Appl Microbiol Biotechnol (2006) 71: 238–245 DOI 10.1007/s00253-005-0145-0
A P P L I E D M I C R O B I A L A N D C E L L P H Y S I O L O G Y
Chiu-Hsia Chiu . Tzu-Yu Lu . Yun-Yu Tseng . Tzu-Ming Pan
The effects of
Lactobacillus-fermented milk on lipid metabolism
in hamsters fed on high-cholesterol diet
Received: 28 June 2005 / Revised: 13 August 2005 / Accepted: 18 August 2005 / Published online: 20 September 2005 # Springer-Verlag 2005
Abstract The objective of this study was to evaluate the effects of local Lactobacillus strains (NTU 101 and 102) on cholesterol-lowering effects in vivo. Thirty male hamsters were housed, divided into five groups, and fed on a cho-lesterol diet (5 g/kg diet) to induce hyperchocho-lesterolemia. Milk fermented by Lactobacillus paracasei subsp. para-casei NTU 101, Lactobacillus plantarum NTU 102, and Lactobacillus acidophilus BCRC 17010 was administrated for this study. After treatment with different fermented milk, blood was taken and liver was removed for the de-termination of lipoproteins, including total cholesterol, high-density lipoprotein cholesterol (HDL-C), low-density lipoprotein cholesterol (LDL-C), and triglyceride. Lacto-bacilli and bifidobacteria decreased (105) in the control group; when hamsters were fed on fermented milk, the number of lactobacilli (107–108) and bifidobacteria (105– 107) was increased. Serum and liver total cholesterol levels were significantly reduced by about 26.4, 23.5, and 30.1% and by about 17.7, 15.9, and 13.4% when hamsters were given fermented milk. However, serum HDL-C and LDL-C were also reduced. The results of this study showed that the hypocholesterolemic effect of local Lactobacillus strains was attributed to its ability to lower serum and liver total cholesterol levels. Thus, local Lactobacillus strains could significantly increase probiotic count.
Introduction
Dietary habit change toward lipid-rich foods in recent years has resulted in risks for cardiovascular diseases. Serum cholesterol level is commonly recognized as an important factor in disease development. Since Mann and Spoerry (1974) discovered hypocholesterolemic effects arising from the diet of the Massai tribespeople in Africa, who ingested large intakes of milk fermented by Lactobacillus strains, the relationship between lactic acid bacteria or other pro-biotics and cholesterol concentration in serum has become a focus of great interest. Harrison and Peat (1975) reported a reduction in serum cholesterol level in newborns fed on fresh milk when the fecal titer of Lactobacillus acidophilus was raised. A reduction in serum cholesterol level was also associated with yogurt intake in rabbits (Thakur and Jha
1981), and humans (Hepner et al.1979; Keim et al.1981) demonstrated that intestinal lactic acid bacteria, such as L. acidophilus, caused bile salts to deconjugate and coprecip-itate with cholesterol under anaerobic conditions.
In addition to L. acidophilus, Nielson and Gilliland (1985) also showed the cholesterol-reducing activity of Lactobacillus casei. Gilliland et al. (1985) reported that consumption of L. acidophilus RP32, which grew on bile and assimilated cholesterol from a laboratory medium, significantly inhibited increases in the serum cholesterol level of pigs fed on a high-cholesterol diet. Fukushima and Nakao (1995) also reported that a probiotic mixture of Bacillus subtilis, Bacillus natto, Bacillus megaterium, L. acidophilus, Lactobacillus plantarum, Lactobacillus bre-vis, L. casei, Streptococcus faecalis, Streptococcus lactic, Streptococcus thermophilus, Saccharomyces cerevisiae, and Candida utilis at 107–108 CFU/g rice bran showed lower levels of total cholesterol, low-density lipoprotein, and liver cholesterol in hypercholesterolemic rats. Brashears et al. (1998) found that in vitro cultivation of L. casei strains without pH control could reach the maximal amount of removed cholesterol and that L. acidophilus removed the most cholesterol by incorporating the compound into cel-lular membranes. L. casei mainly did this by destabilizing C.-H. Chiu . T.-Y. Lu . Y.-Y. Tseng . T.-M. Pan (*)
Institute of Microbiology and Biochemistry, National Taiwan University,
No. 1, Section 4, Roosevelt Road, Taipei, 10617, Taiwan
e-mail: [email protected] Tel.: +886-2-33664519 Fax: +886-2-23627044 C.-H. Chiu
National Pingtung University of Science and Technology, Pingtung, 91207, Taiwan
cholesterol micelles and coprecipitating cholesterol with deconjugated bile salts at pH <6.0.
