Effects of dietary arginine supplementation on nutrient
metabolism and survival rate in burned mice
Sung-Ling Yeh
a, Hui-Ju Tsai
a, Wan-Chun Chiu
a, Huey-Fang Shang
b,*
aInstitute of Nutrition and Health Sciences, Taipei Medical University, Taipei, Taiwan
b
Department of Microbiology and Immunology, Taipei Medical University, Taipei, Taiwan
Received 6 August 2002; received in revised form 7 November 2002; accepted 18 November 2002
Abstract
This study investigated the effect of arginine (Arg) supplementation on nutrient metabolism and splenocyte response in burned mice. Also, the survival of burned mice complicated with Pseudomonas
aeruginosa was evaluated. Experiment 1: Sixty male BALB/c mice were assigned to 2 groups. One
group was fed with casein as the protein source, the other group was supplemented with 2.4% (w/w) Arg in addition to casein. After 4 weeks, all mice received a 30% body surface area burn injury. Mice in each group were sacrificed for 3 consecutive days after the burn with 10 mice on each respective day. Plasma blood chemistry and amino acid profiles were analyzed. Spleens were removed asepti-cally from mice 1 day after the burn, interferon (IFN)-␥ and interleukin (IL)-4 concentrations secreted by cultured splenocytes were measured. Experiment 2: Thirty mice were divided into 2 groups as described in experiment 1. After 4 weeks, burn injury was induced, and mice were challenged with
P. aeruginosa. Survival of the burned mice was observed for 7 days. In experiment 1, no differences
in plasma glucose, non-esterified fatty acids, and lactic acid concentrations were observed between the 2 groups on each respective day. Arg group had higher levels of alanine and branched-chain amino acids on day 2, also, plasma Arg and glutamine levels were higher in the Arg group on days 2 and 3 after the burn than in the control group. IFN-␥ concentrations in mitogen stimulated splenocyte cultures were higher in the Arg group than the control group. No difference in IL-4 concentrations was observed. In experiment 2, the survival did not differ between the 2 groups. These results suggest that Arg supplementation had no beneficial effect on glucose and lipid metabolism, nor had any effect on survival when burned mice were complicated with P. aeruginosa infection. However, Arg supple-mentation reduced the depletion of plasma Arg and glutamine levels at the hypercatabolic stage after
* Corresponding author. Tel.:⫹8862-27361661 ext. 3170.
E-mail address: [email protected] (H-F. Shang).
www.elsevier.com/locate/nutres
0271-5317/03/$ – see front matter © 2003 Elsevier Science Inc. All rights reserved. doi:10.1016/S0271-5317(02)00534-1
the burn, and may have potential benefit on enhancing cellular immune response. © 2003 Elsevier Science Inc. All rights reserved.
Keywords: Burn; Arginine; Amino acid profiles; Survival; Interferon-␥; Interleukin-4
1. Introduction
Severe burn injury induces local and systemic alterations of tissue functions and metab-olism. The metabolic response to burn is characterized by protein and lipid catabolism, total body protein loss, peripheral insulin resistance and increased energy expenditure [1]. Be-sides, burn injury impairs host defense mechanisms against invading microorganisms. The immunosuppression which occurs after burn injury causes increased susceptibility to infec-tion [2]. These responses are multifactorial. Hormones released from neuroendocrine system and agents released by the immune system including pro and anti-inflammatory cytokines, interacts in the regulation of the metabolic change seen after burn [3].
Arginine (Arg) is not an essential amino acid for healthy adults of most mammalian species. However, it is an important amino acid for protein synthesis, biosynthesis of amino acids and generation of urea by the urea cycle. Study by Cui et al. [4] showed that dietary Arg supplementation improved protein anabolism and attenuated muscle protein catabolism after thermal injury. Studies also showed that Arg has beneficial effects in burned individuals including increase release of growth hormone, stimulation of wound healing, and improved immune functions [5– 8]. Previous work in our laboratory demonstrated that dietary Arg supplementation attenuates the oxidative stress induced by burn injury, and a better in vitro macrophage response was observed when Arg was administered [9]. Arg supplementation also has been reported to improve survival in a noninfected burned animal [10]. However, the roles of Arg on nutrient metabolism in burn injury have not been fully determined. Therefore, the first aim of this study was to evaluate the effects of dietary Arg supplemen-tation on glucose and lipid metabolism as well as amino acid profiles in burned mice.
