Chao Wei Yang & Ching Huei Kao
∗Department of Agronomy, National Taiwan University, Taipei, Taiwan, Republic of China (∗author for correspondence)
Received 10 June 1999; accepted in revised form 25 August 1999
Key words: ammonium, methionine sulfoximine, Oryza sativa, proline accumulation, water stress
Abstract
Ammonium accumulation in relation to proline accumulation in detached rice leaves under stress conditions was investigated. Ammonium accumulation in dark-treated detached rice leaves preceded proline accumulation. Ammonium accumulation caused by water stress coincided closely with proline accumulation in detached rice leaves. Exogenous NH4Cl and methionine sulfoximine (MSO), which caused an accumulation of ammonium in
detached rice leaves, increased proline content. It was found that proline in NH4Cl- or MSO-treated rice leaves
is less utilized than in water-treated rice leaves (controls). These results are in agreement with the observation that a decrease in proline utilization contributes to the accumulation of proline in dark-treated and water stressed rice leaves. Although ammonium content increased in Cd- and Cu-treated rice leaves, the increase in ammonium content was only observed after the increase in proline content.
Abbreviations: MSO – methionine sulfoximine; WS – water stress
1. Introduction
Proline has been shown to accumulate in plants sub-jected to water stress and exposed to excess Cd and excess Cu [1, 2, 9, 12, 26, 27, 29, 32]. Previously, we have reported that proline accumulated in detached rice leaves during dark-induced senescence [10, 31].
The ammonium ion is a central intermediate in the metabolism of nitrogen in plants. Glutamine syn-thetase plays a crucial role in the assimilation of ammonium [16]. It has been shown that water stress results in a decline in the activity of glutamine syn-thetase in leaves [4, 18]. Glutamine synsyn-thetase activity is also known to decrease during dark-induced senes-cence of leaves [7, 13, 20, 28]. We have demonstrated that excess Cu decreased the activity of glutamine synthetase in detached rice leaves [7]. A decline in glutamine synthetase activity in leaves during water stress, dark-induced senescence or when exposed to heavy metals may result, at least in part, in an accu-mulation of ammonium in leaves. In fact, ammonium has been shown to accumulate in leaves subjected to
water stress, when exposed to excess Cu, and during dark-induced senescence [7, 8, 18, 22, 30].
Accumulation of putrescine has been reported in response to environmental stresses [6]. Several lines of evidence suggest that putrescine biosynthesis is linked to the assimilation of ammonium nutrition in both stressed and nonstressed plant tissues. Le Rudulier and Goas [15] showed that ammonium nutrition pro-duced higher tissue putrescine content than did nitrate feeding. Similarly, Klein et al. [14] reported that K+-deficient peas utilizing ammonium as a nitrogen source accumulated putrescine to a greater extent than those maintained on nitrate. Rabe and Lovatt [24] have demonstrated that increased ammonium content in PO4-deficient citrus are accompanied by increased
arginine biosynthesis. Rabe [23] suggested that putres-cine and arginine accumulating during environmental stress conditions might serve as an ammonium detox-ification mechanism. By extension, it seems likely that stress-induced accumulation of proline could rep-resent a manifestation of such an ammonium detox-ification mechanism. To understand the metabolic
140
Figure 1. Influence of NH4Cl on ammonium and proline contents in detached rice leaves. All measurements were made 12 h after treatment in the light. Vertical bars represent standard errors (n = 4).
and cellular phenomena that lead to proline accu-mulation, in detached rice leaves induced by water stress, heavy metals and during dark-induced senes-cence, we have tried to determine the relation, if any, between ammonium and proline accumulations. This paper reports more detailed time-course meas-urements along with some dose-response experiments, designed to describe the relation between ammonium and proline accumulations in detached rice leaves.
