薑黃素和鼠尾草酸下降細胞激素所誘發的人類臍靜脈內皮細胞黏附因子表現; Curcumin and Carnosic acid attenuate cytokine-induced adhesion molecule expression in HUVECs
108
0
0
全文
(2) 謝. 誌. 感謝指導教授余雅美博士兩年來於學業、研究等方面的悉心指導及 待人處事上的諄諄教誨,使本論文得以順利完成。師恩浩瀚,永銘於心, 謹誌卷首,以表誠摯之謝忱。 文稿初成,承蒙台灣大學沈立言教授與中國醫藥大學江素瑛助理教 授於百忙之中撥冗審閱,詳加斧正,並提供寶貴意見使論文內容得以更 加完善,謹此致上萬分謝意。 在學期間,多蒙學長姐的指導與同窗好友杏純、珮玲、馨儀、美瑜、 珊瑩、昇輝及同期研究所同學於實驗與生活上的幫助與支持;實驗室之 學妹于婷、瑜妏、郁婷、珮琪、榆茜的協助與幫忙;再者,要感謝好友 佳蓉、艾倫、閃閃、桓儀、孟青,於艱辛研究過程中給予諸多的協助與 鼓勵,在此致上最誠摯的謝意。 最後由衷感謝我最親愛的爸爸、媽媽以及哥哥的關懷與支持,使我 能安心無慮的完成學業。謹此將本論文獻給所有關心及曾經幫助我的 人,謝謝你們。 林芩卉 謹致於 中國醫藥大學 營養學系 民國 九十六 年 六 月.
(3) Contents 中文摘要 ........................................................................................................................... 1 Abstract ........................................................................................................................... 3 Chapter 1 Introduction ...................................................................................................5 1-1. Atherosclerosis ......................................................................................................... 5. 1-2. Adhesion and atherosclerosis ................................................................................... 7. 1-3 NF-κB and atherosclerosis ....................................................................................... 9 1-4. Oxidative stress and atherosclerosis....................................................................... 1 1. 1-5 Curcumin .................................................................................................................1 2 1-6. Carnosic acid .......................................................................................................... 1 3. 1-7. The purpose of this reasearch................................................................................. 1 4. Chapter 2 Experimentation I ......................................................................................2 5 2-1. Prolegomenon......................................................................................................... 2 6. 2-2. Materials and Methods ........................................................................................... 2 8. 2-3. Results .................................................................................................................... 3 9. 2-4. Discussion .............................................................................................................. 4 3. 2-5. Conclusion.............................................................................................................. 4 7. Chapter 3 Experimentation II .....................................................................................5 9 3-1. Prolegomenon.........................................................................................................6 0. 3-2. Materials and Methods ........................................................................................... 6 2. 3-3. Results .................................................................................................................. 7 1. 3-4. Discussion ............................................................................................................ 7 4. 3-5. Conclusion............................................................................................................ 7 8. Chapter 4 Summary .....................................................................................................8 9 References .....................................................................................................................9 1. I.
(4) Table of contents Table 1-1 Description of adhesion molecules in atherosclerosis ······································24 Table 2-1 Antioxidative capacities of curcumin in vitro ···················································48. II.
(5) Figure of contents. Figure 1-1 Endothelial dysfunction in atherosclerosis··················································· 16 Figure 1-2 Fatty-streak formation in atherosclerosis. ···················································· 17 Figure 1-3 Formation of an advanced, Complicated Lesion of Atherosclerosis. ··········· 18 Figure 1-4 Unstable fibrous plaques in atherosclerosis. ················································ 19 Figure 1-5 Role of the different adhesion molecule in atherosclerosis. ························· 20 Figure 1-6 The signal transduction Pathway of NF-κB activation································· 21 Figure 1-7 Structure of curcumin ·················································································· 22 Figure 1-8 Structure of carnosic acid. ··········································································· 23 Figure 2-1 Cytotoxic effect of curcumin on HUVECs with MTT test··························· 49 Figure 2-2 Effect of curcumin on IL-1 β-induced the adhesion of U937 cells to HUVECs ······································································································································ 50 Figure 2-3 Western blot analysis of ICAM-1 protein levels in cultured HUVECs ········ 52 Figure 2-4 Western blot analysis of VCAM-1 protein levels in cultured HUVECs······· 53 Figure 2-5 Western blot analysis of E-selectin protein levels in cultured HUVECs ······ 54 Figure 2-6 Effect of curcumin on IL-1 β-induced activation of NF-κB p65 in HUVECs ······································································································································ 55 Figure 2-7 Effect of curcumin on IL-1 β-induced activation of NF-κB p50 in HUVECs ······································································································································ 56 Figure 2-8 Effect of curcumin on IL-1 β-induced ROS production in HUVECs··········· 57 Figure 3-1 Cytotoxic effect of carnosic acid on HUVECs with MTT test····················· 79 Figure 3-2 Effect of curcumin on IL-1 β-induced the adhesion of U937 cells to HUVECs ······································································································································ 80. III.
(6) Figure 3-3 Western blot analysis of ICAM-1 protein levels in cultured HUVECs ········ 82 Figure 3-4 Western blot analysis of VCAM-1 protein levels in cultured HUVECs······· 83 Figure 3-5 Western blot analysis of E-selectin protein levels in cultured HUVECs ······ 84 Figure 3-6 Effect of curcumin on IL-1 β-induced activation of NF-κB p65 in HUVECs ······································································································································ 85 Figure 3-7 Effect of curcumin on IL-1 β-induced activation of NF-κB p50 in HUVECs ······································································································································ 86 Figure 3-8 Effect of curcumin on IL-1 β-induced ROS production in HUVECs··········· 87 Figure 4-1 The role of curcumin and carnosic acid in prevention of atherosclerosis····· 90. IV.
(7) Appendix. Table 1 Antioxidative capacities of carnosic acid in vitro ............................................. 101. V.
(8) 中文摘要 內皮細胞表達細胞黏附因子,以及單核球黏附到內皮細胞上,在動 脈粥狀硬化形成的過程中扮演一重要的角色。薑黃素 (curcumin) 是一 種存在薑黃中的多酚類化合物,並且是常用在咖哩的辛香料,先前有研 究顯示薑黃素具有抗氧化的特性。鼠尾草酸 (carnosic acid) 則是存在於 鼠尾草和迷迭香中的多酚類化合物,在過去實驗指出鼠尾草酸也是一種 抗氧化劑。在此次實驗中,主要探討薑黃素和鼠尾草酸個別對於 IL-1 β 所誘導人類臍靜脈內皮細胞黏附因子的表現之影響,以及其影響作用的 機制。結果顯示,薑黃素和鼠尾草酸在體外試驗均有良好的抗氧化能 力,並且薑黃素和鼠尾草酸都可以顯著降低 IL-1 β 所誘導之人類單核球 細胞 (U937) 之黏附。藉由西方點墨法實驗顯示,薑黃素和鼠尾草酸可 降低 IL-1 β 所誘導之 ICAM-1、VCAM-1 和 E-selectin 蛋白質表現。薑 黃素和鼠尾草酸也能減低或阻斷 IL-1 β 所誘導之核轉錄因子 NF-κB p50、 p65 的活化,同時發現薑黃素和鼠尾草酸亦能減少 IL-1 β 所誘導 之細胞中 ROS 的生成。此研究結果證實薑黃素和鼠尾草酸可以降低黏 附因子的表現與單核球細胞的黏附,是藉由抑制 NF-κB P50, P65 的活 化。因此,薑黃素和鼠尾草酸具有抗發炎的特性,並且在預防動脈粥狀 硬化中扮演一重要角色。. 1.
(9) 關鍵字:薑黃素 (curcumin)、鼠尾草酸(carnosic acid)、黏附因子、動脈 粥狀硬化,核轉錄因子-κB,活性氧成分。. 2.
(10) Abstract Expression of cell adhesion molecules by the endothelium and the attachment of monocytes to endothelium may play a major role in the early atherogenic process. Curcumin, which is consumed daily by millions of people, is a polyphenol derived from the plant Turmeric (Curcuma longa) and is used as a spice to give a specific flavor and yellow color to curry. Previous studies indicated that curcumin possesed antioxidant activity in vitro. Carnosic acid, an antioxidant polyphenol derived from Sage (Salvia officinalis) and Rosemary (Rosmarinus officinalis), is a lipophilic antioxidant that scavenges singlet oxygen, hydroxyl radicals, and lipid peroxyl radicals, thus prevents lipid peroxidation and the disruption of biological membranes. In this study, we investigated the antioxidative effects of curcumin and carnosic acid using the TEAC assay, the inhibition of LDL oxidation and DPPH radicals scavenging ability. The effects of curcumin and carnosic acid on the adhesion of U937 cells to IL-1 β-treated human umbilical vein endothelial cells (HUVECs), as well as the expression of adhesion molecules were also examined. In addition, we observed the effect of curcumin and carnosic acid on NF-κB p50, p65 and the formation of intracellular ROS induced by IL-1 β in HUVECs. Our data showed that both of curcumin and carnosic acid had antioxidative effect in vitro. We also found that curcumin and carnosic acid inhibited the adhesion of U937 cells, abolished the expression of ICAM-1, VCAM-1 and E-selectin, attenuated or block nuclear translocation of NF-κB p50, p65 and decreased the production of ROS in IL-1 β treated HUVECs. In conclusion, curcumin and carnosic acid inhibits IL-1 β-induced nuclear translocation of p65 and p50, thereby suppressing the expression of ICAM-1, VCAM-1 and E-selectin, resulting in decreased monocyte adhesion. Thus, curcumin and carnosic have anti-inflammatory properties and may play 3.