Pseudomonas aeruginosa is an opportunistic pathogen that often infects burned patients [11].
Therapy for P. aeruginosa infection is hindered by its well-known antibiotic resistance. Recently, our report demonstrated that dietary Arg supplementation had no appreciable benefit in the production of specific antibodies against P. aeruginosa in burned rats [12]. In order to understand the possible role of Arg on immune response after the burn, survival in burned mice complicated with P. aeruginosa infection, and in vitro cytokine production by splenic cultures were analyzed.
2. Materials and methods
2.1. Animals
Male BALB/c mice weighing 10 –15 g (4 weeks of age) were used in this study. All mice were housed in temperature and humidity controlled rooms, and allowed free access to a
standard chow for 1 week prior to the experiment. The animals included in this study were kept under standard experimental animal care protocols.
2.2. Study protocol
Experiment 1: Sixty mice were randomly assigned to 2 groups, 30 mice in each group. One group was fed with a control diet (Control), in which all amino acids were provided in casein. The other group was fed with Arg (Arg), which 2% total kcal of Arg was supple-mented in addition to casein, the Arg content in the diet was 2.4% (w/w). The 2 diets were isonitrogenous (Table 1). After 4 weeks, all dorsal hair was clipped, and each animal was anesthetized with ether. An asbestos board with a 2.5⫻ 3 cm2 window was pressed firmly against the shaved back. Ethanol (0.5 ml) was evenly spread over the area of the back outlined by the window, ignited, and allowed to burn for 15 sec [13]. Animals were immediately resuscitated by an intraperitoneal injection of sterile 0.9% saline (10 mL/100g body weight) [14]. This procedure produced a full-thickness burn injury on approximately 30% of the total body surface area. They were then food deprived for 24 h with free access to water. Mice on each group were sacrificed on the 3 consecutive days after the burn with 10 mice on each respective day. Mice were anesthetized and killed by exsanguination using heart puncture. The blood samples were collected in tubes containing heparin and immedi-ately centrifuged. All samples were stored at⫺70°C until assay. Immediately after sampling the blood, spleens were removed aseptically from mice 1 day after the burn, and were placed in phosphate buffer saline (PBS) for the pretreatment of in vitro splenocyte cultures.
Table 1
Composition of the experimental diet (g/kg)
Component Arg Control
Casein 200 248 Arg 24 — Protein N1 39.7 39.7 Soybean oil 50 50 Corn starch 470 446 Salt mixture2 35 35 Vitamin mixture3 10 10 Methylcellulose 30 30 Choline chloride 1 1 DL-Methionine 3 3 Sucrose 200 200 1
Total nitrogen calculated from casein and arginine. 2
Salt mixture contains the following (mg/g): calcium phosphate diabasic 500 mg, sodium chloride 74 mg, potassium sulfate 52 mg, potassium citrate monohydrate 220 mg, magnesium oxide 24 mg, manganese carbonate 3.5 mg, ferric citrate 6 mg, zinc carbonate 1.6 mg, curpric carbonate 0.3 mg, potassium iodate 0.01 mg, sodium selenite 0.01 mg, and chromium potassium sulfate 0.55 mg.
3
Vitamin mixture contains the following (mg/g): thiamin hydrochloride 0.6 mg, riboflavin 0.6 mg, pyridoxine hydrochloride 0.7 mg, nicotinic acid 3 mg, calcium pantothenate 1.6 mg, D-biotin 0.02 mg, cyanocobalamin 0.001 mg, retinyl palmitate 1.6 mg, DL-␣-tocopherol acetate 20 mg, cholecalciferol 0.25 mg, menaquinone 0.005 mg.
Experiment 2: Thirty mice were divided into 2 experimental groups, with each group containing 15 mice. Mice in the experimental groups were fed with control or Arg diets for 4 weeks as described in Exp.1. After that, burn injury was induced and 3.2 ⫻ 105 CFU (LD50) P. aeruginosa strain PAO1 (ATCC 15692) was injected into the burned skin of the mice. Survival was noted every 6 h in the first 3 days, and then every 12 h until 7 days.