2. Materials and methods
Rice (Oryza sativa cv. Taichung Native 1) was cul-tured on a stainless net floating on half-stength John-son’s modified nutrient solution (pH 4.2) in a 500 ml beaker [11]. The nutrient solution was replaced every three days. Rice seedlings were grown for 12 d in a greenhouse, where natural light was provided and
Figure 2. Influence of MSO on ammonium and proline contents
in detached rice leaves. All measurements were made 12 h after treatment in the light. Vertical bars represent standard errors (n = 4).
the temperature was controlled at 30◦C during the day and at 25◦C at night. The apical 3 cm segments excised from the third leaves of 12-d-old seedlings were used. A group of 10 segments was floated in a Petri dish containing 10 ml of test solutions or water (controls). Incubation was carried out at 27◦C in light (40 µmol quanta m−2 s−1) or in darkness. Water stress was applied by a previously described method [12]. Briefly, leaf segments were exposed to the vapour above a solution of 0.5 M NaCl to decrease air humidity.
Proline was extracted and its concentration deter-mined by the method of Bates et al. [3]. Leaf segments were homogenized with 3% sulfosalicylic acid and the homogenate was centrifuged. The supernatant was treated with acetic acid and acid ninhydrin, boiled for
Figure 3. Time courses of proline and ammonium contents in
detached rice leaves incubated in distilled water under dark and light conditions. Vertical bars represent standard errors (n = 4).
1 h and then absorbance at 520 nm was determined. Amounts of proline are expressed as µmol g−1initial fresh weight.
Ammonium was extracted by homogenizing leaf segments in 0.3 mM sulphuric acid (pH 3.5). The homogenate was centrifuged for 10 min at 39,000 g and the supernatant was used for determination of ammonium as described previously [17]. Ammonium content was expressed as µmol g−1 initial fresh weight.
3. Results and discussion
Figure 1 shows the effect of NH4Cl on ammonium
and proline contents in detached rice leaves in the
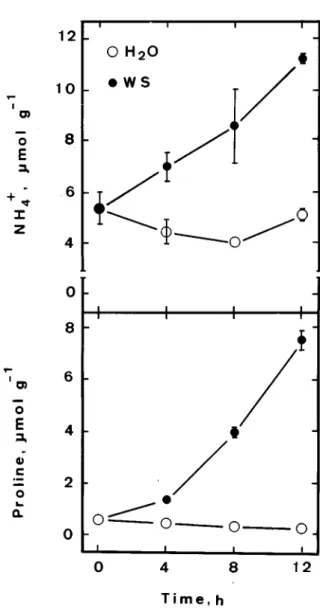
Figure 4. Time courses of proline and ammonium contents in
detached rice leaves incubated in distilled water and water stress (WS) conditions. Vertical bars represent standard errors (n = 4).
light. It is clear that the increase in the ammonium content in NH4Cl-treated detached rice leaves was
associated with the increase in proline content. MSO is a structural analogue of glutamate, and serves as an irreversible inhibitor of glutamine synthetase [25]. There is evidence that the addition of MSO results in an accumulation of ammonium [5, 21]. To char-acterize further the relationship between ammonium and proline contents in detached rice leaves, detached rice leaves were incubated in the presence of vari-ous concentrations of MSO. As indicated in Figure 2, MSO increased endogenous ammonium and proline contents.
142
Table 1. Proline content in ornithine-pretreated detached
rice leaves incubated in distilled water, NH4Cl and MSO
Treatment Proline, µmol g−1
Orn, 3h 18.7± 1.3
Orn, 3h→ H2O, 8h 5.7± 0.4 Orn, 3h→ NH4Cl, 8h 10.6± 0.7 Orn, 3h→ MSO, 8h 9.1± 0.7
Detached rice leaves were pretreated with 50 mM ornithine (Orn) for 3 h in the light and then incubated in distilled water, NH4Cl (200 mM), or MSO (1 mM) for 8 h in the light. Means± SE (n = 4).
Figure 3 shows the change of proline content in detached rice leaves incubated in distilled water in darkness and in the light. In darkness, the amount of proline in detached rice leaves did not increase until 12 h after incubation, but increased about 2-, 2.5-and 6-fold at 12, 48, 2.5-and 72 h, respectively, which confirmed our previous results [10, 31]. However, the amount of proline in detached rice leaves, incubated in the light remained relatively unchanged during 48 h, but subsequently increased slightly. In detached rice leaves, it is apparent that proline accumulates more in darkness than in the light. It is expected that the ammonium content is greater in darkness than in the light. As indicated in Figure 3, this is indeed the case. An increase in the ammonium content of dark-treated detached rice leaves was found to precede the increase in proline contents.
