497
*To whom correspondence and reprint requests should be addressed. Tel: 886-2-23660235. E-mail: hwyuan@ntu.edu.tw
H
abitat fragmentation and habitat loss areconsidered major threats to biodiversity (Wilson 1992). One unavoidable consequence of habitat fragmentation is an increase in edge habitats, which in turn affects abiotic and biotic processes in the landscape (Murcia 1995). Nesting birds may disproportionately favor the edges of forest patch-es due to the availability of forpatch-est cover and abun-dant food, but may suffer high nest predation as a result. Therefore, edges may become“ecological traps”leading to severe population declines in edge-nesting species (Gates and Gysel 1978).
The effects of habitat fragmentation are not homogeneous. There are species-specific responses (e.g., Warkentin et al. 1995, Bourque
and Villard 2001, Flaspohler et al. 2001), guild-specific responses (e.g., Stouffer and Bierregaard 1995), and, theoretically, life history-specific responses (e.g., Donovan and Thompson 2001) to edge habitats. How individuals perceive patchi-ness and how they move among fragments influ-ence how they are affected by fragmentation (Wiens 1994, Ims 1995, Andreassen et al. 1998). Therefore, understanding the pattern of habitat selection, together with the behavioral basis for the selection, can help explain variations in edge effects. Few studies have combined individual-level mechanisms with a population-individual-level demo-graphic structure. Even fewer, if any, multilevel studies have been conducted in tropical and
sub-Habitat Selection of the Cooperative Breeding Taiwan Yuhina (Yuhina
brunneiceps) in a Fragmented Forest Habitat
Pei-Fen Lee1, Sheng-Feng Shen1,2, Tzung-Su Ding3, Chyi-Rong Chiou3, and Hsiao-Wei Yuan3,* 1Institute of Ecology and Evolutionary Biology, and Department of Life Science, National Taiwan University, Taipei, Taiwan 106, R.O.C.
E-mail: leepf@ntu.edu.tw
2Present address: Department of Neurobiology and Behavior, Cornell University, Ithaca, NY 14853, USA
E-mail: ss456@cornell.edu
3School of Forestry and Resource Conservation, National Taiwan University, Taipei, Taiwan 106, R.O.C.
E-mail: ding@ntu.edu.tw (TSD), esclove@ntu.edu.tw (CRC), and hwyuan@ntu.edu.tw (HWY) (Accepted July 22, 2005)
Pei-Fen Lee, Sheng-Feng Shen, Tzung-Su Ding, Chyi-Rong Chiou, and Hsiao-Wei Yuan (2005) Habitat selection of the cooperative breeding Taiwan Yuhina (Yuhina brunneiceps) in a fragmented forest habitat.
Zoological Studies 44(4): 497-504. We used multilevel analyses (individual habitat selection and population
structure) to study edge effects on the Taiwan Yuhina (Yuhina brunneiceps), an endemic subtropical species at Meifeng, central Taiwan. The 95% kernel home range area was used to study habitat preferences, and the source-sink threshold was calculated to determine whether this yuhina population is self-sustainable. Within the study site, woods, open spaces, orchards, buildings, and pond habitats were preferred in that order. Within the home ranges, orchard habitats were preferred to open spaces. Larger breeding groups had larger home ranges with more trees and open spaces. Nest sites were closer to forest edges than random points were. The density of Taiwan cherry trees was significantly higher within home ranges than available areas, especially near edges, suggesting the fruit,s potential value to breeding yuhinas. Although the yuhinas suffered high rates of nest failure at Meifeng, this population still seemed to be self-sustaining due to the long breeding season with multiple broods and a cooperative breeding strategy. http://zoolstud.sinica.edu.tw/Journals/44.4/497.pdf Key words: Edge, Fragmentation, Home range, Source-sink population dynamics.
tropical areas, where ecological contexts (e.g., higher predation risks and longer breeding sea-sons) and life histories (e.g., multiple broods) may widely differ from those of temperate species. These differences may have profound influences on a species, responses to edge habitats (Martin 1996, Russell 2000, Stutchbury and Morton 2001).
