中 國 醫 藥 大 學
專題研究計畫成果報告
計畫名稱:運動訓練合併冬蟲夏草於抗敗血病的作用
計畫編號:CMU 98- N2-12
執行期限:2010 年 4 月 12 日至 2011 年 03 月 31 日
單位名稱:物理治療學系
主持人:鄭宇容
中 華 民 國
100 年 06 月 29 日
附件中文摘要
冬蟲夏草是應用於治療多種發炎性疾病的中草藥,有文獻指出運動訓練可
以降低敗血症帶來的傷害。本篇的目的是研究冬蟲夏草合併運動訓練對於
敗血症引起之肝臟傷害的影響。實驗使用三十二隻小鼠(C57BL/6 mice),
隨機分成八組,其中四組小鼠沒有進行運動訓練,小鼠管餵冬蟲夏草的劑
量分成: 0mg(saline)、20mg、40mg,在運動訓練結束後使用脂多糖(LPS)/
半乳糖胺(D-GalN)誘導敗血症,取出血液與肝臟測量蘇木紫-伊紅染色
(Hematoxylin-Eosin Y staining)、天冬氨酸氨基轉移酶 (aspartate
aminotransferase, AST)、腫瘤壞死因子–α(TNF-α)、介白素-10 (IL-10)、
超氧化物歧化酶(superoxide dismutase, SOD)、一氧化氮(nitric oxide, NO)、肝
臟細胞凋亡表現蛋白、TUNEL陽性細胞。實驗結果表示單獨進行運動訓練
增加了IL-10、SOD表現並降低AST、TNF-α、NO,減少肝臟凋亡表現蛋
白與TUNEL陽性細胞。單獨給予高劑量蟲草可以增加IL-10、SOD表現並降
低AST、TNF-α、NO,減少肝臟凋亡表現蛋白與TUNEL陽性細胞。高劑
量冬蟲夏草合併運動訓練降低了冬蟲夏草的保護效果,增加AST、TNF-α、
NO產生,減少IL-10、SOD並活化了肝臟凋亡表現蛋白與增加TUNEL陽性
細胞產生。總結是單獨進行運動訓練或是高劑量蟲草可以降低敗血症引起
的肝臟傷害,而高劑量冬蟲夏草合併運動訓練則降低了冬蟲夏草的保護效
果。
INTRODUCTION 1. Sepsis
Sepsis is a severe inflammatory disorder, and serious clinical problem with high mortality may lead to severe shock and multiple organ failure. The interaction between infecting
microorganism and host immune, inflammation, and coagulation responses results in sepsis. It is defined as the systemic response to infection, with the most common cause being a
contamination of the blood with bacteria [35].
The infection activates various mechanisms to cause tissue damage, including cytokines production which initiates the inflammatory response and changes in the systemic circulation and coagulation cascade. An unregulated response to multiple organ failure trigger by sepsis is characterized by liver, cardiovascular, pulmonary, renal and gastrointestinal dysfunction. In particular, the severe liver failure after sepsis is associated with a high mortality rate [8] [24] [35].
Lipopolysaccharide (LPS) has been used for induction of endotoxemia in animal
experimentation and is responsible for the multiple organ dysfunctions induced by septic shock. LPS from gram negative bacteria bound with toll-like receptors (TLRs) can arrange other signaling molecules to release cytokines [24]. This process involves the activation of mitogen activated protein kinase (MAPK) [29]. Cytokines then cause oxidant stress production and tissue injury eventually. The pro-inflammatory cytokines can decrease
systemic vascular resistance, resulting in profound hypotension by inducing the expression of inducible nitric oxide synthase (iNOS) and augmenting the production of nitric oxide (NO) [37][44]. D-galactosamine (D-GalN) is a hepatotoxic agent, which increases the susceptibility of mice to LPS-induced shock by impairing liver metabolism [27].
2. Sepsis-induced liver dysfunction
Liver appears to be a key organ in sepsis, and portal blood flow is a subject to
vasoconstriction and bacterial translocation during sepsis. The liver is made mainly of hepatocytes, Kupffer cells, and endothelial sinusoidal cells. All of these cell types are involved in the metabolic, anti-infectious and immune responses through multiple cell interactions. Kupffer cells exposed to endotoxin in vitro produce large amounts of tumor necrosis factor- TNF- . TNF- is also involved in induction of the nitric oxide synthase gene (iNOS) transcription, accounting for nitric oxide hepatic production [8].
Sepsis-induced liver dysfunction is usually attributed to systemic disturbances, endotoxin, the activation of inflammatory cytokines, and mediators. The overproduction of locally secreted and circulating mediators, such as reactive oxygen species, proinflammatory cytokines, and nitric oxide formation leading to tissue damage and cell death which contributed to liver injury [8].
3. Role of pro-inflammatory cytokines in sepsis
Tumor necrosis factor- alpha (TNF-α) is recognized as an important mediator in the development of sepsis and in the systemic inflammatory response syndrome (SIRS) [30].
While Toll like receptor-4 (TLR-4) recognizes the LPS of gram-negative bacteria, binding of TLRs stimulates intracellular signaling which increase the secretion of proinflammatory molecules such as TNF-α and interleukin- 1β, as well as antiinflammatory cytokines such as interleukin-10 [35].
Macrophages represent a major defense system against attack of the host by microorganisms and release a vast group of mediators including cytokines, such as TNF-α and IL-1- , arachidonic acid metabolites, and NO that amplify the inflammatory response. TNF- α has been proposed as a therapeutic target because it plays a key role in arranging the cytokine cascade in many inflammatory diseases and it is a “master-regulator” of inflammatory cytokine production[24][30]. Proinflammatory cytokines up-regulate adhesion molecules in neutrophils and endothelial cells. Activated neutrophils exterminate microorganisms and damage endothelium by releasing mediators that increase vascular permeability. In addition, activated endothelial cells release nitric oxide, a potent vasodilator that acts as a key mediator of septic shock [2] [11].