(11) an important role in the prevention of atherosclerosis. Key word: curcumin, carnosic acid, adhesion molecule, atherosclerosis, NF-κB, ROS. 4.
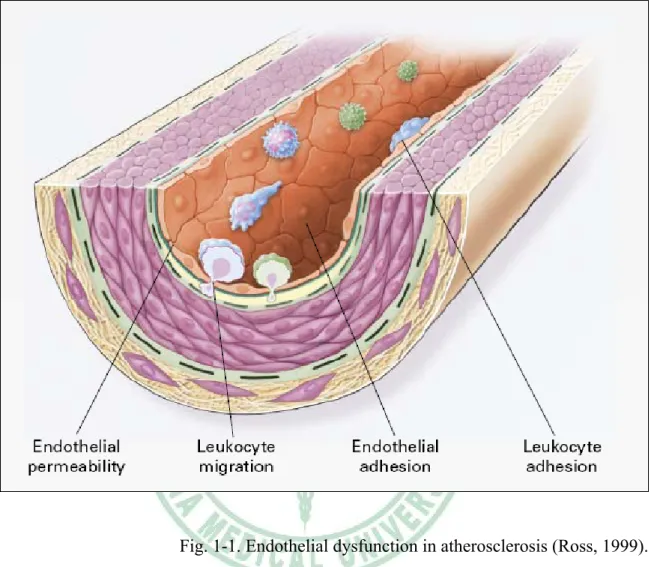
(12) Chapter 1. Introduction. 1.1 Atherosclerosis. Atherosclerosis is the most common cause of death in the Western world, and it is a complex, multifarious disease (Glass & Witztum, 2001). Recent insights into the pathogenesis of atherosclerosis suggest that it may be viewed as a chronic inflammatory disease and inflammatory processes are instrumental in all stages of this disease (Medford, 1995; Offermann & Medford, 1994; Ross, 1995). Ross, 1999 showed that atherosclerosis has four stages. Stage 1: Endothelial dysfunction in atherosclerosis. This stage is the earliest changes that precede the formation of lesions of atherosclerosis take place in the endothelium. These changes include 1.) increased endothelial permeability to lipoproteins and other plasma constituents, which is mediated by nitric oxide, prostacyclin, platelet-derived growth factor (PDGF), angiotensin II, and endothelin; 2.) the up-regulation of leukocyte adhesion molecules, including L-selectin, integrins, and platelet-endothelial-cell adhesion molecule 1 (PECAM-1), and the up-regulation of endothelial adhesion molecules including E-selectin, P-selectin, intercellular adhesion molecule 1 (ICAM-1), and vascular-cell adhesion molecule 1 (VCAM-1); 3.) and the migration of leukocytes into the artery wall, which is mediated by oxidized low-density lipoprotein (ox-LDL), monocyte chemotactic protein 1 (MCP-1), interleukin-8 (IL-8), platelet-derived growth factor (PDGF),. 5.
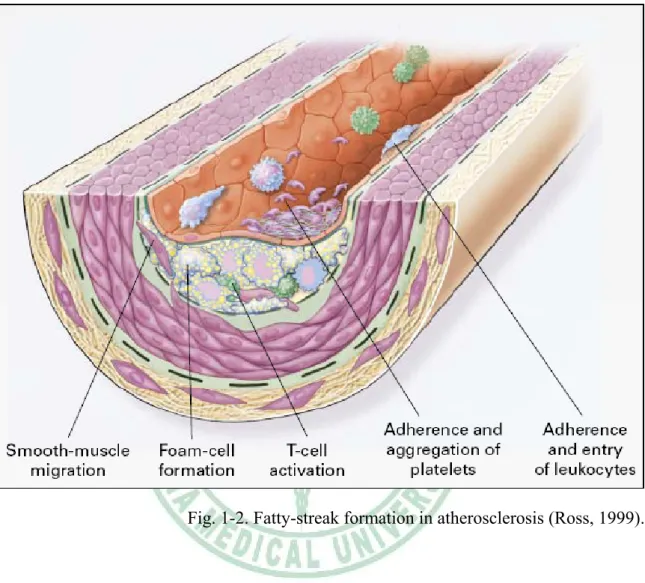
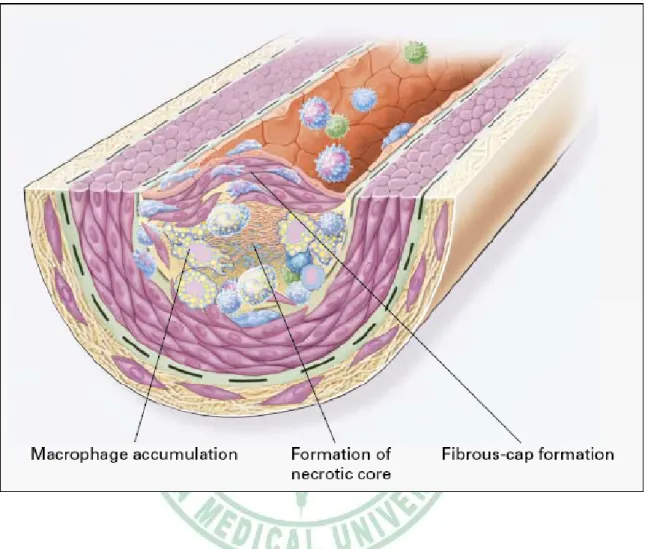
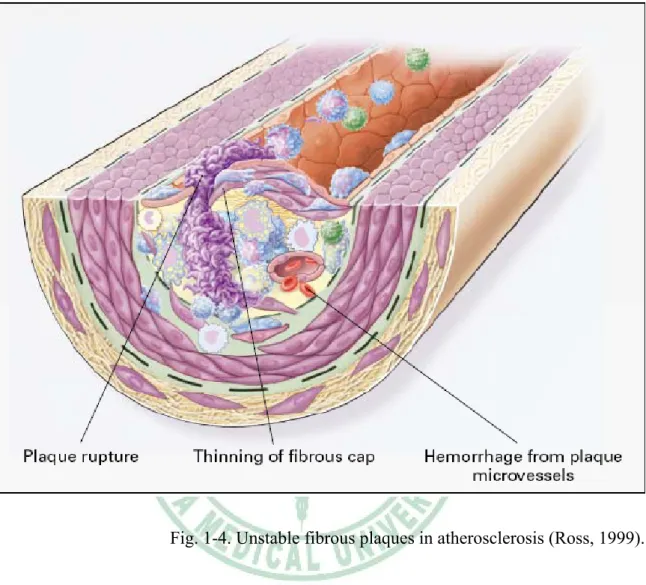
(13) macrophage colony-stimulating factor (MCSF), and osteopontin (Fig 1-1). Stage 2:Fatty-streak formation in atherosclerosis. Fatty streaks initially consist of lipid-laden monocytes and macrophages (foam cells) together with T lymphocytes. Later they are joined by various numbers of smooth muscle cells. These steps include 1.) smooth muscle migration, which is stimulated by PDGF, fibroblast growth factor 2, and transforming growth factor β (TGF-β); 2.) T-cell activation, which is mediated by tumor necrosis factor a (TNF-α), interleukin-2 (IL-2), and granulocyte-macrophage colony -stimulating factor (GM-CSF); 3.) foam cell formation, which is mediated by ox-LDL, MCSF, TNF-a, and interleukin-1 (IL-1); and 4.) platelet adherence and aggregation, which are stimulated by integrins, P-selectin, fibrin, thromboxane A2, tissue factors, and the factors described as responsible for the adherence and migration of leukocytes (Fig 1-2). Stage 3:Formation of an advanced, complicated lesion of atherosclerosis. As fatty streaks progress to intermediate and advanced lesions, they tend to form a fibrous cap that walls off the lesion from the lumen. This represents a type of healing or fibrous response to the injury. The fibrous cap covers a mixture of leukocytes, lipid, and debris, which may form a necrotic core. These lesions expand at their shoulders by means of continued leukocyte adhesion and entry caused by the same factors as stage 1 and 2. The principal factors associated with macrophage accumulation include MCSF, MCP-1, and ox-LDL. The necrotic core represents the results of apoptosis and necrosis, increased proteolytic activity, and lipid accumulation. The fibrous cap forms as a result of increased activity of PDGF, TGF-β, IL-1, TNF-a, and osteopontin and decreased connective tissue degradation (Fig 1-3). Stage 4:Unstable fibrous plaques in atherosclerosis. Rupture of the fibrous cap or. 6.