The Taiwan Yuhina (Yuhina brunneiceps), with monomorphic sexes, is a passerine bird endemic to Taiwan (Cibois et al. 2002). It is widely distributed in forests from 800 to 3000 m in eleva-tion (Koh and Lee 2003). Yuhinas are the only known passerine species to consistently adopt a joint-nesting strategy in a large fraction of nests (Vehrencamp and Quinn 2004). Breeding groups include 2-7 individuals composed primarily of non-kin. Therefore juvenile delayed-dispersal is not observed (Yuan et al. 2004). Living in groups typi-cally grants each individual increased protection from predators and increased chances of finding food (Krebs and Davies 1993). In White-throated Magpie-Jays (Calocitta formosa), a cooperatively breeding species, it has been shown that larger groups possess larger territories containing more food, which contributes to higher reproductive suc-cess (Langen and Vehrencamp 1998).
In this paper, we examined 1) the influence of fragmentation on yuhinas, at the individual level, with respect to (a) habitat selection and (b) the behavioral basis of habitat selection in terms of nest site and food resources; 2) this yuhina popu-lation,s possible status as a source population (in terms of meta-population dynamics (Pulliam 1988); and 3) variations in home range qualities among breeding groups of different sizes.
MATERIALS AND METHODS Behavioral observations
We conducted this study at Meifeng Highlands Experiment Farm of National Taiwan University in Nantou County, central Taiwan (24 05'N, 121
°
10'E, elevation 2150 m; Fig. 1). The study site, established for horticultural research purposes in 1961, a 50 ha area mixed with green-houses, temperate-fruit orchards, meadows, Japanese Cryptomeria (Cryptomeria japonica) plantations, and various ornamental trees sur-rounded by natural forests composed mainly of Fagaceae and Lauraceae.Since 1995, we have banded adult and juve-nile yuhinas at Meifeng to study their breeding
ecology. By 2001, 97% of the adults in our study site were color-banded. We closely monitored the breeding success of 6, 11, and 13 groups in 1998, 2000, and 2001, respectively. Each morning of the 2001 breeding season (early Mar. to late Aug.), we walked along a fixed route that covered the entire study site, weather permitting. When we encoun-tered a banded group, we followed the group and plotted each member,s location on a fine-scale map every 5 min until contact was lost. To facili-tate mapping, we established a grid system with reference points every 20 m at the study site.
In 2001, 91 nests were found from 20 breed-ing groups for nestbreed-ing analyses. We searched for yuhina nests in both natural and man-made habi-tats and mapped the location of each nest. We were confident that most of the nests within the study site had been discovered, as yuhinas are an active and conspicuous species, and every breed-ing group was closely followed.
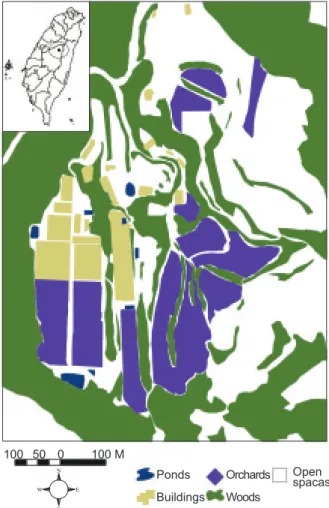
Fig. 1. Study area and land cover map at Meifeng, the Highlands Experiment Farm of National Taiwan Univ., Nantou County, central Taiwan.