4. Role of apoptosis in sepsis
Apoptosis, commonly termed programmed cell death, can proceed by the death receptor or mitochondrial-mediated pathways that ultimately converge to activate executioner caspases including caspase-3 and caspase-6 that mediate the systematic demolition of the cell [6]. Apoptotic cell death plays an important role in LPS-related tissue injuries in clinical systemic inflammatory response syndrome, septic shock, and multiple organ failure [12]. The
inflammatory and metabolic changes develop in septic patients and change the various organs into pathophysiology.
Endotoxin and various ROS elicit DNA strand breaks, which in turn activate Poly (ADP-ribose) polymerase (PARP). The role for PARP in DNA repair suggests that PARP activation serves to help rescue damaged cells and this can occur following mild DNA damage. However, the opposite is true in excessive DNA damage. Cleavage of PARP might occur when cell is no longer able to repair its DNA and it ensures commitment to apoptosis. The excess NO production in Septic shock also activates PARP and damages DNA [32]. 5. Role of nitric oxide (NO) in sepsis
Nitric oxide (NO) is involved in smooth muscle relaxation, and increased NO release contributes to sepsis-induced hypotension. NO is produced by three different NO synthases (NOS); neuronal NOS (nNOS), inducible NOS (iNOS) and endothelial NOS (eNOS). Under normal circumstances, iNOS activity stays in low levels, but its synthesis is rapidly stimulated by a wide range of inflammatory agents with endotoxin [4].
NO is generated from L-arginine and iNOS produces large amounts of NO over prolonged periods of time. The NO metabolites nitrite and nitrate, which is the indicators of NO production, raise progressively in various animal shock models. The enhanced formation of endogenous NO contributes to the hypotension caused by endotoxin and TNF- . NO may also directly contribute to tissue and organ injury by its direct, peroxynitrite-mediated
cytotoxic effects [4] [5] [11].
6. The effect of Cordyceps sinensis in sepsis
Cordyceps fungi, such as Cordyceps sinensis (C. sinensis), is a parasitic complex of fungus and caterpillar, which has been utilized as a traditional medicine and used for medicinal purposes for centuries particularly in Asia. C. sinensis has many bioactive components, such as 3′-deoxyadenosine (cordycepin), cordycepic acid, and Cordyceps polysaccharides [43]. C. sinensis shows various biological effects, e.g. down-regulation of inflammation-related and apoptotic genes [38], anti-tumor activity [7], and suppression of autoimmune diseases and allergy with their immunomodulating effects [43]. C. sinensis has also been used to treat liver, inflammatory and autoimmune diseases [21] [22] [23]. C. sinensis acts to accelerate
macrophage phagocytosis, enhances the immune function and reverse the developing process of liver fibrosis [23].
The anti-inflammatory effect of C. sinensis might result from the inhibition of TNF-α [33] [41]. It has been shown that Cordyceps pruinosa methanol extract suppresses inflammation through suppression of NF- B-dependent inflammatory gene expression, which suggesting that the Cordyceps pruinosa extract may be beneficial for treatment of endotoxin shock or sepsis [14]. However, the mechanisms of C. sinensis contribute to the beneficial effect for hepatic dysfunction induced by sepsis is unknown.
7. The effects of exercise in sepsis
In general, regular exercise can enhance immunological and antioxidant functions [26]. The mechanisms of exercise-associated immune changes are multifactorial and include multiple neuroendocrinological factors. Alterations in metabolism and metabolic factors contribute to exercise-associated changes in immune function. Exercise has important modulatory effects on immunocyte dynamics and possibly on immune function [1]. These effects are mediated by exercise-induced release of proinflammatory cytokines, classical stress hormones, and
hemodynamic effects leading to cell redistribution [31].
Chen et al. [3] suggested that exercise training attenuates septic responses and protects organs from damage in sepsis. The adaptations promoted by swimming training seem to be beneficial, counteracting the cardiovascular abnormalities and pulmonary edema seen in septicemia induced by LPS [25]. The appropriate exercise in humans increases the plasma cytokine IL-6 and suppresses the endotoxemia-induced elevation in TNF-α [39]. They conclude that
exercise exerts anti-inflammatory effects. Because the anti-inflammatory effect of C. sinensis and exercise training attenuates septic responses and protects organs from damage, the present study was designed to test that C. sinensis combined with precondition exercise training can attenuate hepatic injury in sepsis.
MATERIALS AND METHODS Animal groups
Laboratory Animal Center (NLAC), Taiwan. Mice were randomly assigned into control (without LPS combined with D-galactosamine (D-GalN) infusion), saline, CS20mg and CS40mg with or without pre-condition swimming exercise training.
Cordyceps sinensis treatment
The extract of C. sinensis is purchased from TCM Biotech International Corp., Taiwan. Mice were treated with saline or the extract of C. sinensis (20 mg/kg or 40 mg/kg) by oral gavage 4 hours before the precondition swimming exercise training.
Precondition exercise training
Four hours after C. sinensis treatment, mice were treated with precondition exercise training. The exercise protocol is swimming exercise for 30 minutes/day, 5days/week for 4 weeks in 26-28℃ water.
Induction of sepsis
At the end of treatment, mice were induced sepsis by LPS with D-GalN through
intraperitoneally injection (LPS: 10 g/kg. Escherichia coli, B4, 10; D-GalN: 800 mg/kg, Sigma-Aldrich, St. Louis, MO, USA.).
Hematoxylin-Eosin Y staining
The livers were excised and soaked in formalin and processed to paraffin blocks. Slides were prepared by deparaffinization and rehydration. They were passed through a series of graded alcohols (100%, 95%, and 75%), for 15 minutes each. The slides were then dyed with hematoxylin and eosin for 1 minute. After being gently rinsed with water, slides were then soaked with 85% alcohol, two changes of 100% alcohol for 15 minutes each. At the end, the slides were soaked in two changes of Xylene. Photomicrographs were obtained using Zeiss AxioImager M1 microscope (Carl Zeiss MicroImaging GmbH, Jena, Germany).
Biochemistry study
Blood samples were collected at 6 hours after LPS/D-GalN infusion, and immediately centrifuged at 5000g for 10 min. Plasma was stored at 4°c for biochemical examination after collection. Plasma samples were diluted 1:10 with distilled water before measurements. Aspartate aminotransferase (AST) were evaluated of liver failure.