(14) ulceration of the fibrous plaque can rapidly lead to thrombosis and usually occurs at sites of thinning of the fibrous cap that covers the advanced lesion. Thinning of the fibrous cap is apparently due to the continuing influx and activation of macrophages, which release metalloproteinases and other proteolytic enzymes at these sites. These enzymes cause degradation of the matrix, which can lead to hemorrhage from the vasa vasorum or from the lumen of the artery and can result in thrombus formation and occlusion of the artery (Fig 1-4).. 1.2 Adhesion molecules and atherosclerosis. Cellular adhesion molecules are, according to the definition of the Gene Ontology Consortium (http://www.geneontology. org/) “molecules expressed on the surface of a cell that mediate the adhesion of the cell to other cells or to the extracellular matrix”. These proteins are playing numerous crucial functions at the interface of a cell and its environment, whether this environment is another cell from a similar or different cell type (Blankenberg et al., 2003). In the cardiovascular system, the adhesion of cells to each other and to the proteins of the extracellular matrix provides a stable environment for cell growth, differentiation, and migration. Adhesive interactions are also involved in the pathogenesis of cardiovascular disease, regulating thrombus formation, facilitating leukocyte infiltration, mediating the migration and proliferation of smooth muscle cells, and enabling the deposition of fibrotic tissue (Hillis, 1998). Three main groups of adhesion molecules have been implicated in cardiovascular pathology: integrins, selectins, and members of the immunoglobulin superfamily (Table 1-1). 7.
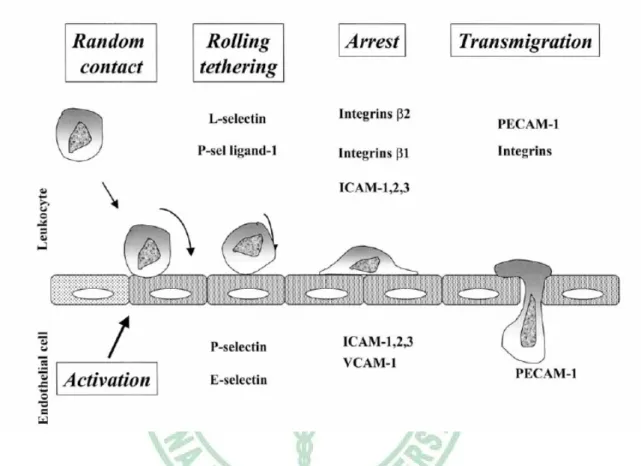
(15) Integrins are glycoproteins composed of two subunits, the α and β chains. As their name suggests they mediate the adhesion of cells to each other and to surround substrates, “integrating” the cell with its environment (Hynes, 1987). In addition, they straddle the cell membrane and play an important role in initiating intracellular events in response to extracellular stimuli (Hynes, 1992). Usually integrins have multiple ligands (Table 1-1). Among these ligands, many extracellular matrix proteins like laminins, collagens, fibronectin, etc., are known to interact with one or more integrins. Selectins are a family of three different glycoproteins sharing a conserved structure and these proteins contain an N-terminal C-type lectin domain (Blankenberg et al., 2003). Unlike other adhesion receptors, selectins bind to carbohydrates and glycopeptides rather than protein ligands (Hillis, 1998) The three members of the selectin family are named according to their main expression site: L-selectin is expressed in leukocytes, E-selectin is specific to endothelial cells and P-selectin is mainly found on platelets but is also expressed in endothelial cells (Bevilacqua & Nelson, 1993). All selectins participate in different, though overlapping, ways to the early steps of leukocyte recruitment at the endothelial surface under shear forces: leukocyte rolling and tethering (Hillis, 1998 ; Blankenberg et al., 2003) (Fig 1-5). The immunoglobulin gene superfamily such as the ICAM-1, ICAM-2 and ICAM3; VCAM-1 and PECAM-1 are principally involved in the adhesion of leucocytes (Hillis, 1998) (table 1-1). Immunoglobulin molecules contain a series of immunoglobulin-like domains of 90 to 100 amino acids that function as endothelial ligands for the integrins expressed on leukocytes and platelets (Muller et al., 1993). ICAM-1 to 3 molecules mediate adhesion of leukocytes to activated endothelium by establishing strong bonds with integrins and inducing firm arrest of inflammatory cells at the vascular surface, and. 8.
(16) participate to leukocyte extravasation (Fig 1-5). VCAM-1 participates to the recruitment of blood cells by activated endothelium by favoring their firm adhesion (Blankenberg et al., 2003) (Fig 1-5). Previous studies have shown that the first step in atherosclerosis development is the adhesion of monocytes to the vascular endothelium (Ross, 1999). The importance of the inflammatory response and the cellular interactions mediated by adhesion molecules in the development of the fatty streak and the fibrous plaque is also evident from experimental data and pathological observations (Daniel et al., 1999). Proinflammatory cytokines such as TNF-α and IL-1 β, commonly found in atherosclerotic lesions, can induce chemotactic factors, other cytokines, and cell adhesion molecules, all of which contribute to the inflammatory process (Zibara et al., 2000; Ross, 1999; Springer, 1994). Previous report has indicated that E-selectin is synthesized by endothelial cells in response to IL-1 β and TNF-α (Bevilacqua et al., 1985). ICAM expression is induced by cytokines, such as TNF-α and IL-1β, or by adherence to extracellular matrix (Dustin et al., 1986; Pober et al., 1986; Daniel et al., 1999) and VCAM-1 is similarly upregulated by IL-1 β and TNF-α (Pober et al., 1986; Marui. et al., 1993).. 1.3 NF-κB and atherosclerosis. Nuclear factor kappa-B (NF-κB) is a redox-sensitive transcription factor regulating a battery of inflammatory genes; it has been implicated as an important factor for initiation and progression of pathogenesis in atherosclerosis, inflammatory bowel disease, autoimmune arthritis, glomerulonephritis, asthma, lung fibrosis, septic shock, carcinogenesis and AIDS (Barnes & Adcock, 1997; Chen et al., 1999; Ghosh et al., 9.
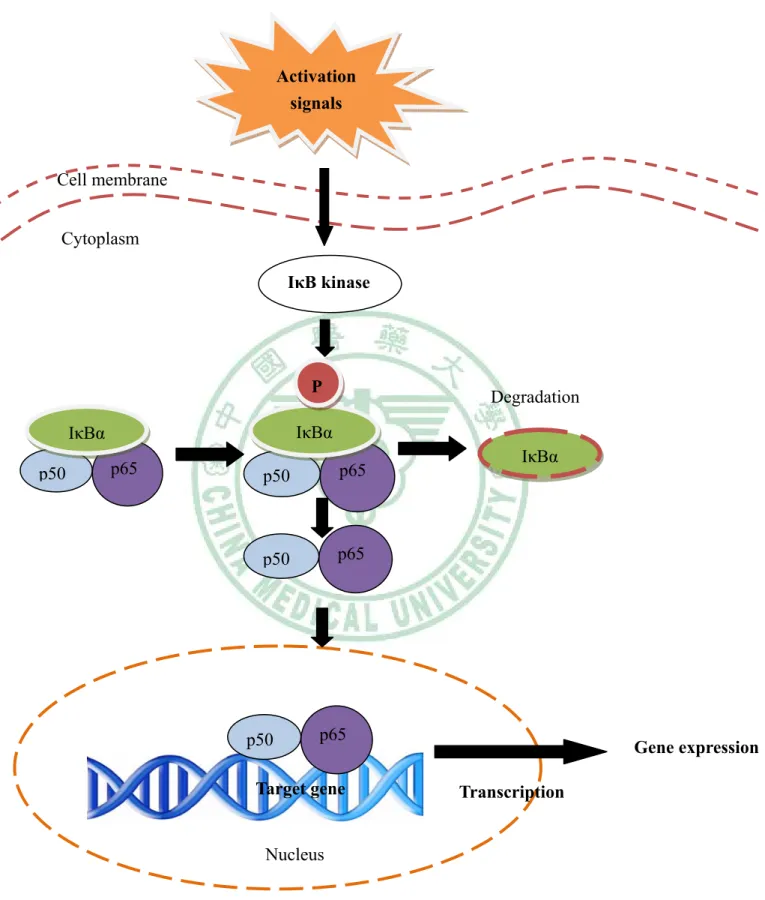
(17) 1998 ). NF-κB was first identified as a regulator of the expression of the kappa light-chain gene in murine B lymphocytes (Sen & Baltimore, 1986). The NF-κB/Rel family includes NF-κB1 (p50/p105), NF-κB2 (p52/p100), p65 (RelA), RelB, and c-Rel NF-κB (Chen et al., 1999), and most members of this family (RelB being one exception) can be homodimers or heterodimers with each other. The activated form of NF-κB is a heterodimer, which usually consists of two proteins, a p65 (also called relA) subunit and a p50 subunit. Other subunits, such as rel, relB, v-rel, and p52, may also be part of activated NF-κB, and it is likely that the different forms of NF-κB may activate different sets of target genes. In unstimulated cells, NF-κB exists in the cytoplasm in an inactive form and is bound to IκBα and IκBβ, which prevent it from entering the nuclei (Baldwin, 1996). Agents that activate NF-κB induce specific phosphorylation events on IκB via IκB kinase activity, which direct IκB to a ubiquitination/proteosomal degradation pathway. Degradation of IkB thus unmasks the nuclear localization sequence of NF-kB and allows NF-κB to enter the nucleus and bind to specific DNA containing the sequence that activates gene transcription (5'-GGGPuNNPyPyCC-3'), where Pu = purine, N = any base; and Py = pyrimidine (Baeuerle & Henkel, 1996) (Fig. 1-6). Many stimuli activate NF-κB, including reactive oxygen intermediates, hypoxia/anoxia, hyperoxia, cytokines, protein kinase C activators, mitogen-activated protein kinase (MAPK) activators, bacterial or viral products, such as lipopolysaccharide (LPS), dsRNA, or the human T-cell leukemia virus type 1 Tax protein, and by UV-irradiation (Barnes & Adcock, 1997; Li et al., 1997; Chen et al., 1999; Ghosh et al., 1998; Thurberg & Collins, 1998; Li & Karin, 1999; Li et al., 1999). NF-κB regulates the expression of many genes involved in both innate and adaptive immunity, among them. 10.
