行政院國家科學委員會專題研究計畫 成果報告
胰島素抗阻、糖尿病與肥胖相關基因之研究(3/3)
計畫類別: 個別型計畫 計畫編號: NSC93-3112-B-002-005- 執行期間: 93 年 05 月 01 日至 94 年 04 月 30 日 執行單位: 國立臺灣大學醫學院內科 計畫主持人: 莊立民 共同主持人: 陳垣崇 計畫參與人員: 葉日弌;、林文星 報告類型: 完整報告 處理方式: 本計畫可公開查詢中 華 民 國 94 年 10 月 28 日
行政院國家科學委員會補助專題研究計畫
□ 成 果 報 告
□期中進度報告
(計畫名稱)
胰島素抗阻、糖尿病與肥胖相關基因之研究
Study on the genes related to insulin resistance, type 2 diabetes and obesity
計畫類別:□
個別型計畫 □
整合型計畫
計畫編號
:NSC 93-3112-B-002-005
執行期間:93 年 5 月 1 日至 94 年 4 月 30 日
計畫主持人:
莊立民
共同主持人:
計畫參與人員:陳垣崇、葉日弌、林文星
成果報告類型(依經費核定清單規定繳交):□精簡報告 □完整報告
本成果報告包括以下應繳交之附件:
□赴國外出差或研習心得報告一份
□赴大陸地區出差或研習心得報告一份
□出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份
處理方式:除產學合作研究計畫、提升產業技術及人才培育研究計畫、
列管計畫及下列情形者外,得立即公開查詢
□涉及專利或其他智慧財產權,□一年 v 二年後可公開查詢
執行單位:
台大醫學院、內科
中 華 民 國 94 年 10 月 27 日
中文摘要
胰島素阻抗常伴隨肥胖、第二型糖尿病、高血壓、脂質代謝異常、冠狀動脈疾病等 臨床表現,臨床上統稱為代謝症候群,是現今全世界健康課題上最常見之重要的慢性 病;吾等提出三年之計劃,旨在剖析胰島素阻抗之致病機轉,並採多種不同之策略來 達成目標。首先我們會採用分子遺傳學之研究,來了解基因之單一核甘酸變異與臨床 表現型之關聯,而候選基因主要是來自我們實驗室自己找出在脂肪細胞與前脂肪細胞 有差異表現之基因,這些基因極可能與肥胖之發生有關,並與胰島素之敏感度有關; 吾等將尋找這些基因之單一核甘酸變異並研究在正常人、肥胖、第二型糖尿病、冠狀 動脈疾病病人之間的差異。而了解這些基因之重要性之後,與這些基因位在同一個路 徑之基因(如位在同一個胰島素信息傳遞路徑)都可能為新的候選基因,我們預期將 可完成數個分子遺傳學之研究報告。 吾等過去之研究發現有些基因之變異與肥胖、第二型糖尿病有關聯,為要證實這個 發現,如能做出此基因之剔除小鼠來研究其表現型,將是最重要之證明;本計劃擬要 剔除之基因為 SH3P12/SORBS1 與本計劃命名為 X 之基因。 前者在別人與吾等之研 究中發現與胰島素促進細胞對葡萄糖之攝取作用有關,且此一基因之變異與國人之肥 胖和第二型糖尿病有關;而後者則尚未在人類找出相對應之基因,然而從這個基因之 表現來判斷是極為有趣之基因,它在脂肪細胞分化時會被誘發表現出來,它在肥胖與 糖尿病鼠之脂肪組織表現量也比正常對照鼠為高,它在不同之組織表現也具特異性, 更重要的是以胰島素增敏劑處理細胞時,它的表現會被抑制下來,暗示它與胰島素阻 抗有極密切之關聯。 經過修正著重目標在: 尋找候補基因之 SNPs,並確定其與胰島素阻抗性及相關臨床第二型糖尿病、肥胖及 冠狀動脈疾病之關係。 我們在臨床結合基因學之研究,顯示與 APM1 基因的 SNP 與胰島素阻抗性有關連, 並與 PPARγ基因型產生交互作用,影響肥胖與胰島素阻抗性(APM1 與肥胖關係之研究,已被JMM 接受刊登,而 APM1 與 PPARγ基因之交互作用,也已被 Diabetologia
接受刊登)。 更重要的是,我們研究 APM1 基因之對偶基因之表達,和 SORBS1 基因 在脂肪組織的表達,與肥胖或胰島素阻抗性有密切之關連(前者在 JMM 中刊登,而後 者則在Ob Res 刊登)。 經由本年的研究發現有更多的有興趣的基因標記,並與臨床複雜的疾病有關,如代謝 症候群。此經驗可提供未來大規模的基因型分析,得出更多的數據資料,以達到對糖 尿病與代謝症候群更深入之瞭解。
英文摘要
Insulin resistance associates with multiple clinical manifestations such as obesity, type 2 diabetes, hypertension, dyslipidemia and coronary artery disease (CAD), collectively termed metabolic syndrome, and is a major health issue in our society and in the whole world. In this limited 3-years project, we propose to dissect pathogenic pathways leading to the major defect underlying the metabolic syndrome, i.e. insulin resistance. We applied several approaches to accomplish the goals. The first is molecular genetic studies of the association between candidate SNPs and clinical phenotypes including insulin resistance, obesity and type 2 diabetes. The candidate genes that we choose to study are mostly identified from a differential display between adipocytes vs. preadipocytes in our own laboratory. Based on our large collection of the samples, we can analyze and correlate the genetic variants with clinical phenotypes in the control, obese, type 2 diabetes and CAD subjects. Once confirmed, the other molecules involved in the same pathway might serve as other candidates for testing. We expected to accomplish several molecular genetic studies for publication in 3 years.