Enzyme-linked immunosorbent assay (ELISAs)
TNF-α and IL-10 was measured using antibody enzyme-linked immunosorbent assays (ELISAs). The experiment was repeated with blood collected at 3and 6 hours for cytokine analysis by intracardiac puncture and immediately allowed to centrifugation for 10 min at 5000g. The liver tissue was collected for analysis at 6 hours after LPS/D-GalN treatment. Liver tissue was homogenated and allowed to centrifugation for 20 min at 5000g. The supernatant was stored at _80°C for determination of IL-10 and TNF-α concentration using commercially available enzyme-linked immunosorbent assay kits (R&D Systems,
Minneapolis, MN, USA. Duo Set ELISA development kit) according to the manufacturer’s instructions. The optical density was measured at 450 nm wavelengths by Multiskan EX Microplate Photometer (Thermos, Waltham, MA, USA).
Measurement of nitrite oxide (NO) concentration
The nitrite concentration in serum were measured after the Griess reaction by incubating 20 L sample with 20 L of Griess reagent (Sigma-Aldrich, St. Louis, MO, USA) and added 160 L deionized water at room temperature for 20 minutes. The absorbance was measured at 550 nm by a spectrophotometer. Nitrite concentration was calculated by comparison with a standard solution of known sodium nitrite concentrations.
Superoxide dismutases (SOD) assay
The enzyme activity of SOD was measured in 10μL of liver sample using kit from Cayman Chemical (Ann Arbor, MI, USA). The method utilizes tetrazolium salt to quantifying
superoxide radicals generated by xanthine oxidase and hypoxanthine. The standard curve was generated using quality controlled SOD standard and the optical density was measured at 450 nm wavelengths.
Western Blot analysis
Mice liver tissue was harvested after 6 hours following LPS/ D-GalN treatment .The tissue were homogenized and lysed for 30 min in lysis buffer (homogenize tissue sample in 1:20 (w/v) of tissue to T-PER Reagent (Thermos, Waltham, MA, USA).
Samples were then centrifuged and supernatant was collected. Protein quantification is measured by Bradford method. Western blotting is analysis for cleaved caspase-3, cleaved caspase-6, cleaved PARP (Poly (ADP-ribose) polymerase), p38 MAPKs and ERK (p44/42) protein expression (Cell Signaling Technology, Danvers, MA, USA). 150 g proteins were separated by 12% sodium dodecyl sulphate-polyacrylamide gel electrophoresis (SDS-PAGE) with a constant voltage 80V and transferred onto polyvinylidene difluoride (PVDF)
membranes (Bio-Rad Laboratories, CA, USA) with a transfer apparatus (Bio-Rad Laboratories, CA, USA).
The membranes were blocked in 0.05% TBS-Tween (TBST) containing 5% milk and then incubated with the primary antibody (1:1,000) in TBST with 5% skim milk at 4 °C overnight. The primary antibody was detected by incubation with horseradish peroxidase-coupled
second antibody (1:2,000) at room temperature for 2 h. The chemiluminescence detection was performed by using Western Chemiluminescence HRP substrate (Millipore, Billerica, MA, USA). The blots were then stripped and incubated with anti -actin antibody (diluted 1:10000; Sigma-Aldrich, St. Louis, MO, USA) to ensure equal loading. Quantitative analysis of bands carried out using Gel-PRO analyzer Imaging System.
Terminal deoxynucleotidyl transferase (TdT)-mediated dUTP nick end labeling (TUNEL) ApopTag Peroxidase Kits (Millipore, Billerica, MA, USA) was used to label apoptotic cells. Paraffin slides were deparaffinized and pretreated with diluted Proteinase K. After washed the specimen in 2 changes of PBS, the specimen was put into 3.0% hydrogen peroxide in PBS for 5 minutes and rinsed twice with PBS for 5 minutes each time. The specimen was applied 75 μL/ 5cm2 of Equilibration buffer immediately and incubated for 10 seconds at room
chamber at 37°C for 1 hour.
After an hour, the specimen was applied Stop/Wash buffer and incubated for 10 minutes. After washed the specimen in 3 changes of PBS for 1 minute each wash, the slides were applied room temperature anti-digoxignenin conjugate and incubated in a humidified chamber for 30 minutes. The specimen was developed color in peroxidase substrate after wash in PBS and counter stained specimen in 0.5% (w: v) methyl green for 10 minutes. The specimen was dehydrated by moving the slide through three changes of xylene for 2 minutes each wash and mount under a glass coverslip in a mounting medium.
Photomicrographs were obtained using Zeiss AxioImager M1 microscope (Carl Zeiss MicroImaging GmbH, Jena, Germany). All counts were performed by at least two independent individuals in a blinded manner.
Statistical analysis
The data are presented as mean ± SEM. Statistical evaluation was performed by ANOVA and Tukey’s post hoc test. p<0.05 was considered to be statistically significant.
RESULTS
Effects of C. sinensis and exercise training on H&E stain in LPS/ D-GalN-treated mice
Light microscopy showed no infiltration or sequestration of neutrophils in liver in control group (Fig. 1), whereas more overt sequestration of neutrophils and coagulative necrosis tissues were found in liver in saline without exercise group. The neutrophil infiltrations and liver necrosis were significantly reduced in 40mg C. sinensis without exercise group (n = 4). Administration of exercise training only decreased LPS/ D-GalN-induced neutrophil
infiltrations and liver necrosis compared with non-exercise group (n = 4). Administration of 20 mg C. sinensis combined with exercise training decreased LPS/ D-GalN-induced
neutrophil infiltrations and liver necrosis (n = 4). However, 40 mg C. sinensis combined with exercise training significantly increased neutrophil infiltrations and liver necrosis (n = 4) compared to 40 mg C. sinensis without exercise group.