(18) pro-inflammatory. cytokines,. chemokines,. leukocyte. adhesion. molecules. and. inflammatory enzymes (Barnes & Adcock, 1997; Chen et al., 1999; Ghosh et al., 1998; Thurberg & Collins, 1998). It is well established that NF-κB in the development of atherosclerotic injury, mediates cell migration, endothelial cell activation and controls the balance between cell proliferation and apoptosis (Tak & Firestein, 2001). Several of the cytokines and growth factors found in the atherosclerotic lesion, such as TNF-α and IL-1β, activate NF-kB in vitro in relevant cell types such as macrophages, SMCs, ECs, and lymphocytes. The activation of NF-κB in endothelial cells is associated with the activation of genes responsible of an increased transcription of adhesion molecules, cytokines and chemokines, which function in regulating critical processes in atherogenesis (True et al., 2000; Valen et al., 2001; Thornburg et al., 2003; Hatada et al., 2003). 1.4 Oxidative Stress and atherosclerosis. The "response to injury" hypothesis developed by Russell Ross in the late 1970s suggested that atherosclerosis resulted from an initial injury to endothelial cells, leading to impaired endothelial function and subsequent macrophage infiltration and smooth muscle dysfunction. More recently, it has become apparent that reactive oxygen species (ROS) play central roles in the pathogenesis of endothelial dysfunction and atherosclerosis (Harrison et al., 2003), the most important of which are nitric oxide (NO•), superoxide (O2–•), hydrogen peroxide (H2O2) and peroxynitrite (ONOO–•). Recent studies have begun to implicate ROS in the cellular responses to inflammatory cytokines such as TNF-α, IL-1 β (Chua et al., 1998; Simon et al., 1998), whereas other studies have 11.
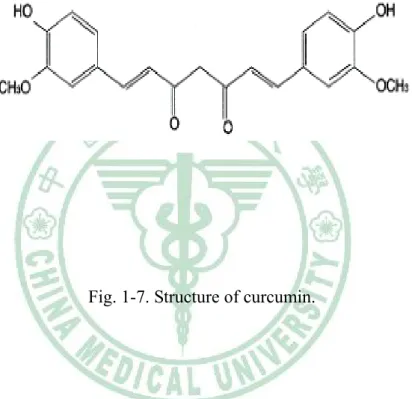
(19) demonstrated that ROS participates in the intracellular signaling initiated during physiological hypoxia (Chandel et al., 1998; Duranteau et al., 1998). ROS in both of these cellular responses suggests that cytokines and hypoxia may interact in the regulation of endothelial barrier function during inflammation (Charles & Russell, 1999). Previous study indicated that the activation mechanism of IκB degradation and NF-κB nuclear translocation have been suggested to involve ROS (Schreck et al., 1991, 1992; Suzuki et al., 1993). Other studies have demonstrated that ROS plays a role as second messengers to regulate signal transduction pathways that ultimately control gene expression and posttranslational modifications of proteins (Charles & Russell, 1999).. 1.5 Curcumin. Curcumin (Fig 1-7), which is consumed daily by millions of people, is a polyphenol derived from the plant turmeric (Curcuma longa) (Fang et al., 2005) and is used as a spice to give a specific flavor and yellow color to curry. In the Indian subcontinent and Southeast Asia, turmeric has traditionally been used as a treatment for inflammation, skin wounds, and tumors (Christopher et al., 2002). Curcumin exhibits a variety of pharmacological effects including antitumor, anti-inflammtory, anti-infectious activities and is currently in clinical trials for AIDS patients (Mazumder et al., 1995; Ruby et al., 1995; Surh, 2002). Molecular actions of curcumin to elucidate these therapeutic effects have been associated with a variety of signaling pathways such as NF-κB, AP-1, and the early growth response gene (EGR-1) (Huang et al., 1995; Pendurthi & Rao, 2000; Grandjean et al., 2002). Curcumin has been shown to inhibit the expression of a sequence of inflammatory cytokines such as TNF-α, IL-1, or IL-8 (Surh, 12.
(20) 2002). Previous study also indicated that supplementation with curcuma longa reduced oxidative stress and attenuated the development of fatty streaks in rabbits fed a high cholesterol diet (Quiles et al., 2002).. 1.6 Carnosic acid. Sage (Salvia officinalis) and Rosemary (Rosmarinus officinalis), popular Labiatae herbs, are increasingly used as food additives with very high antioxidant activity, they have been. proposed as important human dietary factors and inhibitors of skin. tumorgenesis (Singletary & Nelshoppen, 1991; Schwarz et al., 1992; Huang et al., 1994). Carnosic acid (Fig 1-8), an antioxidant polyphenol derived from these plants, is a lipophilic antioxidant that scavenges singlet oxygen, hydroxyl radicals, and lipid peroxyl radicals, thus preventing lipid peroxidation and disruption of biological membranes (Aruoma et al., 1992; Haraguchi et al., 1995). Its radical scavenging activity follows a mechanism analogous to these of other antioxidants such as. -tocopherol and is caused. by the presence of two O-phenolic hydroxyl groups found at C11 and C12 of the molecule (Richheimer et al., 1999). Previous studies indicated that carnosic acid augmented the inhibition of growth and the induction of differentiation of HL60 and U937 leukemia cells by 1 ,25(OH)2D3 and all-trans-retinoic acid (ATRA) (Danilenko et al., 2000). Previous study also demonstated that carnosic acid could inhibit plasma triglyceride elevation in olive oil-loaded mice and reduce the gain of body weight and the accumulation of epididymal fat weight in high fat diet-fed mice (Kiyofumi et al., 2004).. 13.
(21) 1.7 The purpose of this research. Atherosclerosis, a progressive pathological disorder leading to cardiovascular and cerebrovascular diseases, is still the leading cause of mortality and morbidity in industrialized countries, in spite of improved pharmacological and lifestyle approaches (Ross, 1993). The adhesion of circulating leukocytes to the vascular endothelium is a critical early event in the development of atherosclerosis (Joris et al., 1983; Faggiotto et al., 1984). This process depends on the interaction between cell adhesion molecules expressed on the surface of endothelial cells and their cognate ligands on leukocytes (Price & Loscalzo, 1999). Previous studies have indicated that NF-κB/Rel transcription factors may play an important role in the development of the atherosclerosis (Collins, 1993; Qwarnstrom et al., 1994). The activation of NF-κB in endothelial cells is associated with the activation of genes responsible of an increased transcription of adhesion molecules, cytokines and chemokines (True et al., 2000; Valen et al., 2001; Thornburg et al., 2003; Hatada et al., 2003). Plant polyphenols are a large group of naturally-occuring antioxidants and epidemiological studies have suggested that higher polyphenol intake from fruits and vegetables is associated with decreased risk for cardiovascular disease (Ilja & Peter, 2005). Previous studies showed that polyphenolic compounds such as dietary flavonoids or red wine polyphenols could prevent atherosclerosis by inhibiting adhesion molecules expression in endothelial cell (Murase et al., 1999; Silvina & Balz, 2006). Curcumin, which is consumed daily by millions of people, is a polyphenol derived from the plant turmeric (Curcuma longa) (Fang et al., 2005) and is used as a spice to give a specific flavor and yellow color to curry. Curcumin exhibits a variety of pharmacological effects 14.
(22) including antitumor, anti-inflammatory, anti-infectious activities and is currently in clinical trials for AIDS patients (Mazumder et al., 1995; Ruby et al., 1995; Surh, 2002). Previous study also indicated that supplementation with curcuma longa reduced oxidative stress and attenuated the development of fatty streaks in rabbits fed a high cholesterol diet (Quiles et al., 2002). Carnosic acid, an antioxidant polyphenol derived from Sage (Salvia oddicinalis) and Rosemary (Rosmarinus officinalis), is a lipophilic antioxidant that scavenges singlet oxygen, hydroxyl radicals, and lipid peroxyl radicals, thus preventing lipid peroxidation and disruption of biological membranes (Aruoma et al., 1992; Haraguchi et al., 1995). Previous study also demonstrated that carnosic acid can inhibit plasma triglyceride elevation in olive oil-loaded mice and reduce the gain of body weight and the accumulation of epididymal fat weight in high fat diet-fed mice (Kiyofumi et al., 2004). Therefore, we designed to examine the inhibitory effects of curcumin and carnosic acid on monocyte adhesion to cultured human endothelial cells and the expression of adhesion molecules (VCAM-1, ICAM-1 and E-selectin) by IL-1 β in HUVECs and elucidate its possible mechanism of action.. 15.
(23) Fig. 1-1. Endothelial dysfunction in atherosclerosis (Ross, 1999).. 16.
(24) Fig. 1-2. Fatty-streak formation in atherosclerosis (Ross, 1999).. 17.
(25) Fig. 1-3. Formation of an advanced, complicated lesion of atherosclerosis (Ross, 1999).. 18.
(26) Fig. 1-4. Unstable fibrous plaques in atherosclerosis (Ross, 1999).. 19.
(27) Fig. 1-5. Role of the different adhesion molecule in atherosclerosis. (Blankenberg et al., 2003). 20.
