Direct visualization of triplex DNA molecular dynamics by fluorescence resonance
energy transfer and atomic force microscopy measurements
Chia-Ching Chang, Po-Yen Lin, Yen-Fu Chen, Chia-Seng Chang, and Lou-Sing Kan
Citation: Applied Physics Letters 91, 203901 (2007); doi: 10.1063/1.2809406
View online: http://dx.doi.org/10.1063/1.2809406
View Table of Contents: http://scitation.aip.org/content/aip/journal/apl/91/20?ver=pdfcov Published by the AIP Publishing
Articles you may be interested in
Self-assembly of 50 bp poly(dA)·poly(dT) DNA on highly oriented pyrolytic graphite via atomic force microscopy observation and molecular dynamics simulation
J. Chem. Phys. 139, 085102 (2013); 10.1063/1.4818595
Fluorescence resonance energy transfer-based molecular logic circuit using a DNA scaffold Appl. Phys. Lett. 101, 233703 (2012); 10.1063/1.4769812
Fluorescent resonant excitation energy transfer in linear polyenes J. Chem. Phys. 132, 124109 (2010); 10.1063/1.3367896
Electronic couplings and on-site energies for hole transfer in DNA: Systematic quantum mechanical/molecular dynamic study
J. Chem. Phys. 128, 115101 (2008); 10.1063/1.2841421
Using fluorescence resonance energy transfer to measure distances along individual DNA molecules: Corrections due to nonideal transfer
J. Chem. Phys. 122, 061103 (2005); 10.1063/1.1854120
This article is copyrighted as indicated in the article. Reuse of AIP content is subject to the terms at: http://scitation.aip.org/termsconditions. Downloaded to IP: 140.113.38.11 On: Wed, 30 Apr 2014 07:00:45
Direct visualization of triplex DNA molecular dynamics by fluorescence
resonance energy transfer and atomic force microscopy measurements
Chia-Ching Changa兲,b兲
Department of Biological Sciences and Technology, National Chiao Tung University, Hsinchu 30050, Taiwan and National Nano Device Laboratories, Hsinchu 30078, Taiwan
Po-Yen Lin,c兲 Yen-Fu Chen, and Chia-Seng Changa兲,d兲
Institute of Physics, Academia Sinica, Taipei 11529, Taiwan
Lou-Sing Kana兲,e兲
Institute of Chemistry, Academia Sinica, Taipei 11529, Taiwan
共Received 2 August 2007; accepted 22 October 2007; published online 13 November 2007兲 We have detected the dynamics of 17-mer DNA triplex dissociation mechanism at the molecular level. Fluorescence resonance energy transfer 共FRET兲 was used as an indicator of intermolecular interaction in nanometer range, whereas atomic force microscopy共AFM兲 was employed to address single molecule with sub-angstrom precision. The maximum rupture force of DNA triplex was found at pH 4.65, consistent with macroscopic observations. These results indicated that the FRET together with an AFM detection system could be used to reveal the DNA triplex interaction in nanometer scale unambiguously. © 2007 American Institute of Physics.关DOI:10.1063/1.2809406兴
DNA triplexes have attracted considerable attention due to their possible roles played in the control of gene expression.1,2 Gene expression can be regulated via an anti-gene strategy through the formation of DNA triplexes.1,2In this paper, we demonstrated that the reaction mechanism of DNA triplex formation and rupture could be studied by way of direct imaging methods of fluorescence resonance energy transfer3 共FRET兲 and atomic force microscopy 共AFM兲, at single molecule4,5levels and in variant pH conditions.