In addition, some researchers investigated the in vitro cholesterol-reducing ability of bifidobacteria strains and found that Bifidobacterium bifidum was the only strain so far expressing an obvious outcome of cholesterol elimina-tion equivalent to that of L. acidophilus. The main purposes of this study are: (1) to explore the relationship of three Lactobacillus strains (Lactobacillus paracasei subsp. par-acasei NTU 101, L. plantarum NTU 102, and L. acid-ophilus BCRC 17010) with serum cholesterol level in hamsters and their effects on intestinal microflora; and (2) to reduce serum cholesterol level and improve intestinal microflora in host animals with these probiotics.
Materials and methods
Bacterial strains
The bacteria strains used in this study, which were effective in cholesterol reduction in in vitro trials, were two local strains isolated in our laboratory and in type culture (L. acidophilus BCRC 17010). These local strains were L. paracasei subsp. paracasei NTU 101 (which was isolated from human infant feces and showed good survival at low pH, tolerance to high bile concentration, and ability to reduce serum cholesterol in vitro) (Lin et al.2004) and L. plantarum NTU 102 (which was isolated from home-made Korean-style cabbage pickles, was able to survive in vitro at low pH and in the presence of bile salt, and demonstrated pathogen inhibition activities, especially against Pseudomo-nas aeruginosa) (Pan et al. 2002). Growth media included Lactobacilli MRS Broth (Difco Laboratories, Detroit, MI, USA) for the aforementioned strains and Bifidobacteria Iodoacetate Medium-25 (BIM-25) for Bifidobacterium spp.
Animal feeding and grouping
Thirty male Syrian hamsters, 4 weeks of age and weighing about 70 g (mean), were randomly divided into five groups with six members each and fed on high-cholesterol diet and ordinary water supply for 1 month. Then, water supply was replaced with a variety of drinking substitutes: group A, water only; group B, sterilized milk; group C, milk fer-mented by L. paracasei subsp. paracasei NTU 101; group D, milk fermented by L. plantarum NTU 102; and group E, milk fermented by L. acidophilus BCRC 17010. During feeding time, environmental conditions were well con-trolled; relative humidity was 60% and room temperature was 20–25°C, with 12-h light exposure in a daily cycle from 6 a.m. to 6 p.m. Food and liquid were accessible at all times and were replenished everyday. The animals were fed for 8 weeks, during which time body weight and food intake were recorded. After the feeding period, the animals were not fed overnight and were presented for further tests. The feeding material in this study was mainly AIN-76, supplemented with 5% cholesterol and 0.3% bile salt. The
diet formula is shown in Table 1. The experiments were carried out in a qualified animal breeding room in the animal center at our institute. (The protocol complied with guidelines described in the “Animal Protection Law,” amended on 17 January 2001, Hua-Zong-(1)-Yi-Tzi-900000 7530, Council of Agriculture, Executive Yuan, Taiwan.)
Preparation of fermented and nonfermented milk
Fermented milk Skim milk powder was weighed and dissolved in water to constitute 4% skim milk (wt/vol), which was then sterilized using an autoclave at a tem-perature of 121°C and a pressure of 1.2 kg/cm2for 15 min and then cooled to room temperature. The milk was inoculated by adding a 1% bacterial solution (vol/vol) in a lamina flow cabinet and by incubating at 37°C for 18 h. After adding 0.2% carboxylmethyl cellulose as a stabilizer, the fermented milk was homogenized by a blender and fed to the experimental hamsters.
Nonfermented milk Four percent skim milk was prepared, sterilized, and cooled to room temperature, as described above, and then fed to the animals.