(28) Activation signals. Cell membrane Cytoplasm IκB kinase. P IκBα. IκBα p50. Degradation. p65. p50. p65. p50. p65. p50. IκBα. p65. Gene expression. Target gene. Transcription. Nucleus Fig. 1-6. The signal transduction pathway of NF-κB activation (Baeuerle & Henkel, 1996).. 21.
(29) Fig. 1-7. Structure of curcumin.. 22.
(30) Fig.1-8. Structure of carnosic acid.. 23.
(31) Table 1-1. ( Blankenberg et al., 2003 ). 24.
(32) Chapter 2 Experimentation I. Curcumin attenuates the expression of adhesion molecules by IL-1 β in HUVECs. 薑黃素下降介白素-1 β 所誘發的人類臍靜脈內皮細胞黏附 因子表現. 25.
(33) 2.1 Prolegomenon. Atherosclerosis, a progressive pathological disorder leading to cardiovascular and cerebrovascular diseases, is still the leading cause of mortality and morbidity in industrialized countries, in spite of improved pharmacological and lifestyle approaches (Ross, 1993). The adhesion of circulating leukocytes to the vascular endothelium is a critical early event in the development of atherosclerosis (Joris et al., 1983; Faggiotto et al., 1984). This process depends on the interaction between cell adhesion molecules expressed on the surface of endothelial cells and their cognate ligands on leukocytes (Price & Loscalzo, 1999). Previous studies have indicated that NF-κB/Rel transcription factors may play an important role in the development of the atherosclerosis (Collins, 1993; Qwarnstrom et al., 1994). The activation of NF-κB in endothelial cells is associated with the activation of genes responsible of an increased transcription of adhesion molecules, cytokines and chemokines (True et al., 2000; Valen et al., 2001; Thornburg et al., 2003; Hatada et al., 2003). Plant polyphenols are a large group of naturally-occuring antioxidants and epidemiological studies have suggested that higher polyphenol intake from fruits and vegetables is associated with decreased risk for cardiovascular disease (Ilja & Peter, 2005). Previous studies showed that polyphenolic compounds such as dietary flavonoids or red wine polyphenols could prevent atherosclerosis by inhibiting adhesion molecules expression in endothelial cell (Murase et al., 1999; Silvina & Balz, 2006). Curcumin, which is consumed daily by millions of people, is a polyphenol derived from the plant turmeric (Curcuma longa) (Fang et al., 2005) and is used as a spice to give a specific flavor and yellow color to curry. Curcumin exhibits a variety of pharmacological effects including antitumor, anti-inflammatory, 26.
(34) anti-infectious activities and is currently in clinical trials for AIDS patients (Mazumder et al., 1995; Ruby et al., 1995; Surh, 2002). Previous study also indicated that supplementation with curcuma longa reduced oxidative stress and attenuated the development of fatty streaks in rabbits fed a high cholesterol diet (Quiles et al., 2002). Therefore, we designed to examine the inhibitory effects of curcumin on monocyte adhesion to cultured human endothelial cells and the expression of adhesion molecules (VCAM-1, ICAM-1 and E-selectin) by IL-1 β in HUVECs and elucidate its possible mechanism of action.. 27.
(35) 2.2 Materials and Methods. 2.2.1 Instruments. CO2 incubator. NUAIRE, MN, USA. Laminar flow. NUAIRE, MN, USA. Microscope. Nikon, Japan. pH meter. HANNA, RI, USA. Stirrer/Hotplate. Corning, Taiwan. Waterbath tank. TKS, Taiwan. Haemocytometer. Boeco, Germany. Eppendorf centrifugator. Hamburg, Germany. Pipetman. Gilson, France. Spectrophotometer. HITACHI, Japan. Spectrophotometer. Beckman Coulter, CA, USA. MicroPlate fluorescence reader. Bio-Tek, VT, USA. Shaking incubator. Orbital, VA, USA. ELISA plate reader. Bio-Tek, VT, USA. PCR thermocycler. Bio-Rad, CA, USA. Electrophoresis tank. Bio-Rad, CA, USA. Transfer system. Bio-Rad, CA, USA. Electrophoresis chamber. Bio-Rad, CA, USA. Power supply. Hoefer, CA, USA. 28.
(36) 2.2.2 Chemicals. ABTS (2,2’-azino-bis(3-ethylbenzthiazoline-6-sulfonic acid)). Sigma, MO, USA. BSA (Bovine serum albmin). Sigma, MO, USA. Bradford reagent. Bio-Rad, CA, USA. CHCl3 (Chloroform). BDH, Poole, England. Curcumin. Sigma, MO, USA. CuSO4 (Cupric sulfate). Sigma, MO, USA. DCFH-DA (2’, 7’-dichlorofluorescin diacetate). Molecular Probe, Oregon, USA. DEPC (Diethyl pyrocarbonate). Sigma, MO, USA. DMSO (Dimethyl sulfoxide). Sigma, MO, USA. DPPH (2, 2-diphenyl-1-picrylhydrazyl). Sigma, MO, USA. DTT (1, 4-Dithio-D,L-thereitol). Bio-Rad, CA, USA. ECG (Endothelial cell growth supplement). Sigma, MO, USA. EDTA-Disodium (Ethyienediamine Teraacetate Disodium Salt). Bio-Rad, CA, USA. FBS (Fetal Bovine serum). Gibco, NY, USA. Gelatin (2% solution Type B). Sigma, MO, USA. Glucose. Sigma, MO, USA. HBSS (Hank’s balanced salts modified). Sigma, MO, USA. HCl (Hydrochloric acid). Wako, Osaka, Japan. Heparin. Sigma, MO, USA. Hepes buffer solution (1 mol/l). Gibco, NY, USA. H2O2 (Hydrogen Peroxide). Wako, Osaka, Japan. KBr (potassium bromide). SHOWA, Tokyo, Japan. 29.
(37) KCl (potassium chloridem). SHOWA, Tokyo, Japan. KH2 PO4 (potassium dihydrogenphosphate). SHOWA, Tokyo, Japan. L-Glutamine (200 mmol/l). Gibco, NY, USA. MDA (malonaldehyde bis-(dimethyl acetal)). Alderich, WI, USA. MTT (3-[4, 5-Dimethylthiazol-2-yl]-2, 5-diphenyl-terazoliumbromide) Sigma, MO, USA Medium 199. Sigma, MO, USA. Methanol. TEDIA, OH, USA. NaCl (sodium chloride). Sigma, MO, USA. NaHCO3 (sodium bicarbonate). Sigma, MO, USA. Na2HPO4 (Disodium hydrogenphosphate). SHOWA, Tokyo, Japan. NF-κB kit. TransAM, Active Motif, CA, USA. Nuclear Extract kit. TransAM, Active Motif, CA, USA. Penicillin-Streptomycin. Gibco, NY, USA. Peroxidase (Type VI-A). Sigma, MO, USA. Recombinant human interleukin-1 beta. ProSpec-Tany, Rehovot, Isreal. RPMI-1640. Gibco, NY, USA. TBA (2-Thiobarbituric acid). Sigma, MO, USA. TCA (Trichloroacetic acid). Sigma, MO, USA. Trolox (6-Hydroxy-2,5,7,8-treamethylchroman-2-carboxylic acid) Alderich, WI, USA Trypsin-EDTA (0.25 %). Gibco, NY, USA. SDS (Sodium dodecyl sulfate). Bio-Rad, CA, USA. SYBR Green suoermix. Bio-Rad, CA, USA. 30.
(38) 2-propanol. Sigma, MO, USA. 95 % Ethanol. Echo Chemical, Taichung, Taiwan. Mouse anti-human β-actin. Abcam, Cambridge, UK. Mouse anti-human ICAM-1. Abcam, Cambridge, UK. Mouse anti-human VCAM-1. Abcam, Cambridge, UK. Mouse anti-human E-selectin. Abcam, Cambridge, UK. Sheep anti-mouse IgG antibody. Abcam, Cambridge, UK. 2.2.3 Antioxidative capacities of curcumin in vitro. 2.2.3.1 DPPH scavenging assay. Free radical scavenging effect was determined using the free radical generator DPPH (2, 2-diphenyl-1-picrylhydrazyl) by a similar method to Yamaguchi et al. 1998. Briefly, the reaction mixture contained 500 μl of curcumin concentrations (0–200 μmol/l) and 500 μl of DPPH (0.5 mmol/l in methanolic solution). The DPPH radical scavenging activity was evaluated by measuring the decrease of DPPH radical detected at 517 nm and by determining the difference in the peak area between control and reaction mixture. Inhibition % was calculated via Eq. Inhibition % = (B0-B1 / B0) × 100 (where B0: is the absorbance of control. B1: is the absorbance of reaction mixture.) The decoloration was plotted against the sample extract concentration in order to calculate the IC50 values (inhibitory concentration 50 μmol/l), which is the amount of sample needed to decrease the absorbance of DPPH by 50%. 31.