100 50 0 100 M Ponds Buildings Orchards Woods Open spacas N W E S
°
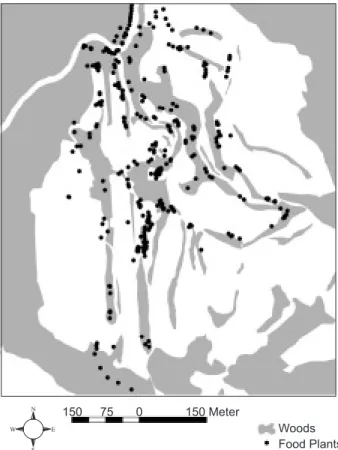
Although the diet of yuhinas varies with the seasons, it consisted primarily of fruits, nectar, and insects within the study site. During the breeding season, yuhinas mainly foraged on 3 plant species: nectar (Jan.-Apr.) and fruits (May-June) of Taiwan cherry (Prunus campanulata), fruits (May-July) of Eastern debregeasia (Debregeasia orien-talis), and nectar (Aug.-Sept.) of mistletoe (Taxillus lonicerifolius) (Liu 1999). We mapped the loca-tions of these 3 species within the study site and used their abundances as food resource indexes for the yuhinas.
Habitat data analysis
A digitized habitat map of the study site was created by interpreting an aerial photo (1:5000, 0.5 x 0.5 m in spatial resolution, taken in 2000 by the Agricultural and Forestry Aerial Survey Institute of the Taiwan Forestry Bureau, Council of Agriculture, Executive Yuan) using ArcGIS 8.1 (ESRI 2001). Five major habitat types were identified by land cover and land use: woods, orchards, buildings, open spaces, and ponds (Fig. 1). Wooded habitat was defined as forest patches and hedgerow strips with trees more than 3 m in height and included primary forests and Cryptomeria plantations. Orchard habitats were plantations of temperate fruit trees, mostly apples, peaches, and pears. Building habitat included greenhouses, residences, and offices. Open space habitat included scat-tered open spaces with shrubs < 3 m in height, scattered individual trees, grass, bare ground, trails, concrete or asphalt roads, and nearby areas. Pond habitat consisted of human-constructed irri-gation ponds. All patches of the 5 habitat types were confirmed by field verification.
Nest sites and all locations where banded group members were observed were mapped using ArcGIS 8.1. A fixed kernel density analysis was applied to delineate the home range of each breeding group. We used a 95% kernel area to avoid potential bias caused by extreme points (Silverman 1986, Powell 2000). To determine nest site preferences in wooded habitat, we generated random points in the wooded habitat within each group,s home range using Animal Movement (ArcView) software (Hooge and Eichenlaub 2000), then calculated the distance from each nest site and random point to the nearest edge.
To determine habitat preferences of each group, we overlaid each home range with the habi-tat map and calculated the area composition of the habitat types. Food resource quality was indexed
by the density of the 3 major food plants within the home range. Those plants shared by home ranges of multiple breeding groups were equally divided by the number of groups and assigned to each home range.
Two levels of use-availability design (Johnson 1980), i.e., utilization vs. home range and home range vs. available area, were conducted to assess habitat selection by yuhinas. We deter-mined the available area for each group separately by generating 100 m circular buffers around all group members, occurrence locations by ArcGIS (McClean et al. 1998, Jones 2001). Because pro-portions of habitat types were not independent, we used compositional analyses to determine the habitat preferences of yuhinas (Aebischer et al. 1993).
Demographic data analysis
The source-sink threshold, as the number of juveniles per pair per year, was calculated accord-ing to the followaccord-ing equation (Trine 1998) for 1998, 2000, and 2001:
Source-sink threshold =
The adult mortality rate was estimated by the overwintering return rate. Because juveniles com-monly disappear from Meifeng (78%, Yuan et al. 2004) and we have observed banded juveniles which had dispersed as far as 7 km away, direct estimation of the juvenile mortality rate was extremely difficult. We used 50% of the adult sur-vival rate to estimate yearling sursur-vival rate, as has been done in many migratory songbird studies (Greenberg 1980, May and Robinson 1985, Thompson 1993, Donovan et al. 1995). If the annual productivity of the population was higher than the calculated source-sink threshold, the pop-ulation was considered to be a demographic source. Alternatively, the average number of fledglings and the adult mortality rate of a given year were applied to the source-sink threshold equation. We could thus obtain the maximum juvenile mortality rate for a self-sustainable popula-tion each year.
