daily methane production of per unit volume reactor, when the COD concentration was 30000 mg/L, the optimal methane production was 51.1 mmole- CH4 / L.day. In term of methane production of per gram influent COD, when the COD concentration was 10000 mg/L, the optimal methane production was got, 1.00 mmole- CH4 /g- CODin. In methane production tests of various temperatures, when the reactor temperature was 35℃, the highest methane production was got, 1.03 mmole- CH4 /g-
CODin. In methane production tests of various recycling flow rates, when recycling flow rate was 1000 ml/min, the highest methane production was got, 1.06 mmole- CH4 /g- CODin.
The general regression equation of methane production from rice husk wastewater after fermentative hydrogenesis is as follows:
(1) methane production of per gram influent COD (mmole- CH4 / g-CODin) = organic loading(kg-COD / m3.day)-0.051˙0.999│1000- recycling flow rate│˙0.834│temp - 35│
˙0.771│pH-7.0│
(2) methane production of per unit volume reactor (mmole- CH4 / L˙day ) = organic loading (kg-COD / m3.day)1.049˙0.999│1000-recycling flow rate│˙0.836│temp - 35│
˙0.771│pH-7.0│
The methanogenic bacteria with flat-end was main bacterial consoria. Being observed by scanning electron microscope (SEM),it was shown that the methanogenic bacteria are smaller and thinner when growing under faster recycling flow rate, possibly due to the higher friction force under higher
V
recycling flow rate, thus the adhesive bacteria may easily fall off.
Most substrate utilization rate P in the tests were around 1.00, indicating that the maximum methane production of the reactors in all tests did not have significant difference when reached balance. This indicated that the maximum production in all tests could fully utilize the substrate. The KS was the lowest in the methane production tests of various COD concentration, indicating that the change of the COD concentrations during the tests could achieve the maximum substrate conversion rate faster. The substrate utilization rate is less likely to be affected by the changes of COD concentration. KS was the highest in the
methane production tests of various temperatures, indicating that changing the temperatures during the tests would require a longer time to achieve the
maximum substrate conversion rate. It was because the substrate conversion rate was affected by temperature changes.
The research of Tseng(2007) reported that when HRT was 6hrs, the daily methane production of methanogenic fluidized bed reactor of rear stage could reach 37.6 mmole- CH4 / L˙day. In this study, when HRT was 6hrs, the daily methane production could reach 39.4 mmole- CH4 / L˙day, indicating that the methane production is satisfactory. Since rice husk wastewater is often
discharged after treatment, if it could be recycled and reused, it could bring more economic benefits. Therefore, rice husk wastewater after fermentative hydrogenesis being reused for methane production is highly feasible.
Keywords: rice husk, re-energization after anaerobic fermentative
hydrogenesis, anaerobic photosynthetic hydrogen production, methanogenic reaction, fluidized bed reactor
VI
VII
目錄
摘要 ... I Abstract ... III 目錄 ... VII 表目錄 ... IX 圖目錄 ... XI 第一章 緒 論 ... 1 第一節 研究背景與動機 ... 1 第二節 研究目的 ... 4 第三節 名詞解釋………...5 第二章 文獻回顧 ... 7 第一節 能源危機與生質能源...7 第二節農業廢棄物探討-稻殼………..………...11 第三節 農業廢棄物產能可行性探討……….………13 第四節 厭氧產能微生物……….14 第五節 厭氧微生物產能之機制……….20 第六節 環境因子對厭氧產能的影響……….25 第七節 各種厭氧產能反應槽之探討……….41 第八節 厭氧產能反應動力學探討……….44 第九節 厭氧醱酵產能之迴歸……….48 第三章 研究方法 ... 49 第一節 試驗基質 ………... 49 第二節 反應槽及試驗設備 ... 49 第三節 試驗設計與流程 ... 56 第四節 分析方法與項目……….59VIII 第四章 結果與討論 ... 65 第一節 光合產氫與甲烷化反應槽之產能比較試驗……….65 第二節 不同水力停留時間之甲烷產率比較試驗……….71 第三節 不同 pH 之甲烷產率比較試驗………..74 第四節 不同 COD 濃度之甲烷產率比較試驗………...80 第五節 不同溫度之甲烷產率比較試驗……….84 第六節 不同迴流速率之甲烷產率比較試驗………...89 第七節 三段式反應槽之總產能評估………...94 第八節 質量平衡…...95 第九節 反應動力學之探討及模擬……….99 第十節 菌相觀察………...105 第五章 結論與建議 ... 109 第一節 結論 ... 109 第二節 建議 ... 111 參考文獻 ... 113