(39) 2.2.3.2 Trolox equivalent antioxidant capacity (TEAC) assay. The assay was carried out using a Spectronic Genesys 5 spectrophotometer by the improved ABTS Briefly, ABTS. +. method as described by Miller et al., (1993) with slight modification. +. radical cation was generated by a reaction of 500 μmol/l. 2,2’-azino-bis(3-ethylbenzthiazoline-6-sulfonic acid) (ABTS) and 450 μmol/l hydrogen peroxide. The reaction mixture was allowed to stand in the dark for 6 min at room temperature and used within 2 days. All samples were diluted appropriately to provide 20–80% inhibition of the blank absorbance. Fifty microliters of the diluted sample were mixed with 1.9 ml of diluted ABTS + solution. The assay with the mixture was carried out in triplicate, the mixture was allowed to stand for 6 min at room temperature and the absorbance was immediately recorded at 734 nm (Li et al., 2007). Trolox solution (final concentration 0~15 μmol/l) was used as a reference standard. A value of 1 TEAC in a sample is defined as a concentration equivalent to 1 μmol/l Trolox, a water-soluble analog of α-tocopherol.. 2.2.3.3 Inhibition of LDL Oxidation. Blood was collected from healthy donors after a 12 h overnight fasting using EDTA as anticoagulant. Plasma was obtained after low-speed centrifugation of the blood, adjusted to a density of 1.21 g/ml with KBr and a discontinuous density gradient was made by overlaying the plasma solution with a 10 mmol/l PBS and 1 mmol/l EDTA pH 7.4. LDL were isolated after ultracentrifugation as described previously (Vieira et al, 1996). The LDL fraction was exhaustively dialyzed against PBS buffer pH 7.4 without. 32.
(40) EDTA in the dark, filtered through a 0.45 μm filter, stored at 4 °C under nitrogen and used in 24-72 h. LDL were oxidised using the classical copper-induced LDL auto-oxidation (Wallin et al., 1993). Briefly, 0.9 mg/ml of LDL total cholesterol was incubated in PBS in the presence of CuSO4 (50 μmol/l). Increasing concentrations of curcumin (0–10 μmol/l) dissolved in DMSO were present in the incubation media. After incubation, 150 µL EDTA (2 mmol/l) was added. A 100 µl portion of the mixture was then transferred to an eppendrof tube containing 0.9 ml of 2-propanol. The precipitates were removed by centrifugation. The concentration of conjugated diene in the supernatant was determined by absorption at 234 nm.. 2.2.4 Cell Culture. Human umbilical vein endothelial cells (HUVECs) were isolated from human umbilical cords using collagenase type II (Jaffe, 1973), and cultured on 0.1 % gelatin-coated culture dishes in medium M199 (Sigma, MO, USA) supplemented with 10 % FBS, 1 % Antibiotic -Antimycotic, Glutamine (2 mmol/l), Heparin (10 U/ml), Hepes (10 mmol/l), Endothelial cell growth supplement (ECG) (12.5 μg/ml) at 37℃ in a humidified atmosphere of 5 % CO2 and 95 % air. After 3 days, the medium was replaced by fresh medium and subcultured at a 1: 4 ratio one time per week. All experiments were performed with HUVEC from passages two to five.. 2.2.5 Cell viability assay (MTT test). The viability of the cells was assessed by MTT (3-[4,5-dimethylthiazol-2-yl] -2,5. 33.
(41) -diphenyl-tetrazolium bromide) assay (Mosmann, 1983), which is based on the reduction of MTT by the mitochondrial dehydrogenase of intact cells to a blue formazan product. Briefly, 1 × 104 cells/well were dispensed within 96-well culture plates and incubated with various concentrations of curcumin (which was dissolved in dimethyl sulphoxide) for 24 h. Four hours before the end of the treatment 10 μl MTT (5 mg/ml) was added to each well. At the end of the treatment the incubation medium was removed and the formazan crystals were dissolved in 100 μl of solution of DMSO. MTT reduction was quantified by measuring the light absorbance with a ELISA plate reader (μQUANT, Bio-Tek, USA) at 590 nm. The reduction in optical density caused by curcumin was used as a measurement of cell viability, normalized to cells incubated in medium with DMSO only, which were considered to be 100 % viable.. 2.2.6 Adhesion of U937 cells to endothelial cells. Adhesion was evaluated using the human leukemia pro-monocytic U937 cells which were labeled with calcein AM (10 nmol/l; Molecular Probe; Invitrogen) (Yu et al., 2007). HUVECs (2 ×105) were distributed into 6-well plates before the assay and allowed to reach confluence. Then the growth medium was supplemented with 10 and 20 μmol/l curcumin for 18 h, followed by incubating 10 ng/ml IL-1 β for 6 h in the continued presence of curcumin. U937 cells were grown in RPMI 1640 medium (Gibco, New York, USA) containing 10 % FBS and subcultured at a 1:5 ratio three times per week. U937 cells were incubated with 10 nmol/l calcein AM in RPMI 1640 medium for 30 min at 37℃, then washed with PBS to remove free dye and resuspended in M-199 mediam containing 10 % FBS. Labeled U937 cells (2×105) were added to each. 34.
(42) HUVEC-containing well and incubation continued for 30 min. Non-adherent cells were removed by two gentle washes with PBS, then the number of bound U937 cells was determined by a fluorescence plate reader at an excitation wavelength of 485 nm and emission at 530 nm; HUVECs cell monnolayers with DMSO only were served as the control.. 2.2.7 Bradford assay. The Bradford assay (Bradford, 1976), a colorimetric protein assay, is based on an absorbance shift in the dye Coomassie when bound to arginine and hydrophobic amino acid residues present in protein. The anionic (bound) form of the dye is blue and has an absorption spectrum maximum historically held to be at 595 nm. The cationic (unbound) forms are green and red. The increase of absorbance at 595 nm is proportional to the amount of bound dye, and thus to the amount (concentration) of protein present in the sample. Standard solutions contain a range of 0 to 25 micrograms protein (BSA) in 800 μl H2O, followed by adding 200 μl dye reagent and incubate 5 min. l μl of sample solution add into 799 μl H2O, followed by adding 200 μl dye reagent and incubated for 5 min. The absorbance was read at 595 nm. The results made a standard curve and the protein concentration of sample was determined by standard curve.. 2.2.8 Western blot. For Western blotting, 3×106 cells were seeded in 10 cm dishes and treated 10 and 20 μmol/l curcumin for 18 h, followed by incubating 10 ng/ml IL-1 β for 6 h in the. 35.
(43) continued presence of curcumin, afterwards cells were scraped with a rubber policeman in PBS and centrifuged at 1,200 rpm for 10 min. Cells were lysed for 1 h at 4 ℃ with lysis buffer (0.32 mol/l Sucrose, 10 mmol/l Tris, 5 mmol/l EDTA, 2 mmol/l DTT, 1 % Triton X-100, and 1 mmol/l PMSF) and centrifuged at 12,000 g for 30 min at 4 ℃. The protein concentration of cell extracts was determined with a Bradford-based assay (Bradford, 1976). Cell extracts were loaded per lane, resolved by 10 % SDS -PAGE and transferred at room temperature by blotting to polyvinylidene difloride (PVDF) membrane (Shishodia et al., 2003). Nonsecific binding was blocked by soacking the membrane in PBS-Tween 20 buffer containing 50 g/L fat-free milk and separately incubated for 1 h at room temperature with mouse anti-human-VCAM-1, ICAM-1, and E-selectin antibodies. Subsequently, the membrane was incubated with a sheep anti-mouse IgG antibody. The protein levels was determined with the enhanced chemiluminescence (Upstate, USA) and High performance chemiluminescence film (Amersham biosciences, USA). Incubation with mouse anti-human β-actin antibody was also performed as an internal control. Results were quantified with scanning densitometer using an image analysis system with software.. 2.2.9 Nuclear extract preparation. Nuclear protein extracts were prepared using a nuclear extract kit (TransAM nuclear extract kit, CA, USA) from HUVECs to assay the NF-κB activity. Nuclear protein were prepared as described previously (Dschietzig et al., 2001). 3×106 cells were seeded in 10 cm dishes and treated 10 and 20 μmol/l curcumin for 18 h, followed by incubating 10 ng/ml IL-1 β for 6 h in the continued presence of curcumin, afterwards. 36.
(44) cells were scraped with a rubber policeman in PBS, collected and centrifuged at 1,200 rpm for 10 min. Cells were lysed in hypotonic buffer and centrifuge suspension for 30 seconds at 14,000×g in a microcentrifuge pre-cooled at 4 °C. Then resuspend nuclear pellet in 50 μl complete lysis buffer containing 10 mmol/l DTT, lysis buffer AM1, and protease inhibitor cocktail by pipetting up and down. The suspension was incubated for 30 min on ice, and centrifuged for 10 min at 14,000×g in a microcentrifuge pre-cooled at 4 °C. Transfer supernatant and stored at -80 °C. The protein concentration was determined with a Bradford-based assay (Bradford, 1976).. 2.2.10 Measurement of NF-κB activity. For analysis of NF-κB activation with the TransAM NF-κB Family kit (TransAM, Active Motif, CA, USA) (Yu et al., 2007). The oligonucleotide containing the NF-κB consensus binding site (5′-GGGACTTTCC-3′) specific for the active form of NF-κB was immobilized to a 96-well plate and the well was filled with 10 μg of nuclear extract. After 1 hour incubation and three washings, the primary antibody against the active form of NF- B recognizing an epitope on p65, p50 that is accessible only when NF- B is activated and bound to its target DNA was added for 1 hour. After washing, the secondary antibody conjugated to horseradish peroxidase was added to achieve a sensitive readout by spectrophotometry at 450 nm and the blank was subtracted from all measurements.. 37.