Since we have characterized some of the candidate genes previously, we will confirm the biological function of the gene(s) that are shown to associated with the diseases. For this purpose, the best way is to knockout the genes and to see if phenotypes occur. Others and we have shown the role of SH3P12/SORBS1 in the insulin-stimulated glucose uptake. In addition, we have found SNP of this gene correlated with obesity and type 2 diabetes in our population. We will create knockout mice of the SH3P12/SORBS1 gene and establish an animal model for studying insulin resistance, obesity, and diabetes mellitus. Another new gene designated X in this proposal is also very interesting in that the expression of this gene is increased in the adipose tissues of heterozygous and homozygous db/db mice as compared to the wild type mice. Moreover, an insulin sensitizer down regulated the expression of this gene in the 3T3-L1 adipocytes, suggesting that this gene might involve insulin sensitivity. Sequence study indicated this gene was a secretory protein that contained a signal peptide. It is therefore speculated that overproduction will cause insulin resistance and the related phenotypes such as obesity and type 2 diabetes.
Isolation of the SNPs of the candidate genes and confirm the association with insulin resistance and the related clinical disorders such as type 2 diabetes, obesity and coronary artery disease in Chinese population. We have confirmed by clinical association studies to show a positive correlation of SNP of the APM1 gene and its interaction with PPARγ gene in subjects with obesity and insulin resistance. More importantly, we showed the allele-specific expression of APM1 gene and expression of SORBS1 gene in the adipose tissue to correlate clinical obesity and insulin sensitivity.
Currently, we have identified more interesting genetic markers that might be related to clinical complex disorder such as the metabolic syndrome. Future large-scale genotyping might yield more remarkable data via high throughput genome center.
Background.
Type 2 diabetes is a multifactorial disorder involving both genetic and environmental factors (7). Detailed studies reveal two interacting basic defects are established in pathophysiology for human type 2 diabetes, i.e., βcell dysfunction (8,9), and insulin resistance (10-12). Understanding the mechanisms of insulin resistance and βcell defect is the key to elucidating the pathogenesis of type 2 diabetes and obesity. However, the genetic susceptibility factors for this disorder remains to be identified, due to a limited success in gene finding for the multifactorial diseases in general (14,15).
From our and others’ observations, obesity is the major determinant for the development of type 2 diabetes (13). In addition to the environmental contribution to the development of obesity, obesity is now can also be considered as a genetic disease of adipose tissue (16). The molecular mechanisms of programming of adipocytes differentiation have been extensively studied and theses studies also provide a basic understanding of obesity in animals and humans (for review, see 17). It is therefore possible to isolate the genes involved in syndrome of obesity and diabetes from the understanding of adipocyte gene expression (18). Interestingly, the induction of adipocyte differentiation is associated with an increase in insulin sensitivity (19).
Aims of the study.
1. To isolate and characterize the genes related to insulin resistance, obesity and type 2 diabetes. 2. Study of the functions of the candidate genes relating to insulin resistance
Study design and methods.
1. Differential display, subtraction cloning, and microarray to isolate the candidate genes that involve insulin signaling/insulin action and adipocytes differentiation.
In the first year, we focused on 3 genes isolated from adipogenesis: i.e. PPARγ, APM1, SORBS1.
Designs and Methods
Subjects. Two independent samples, one of normal non-diabetic and normotensive individuals serving as control and the other of type 2 diabetes mellitus as cases.
