REGULAR ARTICLE
Heat shock-mediated H
2O
2accumulation and protection
against Cd toxicity in rice seedlings
Yi Ting Hsu&Ching Huei KaoReceived: 28 May 2007 / Accepted: 13 August 2007 / Published online: 5 September 2007
# Springer Science + Business Media B.V. 2007
Abstract Rice (Oryza sativa L.) seedlings stressed with CdCl2 (0.5 mM or 50 μM) showed typical Cd toxicity (leaf chlorosis, decrease in chlorophyll content, or increase in H2O2 and malondialdehyde contents). Rice seedlings pretreated with heat shock at 45°C (HS) for 2 or 3 h were protected against subsequent Cd stress. Rice seedlings pretreated with HS had similar Cd concentration in leaves caused by CdCl2 as those non-HS. The content of H2O2 in-creased in leaves 1 h after HS exposure. However, APX and GR activities were higher in HS-treated leaves than their respective control, and it occurred after 2 h of HS treatment. Pretreatment of rice seed-lings with H2O2under non-HS conditions resulted in an increase in APX, GR, and CAT activities and protected rice seedlings from subsequent Cd stress. HS-induced H2O2production and protection against subsequent Cd stress can be counteracted by im-idazole, an inhibitor of NADPH oxidase complex. Results of the present study suggest that early ac-cumulation of H2O2during HS signals the increase in APX and GR activities, which in turn prevents rice seedlings from Cd-caused oxidative damage. Keywords Cadmium . Heat shock .
Hydrogen peroxide . Oryza sativa . Oxidative stress
Abbreviations
APX Ascorbate peroxidase ASC Ascorbate CAT Catalase DAB 3, 3-Diaminobenzidine DW Dry weight GR Glutathione reductase HS Heat shock IMD Imidazole MDA Malondialdehyde ROS Reactive oxygen species SOD Superoxide dismutase
Introduction
Cadmium (Cd), a heavy metal toxic to humans, animals and plants, is a widespread pollutant. In Cd-sensitive plants, it has been shown that (a) Cd can promote the generation of reactive oxygen species (ROS); (b) Cd can inhibit or stimulate the activities of antioxidant en-zymes; and (c) treatment with Cd results in cellular oxidative damage or lipid peroxidation (Chaoui et al. 1997; Chien and Kao2000; Dixit et al.2001; Gallego et al. 1996; Kuo and Kao 2004; Olmos et al. 2003; Piqueras et al.1999; Romero-Puertas et al.2003; Shah et al. 2001; Shaw 1995). These results indicate that oxidative stress is a major component of Cd stress.
Exposure of plants in the range of 15 min to a few hours at temperature 5 to 15°C above the normal growing temperature is usually considered as heat Responsible Editor: Juan Barcelo.
Y. T. Hsu
:
C. H. Kao (*)Department of Agronomy, National Taiwan University, Taipei, Taiwan, Republic of China
shock (HS) treatment. It has been shown that leaf segments exhibited an acquired protection against Cd and other heavy metal toxicity following exposure of the seedlings to HS in the dark (Orzech and Burke 1988). Neumann et al. (1994) demonstrated that a HS treatment preceding Cd stress induced a tolerance effect by preventing the membrane damage. Prior HS exposure of rice seedlings was also observed to in-hibit subsequent Cd-induced ethylene production in detached leaves (Chen and Kao1995a).
In addition to being an endogenous oxidant, H2O2 has also been implicated as a diffusible signal for selective induction of defense mechanisms in plant cells (Chen et al.1993; Prasad et al.1994a,b). H2O2 treatment has been shown to enhance chilling toler-ance in maize (Prasad et al. 1994a, b) and in mung bean and Phalaenopsis (Yu et al.2002;2003). Nodal potato explants subcultured from H2O2-treated mi-croplant were resistant to normally lethal exposure of 42°C (Lopez-Delgado et al. 1998). Recent work also demonstrated that H2O2 pretreatment resulted in an improvement of salt tolerance (Azevedo Neto et al. 2005; Uchida et al. 2002; Wahid et al. 2007). In mustard seedlings, HS at 45°C caused a significant increase in endogenous H2O2 content and improved their tolerance to subsequent heat stress at 55°C (Dat et al. 1998). Gong et al. (2001) reported that a HS pretreatment could simultaneously induce cross adap-tation of maize seedlings to chilling, heat, drought and salt stresses. They also showed that HS-induced H2O2 accumulation was involved in signaling and trigger-ing of this cross adaptation.
It is not known if H2O2treatment induces Cd stress tolerance in rice seedlings. Neither do we know if HS pretreatment results in an accumulation of H2O2 in rice seedlings. In the present study, we showed that a HS pretreatment could induce Cd stress tolerance in rice seedlings and investigated the possible involve-ment of H2O2in Cd stress tolerance.