In order to directly monitor the DNA rupture event, a designed FRET pair of fluorophore关6-carboxyfluorescein 共6-FAM, F兲兴 and dark quencher 关Dabcyl, 共DQ兲兴 was labeled on the designed DNA oligomers at 3
⬘
-end 共synthesized by TriLink, Inc. at San Diego, CA兲 indicated as follows.Oligo1: 5
⬘
-共biotin兲 共T兲10TTCTTCTGATTCTCTCCTTTGGAGAGAATCAGAAGAGAA共DQ兲−3
⬘
. A17-nucleotidyl sequence 共the italic letters above兲 was chosen from the gene of CDC 25 of Pnumernia carinii.6 Incorporat-ing this sequence in our experiments, a hairpin duplex was designed by a corresponding complementary strand 共or W strand, denoted by boldface letters兲 with a TTT double strand linker as shown above. Thus, oligo1 can fold back using the TTT as a loop and form an intramolecular DNA duplex com-posed of 17 base pairs. Although a GA pair introduced in the sequence might disturb the stability a little, it could be sta-bilized by its two long flanking arms composed of 7- and 8-pyrimidine motifs on either side when forming a DNA triplex. During the exploration period, we have checked its circular dichroism spectrum 共data not shown兲 which
indi-cated positively that the ability of triplex formation far ex-ceeded the disrupting effect of inserting a GA pair. A single stranded decathymidylic motif关共T兲10兴 with biotin is attached
at the 5
⬘
-end to serve as a spacer with an anchor to the AFM probe surface. Thus, the hairpin DNA duplex will protrude 共for about 7 nm兲 from the surface of the AFM probe. A DQ moiety was attached to the 3⬘
-end to serve as a quencher when coupling with F in oligo2共see below兲.Oligo2: 5
⬘
-共biotin兲 共T兲10CCTCTCTTAGTCTTCTT共F兲− 3
⬘
. Oligo2 contains a 5⬘
-biotin-共T兲10spacer/anchor moiety and a 17-mer with a F at the 3⬘
-end. The sequence of the 17-mer, CCTCTCTTAGTCTTCTT, was designed to form the Hoogsteen hydrogen bonds with the W strand of oligo1 under triplex formation buffer solution.2Single molecular images of DNA triplex were observed in a solution environment with the MPF-3D 共Asylum Re-search, Santa Barbara, CA兲 in tapping mode. The triplex for-mation can be visualized by AFM, as shown in Fig.1. The lump resembles the structure of triplex that is outlined at the upper left corner according to the study of Hansma et al.7It is known that nonspecific interactions between the probe and the coverslip affect the force curve during the process of probe withdrawal from the coverslip, especially as the two are in close proximity.5In order to reduce these nonspecific interactions during the DNA rupture studies, we tried to en-hance the specific interaction by modifying both the probe and the coverslip surface with biotin and avidin. The un-bound active sites of avidin were blocked and protected with excess free biotin by the method of Wojcikiewicz et al.,8 which can increase the specific interactions among DNA triplexes.
We implemented a dynamic force spectroscopy mode in the software-controlled piezodisplacement by Asylum Re-search and used the functionalized tip, as described previously.8The constant velocity approach-retract cycle was repeated 50 times in triplex formation buffer at 25 ° C. The spring constants of all cantilevers were calibrated by the thermal fluctuation method9 with an absolute uncertainty of ±10%. The measurements presented were performed with
a兲Authors to whom correspondence should be addressed.
b兲Also at: Institute of Physics, Academia Sinica, Taipei. 11529 Taiwan. Tel.:
⫹886-35712121 ext. 56958. FAX: ⫹886-3-5733259. Electronic mail: [email protected]
c兲Also at: Institute of Biophotonics, National Yang Ming University, Taipei,
Taiwan.
d兲Tel.: ⫹886-2-27898550. FAX: ⫹886-2-27884184. Electronic mail:
e兲Tel.: ⫹886-2-27896722. FAX: ⫹886-2—27834187. Electronic mail:
APPLIED PHYSICS LETTERS 91, 203901共2007兲
0003-6951/2007/91共20兲/203901/3/$23.00 91, 203901-1 © 2007 American Institute of Physics This article is copyrighted as indicated in the article. Reuse of AIP content is subject to the terms at: http://scitation.aip.org/termsconditions. Downloaded to IP:
eight different cantilevers with spring constants ranging from 4 to 7 pN/ nm. The thermal fluctuation of the cantilevers was about 1.5 nm. When the DNA rupture process was finished the molecular fluorescence image of DNA triplex was taken on the Olympus IX71 total internal reflection fluorescence microscopy system 共Olympus Optical Co. Ltd., Tokyo, Ja-pan兲 with cool pix charge-coupled device 共iXon DU-887, Andor兲 at −10 °C.