RESULTS Nesting ecology
Most (84%) nests were found during the con-2*adult mortality
struction stage. Seventy-six nests (84%) were built in wooded habitats (Fig. 2); among them, 63 nests (83%) were built in Japanese Cryptomeria. The Manly-Chesson indexes also strongly indicat-ed that woods were the most-important habitat used by yuhinas (Table 1). The distance from
each nest to the nearest wooded edge was signifi-cantly shorter than that from random points (Mann-Whitney U test = 1154.50, p < 0.001, n = 74; Fig. 3).
Home range preference
Of the 20 breeding groups followed, only 13 groups had home ranges completely within the study site, and the following analyses of home range preferences were limited to those groups. We recorded 99 ± 28 (mean ± SD) occurrence locations (i.e., location of individual or group sight-ings) per group during the 2001 breeding season. The area of the 95% kernel home range of the breeding groups was 2.3 ± 2.0 ha (Fig. 4). Most of the breeding groups had home ranges which greatly overlapped those of other groups. The home range of each group, on average, over-lapped with 2.5 other groups, home ranges. The
Fig. 2. Distribution of 91 yuhina nests found at Meifeng during the breeding season of 2001. Most (84%) of the nests were built in trees. The gray area indicates the wooded habitat as shown in figure 1. N W E S 150 Meter 150 75 0 TREE Nest site
Table 1. Manly-Chesson selectivity indexes of nest site selection for yuhinas at
Meifeng in 2001
Habitat type
Buildings Orchards Open spaces Woods Ponds
Percentage available (%) 3.00 11.4 40.5 44.80.31
Percentage of all nests found (%, n = 91) 1.10 6.6 8.80 83.5 0.00
Manly-Chesson indexa 0.37 0.580.22 1.86 0.00
Standardized Manly-Chesson indexb 0.12 0.19 0.07 0.62 0.00
aPercent used / percent available. bEach index was divided by the sum of all indexes.
Fig. 3. Comparison of distances of nests and of randomly selected points within the wooded habitat to the nearest edge.
Random Actual
Propotion of nests (%)
Distances to nearest edges (m) 70 60 50 40 30 20 10 0 >10 10 8 6 4 2 0
average percent of overlap was about 35% (range, 0%-82%). The habitat composition of the 95% kernel home range significantly differed from that of the entire study site (Wilks, lambda = 0.087, F5, 20= 44.0, p < 0.001). The most frequently used habitats were woods, open spaces, orchards, buildings, and ponds, in that order. Similarly, with-in the home range of each breedwith-ing group, habitat composition of the occurrence locations of breed-ing groups significantly differed from the habitat composition of the home ranges (Wilks, lambda = 0.136, F3, 22 = 46.6, p < 0.0001), indicating that yuhinas showed habitat preferences. Within the home ranges, woods, orchards, open spaces, buildings, and ponds were preferred, in that order.
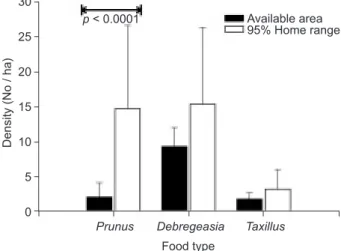
Food plants were abundant within each groups, home ranges (Fig. 5). These plants were usually located within wooded patches, where they typically comprised the subordinate or shrub layers vertically. Only the density of Taiwan cherry was significantly higher within the home ranges than in other available areas (Mann-Whitney U test = 15.00, p < 0.001; Fig. 6). In addition, we found
that 85% of these trees were in wooded patches.
Group size effect of home range quality
Breeding groups did not choose home range areas and habitat types within home ranges at ran-dom (Wilks, lambda = 0.064, F5, 8 = 23.47, p < 0.001). Larger groups used larger home range areas (rs = 0.684, p < 0.01). However, the area per capita did not differ across groups (0.53 ± 0.27 ha, rs = 0.077, p = 0.234). Two habitat types, wooded (rs = 0.712, p = 0.006) and open spaces (rs= 0.712, p = 0.006), made up larger proportions of the habitat within home ranges of larger breed-ing groups. Other habitat types did not vary from group to group (p > 0.05), and neither did food plants (p > 0.05).