Materials and methods Plant material and treatments
Rice (Oryza sativa L., cv. TN1) seeds were sterilized with 2.5% sodium hypochlorite for 15 min and washed extensively with distilled water. These seeds were then germinated in Petri dishes with wetted filter
papers at 37°C in the dark. After 48 h incubation, uniformly germinated seeds were selected and culti-vated in a 250 ml beaker containing half-strength Kimura B solution containing the following macro-and micro-elements: 182.3μM (NH4)2SO4, 91.6μM KNO3, 273.9 μM MgSO4·7H2O, 91.1 μM KH2PO4, 182.5 μM Ca(NO3)2, 30.6 μM Fe–citrate, 0.25 μM H3BO3, 0.2μM MnSO4·H2O, 0.2 μM ZnSO4·7H2O, 0.05 μM CuSO4·5H2O, and 0.07 μM H2MoO4. Kimura B solution contains the desired nutrient ele-ments and has been widely used for growing rice plants. Since young rice seedlings were used for the present study, the nutrient solution contains no silicon, although silicon is essential for rice. The nutrient solutions (pH 4.7) were replaced every 3 days. The hydroponically cultivated seedlings were grown in a Phytotron (Agricultural Experimental Station, National Taiwan University, Taipei, Taiwan) with natural sun-light at 30/25°C day/night and 90% relative humidity. HS pretreatment and Cd stress treatment
Twelve-day-old seedlings with three leaves were ex-posed to 30°C (non-HS) and 45°C (HS) for 3 h in the dark. Non-HS and HS seedlings were then grown in basic nutrient solution with or without CdCl2(0.5 mM or 50μM) at 30/25°C day/night.
Measurement of chlorophyll, H2O2, malondialdehyde, and Cd
Chlorophyll content was determined according to Wintermans and De Mots (1965) after extraction in 96% (v/v) ethanol. H2O2was visually detected in the leaves by using 3,3-diaminobenzidine (DAB) as sub-strate (Orozco-Cárdenas and Ryan 1999). Detached rice leaves were supplied through the cut ends with DAB (1 mg ml−1) solution for 24 h under light at 27°C. Leaves were then decolorized in boiling etha-nol (95%) for 0.5 h. This treatment decolorized the leaves except for the brown polymerization product produced by DAB with H2O2. After cooling, the leaves were extracted at room temperature with fresh ethanol to visualize the brown spots. To verify the specificity of precipitates, before staining with DAB some leaves were immersed for 2 h in solution con-taining the H2O2 scavenger 1 mM ascorbate (ASC) suggested by Romero-Puertas et al. (2004). It was observed that the development of DAB–H2O2
reac-tion product in detached rice leaves could be pre-vented by ASC (data not shown), which demonstrated the specificity of the reaction of DAB with H2O2.The H2O2 staining was repeated four times with similar results. The H2O2content was also measured color-imetrically as described by Jana and Choudhuri (1982). H2O2 was extracted by homogenizing leaf tissue with phosphate buffer (50 mM, pH 6.5) containing 1 mM hydroxylamine. The homogenate was centrifuged at 6,000×g for 25 min. To determine H2O2content, the extracted solution was mixed with 0.1% titanium chloride in 20% (v/v) H2SO4. The mixture was then centrifuged at 6,000×g for 25 min. The absorbance was measured at 410 nm. Using this method, we obtained that absorbance increased linearly with the amount of H2O2 and addition of H2O2to extracts resulted in the predicted increase of absorbance, i.e. added H2O2was fully recovered (data not shown), was calculated using the extinction coefficient of 0.28 μmol−1 cm−1. Malondialdehyde (MDA), routinely used as an indicator of lipid perox-idation, was extracted with 5% (w/v) trichloroacetic acid and determined by the thioburbituric acid reac-tion as described by Heath and Packer (1968). For determination of Cd, leaves were dried at 65°C for 48 h. Dried material was ashed at 550°C for 20 h. The ash residue was incubated with 31% HNO3 and 17.5% H2O2 at 72°C for 2 h, and dissolved in distilled water. Cd concentration was then quantified using an atomic absorption spectrophotometer (Model AA-6800, Shimadzu, Kyoto, Japan). Amount of Cd is expressed on the basis of dry weight (DW).
Enzyme extraction and assays
For extraction of enzymes, leaf tissues were ho-mogenized with 0.1 M sodium phosphate buffer (pH 6.8) in a chilled pestle and mortar. For analysis of ascorbate peroxidase (APX) activity, 2 mM ASC was added to the extraction buffer. The homogenate was centrifuged at 12,000×g for 20 min and the resulting supernatant was used for determination of enzyme activity and protein content. The whole extraction procedure was carried out at 4°C. Super-oxide dismutase (SOD) was determined according to Paoletti et al. (1986). One unit of SOD was defined as the amount of enzyme that inhibits by 50% the rate of NADH oxidation observed in blank sample. Catalase (CAT) activity was assayed according to Kato and
Shimizu (1987). One unit of CAT was defined as the amount of enzyme which degraded 1μmol H2O2per minute. APX activity was determined according to Nakano and Asda (1981). One unit of activity for APX was defined as the amount of enzyme that de-graded 1 μmol of ASC per minute. Glutathione re-ductase (GR) was determined by the method of Foster and Hess (1980). One unit of GR was defined as the amount of enzyme that decreased 1 A340per minute. The enzyme extracts were used for determination of protein by the method of Bradford (1976).
Statistical analysis
Statistical differences between measurements (n=4) on different treatments or on different times were analyzed following LSD test.