Genotyping. (i) Genomic DNA. Total genomic DNA from peripheral blood leukocytes of each individual was extracted using Puregene DNA extraction kit, in accordance with the provider’s instruction (Minneapolis, MN). For the genotyping analysis which need more high quality DNA , the DNA concentration of each sample will be measured more exactly by fluorometric quantitation method using PicoGreen dsDNA Quantitation Reagent (Molecular Probes, Eugene, Oregon) and gel electrophoresis. The genomic DNA was stored at –20 degree C until SNP analysis wsa performed.
(ii) SNP analysis.
a) Candidate genes selection: In the previous study, we isolated 56 candidate genes for diabetes/obesity development in mouse adipogenesis system. Two genes such as Sorbs1/Sh3d5/Cap and Acrp30 had been demonstrated which play some role in the pathogenesis of human disorder with insulin resistance. After human homology searching, we
identified 45 genes for further SNP allele frequencies determination.
b) SNPs selection: In principle, we will select 4 SNPs from public database such as dbSNP in NCBI (http://www.ncbi.nlm.nih.gov/SNP/), JSNPs in Japanese JSNP Database (http://snp.ims.u-tokyo.ac.jp/) and SNPper database (http://bio.chip.org:8080/bio/) for each gene to perform genotyping. Each SNP will been identified the validation and heterozygosity with 96 case and control subjects first then additional subjects genotyping been performed as its real and has more than 10 % polymorphism. The sequence surrounding a SNP site will be obtained from above mentioned SNP databases. PCR primers for evaluating polymorphic sites will be designed by employing SpectroDESIGNER software (MassARRAY, Sequenom). We will try to perform three or four SNPs variant analysis in one genotyping analysis. In this purpose, we will use SpectroDESIGNER to design optimal multiple primer sets for SNPping reaction.
c) MALDI-TOF Mass Spectrometry: After PCR reaction, the terminal phosphate of non-incorporated dNTPs will be removed from the amplification products by shrimp alkaline phosphatase (SAP) treatment. Homogeneous MassExtend (hME) reaction can detect polymorphisms in amplified DNA by specifically extending a specific primer annealing near the SNP site. Then, the analyte DNA of 1 µl will be added to the 384-spectroCHIP by using SpectroPOINT and dried at room temperature. The DNA on the matrix spot will be introduced into the source region of a mass spectrometer (Sequenom). SNP genotyping from a 384-well target will be performed automatically.
Results and Discussion.
a. Selected candidate genes and clinical metabolic diseases: We have demonstrated 3 major genes isolated from differential display, i.e. the APM1 gene, the SORBS1 gene and PPARγ, that showed significant association of metabolic phenotypes in humans. In the 3 papers that we published, we did show significant SNPs of the three genes either alone or interact with each others affect insulin sensitivity, glucose/insulin metabolisms, and clinical diseases such as obesity and type 2 diabetes (See publications #1~3).
b. Mechanism of SNP and clinical disorders. We identified certain genetic interaction of the T/G polymorphism of the APM1 gene with PPARγ Pro12Ala polymorphism in determination of insulin sensitivity, serving as a good model of gene-gene interaction for such a complex disorder or a complex trait. Furthermore, in correlation of the tissue expressions of the genes with clinical phenotypes, we found that tissue mRNA expressions of the APM1 and SORBS1 genes are indeed correlated with clinical phenotypes (See publications #1,3). More interestingly, we found a specific allelic expression of the silent SNP of APM1 gene might be the underlying molecular mechanism that leads to clinical phenotypes (See publication #1). More recently, we found a significant correlation of APM1 genetic polymorphism with coronary artery disease (manuscript 1 in submission). Due to our clinical human association of adiponectin gene, APM1, our manuscript on reviewing the genetic association of APM1 gene has been accepted as a review article in the Journal of Molecular Medicine (See publication #4).
c. Two additional candidate genes, GLUT10 and APOA5 that contribute to clinical phenotypes. We have tested for the contribution of GLUT10 and APOA5 to clinical phenotypes such as type 2 diabetes and hypertriglyceridemia. We found that certain uncommon SNP haplotypes of the GLUT10 gene were associated with type 2 diabetes although with a modest significance level (manuscript 2 in submission). On the other hand, the SNP of the APOA5 gene, which interacts with fasting plasma glucose, was associated with hypertriglyceridemia, indicating another example of complex disease caused by gene-environment interaction (See publication #5).