Is the Meifeng yuhina population self-sustain-able?
Adult mortality rates were estimated to be 28.6%, 18.8%, and 30.6% in 1998, 2000, and
N W E
S
Fig. 4. Distribution of the 95% kernel home ranges of 13 yuhi-na breeding groups. Letters and numbers on the map refer to individual groups, and the gray area indicates the wooded habi-tat as shown in figure 1.
150 Meter
Woods
150 75 0
Fig. 5. Distribution of major food plants, Taiwan cherry (Prunus
campanulata), Eastern debregeasia (Debregeasia orientalis),
and mistletoe (Taxillus lonicerifolius), at Meifeng. The gray area indicates the wooded habitat as shown in figure 1.
N W E S 150 Meter Woods Food Plants 150 75 0 GA GA J4 J4 TR GA RO GA GA J4 J4 J6 WH WH WH PH PH OF RO TR J6 IG IG IG J6 M6 M6 PR PR PH PH RO BR BR WA WA BR M6 M6 TR J6
2001, respectively. The number of adults and fledglings of different groups were pooled for each year to calculate the annual productivity. There were 28, 48, and 49 breeders and 29, 30, and 40 fledglings in 1998, 2000, and 2001, respectively. The average numbers of fledglings per pair were 2.1 and 1.3 in 1998 and 2000, higher than the source-sink thresholds of 1.3 and 0.6, respectively. However, in 2001, the average number of fledg-lings per pair was 1.6, which equaled the source-sink threshold of 1.6.
We used the data given above to calculate the“maximum”juvenile overwintering mortality rates for the population to be self-sustainable (see “Methods”for details). These maximum juvenile mortality rates were 72.8%, 71.0%, and 61.8%, which were 2.5, 3.8, and 2.0 times higher than the adult mortality rates in 1998, 2000, and 2001, respectively.
DISCUSSION
The yuhina population within the fragmented habitats at Meifeng Farm seems to be a self-sus-tainable population based on our demographic analyses. We believe that the conventional assumption of a juvenile survival rate of 50% of the adult survival rate for migratory birds in our analy-ses is a conservative estimate, because yuhinas typically migrate short distances along local eleva-tional gradients (Liu 1999). In addition, the maxi-mum juvenile mortality rates for this population if it were self-sustainable were larger than those of most studies on migratory birds. As in Anders and Marshall,s (2005) review of 7 species with detailed
estimations of fledgling survival rate, the average overwintering survival rate was 52% (range, 25%-72%, generating an average mortality rate of 48%). Therefore, even with such high estimated juvenile mortality rates in our study compared to others, this population is still (or very close to) self-sustainable.
Meifeng Farm is comprised of a mosaic of habitat types. One interesting question is the effect of such spatial heterogeneity on ecological processes for the yuhinas. We found that wooded patches were an important habitat type for provid-ing nest sites and food supply at the individual habitat selection level. This might be“habitat compensation”which indicates that animals may compensate for the loss of their preferred habitat type (woods) by shifting to a less-preferred one (orchards) (Haila et al. 1989, Norton et al. 2000).
Yuhinas frequently selected wooded and orchard patches as nest sites, especially near the edge. Houbara Bustards (Chlamydotis undulata) were found to select edges as nesting sites to increase their ability to detect predators (Yang et al. 2003). Predation was the primary cause of nest failure at Meifeng in 2001 (55%, Yuan HW, unpubl. data). However, the major nest predators were Eurasian Jays (Garrulus glandarius) and Taiwan Sibias (Heterophasia auricularis), which are abun-dant around forest edges. Evidently, nest security was not a factor in the yuhinas, choice of edge habitats. The higher density of Taiwan cherry trees within the home ranges, especially around edges, might be one of the advantages for using edges to breed. In fragmented landscapes, the food supply generally declines with decreasing fragment size (Zanette 2001). But this is not the case at Meifeng where many plants, such as Taiwan cherry, were deliberately planted to enhance the visual landscape and have incidental-ly become food resources for the yuhinas. Abiotic microclimatic factors may also play a role in the yuhinas, preference for habitat edges. The forest interior is usually damp and cool at Meifeng. Higher temperatures and lower humidities at the edges (reviewed in Murica 1995, Roadewald and Yahner 2001) may lower energetic costs for incu-bating adults (Wolf and Walsberg 1996, reviewed in Hansell 2000) and provide a more-suitable microenvironment for the survival of eggs and chicks.