IX
表目錄
表2-1 稻殼之主要成分………...13 表2-2 厭氧產氫微生物之種類………...16 表2-3 各種不同菌種進行醱酵代謝之產物………...17 表2-4 各種厭氧污泥中甲烷菌之種類及特性………...20 表2-5 紫色不含硫光合作用細菌對不同電子供給者利用情形………31表2-6 在各種氣體成分條件下 Rhodobacter sphaeroides O.U.001*其產氫 轉換率及基質消耗量………..33
表2-7 各種厭氧生物廢水處理技術評估表………...43
表3-1 Rhodospiriaceae 培養基成分……….…51
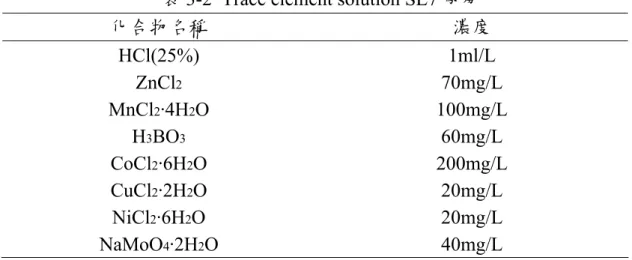
表3-2 Trace element solution SL7 成分………51
表3-3 光合產氫與甲烷化反應槽之產能比較試驗設計……….……57 表3-4 不同水力停留時間 HRT 之甲烷產能比較試驗設計……….….57 表3-5 不同 pH 之甲烷產能比較試驗設計……….58 表3-6 不同進流 COD 濃度之甲烷產能比較試驗設計……….…58 表3-7 不同溫度之甲烷產能比較試驗設計………...58 表3-8 不同迴流速率之甲烷產能比較試驗設計………..….59 表4-1 不同反應槽之累積產氣及產能………...68 表4-2 不同反應槽的進出流水之水質分析………...69 表4-3 甲烷化流體化床反應槽於不同 HRT 下,累積甲烷產量及產氣 組成之比較……….………...72 表4-4 甲烷化流體化床反應槽於不同 HRT 下,其進出流水水質之變 化情形…….………...73 表4-5 甲烷化流體化床反應槽於不同 pH 下, 累積甲烷產量及產氣組 成之比較……….………...76
X 表4-6 甲烷化流體化床反應槽於不同 pH 下,其進出流水水質之變 化情形………...78 表4-7 稻殼廢水不同操作 pH 值下,反應槽中 YpH與pH 校正係數 θ…..80 表4-8 甲烷化流體化床反應槽於不同 COD 濃度,累積甲烷產量及產 氣組成之比較……….………...81 表4-9 甲烷化流體化床反應槽於不同進流 COD 濃度下,其進出流水 水質之變化情形……….………...83 表4-10 有機負荷與甲烷產率之對照表……….83 表4-11 稻殼廢水不同操作有機負荷下,反應槽中 Y單位基質甲烷產率與 Y單位體積甲烷產率………...84 表4-12 甲烷化流體化床反應槽於不同溫度下,累積甲烷產量及產氣 組成之比較………...86 表4-13 甲烷化流體化床反應槽於不同溫度下,其進出流水水質之變 化情形………...87 表4-14 稻殼廢水不同操作溫度下,反應槽中 Y溫度與溫度校正係數θ…..89 表4-15 甲烷化流體化床反應槽於不同迴流流量下,產氣組成及累積 甲烷產量之比較………...91 表4-16 甲烷化流體化床反應槽於不同迴流流量下,其進出流水水質 之變化情形………...91 表4-17 稻殼廢水不同操作迴流流量下,反應槽中 Y溫度與迴流流量校 正係數θ………..………...93 表4-18 各組試驗 COD 質量平衡及回收率………..98 表4-19 以 Michaelis-Menten 模擬連續流各試驗基質利用率之計算值….101 表4-20 以 Michaelis-Menten 迴歸所求得動力學參數……….102
XI
圖目錄
圖2-1 傳統厭氧消化反應過程………..22 圖2-2 主要甲烷化作用途徑………...25 圖2-3 中溫消化與高溫消化之關係圖………..…37 圖2-4 有機物之厭氧分解反應機制圖………..45 圖3-1 柱塞流式反應槽示意圖………..52 圖3-2 完全混和式反應槽示意圖………..53 圖 3-3 甲烷化生物流體化床反應槽示意圖……….55 圖3-4 揮發酸成分分析前處理流程圖……….…..61 圖4-1 不同種類反應槽,其單位體積反應槽每天產能之比較圖…………67 圖4-2 甲烷化流體化床反應槽於不同 HRT 下,每天單位體積反應槽 甲烷產率之比較圖………...72 圖4-3 甲烷化流體化床反應槽於不同 pH 下,每天單位體積反應槽甲 烷產率之比較圖………...76 圖4-4 甲烷化流體化床反應槽於不同 COD 濃度下,每天單位體積反 應槽甲烷產率之比較圖………...81 圖4-5 甲烷化流體化床反應槽於不同溫度下,每天單位體積反應槽甲 烷產率之比較圖………...86 圖4-6 甲烷化流體化床反應槽於不同迴流流量下,每天單位體積反應 槽甲烷產率之比較圖………...90 圖4-7 甲烷化流體化床反應槽之不同 HRT 反應動力學迴歸直線………102 圖4-8 甲烷化流體化床反應槽之不同 pH 反應動力學迴歸直線…………103 圖4-9 甲烷化流體化床反應槽之不同 COD 濃度反應動力學迴歸直線…103 圖4-10 甲烷化流體化床反應槽之不同溫度反應動力學迴歸直線……...104 圖4-11 甲烷化流體化床反應槽之不同迴流速率反應動力學迴歸直線…104XII 圖4-12 甲烷化流體化床反應槽之位相差顯微鏡菌相觀察照片(進流 基質COD 濃度 15000mg/L、HRT=12hrs、放大倍率 600 倍) ………..106 圖4-13 甲烷化流體化床反應槽之螢光顯微鏡菌相觀察照片(進流 基質COD 濃度 15000mg/L、HRT=12hrs、放大倍率 600 倍) ………...106 圖4-14 甲烷化流體化床反應槽中之菌相掃瞄式電子顯微鏡照片(進 流基質COD 濃度 15000mg/L、HRT=12hrs、迴流流量 1000 mg/min、放大倍率 3000 倍)……….………...107 圖4-15 甲烷化流體化床反應槽中之菌相掃瞄式電子顯微鏡照片(進流 基質COD 濃度 15000mg/L、HRT=12hrs、迴流流量 650mg/ min、放大倍率 3000 倍)……….………...107
1
第一章 緒論
本章主要在說明研究的背景、動機及目的。全章共分二節:第一節為 研究背景及動機、第二節為研究目的。第一節 研究背景與動機
能源是人類經濟發展的推手,能源危機常涉及到石油,電力或其他自 然資源的短缺。化石燃料為人類目前最主要的使用能源,然而世界各國蘊 藏量已逐漸匱乏,依據現有資料顯示,預估石油可開採41年、天然氣67年、 煤192年(曲新生,2005)。能源短缺通常會使得經濟發展受到嚴重限制, 很多突如其來的經濟衰退通常就是由能源危機引起的。而在過去能源危機 的主要原因是石油價格過於便宜,以致於使世界對其產生了過度的依賴性 造成石油迅速消耗殆盡。化石燃料的過度開發導致趨於耗竭,不僅對生態 環境造成傷害,且燃燒化石燃料更會造成空氣污染及酸雨、溫室效應等現 象。 生質能源是一種極具開發潛力的再生能源,一般用來轉換成能源的生 物質是經過生物產生的有機質,如木材、農產品、能源作物與農、林、畜 牧業廢棄物等。植物的有機物是由光合作用將吸收的太陽能轉變成化學能 後,將水、二氧化碳與土壤吸收來的各種礦物元素同化成的代謝產物(碳 水化合物、蛋白質、油脂等)。將這些有機質轉換成不同形式的生質能源 後燃燒,又將固定的碳釋放回大氣中循環,因此並不會產生額外的二氧化 碳,可減少環境污染,生產技術困難降低,物料來源生生不息,故為符合 環保與永續之綠色再生能源。目前主要回收的能源有氫及甲烷,在產氫方 面以厭氧醱酵產氫技術最具有發展潛力;而在產甲烷方面,則利用傳統厭2 氧消化反應,有機物經微生物水解、酸化、甲烷化而生成甲烷氣體。 由於農業廢棄物因富含大量的纖維素(cellulose)、半纖維素(hemi- cellulose)及木質素(lignocellulose)…等成分,相當難以分解,必須先使用水 解菌加以分解其中纖維後再以醱酵產氫菌來進行厭氧醱酵加以反應以提 升產H2、產能效果。而纖維素經水解細菌分解反應後,使能源作物基質可 以被產氫菌加以利用,使經過水解、醱酵、酸化等過程得以回收H2能源。 根據樊國恕(2002)的研究指出,厭氧發酵過程可分為三個階段:水解、 酸化、甲烷化,而複雜有機物首先經由非甲烷菌分解為揮發酸及二氧化碳 等中間產物,而後再藉由甲烷菌轉換為甲烷。張嘉修(2008)的研究中指出, 光合菌能夠使用乙酸以及丁酸當做碳源,對於厭氧產氫之代謝物以及其它 發酵程序之代謝產物(如乙酸、丁酸等),光合菌都能直接利用該代謝物並 產生氫氣。由此推估經由水解、醱酵產氫後之出流水,內含豐富的揮發酸 及小分子的有機物質,十分適合光合產氫菌及甲烷菌再加以利用繼續產 能。 根據謝孟宏(2009)研究利用好氧循序批次反應槽( Sequencing Batch Reactor,SBR)串聯厭氧發酵CSTR反應槽,處理農業廢棄物稻殼廢水,搭 配批次試驗所得的最佳操作條件來醱酵產氫,發現其出流水仍含有大量的 有機物質、適量的COD濃度與揮發酸,並且把稻殼由半固體物轉化為液 體,所以十分適合厭氧光合產氫菌與甲烷菌加以利用繼續產能。所以本研 究將利用其出流水於產能(H2+CH4),並且評估可回收再利用部分。 在本實驗室陳嘉正(2003)厭氧光合產氫的研究中指出,進流每克 COD 產氫率及單位體積反應槽產氫率方面以柱塞流式反應槽最佳,且柱塞流式 反應槽長柱狀且橫躺的形式使WP3-5 不易流失而易於增殖,因此柱塞流式 反應槽有較高的生物污泥濃度,加上其光照度也最佳,故為三種反應槽中