Results
Effect of prior HS exposure on the tolerance of rice seedling to Cd
In plants, the most general symptom of Cd toxicity is leaf chlorosis (Das et al. 1997). When rice seedlings were treated with 0.5 mM CdCl2, chlorosis was first shown in the second leaves, but not the third leaves of rice seedlings in a short-term experiment (3 days; Hsu and Kao2003). Thus, in the present study, Cd toxicity was assessed by the chlorosis and the decrease in chlorophyll content of the second leaves.
To test if prior exposure of rice seedlings to HS would affect subsequent Cd-induced toxicity of the second leaves, seedlings were pretreated with HS for 1, 2, and 3 h, respectively, under dark conditions. Rice seedlings pretreated with 1 h HS resulted in no protective effect on subsequent Cd-induced leaf chloro-sis (Fig. 1a) and Cd-decreased chlorophyll content (Fig. 1b). However, a 2-h HS pretreatment exhibited a slight but significant reduction of Cd-induced leaf chlorosis (Fig. 1a) and Cd-decreased chlorophyll content (Fig.1b). Three hours of HS exposure showed a complete inhibition of Cd-induced toxicity in rice seedlings (Fig.1a,b).
In previous work, it has been demonstrated that Cd can induce oxidative stress in rice leaves, characterized by an increase in the contents of H2O2and MDA (an indicator of lipid peroxidation; Hsu and Kao 2004;
Kuo and Kao2004). In the present study, we observed that the increase in the contents of H2O2(Fig.2a) and MDA (Fig. 2b) caused by CdCl2 was more pro-nounced in non-HS leaves than in HS leaves.
The concentration of CdCl2used in the aforemen-tioned study was 0.5 mM. We also conducted experi-ments with lower CdCl2 (50 μM) applied over a longer period (6 days). Rice seedlings pretreated with
3 and 6 h HS was observed to exhibit protective effect on subsequent Cd-induced chlorosis (Fig. 3). Thus, the effects of HS on the response to lower CdCl2 concentration are basically in accordance with those to higher CdCl2concentration.
Effect of HS pretreatment on Cd concentration in leaves of rice seedlings
Figure4shows that rice seedlings pretreated with 3 h HS had similar Cd concentration in the second leaves caused by CdCl2 as those non-HS. These results suggest that the protective effect of HS on subsequent Cd-induced toxicity of the leaves is unlikely due the inhibition of Cd uptake or transport.
Chlorophy ll (mg g -1 FW) 0 1 2 3 a a a a a b bc c Pretreatment HS (h) 0 0 1 2 3 Treatment CdCl2 (0.5 mM, 2 d) – + + + + Pretreatment HS (h) 0 1 2 3 Treatment CdCl2 (0.5 mM, 2 d) – + – + – + – +
a
b
Fig. 1 Effects of CdCl2 (0.5 mM) on chlorosis (a) and chlorophyll content (b) in the second leaves of rice seedlings pretreated with HS (45°C) for 1, 2, and 3 h, respectively, under dark conditions. Arrow indicates the second leaves. Bars indicate standard error (n=4). Values with the same letter are not significantly different at P<0.05
a
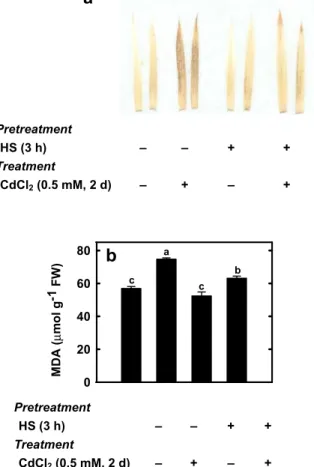
Pretreatment HS (3 h) – – + + Treatment CdCl2 (0.5 mM, 2 d) – + – + MDA ( µ mol g -1 FW) 0 20 40 60 80 a b c cb
Pretreatment HS (3 h) – – + + Treatment CdCl2 (0.5 mM, 2 d) – + – + Fig. 2 Effects of CdCl2(0.5 mM) on H2O2(a) and MDA (b) contents in the second leaves of rice seedlings pretreated with or without HS (45°C) for 3 h under dark conditions. The H2O2 content was visually detected in the second leaves by using DAB as substrate after 2 days of CdCl2treatment. To visualize DAB–H2O2 reaction product, leaves were decolorized in boiling ethanol. Bars indicate standard error (n=4). Values with the same letter are not significantly different at P<0.05H2O2content and antioxidant enzyme activities in response to HS
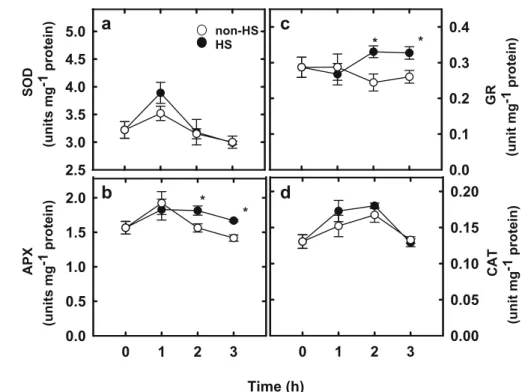
Changes in H2O2 content and antioxidant enzymes (SOD, APX, GR, and CAT) after exposure rice seedlings to HS were evaluated. As shown in Fig.5, H2O2 content increased in leaves 1 h after HS
ex-posure and subsequently remained unchanged. There was no difference in SOD and CAT activities between HS- and non-HS leaves (Fig. 6a,d). However, APX and GR activities in leaves of HS seedlings were higher than their respective non-HS, and it occurred after 2 h of HS treatment (Fig.6b,c).