Effects of C. sinensis and exercise training on AST (Aspartate Aminotransferase) value in LPS/ D-GalN- treated mice
AST expression in serum was measured at 6 hours after LPS/ D-GalN challenge to examine the effect of C. sinensis and exercise training (Figure 2). AST concentration in LPS/ D-GalN- treated groups was significantly increased than control group (p < 0 .05), and significantly decreased with 40mg C. sinensis in non-exercise group (p < 0.05). Administration of exercise training only strongly decreased LPS/ D-GalN- induced AST elevation compared with
non-exercise group (p < 0.05). Administration of 20 mg C. sinensis only did not decreased LPS/ D-GalN -induced AST elevation and there was no difference in 20 mg C. sinensis combined with exercise training group. However, 40 mg C. sinensis combined with exercise
training significantly increased AST concentration compared to 40 mg C. sinensis without exercise group (p < 0 .05).
Effects of C. sinensis and exercise training on serum TNF-α concentration in LPS/ D-GalN -treated mice
To examine the effect of C. sinensis and exercise training on TNF-α level in serum, three hours serum was measured by ELISA (Figure 3). Serum TNF-α concentration at 3 hours after LPS/ D-GalN challenge were significantly increased (p < 0 .05), and increased dose of C. sinensis significantly decreased serum TNF-α in non-exercise group (all p < 0.05). Serum TNF-α concentration was significantly decreased in saline with exercise group (p < 0.05). Administration of 20 mg C. sinensis only decreased LPS/ D-GalN-induced TNF-α elevation(p < 0.05) and there is no difference in 20 mg C. sinensis combined with exercise training group. Administration of 40 mg C. sinensis only strongly decreased LPS/ D-GalN-induced TNF-α elevation (p < 0.05); however, 40 mg C. sinensis combined with exercise training increased TNF-α concentration compared to 40 mg C. sinensis without exercise group (p < 0 .05). Effects of C. sinensis and exercise training on TNF-α concentration of liver tissue in LPS/ D-GalN -treated mice
To examine the effect of C. sinensis and exercise training on TNF-α production induced by LPS/D-GalN treatment, the excised liver of mice was measured by ELISA (Figure 4).Liver TNF-α concentration after LPS/ D-GalN challenge were significantly increased (p < 0 .05), increased dose of C. sinensis significantly decreased liver TNF-α in non-exercise group (all p < 0.05). TNF-α concentration of liver after LPS/ D-GalN challenge were significantly
decreased in saline with exercise group (p < 0.05). Administration of 20 mg C. sinensis only decreased LPS/ D-GalN-induced TNF-α elevation (p < 0.05) but there is no difference in 20 mg C. sinensis combined with exercise training group. Administration of 40 mg C. sinensis only strongly decreased LPS/ D-GalN-induced TNF-α elevation (p < 0.05); however, 40 mg C. sinensis combined with exercise training increased TNF-α concentration(p < 0 .05).
Effects of C. sinensis and exercise training on serum IL-10 concentration in LPS/ D-GalN -treated mice
To examine the effect of C. sinensis and exercise training on IL-10 production of serum, six hours serum of mice was measured by ELISA (Figure 5). Serum IL-10 concentration at 6 hours after LPS/ D-GalN challenge were significantly increased (p < 0 .05), increased dose of C. sinensis significantly increased serum IL-10 in non-exercise group (all p < 0.05). Serum IL-10 concentrations at 6 hours after LPS/ D-GalN challenge were significantly increased in saline with exercise group (p < 0.05). Administration of 20 mg C. sinensis only increased LPS/ D-GalN-induced IL-10 elevation (p < 0.05)and 20 mg C. sinensis with exercise training increased more IL-10 elevation after LPS/ D-GalN treatment (p <0 .05). Administration of 40
mg C. sinensis only strongly increased LPS/ D-GalN -induced IL-10 elevation (p < 0.05); however, 40 mg C. sinensis combined with exercise training decreased IL-10 concentration compared to 40 mg C. sinensis without exercise group (p < 0 .05).
Effects of C. sinensis and exercise training on IL-10 concentration of liver tissue in LPS/ D-GalN -treated mice
To examine the effect of C. sinensis and exercise training on IL-10 production of liver was measured by ELISA (Figure 6). Liver IL-10 concentration after LPS/ D-GalN challenge were significantly increased (p < 0 .05), increased dose of C. sinensis significantly increased liver IL-10 in non-exercise group (all p < 0.05). Liver IL-10 concentrations after LPS/ D-GalN challenge were significantly increased in saline with exercise group (p < 0.05). Administration of 20 mg
C. sinensis only increased LPS/ D-GalN-induced IL-10 elevation (p < 0.05) and 20 mg C. sinensis with exercise training increased more IL-10 elevation after LPS/ D-GalN treatment (p < 0 .05). Administration of 40 mg C. sinensis only strongly increased LPS/ D-GalN-induced IL-10 elevation(p < 0.05); however, 40 mg C. sinensis combined with exercise training decreased IL-10 concentration compared to 40 mg C. sinensis without exercise group (p < 0 .05).
Effects of C. sinensis and exercise training on NO production of three hours serum in LPS/ D-GalN -treated mice
To examine the effect of C. sinensis and exercise training on NO production, three hours serum was measured by Griess Reaction (Figure 7). Serum nitrite concentration at 3 hours after LPS/ D-GalN challenge were significantly increased (p < 0 .05), increased dose of C. sinensis significantly decreased serum nitrite in non-exercise group (all p < 0.05). Serum nitrite concentrations at 3 hours after LPS/ D-GalN challenge were significantly decreased in saline with exercise group (p < 0.05). Administration of 20 mg C. sinensis only decreased LPS/ D-GalN-induced nitrite elevation (p < 0.05); however, 20 mg C. sinensis combined with exercise training increased LPS/ D-GalN-induced nitrite concentration (p < 0.05).
Administration of 40 mg C. sinensis only strongly decreased LPS/ D-GalN-induced nitrite elevation (p < 0.05). However, 40 mg C. sinensis combined with exercise training increased nitrite concentration compared to 40 mg C. sinensis without exercise group (p <0 .05). Effects of C. sinensis and exercise training on NO production of six hours serum in LPS/ D-GalN -treated mice
To examine the effect of C. sinensis and exercise training on NO production, six hours serum was measured by Griess Reaction (Figure 8). Serum nitrite concentration at 6 hours after LPS/ D-GalN challenge were significantly increased (p < 0 .05), increased dose of C. sinensis significantly decreased serum nitrite in non-exercise group (all p < 0.05). Serum nitrite concentrations at 6 hours after LPS/ D-GalN challenge were significantly decreased in saline
with exercise group (p < 0.05). Administration of 20 mg C. sinensis only decreased LPS/ D-GalN-induced nitrite elevation (p < 0.05) and there is no difference in 20 mg C. sinensis combined with exercise training group. Administration of 40 mg C. sinensis only strongly decreased LPS/ D-GalN-induced nitrite elevation (p < 0.05); however, 40 mg C. sinensis combined with exercise training increased nitrite concentration (p < 0 .05).