2.3. Measurements for blood chemistry
Plasma glucose, lactic acid, and nonesterified fatty acid (NEFA) were determined by colorimetric methods after enzymatic reaction (Randox Co., Antrim, Ireland). Amino acids were analyzed by ion exchange chromatography using ninhydrin reaction for detection (Beckman Instruments, model 6300, Palo Alto, CA) [15].
2.4. In vitro cultures of splenocytes
Splenocytes were obtained by mechanical disruption using a homogenizer. Cell suspen-sions then were passed through sterile nylon mesh to remove debris. RBC were lysed by sterile distilled water for 15 sec, and immediately neutralized to isotonic cell suspensions. After washing with PBS for 3 times (300⫻ g for 5 min), splenocytes were resuspended in RPMI-1640 with supplements. The number of isolated splenocytes was determined by hemacytometer count using trypan blue dye exclusion. Phytohemagglutinin (PHA, 10 ng/ mL; Sigma Co., USA) and lipopolysacchride (LPS, 20 ng, 50 ng/mL; Sigma Co., USA) were used to stimulate cytokine production by isolated splenocytes in culture. Triplicate wells of 96-well flat-bottomed microtiter plates (Falcon, Becton Dickinson, Co., Oxford, CA.) were seeded with 100l of splenocytes (2.5 ⫻ 106cells/ml in RPMI-1640) and 100l mitogen. Control wells contained cells plus 100l of medium. The final volume in all wells was 200
l. Splenocytes were incubated in the presence or absence of PHA or LPS for 24 hr at 37o
C in an atmosphere of 95% air and 5% CO2. Supernatants were centrifuged and stored at
⫺70°C until for the analysis of cytokines.
2.5. Cytokine assay
Interferon (IFN)-␥, interleukin (IL)-1, IL-4, tumor necrosis factor (TNF)-␣ concentrations in splenocyte supernatant were determined with commercially available enzyme-linked immunosorbent assay (ELISA) in microtiter plates. Antibodies specific for mice cytokines were coated onto the wells of the microtiter strips provided (Amersham Pharmacia Biotech Inc., UK). Procedures were described in the manufacturer’s instruction.
2.6. Statistics
Data are expressed as mean⫾ SD. Differences among groups were analyzed by analysis of variance using Duncan’s test. Survival rate was measured by Kaplan-Meier survival analysis. A p value⬍ 0.05 was considered statistically significant.
3. Results
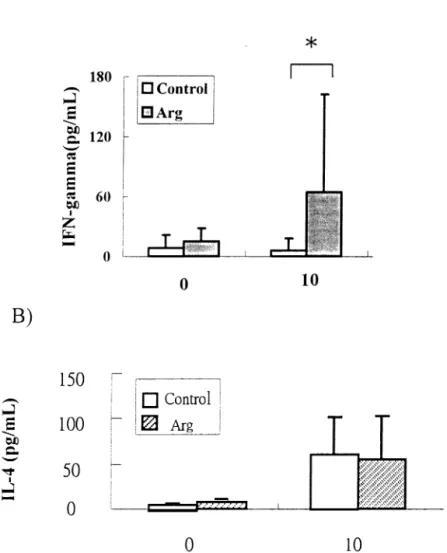
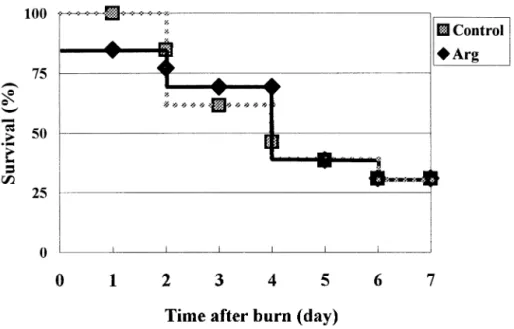
There were no differences in the initial and final body weights between the 2 groups (data not shown). Plasma glucose and lactic acid concentrations were significantly lower on day 1 than days 2 and 3, lactic acid levels were even higher on day 3 than day 2 after the burn, whether mice were fed Arg or not. No significant differences in plasma glucose, lactic acid and NEFA concentrations were observed between the 2 groups on each respective day after the burn (Table 2). Plasma amino acid profiles showed that Arg and glutamine concentrations in the Arg group were significantly higher than the control group on day 2 and 3 after the burn (Fig. 1A). Plasma concentrations of alanine and branch chain amino acids (BCAA), including leucine, isoleucine and valine were significantly higher in the Arg group than the control group on day2 (Fig. 1A & B). IFN-␥ concentrations in PHA-stimulated splenocyte cultures were significantly higher in the Arg group than the control group (Fig. 2A). However, no significant difference in IL-4 concentrations in PHA-stimulated splenocyte cultures were observed (Fig. 2B). IL-1 and TNF-␣ secreted by LPS-stimulated spenocytes did not show any difference between the 2 groups (data not shown). Results in Exp. 2 showed that there was no significant difference in survival between the 2 groups (Fig. 3).