3 最佳的光合產氫率;完全混合式反應槽因有攪拌,而有較佳的質傳效率。 所以本研究選用柱塞流反應槽與完全混和式反應槽進行厭氧光合產氫光 合試驗。 在本研究室過去的研究中,研究結果顯示厭氧醱酵產氫的確有不錯的 產氫效率,且反應過後的出流水中含有大量的揮發性有機酸。再者根據光 合產氫菌群之相關研究文獻顯示,厭氧光合產氫系統非常適合處理厭氧醱 酵產氫之出流廢水,因厭氧醱酵產氫之出流水含有大量之揮發性有機酸, 正是適合於光合產氫菌群之生長基質,且光合產氫菌群亦會產生氫氣,可 達到廢水處理及產生氫氣之雙重效果(紀又華,2006)。 在本實驗室曾智鉉(2007)的研究中指出,以後段甲烷化活性碳流體化 床反應槽,用以處理前段醱酵產氫活性碳棉攪拌式反應槽產氫後所排出之 廢水,能達到第二相甲烷產能的目標。因此本實驗將會比較兩種產能方式 (光合產氫與甲烷化)搭配不同的反應槽以比較何種會有最佳的產能效率。 進流基質方面,本實驗室謝孟宏(2009)反應槽之出流水: COD 濃度大 約為11,000 ~ 17,000 mg/L、揮發酸濃度大約為 2,000 ~ 3,000 mg/L,十分適 合厭氧光合產氫菌與甲烷菌的生長,因此本研究以本實驗室謝孟宏(2009) 反應槽的出流水為進流基質。在厭氧光合產氫微生物方面,以光合作用產 氫細菌中產氫能力最好的菌株Rhodopseudomonas palustris ,菌株編號為 WP3-5 以其生理特性及所需生長環境的各項條件為基礎;在甲烷化反應方 面,以甲烷化活性碳流體化床反應槽最適的生長條件操作,同時比較三種 反應槽的產能情況,並且以產能效率最佳之反應槽,進行後續不同操作條 件下的厭氧產能試驗。
4
第二節 研究目的
本研究一開始分為以下兩部分: 一、以中興大學環工系李季眉教授實驗室研究篩選培養之光合作用產氫細 菌中產氫能力最好的菌株Rhodopseudomonas palustris,菌株編號為 WP3-5,與其生理特性及所需生長環境的各項條件為基礎,添加謝孟 宏(2009)反應槽出流水為基質,結合完全混合式反應槽(CSTR)與柱塞 流式反應槽。 二、以黎明汙水處理廠終沈池污泥與謝孟宏(2009)反應槽出流水做為複合 基質,結合流體化床反應槽。 主要目的為探討完全混合式反應槽與柱塞流式反應槽結合WP3-5,以 及流體化床反應槽結合黎明汙水處理廠終沈池污泥的產能效率比較研究; 接下來以最佳之反應槽來進行試驗,以評估產能之可行性及在何種操作條 件之下,可以得到最佳的產能效率。並進行產能效率之動力學模式,本研 究內容包括: 一、為能了解稻殼被好氧循序批次反應槽串聯醱酵產氫CSTR反應槽處理 之出流水,進行厭氧光合產氫與甲烷化反應之可行性為何。 二、為能了解利用完全混合式反應槽(CSTR)與柱塞流式反應槽進行光合產 氫,以及利用流體化床反應槽來進行甲烷化反應,何者產能效率較佳。 三、為能了解在不同操作條件(HRT、pH、進流COD濃度、溫度與迴流速 率)之下,最佳之反應槽的產能效率為何。 四、為能了解在何種操作條件之下,最佳之反應槽的產能效率最佳。 五、為能了解最佳之反應槽產能結果的反動力參數。5
第三節 名詞解釋
一、稻殼廢水 本研究中所提到之稻殼廢水,乃指謝孟宏(2010)的連續流試驗研究 中,利用循序批次水解反應槽(SBR)串聯醱酵產氫反應槽(CSTR)進行稻殼 的產氫試驗之出流水。而本研究將以其出流水作為光合產氫與甲烷化流體 化床反應槽連續流試驗的進流基質。7
第二章 文獻回顧
本章共分成八節分別為,能源危機與生質能源、農業廢棄物探討-稻 穀、農業廢棄物產能可行性探討、厭氧產能微生物、厭氧微生物產能機制、 環境因子對厭氧產能的影響、各種厭氧產能反應槽之探討、厭氧產能反應 動力學探討、厭氧醱酵產能之迴歸,做回顧探討。第一節 能源危機與生質能源
一、能源危機 能源與經濟的發展有相當密切的關係,在經濟發展與社會進步的過程 中,不管是工業發展、交通運輸或人民生活等,都與能源需求有關,隨著 生活水準的提高,對能源的依賴也越大,且絕大部分都是來自化石燃料, 同時也會產生大量的二氧化碳及廢氣(如硫氧化物、氮氧化物等)排放到 大氣中,及大量的生活廢水與工業廢水排放到河川海洋等水體,對全球自 然資源與生態環境產生莫大的破壞,例如:溫室效應、臭氧層的破壞、酸 雨、河川與海洋的污染等等問題(楊聰宏,2003)。 石油已知的經濟蘊藏量,按目前的價格和使用速度只能使用四十年, 產地又集中,造成供應上潛在的不安定性,經過二次能源危機的教訓,各 國都已經了解到能源多樣化的重要性。換言之,就是要把石油佔總能源使 用的比例降低。台灣目前石油的依存度已達到總能源使用量的53%,自不 宜再增加使用。 煤的蘊藏量豐富,造成的環境污染問題卻是最嚴重的。眾多技術雖然 能去除部分污染,卻無法解決因燃燒而造成的溫室效應問題。溫室效應問 題現已引起全球性的普遍關切,極有可能會簽訂國際公約限制各國二氧化8 碳的排放量,所以煤及其他化石燃料來發電的選擇就受到極大的限制。而 國內民眾對核能發電則多有疑慮。最後,水利發電要建築大型水庫,對環 境衝擊很大,也不是理想的發電方式。 從目前世界能源的消耗情況來看,以非再生能源的化石燃料為主,包 括石油、煤碳、天然瓦斯的比例高達九成,核能發電則佔7%,水力發電僅 佔3%(洪經文,1999)。而我國的經濟成長所帶動的能源需求,全靠化石 能源來提供,使得化石能源的需求逐年增加,二氧化碳排放也超過了經濟 成長速率(楊任徵,2000)。為了達成永續發展的目標,開發新的再生能 源是必須的,可見未來能源政策將由高污染性或高含碳的化石能源,轉為 低污染性或無污染性能源,特別是乾淨的再生能源比例(楊聰宏,2003)。 大量使用化石燃料的結果,除了立即可以感受到的空氣污染問題之 外,使用後所產生大量的二氧化碳,導致全球溫室效應、破壞地球的恆溫 系統,更是無形的殺手。地球上的礦物、石化能源有限,石油的預估可開 採年限平均約剩四十年,天然氣剩六十年,煤仍可210年,核能剩四十五 年(行政院經濟能源局,2006),顯示人類目前高度依賴使用的能源存量未 來逐漸枯竭,應該要朝開發新能源的方向前進,而再生能源的可永續利用 將能夠改善能源有限的問題(石林鍠,2006)。 二、再生能源(Renewable energy) 根據聯合國環境規劃署(UNEP)的定義,「再生能源」係指理論上取之 不盡的天然資源,過程中不會產生污染物,包括:太陽能、風力、生質能、 水力、地熱及海洋能等(經濟部能源局,2005),其均有共通特點,都是可 轉化自然界的能量來進一步運用(張聖宗,2004)。而政府在2005年時,把 再生能源納入能源發展政策,使再生能源至2020 年佔能源需求比率以3% 為目標(行政院經濟部能源局,2005)。再生能源主要包括太陽能、地熱能、
9 潮汐能、水力能、風力能、海洋溫差、海浪能及生質能等,在國內除傳統 水力能已大致被開發殆盡外,其中較有發展及應用潛力為太陽能、風力能 及生質能。 三、生質能源(Biomass energy) 生質能(Biomass energy)泛指由微生物所產生之有機物質,如動植 物、有機污泥等農業、畜牧業、工業、都市能源作物以及廢棄物,經過焚 化、氣化、裂解、醱酵等技術轉換成燃油(酒精汽油、生質柴油)、燃氣 (沼氣、氫氣)與電力等可用之能源,即係指利用生物質(biomass),經 過轉換所獲得的可用能源(工研院能資所,2006)。而我國將生質能定義 為「國內農林植物、沼氣、一般廢棄物與一般事業廢棄物等直接利用或經 處理所產生之能源」(再生能源發展條例草案,2002)。