Enzymatic kits
Enzymatic kits used to quantify the levels of serum cholesterol, high-density lipoprotein cholesterol (HDL-C), low-density lipoprotein cholesterol (LDL-C), and triglyc-eride (TG) were as follows: cholesterol, 1.14830.0001; HDL-C, 1.14210.0001; LDL-C, 1.14992.0001; and TG glycerol phosphate oxidase –phenylperoxidaseaminophe-nozonphenol (GPO-PAP), 1.14856.0001 (Merck, Darm-stadt, Germany).
Blood lipid analysis
The hamsters were euthanized using CO2. Blood
speci-mens were taken with syringe from the celiac vein and transferred to nonheparinized vacuum blood collection tubes. The tubes were held stationary until the blood
Table 1 The composition of high-cholesterol diets
a
Based on AIN-76 formula (American Institute of Nutrition 1977) Ingredient Content (g/kg) Casein 200 Safflower oil 100 Vitamina 10 Minerala 35 Choline chloride 2 Sodium cholate 1 Cellulose 20 Sucrose 579 Methionine 3 Cholesterol 50
obviously appeared in two layers and were then centrifuged at 1,750×g for 15 min. The supernatant was taken and stored in a refrigerator at 4°C for later tests.
Measurement of serum cholesterol, HDL-C, and LDL-C
Cholesterol oxidase–phenylperoxidaseaminophenozonphenol (CHOD-PAP) method was used to measure cholesterol levels in blood specimens (Richmond 1973), whereas GPO-PAP was used to measure TG levels in blood speci-mens (Bucolo and David1973; Fossati and Prencipe1982; McGowan et al.1983).
Measurement of liver cholesterol
After an animal had been killed, the viscera were opened and the liver was removed, rinsed with saline, blotted dry with filter paper, and weighed. A piece of liver tissue weighing about 1 g was placed in a sample bottle, to which Folch solution (chloroform:methanol=2:1; vol/vol) 20 times the tissue volume was then added. After the liver tissue in the sample bottle had been homogenized, the mixture was agitated for 30 min in an orbital shaker at room temperature to facilitate lipid extraction. The homogenate was then filtered with Whatman No. 2 filter paper, quan-tified, and stored in a freezer at −20°C for later use. The CHOD-PAP method was used to analyze samples. One hundred microliters of liver extractant in Folch solution was taken and dried with nitrogen, and 1 ml of chromogemic reagent was added in a 37°C water bath. The reaction of cholesterol in the specimen with cholesterol lipase and cholesterol oxidase produced 4-(p-benzo-quinone-mono-imine) in red. The observed A550was referred to a standard
curve to calculate specimen cholesterol concentration.
Analysis of intestinal microflora
After the feeding period had been completed, the animals were fasted for 12 h and then killed. Once the viscera had been opened and the blood had been collected, the cecum (including a small portion of adjacent colon tissue) of each animal was removed and placed in a capped test tube, which was taken to a lamina flow cabinet where 1 g of cecum tissue was weighed, transferred into a tube with 9 ml of anaerobic diluent, and homogenized by vortexing. The homogenate was taken into a nitrogen-filled glove box where 1 ml of sample was transferred to a tube with 9 ml of anaerobic diluent, and the same procedure was repeated several times to perform a serial dilution. The sample diluents in appropriate dilution factors were added to, and pour-plated with, MRS agar and BIM-25. The plates were placed in anaerobic containers and incubated at 37°C for 48 h. The number of colonies counted after incubation represented the cell counts of Lactobacillus and Bifido-bacterium, respectively (Juang et al.2000).
Measurement of fecal water content
On the last day of the fourth and eighth weeks, feces were collected, packed in airtight bags, weighed, and stored in a freezer at−20°C during the feeding period. For fecal water content measurement, fecal samples that had been weighed were freeze-dried until constant weights have been reached within about 24 h (Juang et al. 2000). The calculation of fecal water content was as follows: fecal water content (%)= [(weight before freeze drying−weight after freeze drying)/ weight before freeze drying]×100%.
Statistical analysis
All data underwent duplicate analysis using one-way analysis of variance in a statistical analysis system. Duncan’s multiple range test was performed to compare any significant differences (p<0.05) in variables between groups.
Results
Growth of hamsters
In Table 2, all groups of hamsters show no significant differences in body weight gain, total food intake, and food efficiency (p>0.05). This indicates that the animals grow in similar patterns.