(45) 2.2.11 Measurement of reactive oxygen species. ROS generation in cells was assessed using the probe 2,7-dichlorofluorescein (DCF) (Molecular Probes Europe BV, Leiden, The Netherlands) according to the method reported by Wang and Joseph (1999). Confluent HUVECs (1×104 cells/well) in 96-well plates were pretreated with 10 and 20 μmol/l curcumin for 18 h, followed by incubation of 10 ng/ml IL-1 β for 6 h in the continued presence of curcumin. After the removal of curcumin or carnosic acid from wells, cells were incubated with 10 μmol/l DCFH-DA for 30 minutes. The fluorescence intensity (relative fluorescence units) was measured at 485-nm excitation and 530-nm emission using a fluorescence microplate reader.. 2.2.12 Statistical analysis. Results are shown as mean ± S.D. Statistical analyses of MTT were performed using One-way ANOVA followed by Dunnett’s test and others were performed using One-way ANOVA followed by Duncan's Multiple Range Test. A value of P <0.05 was considered statistically significant.. 38.
(46) 2.3 Results. 2.3.1 Antioxidative capacities of curcumin in vitro. Free radical scavenging effect was determined using the free radical generator DPPH (2, 2-diphenyl-1-picrylhydrazyl) and the IC50 of DPPH assay was 45.84±0.14 μmol/l. In the inhibition of LDL oxidation assay, LDL were oxidized using the classical copper-induced LDL auto-oxidation and the IC50 of inhibition of LDL oxidation was 2.56±0.12 μmol/l. The TEAC assay is based on the reduction of the ABTS (2,2′-azino-bis-(3-ethylbenzthiazoline-6-sulfonic acid)) radical cation by antioxidants and the TEAC value of curcumin was 2.88±0.04 (Table 2-1). A value of 1 TEAC in a sample is defined as a concentration equivalent to 1 μmol/l Trolox, a water-soluble analog of α-tocopherol.. 2.3.2 Cell viability of curcumin for HUVECs. Cell viability was determined by the MTT test. After 24 h incubation with 10 and 20, 30, 40 and 50 μmol/l curcumin, cell viability was 97.1 ± 5.5, 93.4 ± 3.2, 58.7 ± 4.3, 51.5 ± 3.3 and 48.5 ± 2.8 % of control levels, respectively, with the three highest concentrations causing a significant reduction in cell viability (Fig 2-1). Therefore, according to the MTT test we chose 10 and 20 μmol/l curcumin to do all the following experiments.. 39.
(47) 2.3.3 Curcumin inhibited the adhesion of U937cells to IL-1 β-stimulated HUVECs. In order to determine the effect of curcumin in the adhesion of U937 cells to endothelial cells, HUVECs were treated with 10 and 20 μmol/l curcumin for 18 h, followed by incubation of 10 ng/ml IL-1 β for 6 h and the percentage of cell adhesion was evaluated by the quantification of calcein AM (Fig. 2-2 (B)). The adhesion of U937 cells to HUVECs also was photographed (Fig. 2(A)). The control group showed that minimal binding of HUVECs to U937 cells, but adhesion significantly increased when the HUVECs were treated with IL-1 β (Fig 2-2 A & B). Pretreatment with 10 and 20 μmol/l curcumin could significantly reduce the number of U937 cells adhering to IL-1 β-stimulated HUVECs. The concentration of 20 μmol/l curcumin was more effective on cell adhesion than 10 μmol/l curcumin (Fig 2-2 A & B). These results indicate that curcumin could inhibit monocytes adhesion to endothelial cells.. 2.3.4 Curcumin inhibits IL-1 β-induced cell protein expression of VCAM-1, ICAM-1 and E-selectin in HUVECs. To determine if the inhibition of cell adhesion by curcumin was due to inhibit the expression of adhesion molecules, HUVECs were pretreated for 18 h with 10 and 20 μmol/l curcumin before the addition of 10 ng/ml IL-1 β. The expression of ICAM-1, VCAM-1 and E-selectin was increased after IL-1 β stimulation (Fig 2-3, 2-4, 2-5). Pretreatment of HUVECs with 10 μmol/l curcumin significantly inhibited the expression of ICAM-1 and VCAM-1, but not E-selectin (Fig 2-3, 2-4, 2-5). Pretreatment of. 40.
(48) HUVECs with 20 μmol/l curcumin could significantly inhibit the expression of ICAM-1, VCAM-1 and E-selectin. Therefore, 20 μmol/l curcumin was more effective than 10 μmol/l curcumin on the inhibition of cell adhesion molecular expression (Fig 2-3, 2-4, 2-5).. 2.3.5 Curcumin decrease activation of NF-κB and nuclear translocation of NF-κB p65 and p50 in IL-1 β-stimulated HUVECs. To examine whether the inhibitory effect of curcumin on the cytokine-induced expression of adhesion molecules was medicated via NF-κB, we measured the nuclear translocation of p65 and p50 protein of the NF-κB family. Incubation of IL-1 β (10 ng/ml) for 6 h induced the nuclear translocation of p65 and p50 (Fig. 2-6, 2-7). Pretreatment of HUVECs with 10 and 20 μmol/l curcumin significantly decreased the nuclear translocation of p65 and p50 (Fig. 2-6, 2-7). In addition, pretreatment of HUVECs with 20 μmol/l curcumin was more effective on decreasing the nuclear translocation of p65 and p50 than that of 10 μmol/l curcumin.. 2.3.6 Curcumin inhibits IL-1 β-induced ROS production in HUVECs. To study the effect of curcumin on IL-1 β-induced ROS production in HUVECs, cells were pretreated with 10 and 20 μmol/l curcumin for 18 h and then stimulated by IL-1 β (10 ng/ml) for 6 h. HUVECs were labeled with H2O2-sensitive fluorescent probe and detected by fluorescence microplate reader. The production of ROS induced by IL-1 β was increased and significantly decreased after pretreatment with 10 and 20 μmol/l. 41.
(49) curcumin (Fig. 2-8 A & B). In addition, pretreatment of HUVECs with 20 μmol/l curcumin was more effective on decreasing the production of ROS than 10 μmol/l curcumin.. 42.
(50) 2.4 Discussion. The expression of cell adhesion molecules by the endothelium and the attachment of monocytes to endothelium may play a major role in the early atherogenic process. Curcumin is the principal curcuminoid of the Indian curry spice turmeric. The curcuminoids are polyphenols and are responsible for the yellow color of turmeric. Previous studies indicated that curcumin exhibits a variety of pharmacological effects including antitumor, anti-inflammtory and anti-infectious activities (Mazumder et al., 1995; Ruby et al., 1995; Surh, 2002). In the present study, we found that curcumin had antioxidative effect in vitro. In addition, we also found that curcumin could suppressed IL-1 β-induced intracellular ROS production, the activation of redox-sensitive transcription factors NF-κB p50, p65, the expression of VCAM-1, ICAM-1 and E-selectin; and the adhesiveness to a human monocytic cell line (U937) in HUVECs. These results demonstrated that curcumin had inhibitory effect on proanthersclerotic mechanism in vitro. Curcumin is the most active component of turmeric which contains 2 to 5 % of curcumin. Commercial curcumin is usually isolated from the rhizome of turmeric which contains three major curcuminoids (approximately 77 % curcumin, 17 % demethoxy -curcumin, and 3 % bisdemethoxycurcumin) (Bharat et al., 2005). There are several good sources of curcumin with purity ranging from 60 to 98 % (Bharat et al., 2005). Previous study indicated that the serum concentration was 1.77±1.87 μmol/l after the intake of 8 g curcumin in human (Cheng et al., 2001), therefore, curcumin was absorbable in digestive tract in human. In the present study, we found that 20 μmol/l curcumin did not have any significant effect on the viability of HUVECs from MTT test, therefore, we chose 10 and 43.
(51) 20 μmol/l curcumin to do all the experiments. One of the earliest events in atherogenesis is the adhesion of monocytes to the endothelium, followed by their infiltration and differentiation into macrophages. In the present study, we found that the control group showed minimal binding to U937 cells, but adhesion increased when the HUVECs were treated with IL-1 β. Pretreatment with 10 and 20 μmol/l curcumin reduced the number of U937 cells adhering to IL-1 β-stimulated HUVECs. A similar resulet aws seen when HUVECs were pretreated with other polyphenolic compound, such as vitamin E (40 μmol/l), probucol (50 μmol/l) or tea flavonoid (60 μmol/l), these polyphenolic compound reduced monocytes adhesion to endothelial cells (Islam et al., 1998; Zapolska-Downar et al., 2001; Ludwig et al., 2004). The process by which monocytes become adherent to the endothelium is the result of complex choreography requiring the sequential, yet overlapping, functions of many classes of adhesion molecules (Munro, 1993; Price & Loscalzo, 1999). Tethering and rolling, the first steps of monocyte adhesion to the endothelial surface, appear to depend on the interaction of P- and E- selectin with carbohydrate ligands on leukocytes. Firm adhesion follows if the leukocytes encounter activating signals while rolling along the endothelium, and is facilitated by the interaction of very late antigen-4 (VLA-4) with VCAM-1 or lymphocyte function antigen-1 (LFA-1) with ICAM-1 (Marlin & Springer, 1987; Elices et al., 1990). Previous study showed that adhesion molecules are strong predictors of atherosclerotic lesion development and future cardiovascular events (Blankenberg et al., 2003). It has been reported that expression of adhesion molecules on HUVECs is increased or induced by stimulation with inflammatory cytokines, including TNF-α, IL-1 β, IL-4, and IL-13 (Schleimer & Rutledge, 1986). In the present study, we found that the expression of ICAM-1, VCAM-1 and E-selectin was increased by IL-1 β. 44.