However, with its cooperative breeding strate-gy, yuhinas have some advantages for coping with the high rates of nest failure by reducing the work-load per breeder with each nest attempt. Yuhinas Fig. 6. Comparison of 3 food plant species densities within the
95% kernel home range areas with available areas. Food type
p < 0.0001
Density (No / ha)
Prunus Debregeasia Available area 95% Home range Taxillus 30 25 20 15 10 5 0
exhibit multiple brooding in a single breeding sea-son; greater numbers of breeders can accelerate the renesting process and therefore help spread out the risk of nest failure (Shen 2002). Thus, at the population level, the yuhinas at Meifeng served as the source populations in 1998 and 2000. In those years, the Mayfield nest survival rate was 27% and 25%, respectively (Yuan et al. 2004). In 2001, when predation and adverse weather condi-tion caused an extraordinarily low nest survival rate (14%; Yuan et al. 2004), the population was static.
However, our study area is surrounded by natural forests, and our analyses were only carried at the habitat patch scale. Therefore, the land-scape-scale effect of habitat fragmentation such as disruption of individual dispersal demonstrated by Cooper and Walters (2002) in the Brown Treecreeper (Climacteris picumnus) was not deter-mined in our study. Further studies on multi-scale analyses will be valuable.
Larger groups using larger home ranges con-sisting primarily of wooded and nearby open habi-tat types is consistent with the overall home range habitat preferences of yuhinas, indicating a better home range quality for larger groups. White-throated Magpie-Jays (Langen and Vehrencamp 1998) and Groove-billed Anis (Crotophaga sul-cirostris; Vehrencamp et al. 1988) show similar patterns. However, the area per capita is about the same. Also, there were no differences in the quantities of food plants in the home ranges of yuhina groups of different sizes, suggesting that larger groups using larger home ranges probably does not impact individual fitness in terms of food resource abundance. However, the greater pro-portion of wooded and open habitats within larger home ranges might provide nesting sites for larger groups.
While many studies on edge effects have been conducted in temperate zones, this study of a subtropical species with a unique life history has several implications for future work on edge effects. First, species life history characteristics (e.g., cooperative breeding and multiple broods) are important in understanding the edge effects on bird populations. Linking these mechanisms to larger-scale phenomenological explanations can further complete our understanding of edge effects and avoid misinterpretation of phenomena (like the high nest predation rate, in this case). Second, the ecological contexts of different regions can influ-ence the edge effects (e.g., a long breeding sea-son). Therefore, the study of edge ecologies of
tropical and subtropical regions and in the Southern Hemisphere will enhance our under-standing and help delineate conservation policies (e.g., Schmiegelow and Mönkkönen 2002).
Acknowledgments: We thank LL Lee, SC Lee,
the Chief Editor, and 3 anonymous reviewers for valuable comments on previous drafts of this man-uscript; P Coulter and B Burt for English editing; and MC Tsai, KZ Lin, and other workers at the Meifeng Farm for logistical support. We greatly appreciate the volunteers from the NTU Nature Conservation Students, Club and School of Forestry and Resource Conservation, in particular HY Hung, IH Chang, and KD Zhong, for their help in the field. This study was partially supported by National Science Council and the Council of Agriculture, R.O.C.
REFERENCES
Aebischer NJ, PA Robertson, ER Kenward. 1993. Compositional analysis of habitat use from animal radio-tracking data. Ecology 74: 1313-1325.