Blood lipid analysis
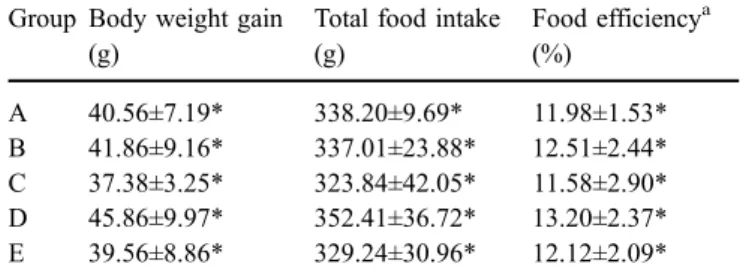
Figure 1 shows the effects of fermented milk diet con-taining Lactobacillus strains on serum cholesterol level. Fermented-milk-feeding groups (groups C, D, and E) displayed significantly lower serum cholesterol levels than those of group A (control) and group B (milk-feeding). In summary, the diet of L. acidophilus BCRC 17010 (group E) achieved the maximal cholesterol reduction of 30.1%, fol-lowed by L. paracasei subsp. paracasei NTU 101 (group C;
Table 2 Body weight gain, total food intake, and food efficiency of hamsters fed on high-cholesterol diet for 8 weeks
Group Body weight gain (g)
Total food intake (g) Food efficiencya (%) A 40.56±7.19* 338.20±9.69* 11.98±1.53* B 41.86±9.16* 337.01±23.88* 12.51±2.44* C 37.38±3.25* 323.84±42.05* 11.58±2.90* D 45.86±9.97* 352.41±36.72* 13.20±2.37* E 39.56±8.86* 329.24±30.96* 12.12±2.09*
A Control group; B high-cholesterol diet + milk; C high-cholesterol diet + fermented milk containing L. paracasei subsp. paracasei NTU 101; D high-cholesterol diet + fermented milk containing L. plantarum NTU 102; E high-cholesterol diet + fermented milk containing L. acidophilus BCRC 17010
*Significantly different at p<0.05
a
26.4%) and L. plantarum NTU 102 (group D; 23.5%). Although the diet of fresh milk (group B) also produced 8% cholesterol reduction, it was regarded as insignificant after statistical analysis.
The contents of other blood lipids (HDL-C, LDL-C, and TG) are shown in Table3. Group B expressed the highest HDL-C concentration, followed by group A and then by groups C, D, and E. The difference among the last three groups was not significant. LDL-C contents in fermented-milk-feeding animals (groups C, D, and E) were sig-nificantly lower than those in animals fed on a diet without lactic acid bacteria (groups A and B). Group E (fed on diet containing L. acidophilus BCRC 17010) expressed the lowest LDL-C content with a reduction of 47.4% compared to the control group, followed by group C (L. paracasei subsp. paracasei NTU 101) with 32.9% reduction and then by group D (L. plantarum NTU 102) with 27.8% reduction. The group ranking in blood TG level was: E < C < D < A < B. The difference between groups C, D, and E and groups A and D appeared small in statistical significance. How-ever, compared to group A, both groups C and E had an obvious reduction (p<0.05).
Liver lipid analysis
Table4shows data on liver weight and liver lipid content. Liver weight remained within a range with little significant change in the hamsters either fed or not fed on high-cholesterol diet. The milk-feeding and fermented-milk-feeding animals (groups B, C, D, and E) had a great reduction in liver cholesterol content compared to the control group (group A). The fermented-milk-feeding groups C, D, and E expressed lower cholesterol contents than milk-feeding group B. Group C had the lowest cholesterol content. The group ranking in liver cholesterol content was: A > B > E > D > C. The difference between two particular groups was significant (p<0.05). In liver TG content, group A expressed a level higher than those of groups B, C, D, and E. However, there existed no significant difference among the four experimental groups (p<0.05).