The effect of NADPH oxidase inhibitor
To test if NADPH oxidase is responsible for H2O2 production during HS, imidazole (IMD), an inhibitor of NADPH oxidase inhibitor, was added to half-strength Kimura B solution at the time when the third leaves was fully expanded. Results in Fig.7show that HS-induced H2O2 production and protection against subsequent Cd stress can be counteracted by IMD (0.1 mM).
Effect of H2O2pretreatment under non-HS condition on the tolerance of rice seedlings to Cd
To test if HS-increased H2O2 content is involved in the protection of rice seedlings against Cd toxicity, rice seedlings were first pretreated with H2O2(in the range of 0.1–1 mM) for 3 h under non-HS conditions and then transferred to nutrient solution with or with-out CdCl2. It was observed that pretreatment of rice
Cd (
µ
g g
-1
DW)
0
2
4
6
8
- - + + - + - + Pretreatment HS (3 h) Treatment CdCl2 (0.5 mM, 2 d) a a b bFig. 4 Effect of CdCl2(0.5 mM) on Cd concentration in the second leaves of rice seedlings pretreated with or without HS (45°C) for 3 h under dark conditions. Cd was determined 2 days after CdCl2treatment. Bars indicate standard error (n=4). Values with the same letter are not significantly different at P<005
H 2 O 2 ( µ mol g -1 FW) 0 30 40 50 60
a
a
a
a
b
b
b
Time (h) 0 1 2 3 non-HS HSFig. 5 Changes in H2O2content in the second leaves of rice seedlings pretreated with or without HS (45°C) under dark conditions. Seedlings were exposed to HS for 1, 2, and 3 h and the second leaves were taken for determination of H2O2 contents. Bars indicate standard error (n=4). Values with the same letter are not significantly different at P<0.05
Pretreatment
HS (h) 0 0 3 3 6 6
Treatment
CdCl2 (50 µµM, 6 d) – + – + – + Fig. 3 Effect of lower CdCl2 concentration (50 μM) on chlorosis in the second leaves of rice seedlings pretreated with HS (45°C) for 3 and 6 h, respectively, under dark conditions
seedlings with H2O2 greatly improved tolerance of rice seedlings to Cd (Fig.8a,b). Figure9a shows that addition of H2O2(0.5 mM) to the basic nutrient so-lution of rice seedlings for 3 h under non-HS con-ditions resulted in an increase in H2O2in the second leaves. Furthermore, rice seedlings pretreated with H2O2under non-HS conditions had no effect on SOD activity (Fig. 9e) but showed an enhancement in APX, GR, and CAT activities in the second leaves (Fig.9b–d).
Discussion
Cd causes chlorosis or chlorophyll loss (Hsu and Kao 2003) and oxidative stress (Kuo and Kao2004) in rice seedlings. In the present study, we evaluated Cd tox-icity by the chlorosis, decrease in chlorophyll content, and increase in H2O2and MDA contents. On the basis of these criteria, we demonstrated that HS pretreatment of rice seedlings was able to protect rice seedlings from subsequent Cd stress [high CdCl2concentration (0.5 mM for 2 days) or low concentration (50μM) for 6 days]. The protective effect of HS against subsequent Cd toxicity has also been described previously (Chen and Kao 1995a, b; Neumann et al.1994; Orzech and Burke1988).
It has been shown that a reduction of Cd-induced ethylene production was less effective in detached leaves from rice seedlings pretreated with HS for 24 h as compared with those for 6 and 12 h (Chen and Kao 1995a). In the present study, we also observed that the protective effect on Cd-induced chlorosis was more pronounced in a 3-h HS pretreatment than a 6-h HS (Fig. 3). All these results are basically in accordance with the idea that only a short duration of HS is re-quired for seedlings to exhibit an acre-quired protection against Cd and other heavy metal toxicity (Gong et al. 2001; Orzech and Burke 1988).
The protective effect of HS against subsequent Cd toxicity of rice seedlings is unlikely due to inhibition of Cd uptake or transport. Because rice seedlings pre-treated with 3 h HS had similar Cd concentration in leaves cause by CdCl2as those non-HS (Fig.4).