4. Discussion
Previous studies have shown that feeding the burned rat immediately ad libitum induces rapid recovery, in contrast to the hypercatabolic state observed in burn patients [16]. Mochizuki et al. [17] reported that early enteral feeding of burned guinea pigs prevents the characteristic hypercatabolic reaction. Since the rodent is relatively resistant to trauma, we administered the experimental diets before burn injury to investigate whether Arg supple-mentation had effects on nutrient metabolism and immune response in metabolic stress. In this study, 24 h starvation after the burn was used to induce the hypermetabolic state of the burned mice. These experimental conditions simulated metabolic disorders observed in burn patient [18]. We sacrificed the mice on the 3 consecutive days after burn, because 3 days after burn was thought to be the time of maximum hypercatabolism in the rodent model [18]. We did not include a non-burn control group in this study, because numerous studies have shown
Table 2
Plasma glucose, non-esterified fatty acids (NEFA), and lactate in mice after burn for 1, 2 and 3 days between the 2 groups
Arg Control
Day 1 Day 2 Day 3 Day 1 Day 2 Day 3
Gluocse (mg/dL) 129.6⫾ 21.3# 200.0⫾ 21.5 184.4 ⫾ 44.9 131.4 ⫾ 19.4# 217.8⫾ 21.7 162.9 ⫾ 47.0 NEFA (mmol/L) 1.30⫾ 0.18# 1.68⫾ 0.17 1.77⫾ 0.17 1.17⫾ 0.16 1.29⫾ 0.17 1.31⫾ 0.59 Lactate (mg/dL) 52.8⫾ 9.6# 76.9⫾ 9.1* 89.9⫾ 14.3 55.7⫾ 7.6# 74.0⫾ 5.5* 97.6⫾ 11.8 #
Significantly different from day 2 and day 3 of the same group. * Significantly different from day 1 and day 3 of the same group.
Fig. 1. Plasma A) arginine, glutamine and alanine B) valine, leucine and isoleucine concentrations between the 2 groups after the burn for 1, 2 and 3 days. n⫽ 10 on each respective day. *Significantly different from the control group at the same time point.
that burn injury results in abnormal nutrient metabolism and impaired immune function [1–3,19].
Results of this study showed that plasma glucose and lactic acid concentrations were significantly lower on day 1 than on day 2 and 3 after the burn, regardless of mice being fed Arg or not. The difference might have resulted from starvation of the mice for the first day or from the ebb phase observed after the burn [19]. Studies in humans and animal models have shown that gluconeogenesis is markedly elevated after a burn. Glucose is metabolized in burn wounds primarily by glycolysis despite the presence of oxygen, with the subsequent release of lactic acid locally and into the systemic circulation [19] as observed in this study, which shows that plasma concentrations of lactic acid are elevated in accordance with the progress of the burn injury. In this study, no differences in plasma glucose, lactic acid and
Fig. 2. A) Interferon (IFN)-␥ concentrations and B) Interleukin (IL)-4 concentrations in PHA-stimulated (0, 10 ng/mL) splenocyte cultures from burned mice. (n⫽ 10) *Significant difference between the 2 groups at the given concentration of PHA.
NEFA concentrations were observed between the 2 groups at various time points after the burn. This result may indicate that Arg supplementation did not influence glucose and lipid metabolism after the burn.