Analysis of intestinal microflora
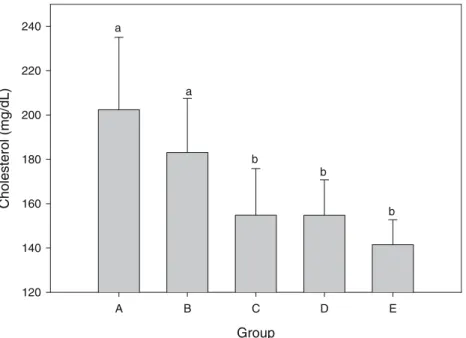
Figure2illustrates the cell counts of bacteria in the ceca of hamsters. In the cecum microflora, the cell numbers of lactic acid bacteria in groups C, D, and E (fed on
Group A B C D E Cholesterol (mg/dL) 120 140 160 180 200 220 240 a a b b b Fig. 1 Serum total cholesterol
concentration of hamsters fed on high-cholesterol diet. Data are presented as mean±SEM (n=6). A Control group; B cho-lesterol diet + milk; C high-cholesterol diet + fermented milk containing L. paracasei subsp. paracasei NTU 101; D high-cholesterol diet + fer-mented milk containing L. plantarum NTU 102; E high-cholesterol diet + fermented milk containing L. acidophilus BCRC 17010.
a,b
Values with different super-scripts are significantly different at p<0.05
Table 3 Serum HDL-C, LDL-C, and TG contents of hamsters fed on high-cholesterol diet Group HDL cholesterol (mg/dl) LDL cholesterol (mg/dl) TG (mg/dl) A 94.36±22.29b 80.34±20.50a 136.52±3.37b B 106.42±5.05a 77.56±17.67a 162.50±28.36a C 81.12±4.33c 53.93±13.17b 113.49±20.22c D 73.45±9.75c 58.00±10.21b 129.70±18.64b,c E 70.70±2.84c 42.26±6.97b 102.09±23.60c
A, B, C, D, and E conditions are the same as in Table2 a,b,c
p<0.05 is significantly different from A
Table 4 Hepatic cholesterol and TG in experimental hamsters
Group Liver weight (g) Cholesterol (mg/g) TG (mg/g)
A 6.23±0.79a 303.81±16.02a 109.50±17.14a
B 6.86±0.68a,b 295.44±13.48a,b 86.26±7.54b
C 6.63±0.59a,b 249.99±11.82c 87.81±18.81b
D 7.12±1.34a,b 255.61±12.73c 93.22±16.51b
E 6.31±0.72a 262.97±10.72b,c 99.62±23.93a,b
A, B, C, D, and E conditions are the same as in Table2 a,b,c
Lactobacillus-fermented milk) were greater than those in group B (fed on milk) and group A (fed on water) (p<0.05). Approximately, the cell numbers of lactic acid bacteria in the animals fed on fermented milk and in those fed on milk were 103 and 101 times the count in the control group, respectively. The difference was significant (p<0.05). The cell counts of Bifidobacterium in the cecum, ranging from 104to 106, showed no significant difference in the animals fed and not fed on fermented milk (p>0.05). However, the colony number of group C (fed on fermented milk con-taining L. paracasei subsp. paracasei NTU 101) was slightly greater than those of the other groups. The group ranking in colony numbers was: C > D > E > A > B.
Fecal water content
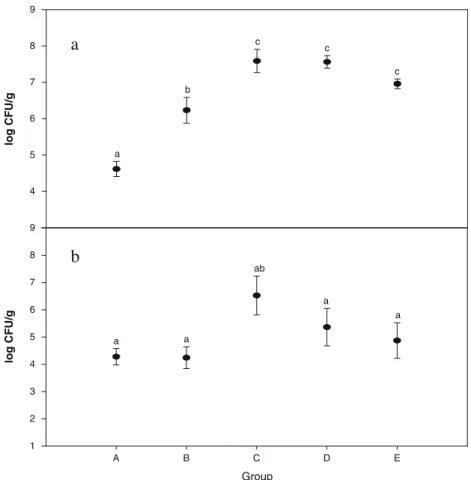
In the middle (after 1 month) and at the end (after 2 months) of the feeding period, animal feces were collected, freeze-dried, and weighed to calculate the fecal water content. The result is shown in Fig.3. Fecal water content can be used as an index of fecal elimination. Since the animals were fed on the same diet and water supply in the first feeding month, all five groups showed no significant difference in fecal water content (p>0.05). The fecal water content ranged between 15 and 30%, and the standard deviation (SD) of animals within the same group was low. At the end of the feeding period, as the supply of water was changed to a variety of substitutes, the fecal water content
varied in the five groups. The group ranking in fecal water content was: D > E > B > C > A. The difference between group D (fed on milk fermented by L. plantarum NTU 102) and control group A was statistically significant. Although the fecal water content still ranged between 15 and 30%, the SD of animals within one group increased. This indicated that the variation of the group members was enlarged.