Doke (1997) showed that early peak in H2O2was observed within 15 min after HS of cell suspensions of potato leaf tissues. Exposure of whole tobacco seedlings to 40°C for 1 h in the light induced a sig-nificant increase in H2O2(Foyer et al.1997). A peak in H2O2 content in mustard seedlings was observed within 5 min transfer to the 45°C acclimation tem-perature (Dat et al. 1998). More recently, Gong et al. (2001) reported that the HS (42°C) pretreatment for 4 h produced an endogenous H2O2 peak in maize
SOD (units mg -1 protein) 2.5 3.0 3.5 4.0 4.5 5.0 Time (h) 0 1 2 3 APX (units mg -1 protein) 0.0 0.5 1.0 1.5 2.0 GR (unit mg -1 protein) 0.0 0.1 0.2 0.3 0.4 0 1 2 3 CAT (unit mg -1 protein) 0.00 0.05 0.10 0.15 0.20 * * * * non-HS HS
a
c
b
d
Fig. 6 Changes in the activities of SOD (a), APX (b), GR (c), and CAT (d) in the second leaves of rice seedlings pretreated with or without HS (45°C) under dark conditions. Rice seed-lings were exposed to HS for 1, 2, and 3 h and the second leaves were taken for determination of each enzyme activity. Bars indicate standard error (n=4). Values with the same letter are not significantly different at P<0.05
seedlings. Here, we show that HS pretreatment of rice seedlings resulted in an increase in H2O2 in 1 h (Fig.5).
There is limited information about the mechanism of HS-induced H2O2 production. HS-induced accu-mulation of H2O2 in mustard seedlings has been suggested to be due to HS-deactivated CAT activity (Dat et al. 1998). This does not seem to be the case in rice seedlings, because HS had no effect on CAT activity (Fig. 6d). In several mode systems investi-gated in plants, the accumulation of H2O2appears to be mediated by the activation of a plasma-membrane-bound NADPH oxidase complex (Orozco-Cárdenas 1999; Pei et al. 2000; Tsai et al. 2004; Zhang et al. 2001). It is not known whether HS-induced H2O2
accumulation in rice seedlings is mediated through NADPH oxidase complex. The fact that HS-induced H2O2 production in leaves of rice seedlings can be inhibited by IMD (Fig. 7a) suggests that HS-dependent H2O2generation in rice leaves originated, at least in part, from plasma membrane NADPH oxidase. The involvement of NADPH oxidase in HS-induced H2O2 production in Arabidopsis has also been described recently (Volkov et al.2006).
Chlorophy ll (mg g -1 FW) 0 1 2 3 4 5 - - + + - + - + Pretreatment IMD (0.1 mM, 3 h) Treatment CdCl2(0.5 mM, 2 d) a b c c H2 O2 ( µ mol g -1 FW) 0 5 30 35 40 45 50 55 60 - +
a
Pretreatmnet IMD (0.1 mM, 3 h) a bb
Fig. 7 Effect of IMD (0.1 mM) on HS-induced H2O2 production (a) and CdCl2-incuced chlorophyll loss (b) in the second leaves of rice seedlings. Rice seedlings were pretreated with 3 h HS under dark conditions and then treated with or without 0.5 mM CdCl2for 2 days. IMD was added to half-strength Kimura B solution during 3 h HS. Bars indicate standard error (n=4). Values with the same letter are not significantly different at P<0.05 0 1 2 3 Chlorophy ll (mg g -1 FW) ab a b c b b
a
Pretreatment H2O2 (mM, 3 h) 0 0 0.1 0.3 0.5 1.0 Treatment CdCl2 (0.5 mM, 2 d) – + + + + + Pretreatment H2O2 (0.5 mM, 3 h) – – + Treatment CdCl2 (0.5 mM, 2 d) – + +b
Fig. 8 Effects of CdCl2(0.5 mM) on chlorophyll content (a) and chlorosis (b) in the second leaves of rice seedlings pretreated with H2O2for 3 h under non-HS (30°C) and dark conditions. Seedlings were pretreated with various H2O2 concentrations (0.1, 0.3, 0.5, and 1 mM). Arrow indicates the second leaves. Bars indicate standard error (n=4). Values with the same letter are not significantly different at P<0.05
Gong et al. (2001) proposed that H2O2 is in-volved in HS-induced cross adaptation to heat, chil-ling, drought, and salt stress in maize seedlings. The present study also indicated that H2O2was involved in HS-induced protection against subsequent Cd stress of rice seedlings. This conclusion was based on the observations that (a) endogenous H2O2 was higher in leaves of rice seedlings during HS than non-HS (Fig. 5), (b) HS-induced H2O2 production and protection against subsequent Cd stress can be counteracted by IMD, an inhibitor of NAPDH oxidase (Fig. 7a,b), and (c) pretreatment of rice seedlings with exogenous H2O2under non-HS con-ditions, which increased endogenous H2O2 content, greatly improved tolerance of rice seedlings to Cd (Figs.8 and9a).
It has been shown that various abiotic stresses induce oxidative stress and improvement of stress tolerance is often related to increase in activities of antioxidant enzymes (Alscher et al.1997; Noctror and Foyer1998). Heat protection of wheat is improved by keeping APX and GR activities high (Kraus and Fleccher 1994), and GR activity is significantly enhanced during heat-induced thermoprotection in mustard seedlings (Dat et al.1998). Exposure of alfalfa to supra-optimal tem-perature leads to increases in APX and CAT (Badiani et al. 1997). Evidence was also provided to show that HS induction of APXa gene could be the possible cause of reduced chilling injury in rice seedlings (Sato et al. 2001). Here, we report that HS pretreatment in rice seedlings resulted in higher APX and GR activities than non-HS (Fig.6b,c).