Previous studies showed that burn injury exerts a great stimulus for muscle protein catabolism. BCAAs are oxidized primarily in skeletal muscle [20]. In the catabolic state, BCAAs are produced as a result of muscle protein breakdown, and the amino groups of BCAAs transfer to other amino acids for the de novo synthesis of alanine and glutamine (Gln) [21]. Many in vivo studies indicated that the peripheral uptake or clearance of BCAAs is significantly increased after trauma, suggesting an increased requirement for oxidation in muscle tissue [22,23]. Wolfe et al. [24] demonstrated that burn patients had an increased rate of leucine oxidation and an increased rate of leucine utilization for protein synthesis. In addition, degradation of Arg and Gln increased and plasma levels of Arg and Gln were significantly reduced after severe burn injury [25–27]. The present results show that the Arg group had higher plasma levels of alanine and BCAAs on day 2; Arg and Gln levels were higher on days 2 and 3 after the burn than were those levels in the control group. This finding may indicate that Arg supplementation reduces the depletion of plasma Arg and Gln in burn injury. Yoshida et al. [28] demonstrated that under catabolic conditions, the proportion of Gln derived from leucine is increased. It is possible that plasma BCAAs and alanine were used on day 3 in the Arg group to maintain Gln levels after the burn. Although the removal of BCAAs from circulation was accelerated in burned mice fed the control diet, the lower levels of plasma Arg and Gln resulting from a burn-induced catabolic reaction, however, can not be compensated. A previous study carried out by our laboratory showed that antioxidant enzyme activities and lipid peroxides in tissues tended to be lower in the Arg group than in the control group after a burn [9] Since burn injury is a trauma with high oxidative stress
Fig. 3. Survival curves of the arginine and control groups after challenge with Pseudomonas aeruginosa at burn area (n⫽ 15 for each group). There was no significant difference in the survival rate between the 2 groups.
[19], it is possible that the beneficial effects of Arg supplementation on reducing burn-induced catabolic reaction is due to its attenuation of oxidative stress. The effect of Arg on the immune response may also play a role.
Saito et al. [10] demonstrated that Arg supplementation improved survival in a non-infected animal. In this study, we non-infected the burned mice with P. aeruginosa to increase their mortality, since it is usually a major problem in most thermal injury caring centers. However, we observed no difference in survival between the 2 groups, this result was consistent with our previous report in a burned infected rat model. In the previous report we found that the production of specific antibody against P. aeruginosa was not enhanced after Arg supplementation [12]. In order to further understand the effect of Arg on immune response, we analyzed the cytokines production of splenocytes after mitogen stimulation. IFN-␥ is primarily produced by Th1 lymphocytes, it enhances cellular immunity and promotes differentiation of the Th1 cells [29]. IL-4 is produced by Th2 lymphocytes. IL-4 activates B cells and promotes humoral immunity [29]. In this study, we found that splenocytes from mice on an Arg-supplemented diet secreted significantly greater amount of IFN-␥ than that of the control group when the same concentration of PHA was used. However, no significant difference in IL-4 concentrations in PHA-stimulated cultures was observed. These results indicate that Arg supplementation may have potential benefit on enhancing cellular immune response instead of humoral immunity. Because the defense mechanism against P. aeruginosa is dependent upon the neutralization and opsonization of antibodies [30], the production of specific antibodies against P. aeruginosa will be a major factor to determine the survival of burned infected mice. Since Arg had no beneficial effects on IL-4 secretion and specific antibody production in burned rodent models, this may explain why survival in burned mice complicated with infection were not improved when Arg was administered.
In conclusion, this study demonstrates that, compared with the control diet, Arg supple-mentation had no beneficial effects on glucose and lipid metabolism, nor did it have an effect on survival when burned mice were complicated with P. aeruginosa infection. However, Arg supplementation reduces the depletion of plasma Arg and Gln levels at the hypercatabolic stage after the burn and in vitro IFN-␥ production is significantly elevated in the Arg group compared to the control group when splenocytes are mitogen-stimulated. Whether Arg supplementation plays a role in cellular immune response in rodents subjected to burn trauma requires further investigation.
Acknowledgments
This study was supported by research grant NSC 90-2320-B-038-037 from the National Science Council, R.O.C.
References
[2] O’Sullivan ST, O’Connor TPF. Immunosuppression following thermal injury: the pathogenesis of immu-nodysfunction. Br J Plastic Surg 1997;50:615–23.
[3] Tredget EE, Yu YM. The metabolic effects of thermal injury. World J Surg 1992;16:68 –79.