Discussion
High concentrations of cholesterol and LDL-C in the blood account for increasing risk for cardiovascular diseases. According to Frick et al. (1987), every 1% reduction in body cholesterol content lowers the risk for cardiovascular diseases by 2%. A change in dietary habit, such as eating fermented products containing lactic acid bacteria, can lead to cholesterol reduction. In this study, hamsters in which high blood cholesterol levels were induced with a choles-terol-rich diet were fed on milk fermented by L. paracasei subsp. paracasei NTU 101, L. plantarum NTU 102, and L. acidophilus BCRC 17010. The results showed that all the three Lactobacillus strains were effective in reducing cholesterol and LDL-C levels. This was in agreement with other studies (Harrison and Peat 1975; Grunewald 1982; Gilliland et al.1985; Danielson et al.1989). In the hamsters fed on skim milk, although blood cholesterol and LDL-C were also lowered, the difference was not significant
4 5 6 7 8 9 a b c c c Group A B C D E log CFU/g log CFU/g 1 2 3 4 5 6 7 8 9 a a ab a a
a
b
Fig. 2 Effect of nonfermented milk, fermented milk produced from Lactobacillus, and control group on lactobacilli and bifi-dus. a Count of Lactobacillus colony in cecum. b Count of Bifidus colony in cecum. A, B, C, D, and E conditions are the same as in Fig.1.a,b,cp<0.05 is significantly different from A
compared to that of the control group. A similar result was also observed by Grunewald (1982) and Mann (1977).
In this study, HDL-C and TG contents were reduced in the hamsters fed on diet containing lactic acid bacteria. Similar results in humans and swines were also reported by Keim et al. (1981) and Rossouw et al. (1981). However, in a study performed by Hashimoto et al. (1999), a diet containing L. casei TMC 0409 was found to raise the concentration of HDL-C in the blood. Besides, Fukushima and Nakao (1996) indicated that no significant difference was found in the HDL-C content corresponding to supplement of probiotics, including Lactobacillus and Streptococcus, in lipid-rich and cholesterol-rich diets.
The cholesterol reduction produced by lactic acid bacteria can be explained by five mechanisms (Rao et al. 1981; Grunewald1982; Suzuki et al.1991; Fukushima and Nakao
1995; Beena and Prasad1997; Hashimoto et al. 1998), as follows: (1) fermentation products of lactic acid bacteria inhibit the activity of enzymes for cholesterol synthesis and thus reduce cholesterol production; (2) the bacteria facilitate the elimination of body cholesterol in feces; (3) the bacteria inhibit the absorption of cholesterol back into the body by binding with cholesterol; (4) the bacteria interfere with the
recycling of bile salt (a metabolic product of cholesterol) and facilitate its elimination, which raises the demand for bile salt made from cholesterol and thus results in body cholesterol consumption; and, (5) due to the assimilation of lactic acid bacteria, cholesterol in the host body is incorporated into the cell membrane or cell wall of bacteria to increase the resistance of bacterial cell membrane to environmental challenge; thus, the host cholesterol content is reduced.