During the probe withdrawal, a nonspecific probe-substrate interaction can still be detected when the piezodis-placement was around 10 nm关Fig.2共a兲兴, owing possibly to the extension of the polyglutaraldehyde-avidin complex.10 Unlike the wormlike-chain model of polyprotein experiments,5the force extension curve remains in a plateau 共from 10 to 15 nm piezodisplacement兲 when the nonspecific interaction is relieved. This may have been caused by stretching of the glutaraldehyde-avidin-共T兲10linker and
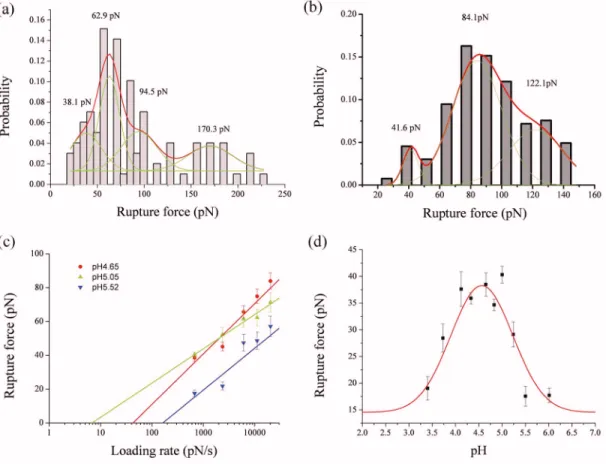
con-formational change of the ribose in DNA.4 The stretching force is released suddenly when the probe is pulled 15 nm away from the contact point. Meanwhile, fluorescence signal can still be observed after pulling the DNA molecule for 50 times 关right panel of Fig. 2共b兲兴. This fluorescence image showed a sudden drop of the extension force which we as-cribed to a rupturing event of the DNA triplex. The rupture force histogram analysis for 50 approach-retract cycles of AFM indicated that this fluorescence image may be caused by 24 rupture molecules. Therefore, this is a fluorescence image at the molecular level. By applying a different loading rate, from 676 to 20 280 pN/ s, the rupture force exerted on the DNA triplex varies accordingly. The rupture force can be revealed from the frequency histograms of the Gaussian dis-tribution fitting at different loading rates关Figs3共a兲and3共b兲兴. In the case of DNA triplex environment in pH 4.65, the rupture force increases from 38.1 pN 共at a loading rate of 676 pN/ s兲 to 83.9 pN 共at a loading rate of 20 280 pN/s兲 关the red line of Fig.3共c兲兴. There is a linear relation between the rupture force and the logarithm of the loading rate, indicative of a single potential well model for the DNA rupture.11,12 Furthermore, we observed a single rupture force共i.e., from all to none兲 at a piezodisplacement of 15 nm during the dis-sociation process 关Fig. 2共a兲兴. Therefore, we concluded that the DNA rupture, in this occasion, is a two-state process; namely, all the hydrogen bonds and the stacking force of the base pairs between the Hoogsteen and Watson strands broke
at the same time 共i.e., from all to none兲 during the DNA triplex dissociation process. The inclusion of the GA step is destabilizing and perhaps the reason for a two-state rupture. The Koffvalues of triplex DNA rupture event, derived from
Evans’ study,12 were 3.13⫻10−2 and 9.38⫻10−2s−1 at pH
4.33 and 3.48, respectively.
In order to understand the effect of pH on the single molecular mechanism during the triplex formation, we per-formed DNA triplex rupture experiments by varying pH val-ues from 3.40 to 6.01 systematically. The maximum rupture force occurred in the range from pH 4.12 to 5.00, with mag-nitude between 37.6± 2.2 and 40.3± 1.6 pN 关Fig.3共d兲兴. The data points fit a Gaussian curve nicely with a maximum at
pH 4.56. However, the rupture force declines rapidly to
17.6± 1.9 pN when the pH is raised to 5.5, indicating that the cytosines of the Hoogsteen strand are no longer protonated and, consequently, the ability to form the hydrogen bonds with guanine of the Watson strand is reduced.1,2
Although the pKaof cytosine is 4.2, the pKaof the N3on
cytosine base in a nucleotide chain varies with its local environment,13 such as the dielectric constant 共of interior DNA兲 and electrostatic potential 共of neighboring bases兲.14 Thus, the pKa’s of C+ 共shown by the midpoint of the
asso-ciation兲 in different sequences and chain lengths are usually measured experimentally.1According to previous studies,1,14 it is not uncommon that the pKaof N3on the cytosine base
in a nucleotide chain may rise to 5.6, consistent with our experimental observation here. The peak at 4.56 is due to the
FIG. 1. 共Color online兲 Single-molecule image and height of DNA triplex oligo 1⫹ oligo 2. The inset of denotes the possible conformation of DNA triplex.
FIG. 2.共a兲 The rupture force curve of DNA triplex. The first rupture force at 10 nm from the contact point may be caused by the nonspecific binding between the tip and the glutaraldehyde-avidin modified coverslip surface. The last rupture force, at 15 nm from the contact point, comes from break-age of the hydrogen bonds between the Watson strand of oligo1 and oligo2 共H strand兲. 共b兲 The fluorescence images before 共left panel兲 and after 共right panel兲 the probe withdrawal process. The left image denotes the fluores-cence image observed before the probe retraction process when W and H are associated as triplex and F is quenched. It serves as a background intensity image. A brighter spot at the right denotes the fluorescence image observed after the probe retraction process.