It has long been recognized that water stress is effective in increasing proline content in detached rice leaves [12]. The effect of water stress on proline con-tent in relation to ammonium concon-tent is presented in Figure 4. Water stress effectively increased proline and ammonium content.
Recently, we have shown that a decrease in proline utilization contributes to the accumulation of proline in dark-treated and water stressed rice leaves [33]. If ammonium accumulation is indeed important in regu-lating dark- and water stress-induced proline accu-mulation in detached rice leaves, NH4Cl or MSO is
expected to decrease proline utilization. To test this possibility, detached rice leaves were pretreated with 50 mM ornithine for 3 h to increase the endogenous proline content and then transferred to NH4Cl or MSO
for 8 h. Proline content was then determined. As indi-cated in Table 1, proline content in NH4Cl- or
MSO-treated rice leaves was greater than in water-MSO-treated control rice leaves, suggesting that proline in NH4
Cl-or MSO-treated rice leaves is less utilized than in
con-Figure 5. Time courses of proline and ammonium contents in
detached rice leaves incubated in distilled water, CdCl2(5 mM) and CuSO4(10 mM). Vertical bars represent standard errors (n = 4).
trols. These results supported further that ammonium accumulation in dark-treated and water stressed rice leaves is associated with proline accumulation.
Proline has been shown to accumulate in plants exposed to excess Cd or Cu [2, 9, 26, 27, 32]. In detached rice leaves, treatment of Cd or Cu also resul-ted in an accumulation of proline (Figure 5), However, proline accumulation in Cd- and Cu-treated detached rice leaves preceded ammonium accumulation (Fig-ure 5). It is apparent that ammonium accumulation is
deficiency and cold treatment showed increased ammonium content and increased rate of arginine biosynthesis in leaves exposed to stress [19, 23]. Susceptible plants had lower rates of arginine biosyn-thesis and higher ammonium content; tolerant plants had greater rates of arginine biosynthesis and lower ammonium content. These data suggest that there is a positive correlation between detoxification of ammonium, arginine biosynthesis and tolerance to several types of environmental stress. Similar findings were reported for studies conducted on alfalfa suspen-sion cultures subjected to osmotic shock as well as cucurbits exposed to salinity stress [19]. In the present investigation, we are able to show that ammonium accumulation in dark-treated and water stressed rice leaves is associated with proline accumulation. Future work should be focused on the correlation between conversion of ammonium into proline and tolerance to dark-induced senescence and water stressed in rice leaves.
Acknowledgement
This work was supported financially by the National Science Council of the Republic of China to CHK (NSC 89-2313-B-002-111).
References
1. Aspinall D and Paleg LG (1981) Proline accumulation: physiological aspects. In: Paleg LG and Aspinall D (eds) Physiology and Biochemistry of Drought Resistance in Plants, 205 pp. Sydney: Academic Press
2. Bassi R and Sharma SS (1993) Changes in proline content accompanying the uptake of zinc and copper by Lemna minor. Ann Bot 72: 151–154
3. Bates LS, Waldren RP and Teare ID (1973) Rapid determin-ation of free proline for water stress studies. Plant Soil 39: 205–207
4. Becker TW, Hoppe M and Foch PH (1986) Evidence for the participation of dissimilatory processes in maintaining high carbon fluxes through the photosynthetic carbon reduction and oxidation cycle in water-stressed bean leaves. Photosynthetica 20: 153–157
5. Berger MG and Fock HP (1983) Effect of methionine sulfox-imine and glycine on nitrogen metabolism of maize leaves in the light. Aust J Plant Physiol 10: 187–194
6. Bouchereau A, Aziz A, Larher F and Martin-Tanguy J (1999) Polyamines and environmental challenges: recent develop-ment. Plant Sci 140: 103–125
tion is associated with senescence of rice leaves. Plant Growth Regul 21: 195–201