Anders AD, MR Marshall. 2005. Increasing the accuracy of productivity and survival estimates in assessing landbird population status. Conserv. Biol. 19: 66-74.
Andreassen HP, K Hertzberg, RA Ims. 1998. Space-use responses to habitat fragmentation and connectivity in the root vole Microtus oeconomus. Ecology 79: 1223-1235. Bourque J, MA Villard. 2001. Effects of selection cutting and
landscape-scale harvesting in the reproductive success of two neotropical migrant bird species. Conserv. Biol. 15: 184-195.
Cibois A, MV Kalyakin, LX Han, E Pasquet. 2002. Molecular phylogenetics of babblers (Timaliidae): revaluation of the genera Yuhina and Stachyris. J. Avian Biol. 33: 380-390. Cooper CB, JR Walters. 2002. Independent effects of
wood-land loss and fragmentation on Brown Treecreeper distri-bution. Biol. Conserv. 105: 1-10.
Donovan TM, HR Lamberson, A Kimber, FR Thompson III, J Faaborg. 1995. Modeling the effects of habitat fragmen-tation on source and sink demography of neotropical migrant birds. Conserv. Biol. 9: 1396-1407.
Donovan TM, FR Thompson III. 2001. Modeling the ecological trap hypothesis: a habitat and demographic analysis for migrant songbirds. Ecol. Appl. 11: 871-882.
ESRI. 2001. ArcGIS 8.1. Redlands, MI.: Environmental Systems Research Institute.
Flaspohler DJ, SA Temple, RN Rosenfield. 2001. Species-specific edge effects on nest success and breeding bird density in a forested landscape. Ecol. Appl. 11: 32-46. Gates JE, LS Gysel. 1978. Avian nest dispersion and fledgling
success in field-forest ecotone. Ecology 59: 871-883. Greenberg R. 1980. Demographic aspects of long-distance
migration. In A Keast, ES Morton, eds. Migrant birds in the Neotropics: ecology, behavior, distribution, and con-servation. Washington DC: Smithsonian Institution Press, pp. 493-504.
Haila Y, IK Hanski, S Raivio. 1989. Methodology for studying the minimum habitat requirements of forest birds. Ann. Zool. Fenn. 26: 173-180.
Hansell M. 2000. Bird nests and construction behavior. Cambridge, UK: Cambridge Univ. Press.
Hooge PN, B Eichenlaub. 2000. Animal movement extension to Arcview, ver. 2.0. Anchorage, AL: Alaska Science Center - Biological Science Office, US Geological Survey. Ims RA. 1995. Movement patterns in relation to landscape
structures. In L Hansson., L Fajrig, G. Merriam, eds. Mosaic landscapes and ecological processes. Berlin: Springer-Verlag, pp. 89-109.
Johnson DH. 1980. The composition of usage and availability measurements for evaluating resource preference. Ecology 6: 65-71.
Jones J. 2001. Habitat selection studies in avian ecology: a critical review. Auk 118: 557-562.
Koh CN, PF Lee. 2003. Elevational gradients in breeding birds in northern Taiwan. Taiwan J. For. Sci. 18: 349-361. (in Chinese with English summary)
Krebs JR, NB Davies. 1993. An introduction to behavioural ecology. London: Blackwell Scientific Publication. Langen TA, SL Vehrencamp. 1998. Ecological factors
affect-ing group and territory size in White-throated Magpie-Jays. Auk 115: 327-339.
Liu M. 1999. The cooperative breeding of Taiwan Yuhina (Yuhina brunneiceps) at Meifeng area, Taiwan. Master,s thesis, National Taiwan Univ., Taipei, Taiwan. (in Chinese with English abstract)
Martin TE. 1996. Life history evolution in tropical and south temperate birds: What do we really know? J. Avian Biol. 27: 263-272.
May RM, SK Robinson. 1985. Population dynamics of avian brood parasitism. Am. Nat. 126: 475-494.