Effects of C. sinensis and exercise training on nitrite oxide production of liver in LPS/ D-GalN-treated mice
To examine the effect of C. sinensis and exercise training on NO production, liver tissue was measured by Griess Reaction (Figure 9). Liver nitrite concentration after LPS/ D-GalN challenge were significantly increased (p < 0 .05), increased dose of C. sinensis significantly decreased liver nitrite in non-exercise group (all p < 0.05). Liver nitrite concentrations after LPS/ D-GalN challenge were significantly decreased in saline with exercise group (p < 0.05). Administration of 20 mg C. sinensis only decreased LPS/ D-GalN-induced nitrite elevation (p < 0.05) and there is no difference in 20 mg C. sinensis combined with exercise training group. Administration of 40 mg C. sinensis only strongly decreased LPS/ D-GalN-induced nitrite elevation (p < 0.05); however, 40 mg C. sinensis combined with exercise training increased nitrite concentration
(p < 0 .05).
Effects of C. sinensis and exercise training on SOD (superoxide dismutases) levels in LPS/ D-GalN -treated mice
To examine the effect of C. sinensis and exercise training on SOD production, liver homogenate was measured by ELISA (Figure 10). SOD concentration after LPS/ D-GalN challenge were significantly decreased (p < 0 .05), increased dose of C. sinensis significantly increased SOD level in non-exercise group (all p < 0.05). SOD concentrations after LPS/ D-GalN challenge were significantly increased in saline with exercise group (p < 0.05). Administration of 20 mg C. sinensis only increased LPS/ D-GalN-induced SOD elevation (p < 0 .05) and there is no difference in 20 mg C. sinensis combined with exercise training group. Administration of 40 mg C. sinensis only strongly increased LPS/ D-GalN -induced SOD elevation (p < 0.05); however, 40 mg C. sinensis combined with exercise training decreased SOD concentration (p < 0 .05).
Effects of C. sinensis and exercise training on ERK (P44/42) protein level in LPS/D-GalN -treated mice
The effect of C. sinensis and exercise training on ERK pathway was measured by Western Blotting. Liver tissue was harvested and homogenate was analyzed after 6 hour of LPS/ D-GalN infusion (Figure 11). Increased dose of C. sinensis significantly increased ERK protein in non-exercise group (all p < 0.05). The phosphorylation of ERK after LPS/ D-GalN
challenge increased in saline with exercise group (p < 0.05). Administration of 20 mg C. sinensis only increased LPS/ D-GalN-induced ERK elevation (p < 0 .05). However, the phosphorylation of ERK after LPS/ D-GalN challenge were no different in 20 mg C. sinensis combined with exercise training group. Administration of 40 mg C. sinensis only strongly increased LPS/ D-GalN -induced ERK phosphorylation (p < 0.05); however, 40 mg C. sinensis combined with exercise training decreased ERK activation (p < 0 .05).
Effects of C. sinensis and exercise training on p38 mitogen-activated protein kinase (MAPK) protein level in LPS/ D-GalN -treated mice
To examine the effect of C. sinensis and exercise training on activation p38 pathway, the liver tissue were analyzed by Western Blotting (Figure 12). Increased dose of C. sinensis
significantly decreased p38 protein in non-exercise group (all p < 0.05). The phosphorylation of p38 after LPS/ D-GalN challenge was no different in saline with exercise group (p < 0.05). Administration of 20 mg C. sinensis only decreased LPS/ D-GalN-induced p38 elevation (p < 0 .05). However, 20 mg C. sinensis combined with exercise training increased LPS/
D-GalN-induced p38 activation (p < 0.05). Administration of 40 mg C. sinensis only strongly decreased LPS/ D-GalN-induced p38 phosphorylation (p < 0.05); however, 40 mg C. sinensis combined with exercise training increased p38 activation (p < 0 .05).
Effects of C. sinensis and exercise training on cleaved-caspase 3 protein level in LPS/ D-GalN -treated mice
The effect of C. sinensis and exercise training on cleaved-caspase 3 pathway was measured by Western Blotting. Liver tissue was harvested and homogenate was analyzed after 6 hours of LPS/ D-GalN infusion (Figure 13). Cleaved-caspase 3 protein levels after LPS/ D-GalN challenge were significantly increased (p < 0 .05) and increased dose of C. sinensis significantly decreased cleaved-caspase 3 protein activation in non-exercise group (all p < 0.05). Cleaved-caspase 3 protein after LPS/ D-GalN challenge was significantly decreased in saline with exercise group (p < 0.05). There was no difference between 20 mg C. sinensis alone treatment and 20 mg C. sinensis combined with exercise training group. Administration of 40 mg C. sinensis only strongly decreased LPS/ D-GalN -induced cleaved-caspase 3 activation (p < 0.05); however, 40 mg C. sinensis combined with exercise training increased cleaved-caspase 3 activation (p < 0 .05).
Effects of C. sinensis and exercise training on cleaved-caspase 6 protein level in LPS/ D-GalN -treated mice
The effect of C. sinensis and exercise training on cleaved-caspase 6 pathway was measured by Western Blotting. Liver tissue was harvested and homogenate was analyzed after 6 hours of LPS/ D-GalN infusion (Figure 14). Cleaved-caspase 6 protein level after LPS/ D-GalN challenge were significantly increased (p < 0 .05) and 40mg of C. sinensis significantly
decreased cleaved-caspase 6 protein activation in non-exercise group (p < 0.05). Cleaved-caspase 6 protein after LPS/ D-GalN challenge were significantly decreased
activation in saline with exercise group (p < 0.05). There was no difference between 20 mg C. sinensis alone treatment and 20 mg C. sinensis combined with exercise training group.