The Lactobacillus strains used in this study (L. pa-racasei subsp. papa-racasei NTU 101, L. plantarum NTU 102, and L. acidophilus BCRC 17010) lowered the cho-lesterol content in the growth medium and increased in-corporated cholesterol in the cell membrane when bile salt was added in an in vitro trial (data not shown). This indicated that these bacteria were effective in cholesterol reduction in the presence of bile salt. According to the study of Noh et al. (1997), the reduction in bile salt content was due to the activity of bile salt hydrolase in lactic acid bacteria. Bile salt is first hydrolyzed as bile acid, which is then incorporated into lactic acid bacteria, where bile acid is converted to cholesterol. Therefore, the mechanism of cholesterol reduction in the three Lactobacillus strains used in this study is likely to convert bile salt to free bile acid
% 10 20 30 40 50 60 70 a a a a a
b
Group A B C D E % 0 10 20 30 40 50 60 70 a ab ab b aba
Fig. 3 Effect of nonfermented milk, fermented milk produced from Lactobacillus strains, and control group on fecal water content. a Water contained in feces during half of the feeding period. b Water contained in feces at the end of the feeding period. A, B, C, D, and E conditions are the same as in Table2.a,bp<0.05 is signifi-cantly different from A
through an enzymatic activity that deconjugates conjugated bile acid in the intestinal lumen. As bile acid is ready to bind cellulose or intestinal bacteria, it can be eliminated quickly (Chikai et al. 1987). This lowers the bile acid content and causes the liver to use cholesterol to produce bile acid, which is later converted to bile salt to meet the demands of lipid digestion and absorption. Thus, the out-come is reduction in body cholesterol. The three Lactoba-cillus strains used in this study reduced blood and liver cholesterol contents in the in vivo trial.
At the end of the feeding period, the cell counts of lactic acid bacteria in the cecum in groups C, D, and E were 103 times greater than that in the control group. This indicated that the Lactobacillus strains fed to the animals in this study could successfully tolerate gastric acid and bile salt, adhere to the intestinal wall, grow, and proliferate. The result was in agreement with a study of Usman and Hosono (2000). There was no significant difference in the colony numbers of Bifidobacterium among the five groups when Lactobacillus strains were not fed to the animals. Many studies have shown that lactic acid bacteria inhibit the proliferation of pathogenic bacteria, improve intestinal microflora, reduce the risk for digestive diseases such as diarrhea and ulcer, and promote the health of host animals. From the effects of cholesterol reduction shown by the three Lactobacillus strains (L. paracasei subsp. paracasei NTU 101, L. plantarum NTU 102, and L. acidophilus BCRC 17010) in hamsters, it is deduced that the activity of bile salt hydrolase might be involved. The lactic acid bacteria could grow and proliferate in the cecum; thus, they might interfere with pathogenic bacteria in the stomach. Group D (fed on L. plantarum NTU 102) showed fecal water content higher than that of the control group, sug-gesting that the Lactobacillus strain might facilitate fecal elimination. Therefore, lactic acid bacteria may improve food digestion, food absorption, and fecal elimination in the host.
Conclusion
In this study, both Lactobacillus strains isolated from the human gut and pickled vegetables, respectively, were ef-fective in reducing cholesterol in the blood and in the liver. We further plan to initiate a toxicity trial and a clinical trial to confirm the hypocholesterolemic effects of these Lacto-bacillus strains.
References
American Institute of Nutrition (1977) Report of the American Institute of Nutrition ad hoc committee on standards for nutritional studies. J Nutr 107:1340–1348
Beena A, Prasad V (1997) Effect of yogurt and bifidus yogurt fortified with skim milk powder, condensed whey and lactose-hydrolysed condensed whey on serum cholesterol and triacyl-glycerol levels in rats. J Dairy Res 64:453–457
Brashears MM, Gilliland SE, Buck LM (1998) Bile salt deconjuga-tion and cholesterol removal from media by Lactobacillus casei. J Dairy Sci 81:2103–2110