d. Large-scale SNP genotyping for the genes involved in adipogenesis in human type 2 diabetes. We demonstrated the feasibility of finding genes from the differentially expressed genes during a pathway of adipogenesis, which is involved in pathogenesis of clinical disorders such as insulin resistance, obesity and type 2 diabetes. For now, we have chosen 4 SNP sites for each of the genes isolated from differential display that resulted in 106 SNP which were informative and successfully genotyped. In the first set of 380 normal and 380 diabetic samples, total genotyping of 60,040, we found 7 SNPs that were significantly correlated with type 2 diabetes. For replication, we genotype another set of 380 normal and 380 diabetic samples and found two of the SNPs that were significant associated with type 2 diabetes. The following summarizes the processes that we conducted for this association study:
• Overall success rate for the SNP genotyping : 97.02% for the control samples; 90.78% for the diabetic samples
• Testing allelic and genotypic distribution difference (association) between control and patient groups;
• Testing HWE for all SNPs in the control group (http://www.biostat-resources.com/stata/); • Significant level determined by Monte Carlo simulation (with CLUMP from
http://www.gene.ucl.ac.uk/~dcurtis). SNPs with p<0.05 after correction for multiple
testing are labeled significant.
• The final two SNPs that were significant in association with type 2 diabetes are now under more SNP typing and for further haplotype analyses.
Some statistical results are shown in the following:
1. Firstly, we can analyzed the different SNP/gene on the quantitative traits, such as fasting plasma glucose, HbA1c, HOMA-IR or –beta, and triglyceride levels (Table 1~4) in the normal non-diabetic and normotensive individuals. These examples illustrate a multigenetic nature for the complex disorder.
2. There are good examples to show a diverse effect of one single gene on metabolic phenotypes, genetic pleiotropism- a common feature of complex disorder (Table 5,6).
3. Among the two genes that had associated with type 2 diabetes, we further used STRUCTURE and STRAT software to adjust for population structure in association study.
HMGA2 remained highly significant after adjustment for population structure. Interestingly, this encoded protein contains structural DNA-binding domains and may act as a transcriptional regulating factor. Identification of the deletion, amplification, and rearrangement of this gene that are associated with myxoid liposarcoma suggests a role in adipogenesis and mesenchymal differentiation. A gene knock out study of the mouse counterpart demonstrated that this gene is involved in diet-induced obesity, making this finding highly sensible. The other GM2A, though showed significant association with insulin resistance with HOMA as shown in the table 3 and association with type 2 diabetes, the explanation of its functional significance remains to be further explored (manuscript 3 in preparation).
Table 1. The SNPs that are associated with fasting plasma glucose in the normal individuals with quantitative analyses. The SNP numbering (the first two rows), gene names, and p-values are shown.
s48 6150 rs3205421 CHPT1 0.0013 s28 6115 rs340141 TRIP10 0.0113 s39 6133 rs2256923 IGFALS 0.0255 s50 6153 rs2695281 CHPT1 0.029 s94 7546 rs2303873 SORT1 0.0427 s48 6150 rs3205421 CHPT1 0.0006 s39 6133 rs2256923 IGFALS 0.02 s3 6065 rs3740297 GDF2 0.0331 s28 6115 rs340141 TRIP10 0.0362 s50 6153 rs2695281 CHPT1 0.0479
Table 2. The SNPs that are associated with HbA1c in the normal individuals with quantitative analyses. The SNP numbering (the first two rows), gene names, and p-values are shown.
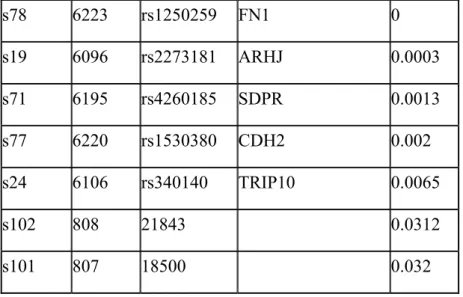
s78 6223 rs1250259 FN1 0 s19 6096 rs2273181 ARHJ 0.0003 s71 6195 rs4260185 SDPR 0.0013 s77 6220 rs1530380 CDH2 0.002 s24 6106 rs340140 TRIP10 0.0065 s102 808 21843 0.0312 s101 807 18500 0.032
Table 3. The SNPs that are associated with HOMA-IR or HOMA-beta in the normal individuals with quantitative analyses. The SNP numbering (the first two rows), gene names, and p-values are shown.