Administration of 40 mg C. sinensis only strongly decreased LPS/ D-GalN -induced
cleaved-caspase 6 activation (p < 0.05); however, 40 mg C. sinensis combined with exercise training increased cleaved-caspase 6 activation (p < 0 .05).
Effects of C. sinensis and exercise training on cleaved-PARP (Poly ADP ribose polymerase) protein level in LPS/ D-GalN -treated mice
The effect of C. sinensis and exercise training on cleaved-PARP pathway was measured by Western Blotting. Liver tissue was harvested and homogenate was analyzed after 6 hours of LPS/ D-GalN infusion (Figure 15). Cleaved-PARP protein level after LPS/ D-GalN challenge were significantly increased (p < 0 .05) and increased dose of C. sinensis significantly
decreased cleaved-PARP activation in non-exercise group (all p < 0.05). Cleaved-PARP protein after
LPS/ D-GalN challenge were significantly decreased in saline with exercise group (p < 0.05). Administration of 20 mg C. sinensis only decreased LPS/ D-GalN-induced cleaved-PARP activation (p < 0 .05) and there was no difference in 20 mg C. sinensis combined with exercise training group. Administration of 40 mg C. sinensis only strongly decreased LPS/ D-GalN -induced cleaved-PARP protein (p < 0 .05); however, 40 mg C. sinensis combined with exercise training increased cleaved-PARP elevation (p < 0 .05).
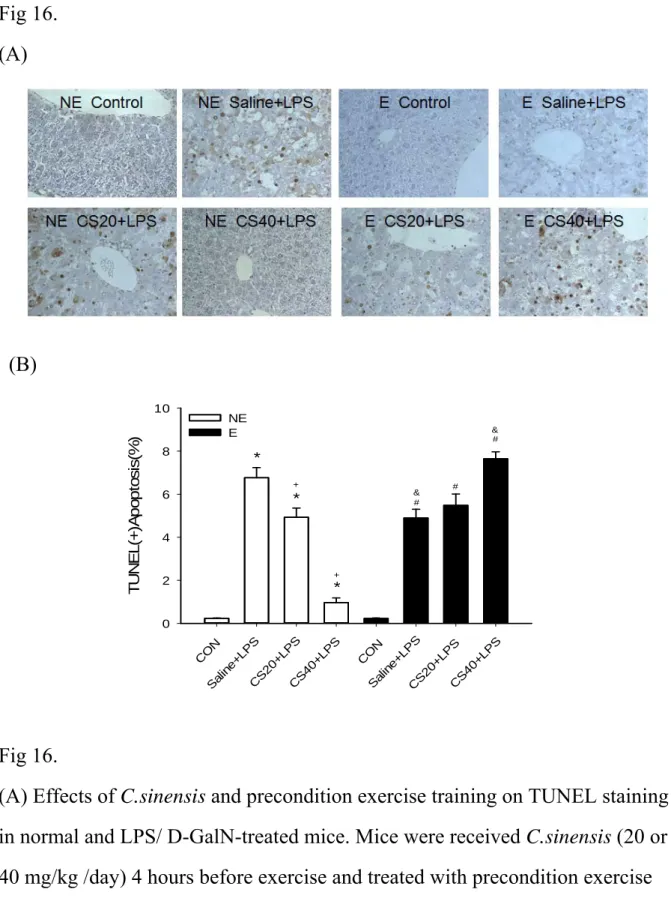
Effects of C. sinensis and exercise training on terminal deoxynucleotidyl transferase (TdT)-mediated dUTP nick end labeling (TUNEL) staining in
LPS/ D-GalN -treated mice
To examine the effect of C. sinensis and exercise training on hepatic cell apoptosis induced by LPS/ D-GalN treatment, the liver tissue were measured by TUNEL staining (Figure 16). TUNEL positive cells after LPS/ D-GalN challenge were significantly increased (p < 0 .05) and increased dose of C. sinensis significantly decreased TUNEL positive cells in
non-exercise group (all p < 0.05). TUNEL positive cells after LPS/ D-GalN challenge were significantly decreased in saline with exercise group (p < 0.05). Administration of 20 mg C. sinensis only decreased LPS/ D-GalN-induced TUNEL positive cells elevation (p < 0 .05) and there is no difference in 20 mg C. sinensis combined with exercise training group.
Administration of 40 mg C. sinensis only strongly decreased LPS/ D-GalN -induced TUNEL positive cells (p < 0 .05); however, 40 mg C. sinensis combined with exercise training
increased TUNEL positive cells (p < 0 .05). DISCUSSION
The main findings can be summarized as follows: (1) High dose C. sinensis treatment alone decreased the tissue damage induced by sepsis. The AST value, TNF- , and nitrite
concentration in serum and liver were significantly decreased after LPS/D-GalN infusion. The phosphorylation of p38 MAPK and apoptotic proteins including cleaved-caspase-3, 6 and cleaved-PARP activation in liver tissue were significantly decreased. SOD concentration and phosphorylation of ERK protein in liver were significantly increased. IL-10 level in serum and liver was also increased. (2) Precondition exercise training alone could decrease the damage induced by sepsis. The AST value, TNF- level in serum and liver, nitrite
concentration in serum and liver were significantly decreased after LPS/D-GalN infusion. The apoptotic proteins including cleaved-caspase-3, 6 and cleaved-PARP levels in liver were significantly decreased after LPS/ D-GalN infusion. Phosphorylation of ERK protein levels in liver tissue were significantly increased after LPS/ D-GalN infusion. SOD concentration and IL-10 level in serum and liver was significantly increased after LPS/D-GalN infusion.