H2 O2 ( µ mol g -1 FW) 0 5 35 40 45 50 a b APX (units mg -1 protein) 0.0 0.5 1.0 1.5 2.0 a b GR (unit mg -1 protein) 0.00 0.05 0.10 0.15 0.20 0.25 0.30 a b - + Pretreatment H2O2 (0.5 mM, 3 h)
a
b
c
0.0 0.1 0.2 0.3 a b SOD (units m g -1 p rotein ) 0.0 0.5 1.0 1.5 2.0 2.5 a a - +d
e
CAT (unit m g -1 p rotein ) - + Pretreatment H2O2 (0.5 mM) Fig. 9 Effects of H2O2 on the content of endogenous H2O2(a) and the activities of APX (b), GR (c), CAT (d), and SOD (e) in the second leaves of rice seed-lings. Seedlings pretreated with 0.5 mM H2O2for 3 h under non-HS (30°C) and dark conditions. Bars indi-cate standard error (n=4). Values with the same letter are not significantly different at P<0.05Because H2O2 is relatively stable and diffusable through membranes (in contrast with superoxide), it may be a perfect candidate to act as a signal molecule during stress responses (Van Breusegem et al.2001). In fact, H2O2is now considered as a signal molecule that induces gene expression of antioxidant enzymes related to stress tolerance (Foyer et al. 1997; Morita et al.1999; Prasad et al.1994a,b).
Elevated levels of SOD, CAT, and APX have been shown to be correlated with the development of HS-induced chilling tolerance (Kang and Saltveit 2001). In previous work, we demonstrated that Cd can induce oxidative stress in rice leaves (Hsu and Kao2004; Kuo and Kao 2004). Cho and Seo (2005) reported that seedlings of Cd-resistant Arabidopsis had higher activ-ities of SOD, APX, and GR and experienced lower oxidative stress from Cd exposure.
The time-course analyses of HS in rice seedlings clearly indicated that H2O2 accumulation occurs first and then APX and GR activities increase (Figs. 5 and 6). Exogenously supplied H2O2 to rice seedlings under non-HS conditions also increased APX and GR activities (Fig. 9b,c) and protected against subsequent Cd stress (Fig. 8). All these results have led us to conclude that early accumulation of H2O2 during HS signals the increase in APX and GR activities, which in turn prevents rice seedlings from the oxidative damage caused by Cd.
In the present study, we show that HS pretreatment had no effect on CAT activity in rice leaves (Fig.6d). However, exogenous application of H2O2under non-HS conditions enhanced CAT activity (Fig. 9d). The discrepancy in the regulation of CAT activity of rice seedlings in response to HS and exogenous H2O2 under non-HS conditions is more probable due to the difference in the amount of H2O2 in leaves (Figs. 5 and 9a).
Glutathione (GSH) is an important compound of the antioxidant system, which scavenge ROS under oxidative conditions. One mechanism of increasing tolerance to Cd or other heavy metals may be the for-mation of GSH. The protective role of GSH on chilling stress and in reducing Cd or Ni toxicity has been reported (Chen and Kao 1995b; Freeman et al.2004; Kocsy et al.2000; Xiang et al.2001). Nieto-Sotelo and Ho (1986) demonstrated that maize roots under HS conditions had high GSH content than those under non-HS. Furthermore, H2O2 treatment induced GSH accumulation has also been described previously
(Yu et al. 2002; 2003). An alternative mechanism to explain the HS-acquired Cd tolerance may be related to HS-produced heat shock proteins (Kochhar and Kochhar 2005; Neumann et al. 1994; Susuki et al. 2001; Volkov et al.2006). It is important to note that the participation of H2O2in HS-induced expression of heat shock proteins has been demonstrated (Volkov et al.2006). Whether HS-induced Cd tolerance of rice seedlings is due to H2O2-mediated GSH biosynthesis and/or expression of heat shock proteins remains to be established.
Acknowledgements This work was supported by a research grant the National Science Council of the Republic of China (NSC 95-2313-B-002-046).