203901-2 Chang et al. Appl. Phys. Lett. 91, 203901共2007兲
This article is copyrighted as indicated in the article. Reuse of AIP content is subject to the terms at: http://scitation.aip.org/termsconditions. Downloaded to IP: 140.113.38.11 On: Wed, 30 Apr 2014 07:00:45
formation of a fully protonated triplex, this means that the
pKaof cytosine is higher than 5.6 and consistent with NMR
observations of the literature.1One should also pay attention to the underlying DNA duplex stability.15Although the for-mation of DNA duplex may be less favorable in a lower pH environment, however, weaker duplexes may yield stronger triplexes. Therefore, we observed the strongest triplex forma-tion at pH 4.56.
The rupture force decreases to 19.0± 2.2 pN at pH 3.40, which means that the overall hydrogen bonds of the triplex are weakened in a low pH environment. The baseline of Gaussian distribution curve fitting is around 14.57 pN. This indicates that the DNA triplex may dissociate spontaneously at a pH level lower than 2.5 or higher than 6.5. The forma-tion mechanism of DNA triplex, revealed here by AFM at the single-molecule level, is consistent with our previous macro-scopic results1,2 that both protonated cytosine and pH envi-ronment play vital roles in the DNA triplex formation.
In summary, we have designed a pair of specific nucle-otide sequences, with which a 17-mer DNA triplex can be formed. By combining AFM and FRET techniques the dy-namics of its dissociation mechanism has been revealed at the single-molecular level. DNA dynamics and the related molecular mechanism of other sequences, at the single-molecule level, are under investigation following similar de-sign and methods.
Grant supports were from National Science Council, Tai-wan, R.O.C. 共Nos. NSC 95-2112-M-009-019 for C.C.C. NSC 95-2120-M-001-007 for C.S.C. and NSC 94-2120-M-001-008 for L.S.K.兲
1D. E. Callahan, T. L. Trapane, P. S. Miller, P. O. P. Ts’o, and L. S. Kan,
Biochemistry 30, 1650共1991兲.
2L. S. Kan, L. Pasternack, M. T. Wey, Y. Y. Tseng, and D. H. Huang,
Biophys. J. 91, 2552共2006兲.
3A. Hillisch, M. Lorenz, and S. Diekmann, Curr. Opin. Struct. Biol. 11,
201共2001兲.
4P. E. Marszalek, H. Li, A. F. Oberhauser, and J. M. Fernandez, Proc. Natl.
Acad. Sci. U.S.A. 99, 4278共2002兲.
5T. E. Fisher, P. E. Marszalek, and J. M. Fernandez, Nat. Struct. Biol. 7,
719共2000兲.
6M. P. Gustafson, C. F. Thomas, Jr., F. Rusnak, A. H. Limper, and E. B.
Leof, J. Biol. Chem. 276, 835共2001兲.
7H. G. Hansma, I. Revenko, K. Kim, and D. E. Laney, Nucleic Acids Res.
24, 713共1996兲.
8E. P. Wojcikiewicz, X. Zhang, and V. T. Moy, Biol. Proced. Online 6, 1
共2004兲.
9J. L. Hutter and J. Bechhoefer, Rev. Sci. Instrum. 64, 1868共1993兲. 10G. T. Hermanson, Bioconjugate Techniques共Academic, Burlinton, MA,
1996兲, 16, p. 630.
11A. F. Oberhauser, B. F. Carmelu, C. V. Mariano, and J. M. Fernandez, J.
Mol. Biol. 319, 433共2002兲.
12E. Evans, Annu. Rev. Biophys. Biomol. Struct. 30, 105共2001兲. 13A. S. Petrov, G. Lamm, and G. R. Pack, Biophys. J. 87, 3954共2004兲. 14G. R. Pack, L. Wong, and G. Lamn, Int. J. Quantum Chem. 70, 1177
共1998兲.
15D. E. Wemmer and A. S. Benight, Nucleic Acids Res. 13, 8611共1985兲.
FIG. 3.共Color online兲 Frequency histograms of rupture forces for DNA triplex at 共a兲 the loading rates of 676 pN/s 共n=309 rupture events兲 and 共b兲 2366 pN/s 共n=261 rupture events兲. Red lines: The Gaussian distribution fitting of the rupture forces. Green lines: The deconvolution of the Gaussian distribution which revealed the rupture force of the single molecule.共c兲 The loading rate vs the rupture forces of DNA triplex as a function of pH. The red, green, and blue dots 共with least square fittings兲 denote the rupture forces in pH 4.65, 5.05, and 5.52 buffer conditions, respectively. 共d兲 The rupture forces of DNA triplex 共black squares with standard error bars兲 and their Guassian distribution fittings in different pH environments. The loading rate is 676 pN/s.
203901-3 Chang et al. Appl. Phys. Lett. 91, 203901共2007兲
This article is copyrighted as indicated in the article. Reuse of AIP content is subject to the terms at: http://scitation.aip.org/termsconditions. Downloaded to IP: 140.113.38.11 On: Wed, 30 Apr 2014 07:00:45