In the current study, C. sinensis suppresses the expression of TNF- , NO formation, and the apoptotic injury in liver of LPS/D-GalN stimulated mice. C. sinensis increases the superoxide dismutase activity decreased by LPS/ D-GalN stimulation. It implicated that high dose C. sinensis treatment might decreased the tissue damage induced by sepsis through decreasing TNF- production and NO formation, increasing IL-10 and SOD production and reducing cell apoptosis
Our results showing that precondition exercise training may decrease the tissue damage induced by sepsis through decreasing TNF- production, increasing IL-10 production, further reducing hepatic apoptosis. It is generally known that adequate and appropriate exercise is essential to maintain health and release work pressure. Some studies have
demonstrated that regular exercise can enhance immunological and antioxidant functions [3] [13] [25]. Regular exercise induces anti-inflammatory actions. During exercise, IL-6 is produced by muscle fibres. IL-6 stimulates the appearance in the circulation of other anti-inflammatory cytokines such as IL-10 and inhibits the production of the
pro-inflammatory cytokine TNF- [31].
Interestedly, exercise decreased the protective effect of high dose C. sinensis on liver damage induced by sepsis. AST value in mice received 40 mg C. sinensis alone were significantly decreased than saline of non-exercise group. However, ASL level in 40 mg C. sinensis with exercise increased significantly to the same level of septic mice. The cleaved-caspase-3, 6 and cleaved-PARP protein levels in liver tissue of 40 mg C. sinensis with exercise group were also markedly increased. Because the TNF- level in serum of 40 mg C. sinensis with exercise group was significantly increased than 40 mg C. sinensis alone.
The data suggested that higher caspases activation by exercise on high dose CS in liver might due to TNF-α induction. The TNF- cleaved-caspase-3, 6 and cleaved-PARP protein levels was no difference between 20 mg C. sinensis with exercise group and 20 mg C. sinensis without exercise group. It implicated that the reverse effect of exercise on 20 mg C.
sinensis is not obvious.
The controversial results of exercise combine C. sinensis showed that physical exercise is new host factor with impact on hepatic drug metabolism. Moderate to heavy physical exercise for a few hours reduces liver blood flow leading to a decreased elimination of drugs
exhibiting flow-limited metabolism [10]. The moderate physical exercise can prolong the half-life of indocyanine green for a few minutes, and the amount of prolongation increases with the duration of physical exercise. A reduction of liver blood flow amounting to 60% has been reported [34]. The physical exercise might change the metabolism of C. sinensis in liver and may lead to changes in drug efficacy and drug toxicity.
In summary, high dose C. sinensis treatment alone and precondition exercise training alone reduced the tissue damage induced by sepsis. The reverse effect of exercise on high dose C. sinensis in liver might due to high expression of TNF- and nitric oxide accumulation, and caspases activation.
REFERENCES
[1] Bruunsgaard H and Pedersen BK. Effects of exercise on the immune system in the elderly population. Immunol Cell Biol. 78(5):523-31, 2000.
[2] Cai B, Deitch EA, Ulloa L. Novel insights for systemic inflammation in sepsis and hemorrhage. Mediators Inflamm. 2010:642462,2010.
[3] Chen HI, Hsieh SY, Yang FL, Hsu YH, Lin CC. Exercise Training Attenuates Septic Responses in Conscious Rats. Med. Sci. Sports Exerc. Vol. 39, No. 3, 435–442, 2007. [4] Cauwels A. Nitric oxide in shock. Kidney Int.72 (5):557-65, 2007.
[5] Cauwels A, Brouckaert, P. Nitrite regulation of shock. Cardiovasc Res. 89(3):553-9, 2011.
[6] Chang KC, Unsinger J, Davis CG, Schwulst SJ, Muenzer JT, Strasser A, Hotchkiss RS. Multiple triggers of cell death in sepsis: death receptor and mitochondrial-mediated apoptosis. FASEB J. 21(3):708-19, 2007.
[7] Chen J, Zhang W, Lu T, Li J, Zheng Y, Kong L. Morphological and genetic
characterization of a cultivated Cordyceps sinensis fungus and its polysaccharide component possessing antioxidant property in H22 tumor-bearing mice. Life Sci. 78(23): 2742-8, 2006. [8] Dhainaut JF, Marin N, Mignon A, Vinsonneau C: Hepatic response to sepsis: interaction between coagulation and inflammatory processes. Crit Care Med 29:S42YS47, 2001.
[9] Dong CH, Yao YJ. In vitro evaluation of antioxidant activities of aqueous extracts from natural and cultured mycelia of Cordyceps sinensis. LWT: Food Sci. Technol. 2007.
[10] Døssing M. Effect of acute and chronic exercise on hepatic drug metabolism. Clin Pharmacokinet. 10(5):426-31, 1985.
[11] Feihl F, Waeber B, Liaudet L. Is nitric oxide overproduction the target of choice for the management of septic shock? Pharmacol Ther. 91(3):179-213, 2001.
Dis. 35(9): 585-92, 2003.
[13] Ji LL. Modulation of skeletal muscle antioxidant defense by exercise: Role of redox signaling. Free Radic Biol Med. 44(2):142-52, 2008.
[14] Kim KM, Kwon YG, Chung HT, Yun YG, Pae HO, Han JA, Ha KS, Kim TW, Kim YM. Methanol extract of Cordyceps pruinosa inhibits in vitro and in vivo inflammatory mediators by suppressing NF-kappaB activation. Toxicol. Appl. Pharmacol. 190: 1–8, 2003.
[15] Kuo CF, Chen CC, Lin CF, Jan MS, Huang RY, Luo YH, Chuang WJ.
Abrogation of streptococcal pyrogenic exotoxin B-mediated suppression of phagocytosis in U937 cells by Cordyceps sinensis mycelium via production of cytokines. FoodChem. Toxicol. 45, 278–285, 2007.
[16] Kuo HC, Su YL, Yang HL, Huang IC, Chen TY. Differentiation of Cordyceps sinensis by a PCR-single-stranded conformation polymorphism-based method and characterization of the fermented products in Taiwan. Food Biotechnol. 20, 161–170, 2006.
[17] Kuo CF, Chen CC, Luo YH, Huang RY, Chuang WJ, Sheu CC, Lin YS. Cordyceps sinensis mycelium protects mice from group A streptococcal infection. J. Med. Microbiol. 54, 795–802, 2005.
[18] Keller C, Keller P, Giralt M, Hidalgo J, and Pedersen BK. Exercise normalises overexpression of TNF-alpha in knockout mice. Biochem Biophys Res Commun 321: 179–182, 2004.