References
Alscher RG, Donahue JL, Cramer CK (1997) Reactive oxygen species and antioxidants: relationships in green cells. Physiol Plant 100:224–233
Azevedo Neto AD, Prisco JT, Enéas-Filho J, Medeiros J-VR, Gomes-Filho E (2005) Hydrogen peroxide pre-treatment induces salt-stress acclimation in maize plants. J Plant Physiol 162:114–1122
Badiani M, Paolacci AR, Fusari A, D’Ovidio R, Scandalios JG, Porceddu E, Sermanni G (1997) Non-optimal growth temperatures and antioxidants in the leaves of Sorghum bicolor (L.) Moench. II. Short-term acclimation. J Plant Physiol 151:409–421
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72:248–254
Chaoui A, Mazhoudi S, Ghorbal MH, Ferjani EE (1997) Cadmium and zinc induction of lipid peroxidation and effects on antioxidant enzyme activities in bean (Phaseolus vulgaris L.). Plant Sci 127:139–147
Chen SL, Kao CH (1995a) Prior temperature exposure affects subsequent Cd-induced ethylene production in rice leaves. Plant Sci 104:135–138
Chen SL, Kao CH (1995b) Glutathione reduces the inhibition of rice seedling root growth caused by cadmium. Plant Growth Regul 16:249–252
Chen Z, Silva H, Klessig RF (1993) Active oxygen species in the induction of plant systemic acquired resistance by salicylic acid. Science 262:1883–1886
Chien HF, Kao CH (2000) Accumulation of ammonium in rice leaves in response to excess cadmium. Plant Sci 156:111– 115
Cho U-H, Seo N-H (2005) Oxidative stress in Arabidopsis thaliana exposed to cadmium is due to hydrogen peroxide accumulation. Plant Sci 168:113–120
Das P, Samantaray S, Rout GR (1997) Studies on cadmium toxicity in plants: a review. Environ Pollut 98:2229–2236
Dat JF, Foyer CH, Scott IM (1998) Changes in salicylic acid and antioxidants during induced thermotolerance in mustard seedlings. Plant Physiol 118:1455–1461
Dixit V, Pandey V, Shyam R (2001) Differential antioxidative responses to cadmium in roots and leaves of pea (Pisum sativum L. cv. Azad). J Exp Bot 52:1101–1109
Doke N (1997) The oxidative burst: roles in signal transduction and plant stress. In: Scandalios J (ed) Oxidative stress and the molecular biology of antioxidant defenses. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, pp 785–813
Foster JG, Hess JL (1980) Response of superoxide dismutase and glutathione reductase activities in cotton leaf tissue exposed to an atmosphere enriched in oxygen. Plant Physiol 66:482–487
Foyer CH, Lopez-Delgado H, Dat JF, Scott IM (1997) Hydrogen peroxide- and glutathione-associate mechanism of acclamatory stress tolerance and signaling. Physiol Plant 100:241–254
Freeman JL, Persan MW, Nieman K, Albrecht C, Peer W, Pickering IJ, Salt DE (2004) Increased glutathione biosyn-thesis plays a role in nickel tolerance in Thalspi nickel hyperaccumulators. Plant Cell 16:2129–2176
Gallego SM, Benavides MP, Tomaro ML (1996) Effect of heavy metal ion excess on sunflower leaves: evidence for involvement of oxidative stress. Plant Sci 121:151–159 Gong M, Chen B, Li X-G, Guo L-H (2001)
Heat-shock-induced cross adaptation to heat, chilling, drought and salt stress in maize seedlings and involvement of H2O2. J Plant Physiol 158:1125–1130
Heath RL, Packer L (1968) Photoperoxidation in isolated chloroplasts I. Kinetics and stoichiometry of fatty acid peroxidation. Arch Biochem Biophys 125:189–198 Hsu YT, Kao CH (2003) Role of abscisic acid in cadmium
tolerance of rice (Oryza sativa L.) seedlings. Plant Cell Environ 26:867–874
Hsu YT, Kao CH (2004) Cadmium toxicity is reduced by nitric oxide in rice leaves. Plant Growth Regul 42:227–238 Jana S, Choudhuri MH (1982) Glycolate metabolism of these
submerged aquatic angiosperms during aging. Aquat Bot 12:345–354
Kang H-M, Saltveit ME (2001) Activity of enzyme antioxidant defense systems in chilled and heat shocked cucumber seedling radicles. Physiol Plant 1123:548556
Kato M, Shimizu S (1987) Chlorophyll metabolism in higher plants VII. Chlorophyll degradation in senescing tobacco leaves: phenolic-dependent peroxidative degradation. Can J Bot 65:729–735
Kochhar S, Kochhar VK (2005) Expression of antioxidant enzymes and heat shock proteins in relation to combined stress of cadmium and heat in Vigna mungo seedldings. Plant Sci 168:921–929
Kocsy G, Von Ballmoss P, Suter M, Rüegseggger A, Galli U, Szalai G, Galiba G, Brunold C (2000) Inhibition of glutathione synthesis reduces chilling tolerance in maize. Planta 211:528–536
Kraus TE, Fleccher RA (1994) Paclobutrazol protects wheat seedlings from heat and paraquat injury: is detoxification of active oxgen involved? Plant Cell Physiol 35:45–52 Kuo MC, Kao CH (2004) Antioxidant enzyme activities are
upregulated in response to cadmium in sensitive, but not in
tolerant rice (Oryza sativa L.) seedlings. Bot Bull Acad Sin 45:291–299
Lopez-Delgado H, Dat JF, Foyer Ch, Scott IM (1998) Induction of thermotolerance in potato microplants by acetylsalicylic acid and H2O2. J Exp Bot 49:713–720