s58 6167 rs1048719 GM2A 0.0458
s58 6167 rs1048719 GM2A 0.0355
s66 6184 rs153477 GM2A 0.0259
s66 6184 rs153477 GM2A 0.0284
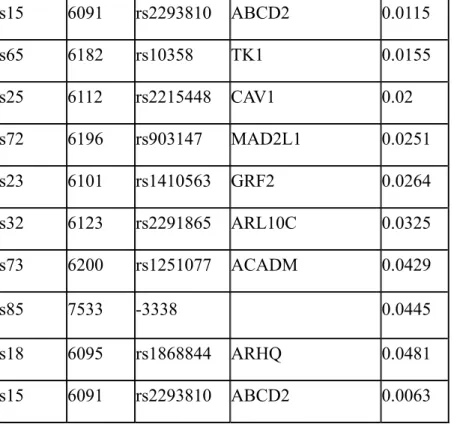
Table 4. The SNPs that are associated with triglyceride levels in the normal individuals with quantitative analyses. The SNP numbering (the first two rows), gene names, and p-values are shown. s15 6091 rs2293810 ABCD2 0.0115 s65 6182 rs10358 TK1 0.0155 s25 6112 rs2215448 CAV1 0.02 s72 6196 rs903147 MAD2L1 0.0251 s23 6101 rs1410563 GRF2 0.0264 s32 6123 rs2291865 ARL10C 0.0325 s73 6200 rs1251077 ACADM 0.0429 s85 7533 -3338 0.0445 s18 6095 rs1868844 ARHQ 0.0481 s15 6091 rs2293810 ABCD2 0.0063
Table 5. Correlation of the SNPs of caveolin 1 with various metabolic phenotypes. ht s26 6113 rs6867 CAV1 0.0103 ht_r s26 6113 rs6867 CAV1 0.007 ltg s25 6112 rs2215448 CAV1 0.02 ltg_r s25 6112 rs2215448 CAV1 0.039 ua s25 6112 rs2215448 CAV1 0.0143 ua_r s25 6112 rs2215448 CAV1 0.0068 Ht: height; tg: triglyceride, ua: uric acid; _r: data transformed
Table 6. Correlation of the SNPs of CCRN4L with various metabolic phenotypes.
bmi s8 6081 rs2292839 CCRN4L 0.0427
pc s11 6085 rs2271777 CCRN4L 0.0296
ua s8 6081 rs2292839 CCRN4L 0.0379
Papers published:
1. Yang WS,Tsou PL, Lee WJ, Tseng DL, Chen CL, Peng CC, Lee KC, Chen MJ, Huang CJ, Tai TY, Chuang LM* (2003) Allele-specific differential expression of a common adiponectin gene polymorphism related to obesity. J Mol Med 81:428-434
2. Yang WS, Hsiung CA, Ho LT, Chen YT, He CT, Curb JD, Grove J, Quertermous T,Chen YDI, Kuo SS, Chuang LM* for the SAPPHIRe Study Group (2003) Genetic Epistasis of Adiponectin and PPARγ2 Genotypes in Modulation of Insulin Sensitivity: a Family-based Association Study. Diabetologia 46:977-983.
3. Yang WS, Lee WJ, Huang KC, Lee KC, Chao CL, Chen CL, Tai TY, Chuang LM* (2003) mRNA Levels of the insulin-signaling molecule SORBS1 in the adipose depots of nondiabetic women. Ob Res 11:586-90.
4. Yang WS, Chuang LM* (2005). Human Genetics of Adiponectin in the Metabolic Syndrome. J Mol Med (in press)
5. Jiang YD, Yen CJ, Chou WL, Kuo SS, Lee KC, Chiu KC, Chuang LM* (2005) Interaction of the G182C polymorphism in the APOA5 gene and fasting plasma glucose on plasma triglycerides in Type 2 diabetic subjects. Diabetic Medicine (in press)
Manuscript in submission/preparation:
1. JY Jiang,YD Jiang, FT Chiang,JJ Hwang, WP Lien, LM Chuang*. Interaction of the Genetic Variant of the APM1/ACDC Gene and serum cholesterol level in Angiography-Proven
Coronary Artery Disease. (submitted)
2. WH Lin, LM Chuang (equal to first author), CH Chen, JI Yeh, PS Hsieh, CH Cheng, YT Chen. The genetic variation of human SLC2A10 is associated with type 2 diabetes mellitus in Taiwanese population. (submitted)
3. Yeh JI, Lin WS, Chen YT, Chuang LM*. SNP-based association study of the genes involved in adipogenesis in human type 2 diabetes. (in preparation)
Self evaluation. Based on the first year’s experience, we demonstrated the feasibility of finding genes from the differentially expressed genes during a pathway, which is involved in pathogenesis of clinical disorders for a complex disorder. This would provide us a ground base for a plan in the next two years for a larger set of the genes that are isolated during adipogenesis for better understanding of the pathogenetic mechanism of insulin resistance and the related clinical disorders. In addition, we have many original papers and one review paper accepted for publications in the prestigious journals.