[19] Kuo YC, Lin CY, Tsai WJ, Wu CL, Chen CF, Shiao MS. Growth inhibitors against tumor cells in Cordyceps sinensis other than cordycepin and polysaccharides. Cancer Invest. 12, 611–615, 1994.
[20] Li SP, Yang FQ, Tsim KW. Quality control of Cordyceps sinensis, a valued traditional Chinese medicine. J Pharm Biomed Anal. 28; 41(5):1571-84, 2006.
[21] Liu C, Lu S, Ji MR. Effects of Cordyceps sinensis (CS) on in vitro natural killer cells. Zhongguo Zhong Xi Yi Jie He Za Zhi 12: 267–269, 259, 1992.
[22] Liu P, Zhu J, Huang Y, Liu C. Influence of Cordyceps sinensis (Berk.) Sacc. and rat serum containing same medicine on IL-1, IFN and TNF produced by rat Kupffer cells. Zhongguo Zhong Yao Za Zhi 21: 367–369, 384, 1996.
[23] Liu YK, Shen W. Inhibitive effect of Cordyceps sinensis on experimental hepatic fibrosis and its possible mechanism. World J. Gastroenterol. 9: 529–533, 2003.
[24] Marsh, C. B., & Wewers, M.D. The pathogenesis of sepsis. Factors that modulate the response to gram-negative bacterial infection. Clin Chest Med 17, 183– 197, 1996.
[25] Mehanna A, Vitorino DC, Panis C, Blanco EE, Pinge-Filho P, Martins-Pinge MC. Cardiovascular and pulmonary effects of NOS inhibition in endotoxemic conscious rats subjected to swimming training. Life Sci.81:1301–1308, 2007.
[26] Mackinnon LT. Current challenges and future expectations in exercise immunology: back to the future. Med. Sci. Sports Exerc. 26:191–194, 1994.
T. Apoptotic cell death in the response of D-galactosamine-sensitized mice to
lipopolysaccharide as an experimental endotoxic shock model. Infect.Immun. 64(3): 734-8, 1996.
[28] Nieman DC, Henson DA, Gusewitch G, Warren BJ, Dotson RC, Butterworth DE, Nehlsen-Cannarella SL. Physical activity and immune function in elderly women. Med. Sci. Sports Exerc. 25:823–831, 1993.
[29] O’Sullivan AW, Wang JH, Redmond HP. NF- B and p38 MAPK inhibition improve survival in endotoxin shock and in a cecal ligation and puncture model of sepsis in
combination with antibiotic therapy. J .Surg. Res. 152(1):46-53, 2009.
[30] Parameswaran N, Patial S. Tumor necrosis factor-α signaling in macrophages. Crit Rev Eukaryot Gene Expr. 20(2): 87-103, 2010.
[31] Pedersen BK. The anti-inflammatory effect of exercise: its role in diabetes and cardiovascular disease control. Essays Biochem; 42:105-17, 2006.
[32] Pieper AA, Verma A, Zhang J, Snyder SH. Poly (ADP-ribose) polymerase, nitric oxide and cell death. Trends Pharmacol Sci. 20(4):171-81. 1999.
[33] Rao YK, Fang SH, Tzeng YM. Evaluation of the anti-inflammatory and anti-proliferation tumoral cells activities of Antrodia camphorata, Cordyceps sinensis, and Cinnamomum
osmophloeum bark extracts. J Ethnopharmacol.114 (1):78-85. 2007.
[34] Rowell LB. Human cardiovascular adjustments to exercise and thermal stress. Physiol Rev. 54(1):75-159, 1974.
[35] Russell JA. Management of sepsis. N Engl J Med 355:1699-1713, 2006.
[36] Sakaguchi S and Furusawa S. Preventive Effects of a Traditional Chinese Medicine (Sho-saiko-to) on Endotoxin-Induced Cytotoxicity and Tumor Necrosis Factor- Production in J774A.1 Cells. Biol. Pharm. Bull. 27(9): 1468-1470, 2004.
[37] Sakaguchi S and Furusawa S. Oxidative stress and septic shock: metabolic aspects of oxygen derived free radicals generated in the liver during endotoxemia. FEMS Immunol Med Microbiol. 47(2):167-77, 2006.
[38] Shahed AR, Kim SI, Shoskes DA. Down-regulation of apoptotic and inflammatory genes by Cordyceps sinensis extract in rat kidney following ischemia/reperfusion. Transplant. Proc. 33: 2986–2987, 2001.
[39] Starkie R, Ostrowski SR, Jauffred S, Febbraio M, Pedersen BK. Exercise and IL-6 infusion inhibit endotoxin induced TNF-alpha production in humans. FASEB J. 17:884– 886, 2003.
[40] Ulich TP, Guo K, del Castillo J: Endotoxin-induced cytokine gene expression in vivo. I. Expression of tumor necrosis factor mRNA in visceral organs under physiologic conditions and during endotoxemia. Am J Pathol. 134:11–14, 1989.
[41] Wu JY, Zhang QX, Leung PH. Inhibitory effects of ethyl acetate extract of Cordyceps sinensis mycelium on various cancer cells in culture and B16 melanoma in C57BL/6 mice. Phytomedicine .14: 43–49, 2007.
[42] Wong F, Bernardi M, Balk R, Christman B, Moreau R, Garcia-Tsao G, Patch D, Soriano G, Hoefs J, Navasa M. International Ascites Club. Sepsis in cirrhosis: report on the 7th meeting of the International Ascites Club. Gut. 54(5):718-25, 2005.
[43] Zhou X, Luo L, Dressel W, Shadier G, Krumbiegel D, Schmidtke P, Zepp F, Meyer CU. Cordycepin is an immunoregulatory active ingredient of Cordyceps sinensis. Am J Chin Med. 36(5):967-80,2008.
[44] Zhao Q, Wang X, Nelin LD, Yao Y, Matta R, Manson ME, Baliga RS, Meng X, Smith CV, Bauer JA, Chang CH, Liu Y. MAP kinase phosphatase 1 controls innate immune responses and suppresses endotoxic shock. J Exp Med. 203(1):131-40, 2006.