9. Chen SL and Kao CH (1995) Cd induced changes in proline content and peroxidase activity in roots of rice seedlings. Plant Growth Regul 17: 67–71
10. Chou IT, Chen CT and Kao CH (1990) Regulation of proline accumulation in detached rice leaves. Plant Sci 70: 43–48 11. Kao CH (1980) Senescence of rice leaves IV. Influence of
ben-zyladenine on chlorophyll degradation. Plant Cell Physiol 21: 1255–1262
12. Kao CH (1981) Senescence of rice leaves VI. Comparative study of the metabolic changes of senescing and turgid and water-stressed excised leaves. Plant Cell Physiol 22: 683– 688
13. Kar M and Feierabend J (1984) Changes in the activities of enzymes involved in amino acid metabolism during the senescence of detached wheat leaves. Physiol Plant 62: 39– 44
14. Klein H, Priebe A and Jager HJ (1979) Putrescine and sper-midine in peas: effects of nitrogen source and potassium supply. Physiol Plant 45: 497–499
15. Le Ruduller D and Goas G (1971) Mise en evidence et dosage de guelgues amines dans les plantules de Soja hispida Moench. Privees de lenrs cotyledons et cultivees en presence de nitrates, d’uree et de chlorure d’ammonium. CR Acad Sci (Paris) Ser D 273: 1108–1111
16. Lea DJ and Miflin BJ (1974) Alternative route for nitrogen assimilation in higher plants. Nature 251: 614–616
17. Lin CC and Kao CH (1996) Disturbed ammonium assimilation is associated with growth inhibition of roots in rice seedlings caused by NaCl. Plant Growth Regul 18: 233–238
18. Lin J-N and Kao CH (1998) Water stress, ammonium, and leaf senescence in detached rice leaves. Plant Growth Regul 26: 165–169
19. Lovatt CJ (1991) Stress alters ammonia and arginine meta-bolism. In: Flores HE, Arteca RN and Shannon JC (eds) Polyamines and Ethylene: Biochemistry, Physiology, and Interactions, pp 166–179. Maryland: American Society of Plant Physiologists
20. Peeters KMU and Van Laere AJ (1992) Ammonium and amino acid metabolism in excised leaves of wheat (Triticum
aestivum) senescing in the dark. Physiol Plant 84: 243–249
21. Platt SG and Anthon GE (1981) Ammonium accumulation and inhibition of photosynthesis in methionine sulfoximine treated spinach. Plant Physiol 67: 509–513
22. Postitus C and Jaccobi G (1976) Dark starvation and plant metabolism. VI. Biosynthesis of glutamic acid dehydrogenase in detached leaves of Cucurbita maxima. Z Pflanzenphysiol 78: 133–140
23. Rabe E (1990) Stress physiology: the functional significance of the accumulation of nitrogen containing compounds. J Hort Sci 65: 231–243
24. Rabe E and Lovatt CJ (1986) Increased arginine biosynthesis during phosphorus deficiency. Plant Physiol 81: 774–779 25. Ronzio RA, Rowe WB and Meister A (1969) Studied on the
mechanism of inhibitor of glutamine synthetase by methionine sulfoximine. Biochemitry 8: 1061–1075
26. Saradhi A and Saradhi PP (1981) Proline accumulation under heavy metal stress. J Plant Physiol 138: 554–558
144
27. Schat H, Sharma SS and Vooijs R (1997) Heavy metal-induced accumulation of free proline in a metal-tolerant and a nontolerant ecotype of Silene vulgaris. Physiol Plant 101: 477–482
28. Simpson RJ and Dalling MJ (1981) Nitrogen redistribution during grain growth in wheat (Triticum aestivum L.). III. Enzymology and transport of amino acids from senescing flag leaves. Planta 151: 447–456
29. Singh TN, Paleg LG and Aspinall D (1973) Stress metabolism I. Nitrogen metabolism and growth in the barley plant during water stress. Aust J Biol Sci 26: 45–56
30. Thomas H (1978) Enzymes of nitrogen mobilization in detached leaves of Lolinm temulentum during senescence. Planta 142: 161–169
31. Wang CY, Cheng SH and Kao CH (1982) Senescence of rice leaves VII. Proline accumulation in senescing excised leaves. Plant Physiol 69: 1348–1349
32. Wu J-T, Chang SJ and Chou T-L (1995) Intracellular proline accumulation in some algae exposed to copper and cadmium. Bot Bull Acad Sin 36: 69–93
33. Yang C-W, Lin CC and Kao CH (1999) Endogenous ornithine and arginine contents and dark-induced proline accumulation in detached rice leaves. J Plant Physiol (in press)