根據國際能源總署(International Energy Agency)的統計資料顯示, 目前生質能為全球第四大能源,僅次於石油、煤及天然氣,供應了全球約 14%的初級能源需求,也提供了開發中國家 35% 的能源,同時也是目前 最廣泛使用的一種再生能源,約佔世界所有再生能源應用的80%。 若以生質能利用技術來區分,生質能運用技術大致可分為:燃料酒精 技術、生質柴油技術、沼氣利用技術、液化與氣化技術,以下就各技術來 說明(吳耿東、李宏台,2004): (1)燃料酒精技術 生質酒精(乙醇)是一個比較環保的燃料,過去數十年,主要採用 發酵法生產酒精,目前世界各國主要生產生質酒精的國家,除了巴西自 1970 年代石油危機起,以甘蔗為主要能源作物從事酒精燃料之研發與生 產外,另一主要國家為美國利用玉米澱粉來醱酵以生產酒精。而生質酒 精最經濟且實用的生產途徑有二,一是利用廢棄的農作副產品(稻梗、
10 廢糖蜜等)為原料生產乙醇;二是養殖綠藻生產乙醇,其中以廢棄農作 物來生產酒精最具潛力。 (2)生質柴油技術 利用油脂作物或廢食用油與甲醇(或乙醇)進行轉酯化反應,可產 生脂肪酸甲酯(或乙酯)及甘油等產物;經分離甘油後,以蒸餾去除未 反應完全的油脂,產生與一般柴油品質相當的液態燃料,稱為生質柴油。 (3)沼氣利用技術 沼氣的產生主要是藉由細菌把廢棄物中的有機物質分解以得到可燃 性氣體,主要成分是氫、甲烷、二氧化碳及少量硫化氫。 沼氣產生反應主要分成厭氧醱酵產氫反應與甲烷化反應兩部分。厭 氧醱酵反應主要是由厭氧產酸菌,在厭氧的環境下,利用基質中的大分 子有機物,經由醱酵反應,將大分子有機物分解為較小分子有機酸的過 程而產氫。而甲烷化反應是由甲烷菌,在嚴格厭氧的環境下,利用醱酵 反應中產生代謝產物,包括:揮發酸、H2、CO2等,轉化為甲烷。 目前生物法產氫技術主要分為三類,包括:暗醱酵法、光醱酵法與光 合作用法。而其中厭氧醱酵產氫法可分解有機物同時產生氫氣及CO2,而 氫氣是屬於乾淨的能源之一,其產能效率每克的氫氣中約有122千焦耳, 約為石油的3倍,甲烷的2.4倍,且燃燒過後只產生熱能和水,不會形成溫 室氣體(二氧化碳)而造成環境暖化,因此氫氣是未來最具發展潛力技術之 一。 甲烷化反應則是甲烷菌在嚴格厭氧狀態下,包括:中性 pH 質、揮 發酸…等適合的條件,利用醱酵產酸反應過程所產的揮發酸、H2、CO2, 來進行甲烷化反應轉化為甲烷氣體。而光合產氫菌可以利用醱酵產酸菌所 產生的揮發酸,在光源充足的厭氧環境下,利用產生氫氣。
11
第二節 農業廢棄物探討-稻殼
農業廢棄物包含農產廢棄物與農業資材廢棄物,這些農產廢棄物泰半 均可回收再利用,製成堆肥、廄肥,再回歸到農田,惟若隨意傾倒,丟棄 或燃燒亦或處理不當則均會造成環境污染。而根據楊紹榮(1997)的研究中 指出目前農業生產過程中所產生的廢棄物有:穀類廢棄物、特用作物廢棄 物、蔬果廢棄物、食品工廠廢棄物、菇類栽培介質廢棄物、禽畜及養殖廢 棄物、樹皮、庭園及行道樹修剪之廢棄物等七大類。而目前主要有幾種農 業廢棄物的處理機制,不外乎:在田間進行堆肥處理,使其腐熟、直接焚 燒成灰燼,讓其營養份回歸土壤中、或翻入田間土壤中,令其自然腐化作 為綠肥。於是有些學者開始考慮農業廢棄物的再利用技術,甚至能源化技 術。 目前農業方面每年都產生大量的廢棄物,但其中大部分沒有得到充分 利用,若能選擇適當的處理技術,加以再利用,將能減輕能源之使用量, 也可減少大量之農業廢棄物。 一、農業廢棄物可應用的前處理技術 農業廢棄物因富含大量的纖維素(cellulose)、半纖維素(hemicellulose) 及木質素(lignocellulose)…等成分,相當難以分解,而稻殼一般成份為粗蛋 白質3.25%、纖維素 43.3%、木質素 22%、木醣 17.5%,因此必須採取適 當的前處理技術,而國內主要的農業廢棄物之前處理技術,目前計有:加 熱處理(李傳華,2003),鹼化處理(Beckmann,1919),酵素前處理(張殿傑、 武雲東,2001),化學處理(Ngu et al., 2001),超音波處理(張永和,2004), 以及冰凍處理(林月惠等,2001)等技術。 根據李傳華(2003)的研究指出利 用 水 解 技 術 , 以甘 蔗 渣 、 樹 葉 和 菜 皮 三 種 物 質 為 材 料 , 並 加 入 催 化 劑 Na2CO3 的 條 件 下 , 上 述 廢 棄12 物 可 轉 化 得 到 含 有 大 量 的 腐 植 酸 物 質 , 最 高 能 達 到 45% 左 右 , 而 這 類 腐 植 酸 物 質 可 以 作 為 生 態 肥 料,具 有 良 好 的 肥 效 和 經 濟 價 值 。 而臧金燦、樊國燕(2003)研究中指出,稻桿的鹼化處理可使植物細胞壁 變得鬆散,易於分解處理。張殿傑、武雲東(2001)的研究中指出,採用 BYIM(農用酵素)處理稻桿製成酵素菌堆肥,施放於田間可增加土壤有 機質,促進土壤團粒結構形成,改良土壤結構;提高土壤肥力,抑制土傳 病害,提高作物的產量,改善產品品質。 在Ngu et al.,(2001)的研究指出將化學氧化劑,諸如過氧化氫(Hydrogen Peroxide)、高錳酸鹽(Permanganate)及臭氧(Ozone)等加入受污染之作物 中,藉由氧化作用將污染作物氧化產生二氧化碳或轉化污染作物為無毒性 之化合物。在Beckmann(1919)研究中提出以 1.5%的 NaOH 溶液將稻桿在 室溫下浸泡3 天後,再將多餘的鹼用水沖掉,結果麥稻的有機物消化率由 45.7%提高到 71.2%。 由上述的文獻可以發現農業廢棄物若含大量的纖維素(cellulose)、半纖 維素(hemicellulose)及木質素(lignocellulose)等難以分解成分,若能選擇適 當的前處理技術,將可有效分解像纖維素等難以分解有機物質,而經分解 後之有機物可經由後段的能源化技術來產生能源,使能源作物之產能效益 提升。 二、稻殼成分分析及用途 水稻為本省最重要的農作物,無論種植面積或者是產值均高居各類 農畜產品之首位,其主要廢棄物有稻草及稻殼。以下主要探討對象為稻 殼。 (一)稻殼成分 加工時所產生的廢棄物主要有稻殼、米糠、脫脂米糠及白米糠,其
13 成分如表2-1所示: 表2-1 稻殼之主要成分 項目 百分比(%) 粗蛋白質 3.25 粗纖維 44.30 乙醚抽出物 0.85 灰分 20.75 無氮抽出物 30.85 (二)稻殼的處理方式 稻殼是加工生產之副產品,台灣省每年之稻殼廢棄物約有60萬公噸, 過去的碾米場和農會是採取露天燃燒或者傾到在河川空地,造成環境相當 大的影響,對於農民而言,過去的稻殼為主要的農家燃料。目前的稻殼僅 有少量作為飼料、建築材料之填充料,極少部分作為果園、花卉及蔬菜的 畦面敷蓋。所以如果能有效的處理稻穀並且加以利用,不但不會對環境造 成汙染,更可以達到廢棄物再產能的效果。
第三節 農業廢棄物產能可行性探討
在林茜羽(2007)的研究中,針對農業廢棄物-稻草桿,利用厭氧生物醱 酵技術進行菌種篩選、溫度、基質濃度及pH 選定等各因子之評估,期望 藉由機制與效能評估等觀點,提供稻桿生質能醱酵之可行性。結果顯示經 過熱前處理之中興新村植種污泥在55℃有較高的累積產氫率 0.7 ml/ VSS g。初始 pH 試驗範圍為 3.5~8,最理想的產氫初始 pH 為 6.5,其產氫率為 0.79 ml/ VSS g。將稻桿基質粒徑分為<0.297mm、0.297~0.58mm、0.5814 ~1.19mm、大於 1.19mm 及 10 mm(1 cm),得知粒徑小於 0.297 mm 能獲 得較大的產氫率0.67 ml/ VSS g。基質濃度測試範圍在 30~120 g 稻桿/L, 當基質濃度為90 g 稻桿/L 有最大的氫氣產率,一克的總揮發性固體量有 77.2 ml 的氫產量(77.2 ml/g TVS)相當於 0.77 mmol/g cellulose。 在蘇偉昌(2008)的研究中指出,微波酸水解液經過量鹼處理後均可有 效轉化成氫氣。其中柳杉及蔗渣以磷酸水解液可獲致最大氫氣產率,分別
為5.22 及 4.91 mmol H2/g total sugar。而稻稈以鹽酸水解液可獲致最大氫氣
產率4.02 mmol H2/g total sugar。在二階段微波酸水解液醱酵產氫試驗中,
經過量鹼處理之柳杉及稻稈之水解液,其氫氣產率分別為5.56 及 5.51
mmol H2/g total sugar。
在謝孟宏(2009)的研究中指出,在SBR+CSTR 以稻殼當作進流基質 中,搭配水解菌與厭氧醱酵產氫菌,當總循序時間+HRT 為 6+18hr 時, 有較佳的單位體積產氫率,為19.8 mmole-H2/L.day。由以上的文獻得知, 只要農業廢棄物在適宜的操作條件與環境下,便可有效的加以利用,並且 達到再能源化的目標。