Morita S, Kaminaka H, Masumura T, Tanaka C (1999) Induction of rice cytosolic ascorbte peroxidase mRNA by oxidative stress: involvement of hydrogen peroxide in oxidative stress signaling. Plant Cell Physiol 40:417– 422
Nakano Y, Asda K (1981) Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol 22:807–880
Neumann D, Lichtenberger O, Günther D, Tschiersch K, Nover L (1994) Heat-shock proteins induce heavy-metal tolerance in higher plants. Planta 194:360–367
Nieto-Sotelo J, Ho T-HD (1986) Effect of heat shock on the metabolism of glutathione in maize roots. Plant Physiol 82:1031–1035
Noctror G, Foyer CH (1998) Ascorbate and glutathione: keeping active oxygen under control. Annu Rev Plant Physiol Plant Mol Biol 49:249–279
Olmos EO, Martínez-Solano JR, Piqueras A, Hellín E (2003) Early steps in the oxidative burst induced by cadmium in cultured tobacco cells (BY-2 line). J Exp Bot 54:291–301 Orozco-Cárdenas ML, Ryan CA (1999) Hydrogen peroxide is generated systemically in plant leaves by wounding and systemin via the octadecanoid pathway. Proc Natl Acad Sci USA 96:6553–6557
Orzech KA, Burke JJ (1988) Heat shock and the protection against metal toxicity in wheat leaves. Plant Cell Environ 11:711–714
Paoletti F, Aldinucci D, Mocali A, Capparini A (1986) A sensitive spectrophotometric method for the determination of superoxide dismutase activity in tissue extracts. Anal Biochem 154:536–541
Pei X, Murata Y, Benning G, Thomine S, Klusener B, Allen G, Grill E, Schroeder J (2000) Calcium channels activated by hydrogen peroxide mediate abscisic acid signaling in guard cells. Nature 406:731–734
Piqueras A, Olmos E, Martínez-Solano JR, Hellín E (1999) Cd induced oxidative burst in tobacco BY-2 cell: time-course, subcellular location and antioxidant response. Free Radic Res 31:S25–S31
Prasad TK, Anderson MD, Martin BA, Stewart CR (1994a) Evidence for chilling-induced oxidative stress in maize seedlings and a regulatory role for hydrogen peroxide. Plant Cell 6:65–74
Prasad TK, Anderson MD, Stewart CR (1994b) Acclimation, hydrogen peroxide, and abscisic acid protect mitochondria against irreversible chilling injury in maize seedlings. Plant Physiol 105:619–627
Romero-Puertas MC, Zablza A, Rodriguez-Serrano M, Gómez M, del Río LA, Sandalio LM (2003) Antioxidative response to cadmium in pea roots. Free Radic Res 37:44 Romero-Puertas MC, Rodríguez-Serrano M, Corpas FJ, Gómez
M, del Río LA, Sandalio LM (2004) Cadmium-induced subcellular accumultion of O2 and H2O2in pea leaves. Plant Cell Environ 27:1122–1134
Sato Y, Murakami T, Funatsuki H, Matsube S, Saruyama H, Tanida M (2001) Heat shock-mediated APX gene expression
and protection against chilling injury in rice seedlings. J Exp Bot 52:145–151
Shah K, Kumar RG, Verma S, Dubey RS (2001) Effect of cadmium on lipid peroxidation, superoxide anion generation and activities of antioxidant enzymes in growing rice seedlings. Plant Sci 161:1135–1144
Shaw BP (1995) Effect of mercury and cadmium on the activities of antioxidative enzymes in the seedlings of Phaseolus aureus. Biol Plant 37:587–596
Susuki N, Koizumi N, Sano H (2001) Screening of cadmium-responsive genes in Arabidopsis thaliana. Plant Cell Environ 24:1177–1188
Tsai Y-C, Hong C-Y, Liu L-F, Kao CH (2004) Relative importance of Na+and Cl−in NaCl-induced antioxidant sytems in roots of rice seedlings. Physiol Plant 122:86– 94
Uchida A, Jagendorf AT, Hibino T, Takabe T, Takabe T (2002) Effects of hydrogen peroxide and nitric oxide on both salt and heat stress tolerance in rice. Plant Sci 163:5115–5523 Van Breusegem F, Vranová E, Dat JF, Inzé (2001) The role of active oxygen species in plant signal transduction. Plant Sci 161:405–414
Volkov RA, Panchuk II, Mullineaux PM, Schöffl F (2006) Heat stress-induced H2O2is required for effective expression of
heat shock genes in Arabidopsis. Plant Mol Biol 61:733– 746
Wahid A, Perveen M, Gelani S, Basra SMA (2007) Pretreatment of seed with H2O2improves salt tolerance of wheat seed-lings by alleviation of oxidative damage and expression of stress proteins. J Plant Physiol 164:83–294
Wintermans JFGM, De Mots A (1965) Spectrophotometric characteristics of chlorophyll a and b and their pheophytins in ethanol. Biochim Biophys Acta 109:448–453
Xiang C, Werner BL, Christensen EM, Oliver DJ (2001) The biological functions of glutathione revisited in Arabidopsis transgenic plants with altered glutathione levels. Plant Physiol 126:564–574
Yu C-W, Murphy TM, Sung W-W, Lin C-H (2002) H2O2 treatment induced glutathione accumulation and chilling tolerance in mung bean. Funct Plant Biol 29:1081–1087 Yu C-W, Murphy TM, Lin C-H (2003) Hydrogen
peroxide-induced chilling tolerance in mung beans mediated through ABA-independent glutathione accumulation. Funct Plant Biol 30:955–963
Zhang X, Zhang L, Dong F, Gao J, Galbraith DW, Song C-P (2001) Hydrogen peroxide is involved in abscisic acid-induced stomatal closure in Vicia faba. Plant Physiol 126:1438–1448