Bucolo G, David H (1973) Quantitative determination of serum triglyceride by the use of enzyme. Clin Chem 19:476–482 Chikai T, Nakao H, Uchida K (1987) Deconjugation of bile acids by
human intestinal bacteria implanted in germ free rats. Lipids 22:669–671
Danielson AD, Peo ERJ, Shahani KM, Lewis AJ, Whalen PJ, Amer MA (1989) Anticholesterolemic property of Lactobacillus acidophilus yogurt fed to mature boars. J Anim Sci 67:966–974 Fossati P, Prencipe L (1982) Serum triglycerides determined colorimetrically with an enzyme that produce hydrogen perox-ide. Clin Chem 28:2077–2080
Frick M, Elo O, Haapa K (1987) Helsinki heart study: primary prevention trial with gemfibrozil in middle-age men with dyslipemia. N Engl J Med 317:1237–1245
Fukushima M, Nakao M (1995) The effect of a probiotic on faecal and liver lipid classes in rats. Br J Nutr 73:701–710
Fukushima M, Nakao M (1996) Effects of mixture of organisms, Lactobacillus acidophilus or Streptococcus faecalis on choles-terol metabolism in rats fed on a fat- and cholescholes-terol-enriched diet. Br J Nutr 76:857–867
Gilliland SE, Nelson CR, Maxwell C (1985) Assimilation of cholesterol by Lactobacillus acidophilus. Appl Environ Microbiol 49:377–381
Grunewald KK (1982) Serum cholesterol levels in rats fed skim milk fermented by Lactobacillus acidophilus. J Food Sci 47:2078–2079
Harrison VC, Peat G (1975) Serum cholesterol and bowel flora in the newborn. Am J Clin Nutr 28:1351–1355
Hashimoto H, Yamazaki K, Arai Y, Kawase M, He F, Hosoda M, Hosono A (1998) Effect of lactic acid bacteria on serum cholesterol level in rats fed cholesterol diet. Anim Sci Technol 69:702–707
Hashimoto H, Yamazaki K, He F, Kawase M, Hosoda M, Hosono A (1999) Hypocholesterolemic effects of Lactobacillus casei subsp. casei TMC 0409 strain observed in rats fed cholesterol contained diets. Anim Sci J 72:90–97
Hepner G, Fried R, Jeor S, Fusetti St L, Morin R (1979) Hypocholesterolemic effect of yogurt and milk. Am J Clin Nutr 32:19–24
Juang IL, Pan TM, Huong HP, Huang CJ (2000) The effect of oligosaccharide beverage on microflora in rats. Chin J Nutr 25:232–242
Keim NL, Marlett JA, Amundsopn CH (1981) The cholesterolemic effect of skim milk in young men consuming controlled diets. Nutr Res 1:422–429
Lin FM, Chiu CH, Pan TM (2004) Fermentation of a milk–soymilk and Lycium Chinese Miller mixture using a new isolate of Lactobacillus paracasei subsp. paracasei NTU 101 and Bifidobacterium longum. J Ind Microbiol Biotech 31:559–564 Mann GV (1977) A factor in yogurt which lowers cholesteremia in
man. Atherosclerosis 26:335–340
Mann GV, Spoerry A (1974) Studies of a surfactant and cholester-emia in the Massai. Am J Clin Nutr 27:464–469
McGowan MW, Artiss JD, Strandbergh DR, Zak B (1983) A peroxidase-coupled method for the colorimetric determination of serum triglycerides. Clin Chem 29:512–538
Nielson JW, Gilliland SE (1985) Variations in cholesterol assimi-lation by individual strains of Lactobacillus acidophilus and Lactobacillus casei from human intestines. J Dairy Sci 68 (Suppl 1):83 (Abstract)
Noh DO, Kim SH, Gilliland SE (1997) Incorporation of cholesterol into the cellular membrane of Lactobacillus acidophilus ATCC 43121. J Dairy Sci 80:3107–3113
Pan TM, Chiu CH, Gun YK (2002) Characterization of Lactoba-cillus isolated from pickled vegetables for use as dietary or pickle adjuncts. Food Food Ingred Japan 206:45–51
Rao DR, Chawan CB, Pulusani SR (1981) Influence of milk and Thermophilus milk on plasma cholesterol levels and hepatic cholesterogenesis in rats. J Food Sci 46:1339–1341
Richmond W (1973) Preparation and properties of a cholesterol oxidase from Nocardia sp. and its application to the enzymatic assay of total cholesterol in serum. Clin Chem 19:1350–1356 Rossouw JE, Burger E, Van der Vyver P, Ferrerira JJ (1981) The
effect of skim milk, yogurt and full cream on human serum lipids. Am J Clin Nutr 34:351–356
Suzuki Y, Kaizu H, Yamauchi Y (1991) Effect of cultured milk on serum cholesterol concentrations in rats which fed high-cholesterol diets. Animal Sci Technol 62:565–571
Thakur CP, Jha AN (1981) Influence of milk, yogurt and calcium on cholesterol induced atherosclerosis in rabbits. Atherosclerosis 89:211–215
Usman, Hosono A (2000) Effect of administration of Lactobacillus gasseri on serum lipids and fecal steroids in hypercholesterol-emic rats. J Dairy Sci 83:1705–1711