第四節 厭氧產能微生物
本研究在一開始的試驗一中,將會進行厭氧光合產氫反應與甲烷化反 應的產能比較試驗,因此本研究有必要對產氫菌及甲烷菌進行探討,以瞭 解其生長特性。 一、厭氧產氫微生物早期Toerien (1967)、Iannotti (1973)等人曾利用Vibrio succinogenes菌 株,將葡萄糖等碳水化合物,於厭氧環境下,醱酵產生氫氣。另外Karube
15 (1982)則將Clostridium butyricum菌種利用固定化技術,於厭氧酸化過程之 酸形成相中,亦能有效產生氫氣。因此,由過去許多的研究中發現,有很 多微生物均具有產氫能力,如表2-2 (Das & Veziroğlu, 2001)。但因為各產 氫菌生理特性的不同,其產氫能力也不同,對於環境的要求亦有所差異 (林 秋裕、林明正、陳晉照,1999)。 根據Yokoi et al. (1995)的研究指出,產氫菌主要分為光合作用產氫菌 (Photosynthetic microorganisms)和厭氧醱酵產氫菌 (Fermentative hydrogen producing microorganisms)二大類。
光合作用微生物部份包括:紫色不含硫菌、藻類等。醱酵產氫微生物 又依其對氧需求性分為兼性厭氧菌 (如Bacillus、Enterobacter aerogenes 等),及絕對厭氧菌 (如Clostridium等)。各方資料 (Yokoi, 1997;Odom, 1983) 均顯示:目前研究以紫色不含硫菌產氫效率最佳,其產氫量為厭氧菌的 1.5-2.0倍,但需要照光及靜置的環境中生長;而醱酵產氫微生物中以 Clostridium菌屬為主,此菌屬大部分都可以產氫,其中以Clostridium butyricum菌種產氫更是可觀,醱酵產物為butyrate、butanol及Acetone。表 2-2為各種不同產氫菌進行厭氧醱酵之產物 (Göttschalk, 1986)。而本實驗產 氫反面主要以紫色不含硫菌為實驗菌種,所以以下將以紫色不含硫菌為主 進行探討。
16 表2-2 厭氧產氫微生物之種類 (Das & Veziroğlu, 2001) 微生物分類 微生物名稱 Green algae (綠藻) Scenedesmus obliquus Chlamydomonas reinhardii C. movewusii Cynobacteria Heterocystous (藍綠菌) Anabaena azollae Anabaena CA A. variabilis A. cylindrical Nostoc muscorum N. spongiaeforme Westiellopsis prolifica
Nonheterocystous Plectonema boryanum Oscillotoria Miami BG7 O. limnetica Synechococcus sp. Alphanothece halophytico Mastidocladus Phormidium valderianum Photosynthetic bacteria (厭氧光合細菌) Rhodobater sphaeroides R. palustris R. capsulata Rhodospirillum rebnum Chromatium sp. Miami PSB 1071 Chlorobium limicola Chloroflexu aurantiacus Thiocapsa roseopersicina Halobacterium halobium Fermentative bacteria (厭氧醱酵菌) Enterobacter aerogenes E. cloacae Clostridium butyricum C. pasteurianum Desulfovibrio vulgaris Magashaera elsdenii Citrobacter intermedius
17 表2-3 各種不同菌種進行醱酵代謝之產物 (Göttschalk, 1986) 醱酵種類 菌種 產氫量及產物 乙醇 Zymomonas anaerobica Sarcina ventriculi Erwinia amylovora 少量氫氣 醋酸 乳酸 丁酸 (酯) 丁醇 丙酮 Clostridium C. acetobutylicum C. beijerinckii C. tetanomorphum C. aurantibutylicum C. kluyveri Butyrivibrio Eubacterium Fusobacterium 大量的氫氣 丁酸 (酯) 己酸 混酸 丁二醇 Escherichia coli Enterbacter aerogenes 中量的氫氣
(一)紫色不含硫光合作用細菌(purple non-sulfur bacteria)
紫色不含硫光合作用細菌(Rhodospirillaceae)主要以簡單的有機物為 電子供給者,如:乙酸(acetate)、琥珀酸(succinate)等,也可以利用氫
氣將二氧化碳還原,為光合有機營生物(photoorganotrophic),它們通常是
兼性光合營生物(facultative phototrophs)及兼性厭氧生物(facultative anae- robes)。大部分的種類均能在厭氧光照條件下生長或是在暗處而有氧的條
件下利用有機物作為碳源及能源來生長(即化學異營性)。在氧濃度0.5~5
﹪之間,光合作用和氧化性代謝(oxidative metabolism)有可能同時進行,
紫色不含硫光合作用細菌依型態不同可分為:Rhodospirillum(R.)、
Rhodobacter(Rb.)、Rhodomicrobium(Rm.)、Rhodopseudomonas(Rps.)
及Rhocyclus等(Brock & Madigan,1991)。
18 產氫能力佳,大部分的研究均指向其藉由固氮酵素(nitrogenase)的作用 來產生氫氣;而厭氧醱酵產氫微生物則是在分解複雜有機物情形下,在水 解、酸化及乙酸的情況,會伴隨著氫氣的產生;亦有部份文獻指出,在厭 氧醱酵產氫菌與厭氧光合產氫菌混合共生的系統中,其產氫效率極佳(許 淳鈞,1990)。本研究中所使用到的菌株為中興大學李季眉教授實驗室中 產氫能力最佳,菌株編號為WP3-5號菌株,其經鑑定為Rhodopseudomonas palustris。 二、厭氧甲烷化微生物 甲烷菌(methangogens)為執行甲烷化作用之專責微生物,為一類相 當特別的古細菌。甲烷生成菌為絕對厭氧微生物,所需生長環境亦較酸化 菌嚴格。目前研究中能夠被產甲烷菌利用的基質而生成甲烷的有:二氧化
碳(CO2)、甲酸(HCOOH)、一氧化碳(CO)、甲醇(CH3OH)、甲
胺(CH3NH3)、二甲胺、三甲胺、及醋酸(CH3COOH)等。而其中CO2 則幾乎能被所有產甲烷菌利用,最主要甲烷化反應是由甲烷生成菌二個族 群來進行,即嗜氫甲烷生成菌及嗜乙酸甲烷生成菌,其反應機制如下: CO2+ 4H2→ CH4(甲烷) + 2H2O………...…(2-1) (嗜氫甲烷生成菌) CH3COOH → CH4(甲烷) + CO2………..(2-2) (嗜乙酸甲烷生成菌) 其中,利用氫氣之甲烷生成菌可協助維持轉化揮發有機酸及乙醇為乙 酸根所需之低氫氣分壓條件(Speece,1983)。而經由甲烷化反應中產生 之甲烷大約有2/3之甲烷是由嗜乙酸甲烷生成菌轉化乙酸所產生,而另外 1/3則由氫還原二氧化碳所生成(Mackie & Bryant,1981)。
根據美國ATCC (American Type Culture Collection, 1989)的分類,甲烷 生成菌之種類可由其型態之特徵區分為四屬: (1)桿菌