McClean SA, MA Rumble, RM King, WL Baker. 1998. Evaluation of resource selection methods with different definitions of availability. J. Wildlife Manage. 62: 793-801. Murcia C. 1995. Edge effects in fragmented forests:
implica-tions for conservation. Trends Ecol. Evol. 10: 58-62. Norton MR, SJ Hannon, FKA Schmiegelow. 2000. Fragments
are not islands: patch vs landscape perspectives on song-bird presence and abundance in a harvested boreal for-est. Ecography 23: 209-223.
Powell RA. 2000. Animal home ranges and territories and home range estimators. In L Boitani, TK Fuller, eds. Research techniques in animal ecology. New York: Columbia Univ. Press, pp. 65-110.
Pulliam HR. 1988. Sources and sinks, and population regula-tion. Am. Nat. 132: 652-661.
Rodewald AD, RH Yahner. 2001. Influence of landscape com-position on avian community structure and associated mechanisms. Ecology 82: 3493-3504.
Russell EM. 2000. Avian life histories: Is extended parental care the southern secret? Emu 100: 377-399.
Schmiegelow FKA, M. Mönkkönen. 2002. Habitat loss and fragmentation in dynamic landscapes: avian perspectives from the boreal forest. Ecol. Appl. 12: 398-411.
Shen SF. 2002. The ecology of cooperative breeding Taiwan Yuhinas (Yuhina brunneiceps) at Meifeng area. Master,s thesis, National Taiwan Univ., Taipei, Taiwan. (in Chinese with English abstract)
Silverman BW. 1986. Density estimation for statistics and data analysis. London: Chapman and Hall.
Stouffer PC, RO Bierregaard. 1995. Effects of forest fragmen-tation on understory hummingbirds in Amazonian Brazil. Conserv. Biol. 9: 1085-1094.
Stutchbury BJM, ES Morton. 2001. Behavioral ecology of trop-ical birds. London: Academic Press.
Thompson FR. III. 1993. Simulated responses of a forest-inte-rior bird population to forest management options in cen-tral hardwood forests of the United States. Conserv. Biol. 7: 325-333.
Trine CL. 1998. Wood thrush population sinks and implica-tions for the scale of regional conservation strategies. Conserv. Biol. 12: 576-585.
Vehrencamp SL, RR Koford, BS Bowen. 1988. The effect of breeding-unit size on fitness components in Groove-billed Anis. In TH Clutton-Brock, ed. Reproductive success. Chicago, IL: Chicago Univ. Press, pp. 291-304.
Vehrencamp SL, JS Quinn. 2004. The evolution of joint-nest-ing systems: mutual cooperation or conspecific brood par-asitism? In WD Koenig, J Dickinson, eds. Cooperative breeding in birds: recent research and new theory. Cambridge, UK: Cambridge Univ. Press, pp. 177-196. Warkentin IG, R Greenberg, OJ Salgado. 1995. Songbird use
of gallery woodlands in recently cleared and older settled landscapes of the Selva Lacandona, Chiapas, Mexico. Conserv. Biol. 9: 1095-1106.
Wiens JA. 1994. Habitat fragmentation: island vs. landscape perspectives on bird conservation. Ibis 137: 97-104. Wilson EO. 1992. The diversity of life. Cambridge, UK:
Belknap Press of Harvard Univ. Press.
Wolf BO, GE Walsberg. 1996. Thermal effects of radiation and wind on a small bird and implications for microsite selec-tion. Ecology 77: 2228-2236.
Yang WK, JF Qiao, O Combreau, XY Gao, WQ Zhong. 2003. Breeding habitat selection by the Houbara Bustard
Chlamydotis (undulata) macqueenii in Mori, Xinjiang,
China. Zool. Stud. 42: 470-475.
Yuan HW, M Liu, SF Shen. 2004. Joint nesting in Taiwan yuhi-nas: a rare passerine case. Condor 106: 862-872. Zanette L. 2001. Indicators of habitat quality and the
reproduc-tive output of a forest songbird in small and large frag-ments. J. Avian Biol. 32: 38-46.