(52) and the pretreatment with curcumin could decreased the inducetion of expression by IL-1 β in HUVECs. These results indicated that curcumin could inhibit the rolling, tethering and firm adhesion of the monocytes on the vascular wall. NF-κB is a redox-sensitive transcription factor which mediates cell migration, endothelial cell activation and the balance between cell proliferation and apoptosis (Tak & Firestein, 2001). The activation of NF-κB in endothelial cells is associated with the activation of genes responsible for an increased transcription of adhesion molecules, cytokines and chemokines (True et al., 2000; Valen et al., 2001; Thornburg et al., 2003; Hatada et al., 2003). The activated form of NF-κB is a heterodimer, which usually consists of two proteins, a p65 (also called relA) subunit and a p50 subunit (Baldwin, 1996). Udalova et al., 2000 described that the p50–p65 heterodimers are involved in enhancing the transcription of adhesion molecules. In the present study, we examined whether the inhibitory effect of curcumin on the cytokine-induced expression of adhesion molecules is medicated via NF-κB, therefore, we measured the nuclear translocation of p65 and p50 protein of the NF-κB family. We found that the nuclear translocation of p65 and p50 induced by IL-1 β was increased and the pretreatment with curcumin could decreased the inducetion of translocation by IL-1 β in HUVECs. Previous studies also showed that curcumin could inhibit the activation of NF-κB in human lung epithelial cells and human myelomonoblastic leukemia cells (Sanjaya & Bharat, 1995; Shishodia et al., 2003). Therefore, we confirmed that curcumin has an anti-inflammatory effect through the partial interference of NF-κB activation. Several studies have indicated that ROS are implicated in the activation of NF-κB (Muller et al., 1997). In the present study, we found that the production of ROS induced by IL-1 β was increased and the pretreatment with curcumin could decreased the. 45.
(53) inducetion of expression by IL-1 β in HUVECs. Previous studies showed that antioxidants such as PDTC and NAC could inhibit the activation of NF-κB, it is strongly suggested that endogenous ROS may play an important role in these redox-sensitive transcription pathways in atherogenesis (Schrect et al., 1992; Weber et al., 1994). Based on the present results, we proposed that the inhibitory effect of curcumin on adhesion molecules expression and NF-κB activation may be due to its antioxidant and anti-inflammatory properties and that it may act by directly scavenging free radicals. In the present study, we found that curcumin was approximately 2-3-folds more potent than Trolox in antioxidative ability and it also could scavenge DPPH, alkoxyl radical (RO‧) and peroxyl radical (ROO‧) (Table 2-1). Since atherosclerosis is a chronic inflammatory disease associated with increased oxidative stress in the vascular endothelium, it is possible that the antiatherogenic effects of curcumin may due to its antioxidative properties.. 46.
(54) 2.5 Conclusion. In the present study, we found that curcumin inhibited monocyte adhesion to endothelial cell, the expression of adhesion molecules, the translocation of NF-κB and the production of ROS in HUVECs. These findings may provide a rationale for the in vitro antiatherosclerosis effect of curcumin and support its potential use in the prevention of atherosclerosis.. 47.
(55) Table 2-1. Antioxidative capacities of curcumin in vitro.. Inhibition of LDL Oxidation (IC50, μmol/l). curcumin. DPPH radicals. TEAC. scavenging ability (IC50, μmol/l). 2.56±0.12. 45.8±0.1. 2.88±0.04. All values are mean ± S.D. IC50 values were obtained from the concentration response curves; n =3.. 48.
(56) Fig. 2-1. 120. % of control. 100 80. *. 60. *. *. 40. 50. 40 20 0 0. 10. 20. 30. curcumin (μmol/l). Figure 2-1. Cytotoxic effect of curcumin on HUVECs with MTT test. HUVECs were treated with verious concentration (0~50 μmol/l) of curcumin for 24 h in 10 % FBS-M199. Values are mean ± S.D., n=4. *p < 0.05 compared with control .. 49.
(57) Fig. 2-2. (A). C. IL-1. IL-1+CU 10. IL-1+CU 20. 50.
(58) Fig. 2-2. (B). a. 300. b. % of control. 250 200. c. 150 d 100 50 0 C. IL-1. IL-1+ CU 10. IL-1+ CU 20. Figure 2-2. Effect of curcumin on IL-1 β-induced adhesion of U-937 cells to HUVECs. (A) Representative images of the reduction of IL-1 β-induced adhesion of U-937 cells to HUVECs monolayers after pretreatment of 10 and 20 μmol/l curcumin for 18 hours. (B) HUVECs were pretreated with 10 and 20 μmol/l curcumin for 18 hours and induced by IL-1 β (10 ng/ml) for 6 hours. Fluorescence-labeled U-937 cell were added to the HUVECs monolayer and allowed to adhere for 30 min. Values are mean ± S.D., n=3. a-d. Means with different letters are significantly different at p < 0.05. Abbreviation: C. (control), IL-1 (IL-1 β), CU10 (curcumin 10 μmol/l), CU20 (curcumin 20 μmol/l).. 51.
(59) Fig. 2-3. (A) ICAM-1. β-actin. (B). 300 a. % of control. 250. b. 200 150. c. d. 100 50 0 C. IL-1. IL-1+ CU 10. IL-1+ CU 20. Figure 2-3. Effect of curcumin on the protein levels of ICAM-1 in cultured HUVECs. HUVECs were pretreated with 10 and 20 μmol/l curcumin for 18 hours and induced by IL-1 β (10 ng/ml) for 6 hours. (A) Representative images of the reduction of IL-1 β-induced the expression of ICAM-1 by curcumin in HUVECs. (B) Densitometric analysis was conducted with image analysis system software to quantify Western blot data. The summarized data (mean ± S.D.) from 3 separate experiments is shown in the bar graph. a-dMeans with different letters are significantly different at p < 0.05. Abbreviation: C (control), IL-1 (IL-1 β), CU10 (curcumin 10 μmol/l), CU20 (curcumin 20 μmol/l).. 52.
(60) Fig. 2-4. (A) VCAM-1. β-actin. a 350. (B). b. % of control. 300 250 200 150. c. c. 100 50 0 C. IL-1. IL-1+ CU 10. IL-1+ CU 20. Figure 2-4. Effect of curcumin on the protein levels of VCAM-1 in cultured HUVECs. HUVECs were pretreated with 10 and 20 μmol/l curcumin for 18 hours and induced by IL-1 β (10 ng/ml) for 6 hours. (A) Representative images of the reduction of IL-1 β-induced the expression of VCAM-1 by curcumin in HUVECs. (B) Densitometric analysis was conducted with image analysis system software to quantify Western blot data. The summarized data (mean ± S.D.) from 3 separate experiments is shown in the bar graph. a-cMeans with different letters are significantly different at p < 0.05. Abbreviation: C (control), IL-1 (IL-1 β), CU10 (curcumin 10 μmol/l), CU20 (curcumin 20 μmol/l).. 53.
(61) Fig. 2-5. (A) E-selectin. β-actin. 300. (B). a. % of control. 250. a. 200 150 b 100. c. 50 0 C. IL-1. IL-1+ CU 10. IL-1+ CU 20. Figure 2-5. Effect of curcumin on the protein levels of E-selectin in cultured HUVECs. HUVECs were pretreated with 10 and 20 μmol/l curcumin for 18 hours and induced by IL-1 β (10 ng/ml) for 6 hours. (A) Representative images of the reduction of IL-1 β-induced the expression of E-selectin by curcumin in HUVECs. (B) Densitometric analysis was conducted with image analysis system software to quantify Western blot data. The summarized data (mean ± S.D.) from 3 separate experiments is shown in the bar graph. a-cMeans with different letters are significantly different at p < 0.05. Abbreviation: C (control), IL-1 (IL-1 β), CU10 (curcumin 10 μmol/l), CU20 (curcumin 20 μmol/l).. 54.
數據


+7

相關文件
The current study found that high-grade MECs had the highest incidence of strong MUC1 expression in mucous cells, with intermedi- ate- and low-grade tumours showing the
Ulrike Schulz, Peter Munzert, Norbert Kaiser, “Surface modification of PMMA by DC glow discharge and microwave plasma treatment for the improvement of coating adhesion” Surface
- we can put photoresist on a wafer and then selectively expose regions to UV - then we can soak the entire thing in acid and only the parts of the photoresist that.. were exposed to
• 由於細胞代謝、紫外線的電離輻射、不良的生 活習慣 ( 煙、酒、肥胖 ) 、各種感染物所引起 的中性粒細胞、巨噬細胞的激活, 會催化分子 氧發生單價還原產生的 "
SPO has high fatty acids, therefor it has to carry out the acid catalyzed esterification to convert of the fatty acids in SPO to biodiesel first, and then used the
the lymphocyte function-associated antigen 1, or LFA-1, was so named because antibodies recognizing this structure interfere with lymphocyte cell adhesion events and
Promote project learning, mathematical modeling, and problem-based learning to strengthen the ability to integrate and apply knowledge and skills, and make. calculated
Wang, Solving pseudomonotone variational inequalities and pseudocon- vex optimization problems using the projection neural network, IEEE Transactions on Neural Networks 17