19 (Methanobacterium) 、 (2)球菌 (Methanococcus)、 (3)八聯球菌 (Methanosarcina)以及 (4)螺旋菌 (Methanospirillum)。陳是瑩等 (1986)及陳 是瑩、曾怡禎 (1986)研究國內各種厭氧污泥,發現其中所含之甲烷菌主要 有Methanothrix、Methanosarcina、Methanococcus、Methanobacterium、 Methanospirillum等五屬,如表2-4。 甲烷生成菌之成長速率較嗜酸菌慢,所以即使是很小的溫度變化即可 能造成嚴重的影響。因此當甲烷生成菌分解揮發酸時,若降低溫度會使半 飽和常數(half saturation constant)變大而最大比生長速率變小(Lawrence & McCarty,1969)。而一般生活廢水處理廠在操作污泥厭氧消化時,其最 佳操作條件為35℃。大部分之甲烷生成菌在pH值6.7至7.4之範圍內均能保 有其功能,但最佳之pH值則介於7.0至7.2之間,若pH接近6.0時,即對甲烷 菌造成影響(Sahm,1984)。 表2-4 各種厭氧污泥中甲烷菌之種類及特性 甲烷菌種類 (屬) 特性 Methano- thrix Methano- sarcina Methano- coccus Methano- bacterium Methano- spirillum 型態 細桿菌連 接成絲狀 八聯球菌 群體 單一或 雙球菌 粗桿菌 螺旋菌 溫度 (℃) 37 35-37 - - - 形成胞子 × × × × × ORP <-200mv <-333mv pH 7.4-7.8 6.0-7.0 - - - 產氣 CH 4、CO2 CH4、CO2 - - - 對甲烷生成菌而言,當進流基質成分其C:N:P為700:5:1時,能 夠使厭氧消化有最佳的效率(Sahm,1984);而當C/N比值為25/1~30/1
20
時有最佳的產氣率(Polprasert,1989)。甲烷生成菌為絕對之厭氧菌,即 使是低濃度之氧存在也會造成極大之傷害(Oremland,1988)。當NH
3-N
濃度大於1700mg/L時,即對甲烷生成菌造成抑制作用,且隨著濃度增加, 其抑制作用增大(Koster & Lettinga,1984);而當硫化物之濃度達150~
200mg/L時,即對甲烷造成抑制作用(Rinzema & Lettinga,1988)。
第五節 厭氧微生物產能之機制
厭氧消化乃是指在厭氧條件下,藉著兼氧性及厭氧性微生物生化代謝 作用,將複雜之高分子有機物(如蛋白質、碳水化合物、脂質等)降解, 轉換成甲烷與二氧化碳等最終產物的程序。而就產氫反應機制而言,厭氧 醱酵能利用特殊微生物及酵素,在水環境中將複雜有機物水解酸化,分解 出中間產物氫氣;產甲烷反應機制則為甲烷菌利用其中間產物經甲烷化反 應而生成甲烷。本節主要分三個部分來探討厭氧微生物產能之機制:傳統 厭氧消化反應、厭氧醱酵產氫反應、厭氧甲烷化反應。 一、傳統厭氧消化反應 傳統厭氧消化反應是在無氧的環境下,首先,由兼氧性及厭氧性細菌 透過水解、產酸的反應 (即厭氧醱酵階段),將廢水中有機物 (如不溶性有 機物質、碳水化合物、脂肪或蛋白質等)轉化成揮發酸、氫氣、醇類或醛類 等中間代謝產物後;而這些中間代謝產物再轉化為甲烷及二氧化碳等最終 穩定產物,其反應機制如圖2-1 (謝哲松,1995)所示。 由圖2-1可看出,傳統厭氧消化反應最終產物為CO2與CH4,因為一者 為碳的最大氧化態;另一者為碳的最大還原態。在反應過程中,氫氣產生21 後很快被甲烷形成菌轉化成甲烷與二氧化碳,所以氫氣在傳統的厭氧反應 過程中殘留量非常少,可以當作傳統厭氧反應操作不良的重要指標 (林明 瑞,1989),但無法單獨作為系統控制的依據 (黃國豪、黃耀輝、陳致君、 梁德明,1990)。而在整個厭氧消化過程中,所得到的產物與能源(CO2與 CH4)可提供生物作為生長代謝及增殖的來源。 圖2-1 傳統厭氧消化反應過程(謝哲松譯,1995)
22 二、厭氧醱酵產氫反應 在傳統厭氧反應分解有機物的過程中,厭氧微生物群先將有機物經水 解產酸反應,將大部分有機物轉化為揮發酸、醇、醛、氫氣等中間代謝產 物,若生長條件適當,這些中間代謝產物又會被甲烷生成菌分解或轉換成 甲烷。而厭氧醱酵產氫則是設法終止後面的甲烷化反應,使厭氧反應仍持 續進行前面的水解產酸反應而有利氫氣的產生。 由上述可知,一般傳統厭氧消化反應可分為三階段,即水解、酸化、 及甲烷化三階段。而厭氧醱酵產氫反應則發生在水解及酸化階段,以下就 這二階段進行討論。 1.水解階段 此階段是利用細胞外水解 (extracellular enzymes)將複雜的有機物 (如蛋白質、脂肪、多醣類等)分解成簡單的化合物 (如胺基酸、甘油、 單醣等),而水解的效率受到pH值、水力停留時間 (HRT) (Verstraete, deBaere & Rozzi, 1981)及基質種類 (Henze & Harremöes, 1983)等因素影 響。Gujer & Zehnder (1982)指出,若進流基質為一般性有機廢水,在水 解階段不易形成速率限制,但若進流基質含有芳香族類有機化合物、纖 維素等難水解物質時,在水解階段將成為限制反應速率階段 (Kennedy & van den Berg, 1982)。
2. 產酸階段
在此階段主要是將水解後的小分子有機物轉化為氫氣、二氧化碳及 揮發性脂肪酸,包括甲酸、乙酸、丙酸、丁酸、異丁酸、戊酸、異戊酸 及己酸等 (McCarty, 1964),產物之種類依細菌種類及反應條件 (如基 質、溫度、pH、氧化還丙酸、丁酸)及醇類轉化成乙酸根、氫氣及二氧
23 化碳等可直接被甲烷生成菌利用之化合物。因高氫氣分壓時會抑制乙酸 根之生成,而使基質大部分轉化為丙酸、丁酸、乙醇,因此傳統厭氧消 化必需監控氫氣之濃度。 在厭氧生物產氫代謝機制方面,Taguchi(1992)以 Clostridium butyrium 為菌種,反應槽溫度控制在 36℃、pH 值控制在 5.5 的條件下操 作,理想的產氧效率為2.0~4.0 mole-H 2/mole-Glucose,但實際產氫效率 大約只有理想產氫效率的60%。其產氫反應方程式如下所示: C6H12O6→ 2H2+ butyrate + 2CO2……….(2-3) C6H12O6+ 2H2O → 4H2+ acetate + CO2……….(2-4) 式(2-3)的產氫效率為33%,(2-4)的產氫效率為67%,其比值為 1:2,這說明了Clostridium butyricum如果能進一步將葡萄糖分解到乙酸 階段則其產氫量約為分解到丁酸階段的2倍。 Lay et al. (1999)研究中指出,目前利用有機廢污水以連續式試驗進 行醱酵產氫其效果並不佳,原因是氫氣會被甲烷轉換及消耗。因此如能 將反應槽控制在厭氧產氫菌最佳生長條件的操作下,可大幅提昇反應槽 的厭氧產氫效率,而目前常使用的方法有: (1)pH值控制於較酸的環境。 (2)加大有機負荷或縮短水力停留時間。 (3)經常變動操作狀況,如:pH值、溫度、有機負荷、ORP值等, 以對系 統造成突增負荷(shock loading)。 (4)完全混合反應培養。 (5)儘量將系統操作在醱酵產氫菌適合生長,而甲烷菌不適合生長的環 境。 三、厭氧甲烷化反應
24 所謂厭氧甲烷化反應是指甲烷生成菌利用有機物在水解、酸化階段 所產生之乙酸、氫氣、二氧化碳及其他低碳化合物之中間產物,轉化成 甲烷及二氧化碳等氣體的過程。 甲烷生成菌主要可區分為嗜氫 (hydrogenotrophic)及嗜醋酸 (ace- totrophic)甲烷生成菌二個次族群,而嗜氫甲烷生成菌可將氫及二氧化碳 轉化成甲烷;嗜乙酸甲烷生成菌可將乙酸根轉化成甲烷及二氧化碳,其 反應機制如是2-5、2-6所述,而其甲烷作用途徑如圖2-2。 (a)從CO2開始甲烷化途徑 (b)從醋酸開始甲烷化途徑 圖2-2 主要甲烷化作用途徑(謝哲松譯,1995) CO 2 + 4H2 → CH4(甲烷) + 2H2O……….(2-5) (嗜氫甲烷生成菌) CH 3COOH → CH4(甲烷) + CO2………..(2-6)
25 (嗜乙酸甲烷生成菌)
第六節 環境因子對厭氧產能的影響
由於本實驗是以光合產氫反應與甲烷化反應進行,所以本節將分為兩 大方面來探討。 一、光合產氫反應 (一)影響光合作用細菌產氫的因子 紫色不含硫光合作用細菌並非在任何狀況下皆會產氫,是需要在光照 及缺乏氮源的情況下才會產氫,許多的環境因子(如:光照強度、溫度高 低等)均會對產氫造成影響。以下分別對這些環境因子加以探討: 1.酵素的 Cofactor 固氮酵素主要由含鐵(Fe)、鉬(Mo)元素或含鐵、硫的蛋白質所 組成(Woo,1985)。因此有研究指出對於產氫,鐵、鉬、鎂等離子是必 須的。Fascetti, D’addario, Todini and Robertiello(1998)等學者利用
Rhodobacter sphaeroides RV 在含有機酸的都市固體廢棄物中產氫反應 中發現,不加入一些生長基質和微量元素時,氫氣的產量是很低的,而 且產氫的狀況會被Mo、Fe 和 PO42-所影響,但是主要的差異是Mo 的 加入與否,因為Mo-nitrogenase 對於產氫作用,比只有 Fe-nitrogenase 存在時強。 2.光照強度 氫氣產量的多寡與光照強度有關,在光強度 540 lux 下,開始有明 顯的氫氣產生,而氫氣產量會一直隨著光照強度的增加呈正比關係,一
26
Fiβler 利用 Rhodopseudomonas palustris 分解芳香族有機物,發現光 照強度逐漸增加,總產氫量也一直增加,且固氮酵素活性也呈正比關係 逐漸增高,一直到光照強度達10000 lux 後呈現飽和,之後不管光照強 度如何增加,總產氫氣量並不會再增加(Fiβler & Schirra,1995)。 在固氮酵素的生成反應中,需要的ATP 與低能還原劑(low-potential reducing agent)是由光反應所提供。一般紫色不含硫光合作用細菌的氧 化還原電位約為-50 ~ -150 mV,而固氮酵素的 E0=-400 mV,就 Rhodopseudomonas 來說,它可以藉由光能生成-500 ~ -600 mV 來供給固 氮酵素生成,因此光就是固氮酵素生成條件之一(Meyer,Kelley & Vignais,1987)。
Sasikala, Ramana and Rao(1991)三位學者以 Rhodobacter
sphaeroides O.U.001*菌株做不同光照強度試驗,發現光照強度對菌株生 長速率影響比最後生物總質量影響大。當光照強度在3000 lux 以下,微 生物質量與光照強度成正比,但超過3000 lux 以上,微生物總質量呈定 值,也就是達到飽和生長。而光照強度對於總產氫氣量方面,光照強度 在5000 lux 左右,即已達到飽和的產氫總量,超過 5000 lux 以上,其總 產氫氣量方面並沒有太大的差異。由光照強度對菌株生長與產氫試驗可 知,即使光照強度高於最適合的條件,仍不會有抑制的情形發生。 由上述文獻中可以瞭解隨著菌株不同,產生氫氣所需要的光照強 度也略有不同,大致上光照強度至少要540 lux 以上才會有產氫的現 象,而光照強度在到達飽和光照之前,其總產氫氣量與光照強度呈正比 的關係。到達飽和之後,總產氫氣量的差異並不大,也不會有抑制的情 形發生,且利用來產氫的紫色不含硫光合作用細菌菌種不同,其最大光 照強度也就不同。
27
除了利用人為的光照之外,也有研究嘗試利用太陽光作為產氫的 光源,但需要在高強度的太陽光底下才能維持穩定產氫之情形,然而在 廢水處理若需要連續產氫,就必須使用人工光照以提供足夠光照。 Miyake and Kawamura(1987)研究發現 Rhodobacter sphaeroides 在轉
換太陽能效率方面最高可達6.2 ﹪。
3.溫度
部分紫色不含硫光合作用細菌在溫度低於20℃下時,其生長會受
到抑制,有些學者研究顯示,最佳產氫速率發生在30~ 34℃(Zürrer &
Bachhofen,1982;Ohta,Frank & Mitsui,1981)。
在溫度範圍20~ 35℃內,來探討溫度對菌株 Rhodopseudomonas
capsu- lata 的影響,根據試驗結果獲知在這個溫度範圍內,產氫總量以
及轉換成氫氣效率並沒有太大差別,但產氫速率則隨著溫度降低而減 緩,低溫時的產生氫氣時間是高溫狀態下產生氫氣時間的兩倍
(Stevens,Vertoghen,Vos & Ley, 1984)。
Singh and Srivastava(1991)二位學者將紫色不含硫光合作用細菌 TR-22 -R-B 與絕對好氧菌 22TW-S 共同培養,發現就算溫度高達 45℃, 兩菌株仍生長良好,推測可能是有好氧性微生物存在,能快速移除氧 氣,使得兼氣性光合菌生長良好。
以不同溫度培養菌株Rhodobacter sphaeroides O.U.001*,來比較在
各溫度下其產氫量及生物質量產率(biomass yield),瞭解到溫度在30~
35℃時產生最大生物質量,而高於 45℃以上或是低於 20℃,菌種幾乎
不生長。最佳產氫總量發生在30~40℃範圍內,由此可知在熱帶地區進
行產氫試驗是可行的(Sasikala et al.,1991)。
28 (二)營養需求對紫色不含硫光合作用細菌產氫之影響 1.電子供給者 紫色不含硫光合作用細菌可以利用不同的基質來產生氫氣,其氫氣 轉換率的好壞,則視基質的分子量、結構複雜與否及菌體是否容易利用 而決定。 利用固定化細胞法,以Rhodoseudomonas palustris 來測試其對不同 有機酸之產氫能力。測試的有機酸分別為acetate、lactate、pyruvate、 malate 及 succinate,結果發現利用上述有機酸產氫的轉換率分別為 40、 61、52、5 及 67﹪(Vincenzini,Materassi,Tredici & Florenzano,1982)。 其理論方程式為下列之方程式: acetate: C2H4O2 + 2H2O 4H2 + 2CO2 (2-7) lactate: C3H6O3 + 3H2O 6H2 + 3CO2 (2-8) pyruvate: C3H4O3 + 3H2O 5H2 + 3CO2 (2-9) malate: C4H6O5 + 3H2O 6H2 + 4CO2 (2-10) succinate: C4H6O4 + 4H2O 7H2 + 4CO2 (2-11) butyrate: C4H8O2 + 6H2O 10H2 + 4CO2 (2-12)
Fiβler,Schirra,Kohring and Giffhorn(1994)發現 Rhodopseudomonas
palustris 可以分解一些芳香族有機物,如:benzoate、cinnamate、DL- mandelate 及 benzoylformate 等均可以作為生長所需之電子供給者,假 使以固定化方式來處理上述芳香族有機物,其實際的產氫效率分別為 88、86、60 及 57﹪。其理論方程式如下所示: benzoate: C7H6O2 + 12H2O 15H2 + 7CO2 (2-13) cinnamate: C9H8O2 + 16H2O 20H2 + 9CO2 (2-14)
29
DL-mandelate: C8H8O3 + 13H2O 17H2 + 8CO2 (2-15) benzoylformate: C8H6O3 + 13H2O 16H2 + 8CO2 (2-16)
其利用不同濃度benzoate 來探討 Rhodopseudomonas palustris DSM
131 產氫的影響,來尋求出產氫的最佳條件。由研究結果可知,以 benzoate 濃度為 3mM 時,有最大產氣效率,然而當 benzoate 大於 16.5mM 時,則有抑制的情形發生。
於不同基質濃度對產氫之影響方面,Margaritis and Vogrinetz(1983)
等兩位學者以不同濃度之glucose 對於 Rhodobacter sphaeroides VM 81
這株菌加以探討,得到的結果是以初始glucose 濃度為 7 g/L 時,有最
大產氫量。
分別以不同濃度的malate 及 lactate 探討對 Rhodobacter sphaeroides
O.U.001*之影響,發現 malate 濃度為 30mM 時,有最大產氫量,然而 lactate 濃度為 50mM 時的產氫量最大,而且氫氣的產量隨著乳酸濃度增 加而有逐漸上升之趨勢(Sasikala et al.,1991)。
在連續流操作對濃度影響方面,Fascettic and Todini(1995)等兩
位學者以一階段或兩階段chemostat 方式培養菌株 Rhodobacter sphaeroides RV,使用 lactate 作為生長所需的基質,得到氫氣產率與 lactate 濃度有關之結果。當 lactate 濃度為 100mM 時,有最大的氫氣釋 放速率,但lactate 濃度大於 100mM 時則有抑制氫氣產生的現象。 紫色不含硫光合作用細菌因種類不同,可以利用之碳源也跟著有 所不同。故可以根據不同碳源之利用情形,對紫色不含硫光合作用細菌
30
表2-5 紫色不含硫光合作用細菌對不同電子供給者利用情形(Imhoff & Trüper,1992)
Rocy .pur pur eus Romi.vannielii Rops. acidophila Rops.capsulata Rops. gelatinosa Rops. globif or m is Rops. palustr is Rops. sphear oides Rops. sulf idop hila Rops. sulf ovir idis Rops.viridis Rosp. fulvum
Rosp.molischianu Rosp.photometric Rosp.
rubr um Rosp. tenue Acetate + + + + + - + + + + + + + + + + Arginine - - - 0 0 - - 0 - 0 0 - - - + + Aspartate - - - + 0 - - 0 ± 0 0 ± ± ± + - Beznoate + - - - - - + - - - - + - - - - Butyrate - + + + ± - + ± + + - + + + + + Caproate + + ± + - - + ± + 0 - + + - + + Caprylate - + - + 0 - + ± + 0 0 + + - 0 + Citrate - - + - + - - + - - - - - - - - Ethanol - + + - + + + + ± + + + + + + ± Formate - - ± 0 ± - + 0 + - ± 0 + - - - Fructose - - - + 0 + - + ± + 0 - - + ± - Fumarte + + + + + + + + 0 + 0 + + + + + Gluconate 0 0 0 - 0 + 0 + 0 0 0 0 0 0 - 0 Glucose - - ± + + + ± + + + + ± - + - - Glutamate - - - + 0 - + 0 + 0 + 0 0 - + 0 Glycerol - - ± - - - + + 0 + - - - + - - Glycolate - - ± 0 0 - + 0 0 0 0 0 - + 0 - Lacate - + + + + - + + + + - - ± + + + Malate + + + + + + + + + + + + + + + + Malonate - + ± 0 0 - + 0 0 - 0 0 - 0 0 - Mannitol - - - - ± + - + - - 0 - - + - - Methanol - ± ± 0 ± - - 0 - - 0 0 - - ± ± Pelargonate - - - + 0 - - ± + 0 0 + + ± 0 + Propionate - + + + ± - + ± + - - + + ± + + Pyruvate + + + + + ± + + + - + + + + + + Succinate - + + + + + + + + + + + + + + + Tartrate - - ± - ± + - + - - 0 0 - - - - Valerate - + + + 0 - + ± + - - + + + + + Yeast axtract - + + + + + + + + 0 0 + + + + + Casamino Acids - + + + 0 - + 0 + 0 + - - + + + Hydrogen.molecular + + + + ± 0 + + + 0 - + 0 0 + +
31
2.氮源
大部分的文獻指出,氮源對光合作用細菌的產氫能力具有關鍵性 的影響,而紫色不含硫光合作用細菌則是在氮源限制的情況下會產生氫 氣。Woo, Lee,Kwon and Kho(1985)等學者發現 Rhodopseudomonas 在
有NH4+的情況下,不論在光照或暗處均有氫氣的產生。另外Oremerod
et al.(1961)也指出,在有 N2、胺基酸或低濃度的氨氮下也會有氫氣 的產生,但是過高的氨氮、硝酸鹽、尿素、glutamine 及 asparagine 下, 則會對氫氣的產生造成抑制。若缺少氮源,則會使紫色不含硫菌光合作
用細菌無法合成固氮酵素,在接種後的12~16 小時後,固氮酵素的活性
逐漸降低,使得產氫的總時間縮短。Miyake, Tomizuka and Kamibayashi (1982)等學者發現 Rhodospirillum rubrum 添加少量硫酸銨及氮氣可以
延長產氫的時間至5~14 天。
Hillmer and Gest(1977)以 Rhdopseudomonas capsulata 為菌株,
glutamate 作為氮源,有相當高的產氫速率;如果換成以 NH4+或是N2
作為氮源,則出現抑制氫氣產生的現象
Oremerod et al.(1961)發現以紫色不含硫光合作用細菌
Rhodopseudo- monas rubrum 的批次實驗中,氨為單一氮源時,必須要
在氨用完之後的resting cells 狀態下,氫氣才會開始產生,其推測光合 產氫機制需要氨合成蛋白質時所產生的成分(即固氮酵素)。且前培養 使用glutamate 為氮源會比用氨為氮源的產氫量來的高。試驗中提及在 氨過量、不產生氫氣的狀態下,只要將紫色不含硫光合作用細菌 Rhodopseudomonas rubrum 移到無機鹽與光照條件,以蘋果酸為碳源的 條件下,約一小時後就開始產生氫氣,三到四小時後開始有一個線性增 加,若在開始產氫之後加入氯化氨會使產氫終止。
32 Fiβler(1994)以各種不同胺基酸作為菌株 Rhodopseudomonas palustris 生長時所需之氮源,如:L-glutamate、L-alanine、L-aspartate、 L-leucine 等。由菌株的生長狀況及產氫的情形可以瞭解,以 NH4Cl 作 為氮源時,菌株的生長相當快速,但其產氫量不高,如果菌株利用 L-glutamate 來生長時,其產氫速率及生長速率均很快。 (三)其它因子對紫色不含硫光合作用細菌產氫之影響 紫色不含硫光合作用細菌在環境中屬於弱勢的競爭族群,在自然的環 境中有藻類、藍綠細菌硫酸鹽還原菌與甲烷生成菌存在時會影響其生長, 所以光合作用細菌產氣需要在未污染的乾淨環境下來進行。藉著控制藻 類、藍綠細菌、硫酸鹽還原菌的生長,使得紫色不含硫光合作用細菌生長 不受抑制。
表2-6 在各種氣體成分條件下 Rhodobacter sphaeroides O.U.001*其產氫轉
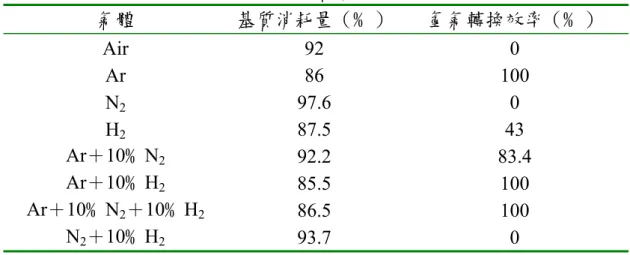
換率及基質消耗量 氣體 基質消耗量(﹪) 氫氣轉換效率(﹪) Air 92 0 Ar 86 100 N2 97.6 0 H2 87.5 43 Ar+10﹪N2 92.2 83.4 Ar+10﹪H2 85.5 100 Ar+10﹪N2+10﹪H2 86.5 100 N2+10﹪H2 93.7 0 在不同氣體狀態下,菌株產氫亦有所影響,Sasikala,Ramana,Rao and Subrahmanyam(1990)以 8 種狀態下之氣體對菌株 Rhodobacter sphaeroides O.U.001*進行測試,測試結果如表 2-8 所示,當氣體狀態以 Ar、Ar+10﹪
33 ﹪,然而菌株卻無法在以Air 或是以 N2為氣態條件下產氫,可能是酵素受 氧氣或是氮氣所抑制。從表2-8 中也可以發現,以 Ar 作為產氫時的氣體狀 態有最佳的結果,不但基質的去除率高而且基質轉換成氫氣轉換率良好。 二、甲烷化反應 (一) pH 厭氧消化反應中,主要參與反應的菌群均有其最適生長的pH值。酸 形成菌的最適pH值為5.0~6.0之間,最適合酸化反應進行。而甲烷生成菌 僅能在pH值維持中性時具有高產甲烷率,大部分的甲烷生成菌最適pH 值在7.0-7.2之間,當pH值低於6.3或大於7.8時,甲烷生成速率可能降低; 厭氧消化系統在不控制pH值時,生物系統內產生揮發性有機酸將會自然 增加反應槽內的酸度[見方程式(2-17)及(2-18)]。當反應槽內揮 發酸產生量超過甲烷菌形成之消耗量時,即會有酸度之累積。 CH3COOH ↔ CH3COO - + H+ (2-17)
H2O + CO2(g) ↔ H2CO3(aq) ↔HCO3-(aq)+ H+(aq) (2-18)
而在進行傳統厭氧消化時,大多建議pH值應控制在中性範圍。因為 低pH值及過量揮發酸累積對甲烷菌抑制將大於醱酵菌。 (二) 溫度 溫度是影響微生物生長與存活最重要的因子之一。在厭氧廢水生物 處理程序中,厭氧反應速率隨溫度增加而增加,適合生長的溫度可達60℃ 左右。因此溫度在厭氧消化反應中,是一個相當重要的控制因子,厭氧 消化的溫度一般可分為中溫消化區(30~35℃)及高溫消化區(50~55℃),