[4] Cui XL, Iwasa M, Iwasa Y, Ohmori Y, Yamamoto A, Maeda H, Kume M, Ogoshi S, Yokoyama A, Sugawara T, Funada T. Effects of dietary arginine supplementation on protein turnover and tissue protein synthesis in scald-burn rats. Nutrition 1999;15:563–9.
[5] Barbul A, Rettura G, Levenson SM, Seifter E. Wound healing and thymotropic effects of arginine: a pituitary mechanism of action. Am J Clin Nutr 1983;37:786 –94.
[6] Barbul A, Lazarou SA, Efron DT, Wasserkrug HL, Efron G. Arginine enhances wound healing and lymphocyte immune response in humans. Surgery 1990;108:331–7.
[7] Daly JM, Reynolds J, Thom A, Kinsley L, Dietrick-Gallagher M, Shou J, Ruggieri B. Immune and metabolic effects of arginine in surgical patients. Ann Surg 1988;208:512–23.
[8] Evoy D, Lieberman MD, Fahey TJ III, Daly JM. Immunonutrition: the role of arginine. Nutrition 1998; 14:611–7.
[9] Tsai HJ, Shang HF, Yeh CL, Yeh SL. Effects of arginine supplementation on antioxidant enzyme activity and macrophage response in burned mice. Burns 2002;28:258 – 63.
[10] Saito H, Trocki O, Wang SL, Gonce SJ, Joffe SN, Alexander JW. Metabolic and immune effects of dietary arginine supplementation after burn. Arch Surg 1987;122:784 –9.
[11] Bodey GP, Bolivar R, Fainstein V, Jadeja L. Infections caused by Pseudomonas aeruginosa. Rev Infect Dis 1983;5:279.
[12] Shang HF, Wang YY, Wu YC, Tsai AY, Chiu WC, Yeh SL. Effects of arginine supplementation on humoral immunity in burned rats. New Taipei J Med 2002 (in press).
[13] Holder IA. The pathogenesis of infection owing to Pseudomonas aeruginosa using the burned mouse model: experimental studies from the Shriners Burns Institute, Cincinnati. Can J Microbiol 1985;31:393– 402. [14] Holder IA, Jogan M. Enhanced survival in burned mice treated with antiserum prepared against normal and
burned skin. J Trauma 1971;11:1041– 6.
[15] Smith RJ, Panico K. Automated analysis of o-phthalaldehyde derivatives of amino acids in physiological fluids by reverse-phase high performance liquid chromatography. J Liq Chromatogr 1985;8:1783–5. [16] Jahoor F, Desai M, Herndon DN, Wolfe RR. Dynamics of the protein metabolic response to burn injury.
Metabolism 1988;37:330 –7.
[17] Mochizuki H, Trocki O, Dominioni L. Metabolism of prevention of post-burn hypermetabolism and catabolism by early enteral feeding. Ann Surg 1984;200:297–310.
[18] Vaubourdolle M, Coudray-Lucas C, Jardel A, Ziegler F, Ekindjian O, Cynober L. Action of enterally administered ornithine␣-ketoglutarate on protein breakdown in skeletal muscle and liver of the burned rat. J Parenter Enter Nutr 1992;15:517–20.
[19] Tredget EE, Yu YM. The metabolic effects of thermal injury. World J Surg 1992;16:68 –79.
[20] Odessey R, Goldberg AL. Oxidation of leucine by rat skeletal muscle. Am J Physiol 1972;223:1376 – 83.
[21] Odessey R, Khairallah EA, Goldberg AL. Origin and possible significance of alanine production by skeletal muscle. J Biol Chem 1974;249:7623–9.
[22] Elia M, Farrell R, Ilie V, Smith R, Williamson DH. The removal of infused leucine after injury, starvation and other conditions in man. Clin Sci 1980;59:275– 83.
[23] Desai SP, Bistrain BR, Moldawere LL, Miller MM, Blackburn GL. Plasma amino acid concentrations during branch-chain amino acid infusion in stressed patients. J Trauma 1982;22:747–52.
[24] Wolfe RR, Goodenough RD, Burke JF, Wolfe MH. Response of protein and urea kinetics in burn patients to different levels of protein intake. Ann Surg 1983;197:163–71.
[25] Yu Y, Ryan CM, Burke JF, Tompkins RG, Young VR. Relations among arginine, citrulline, ornithine, and leucine kinetics in adult burn patients. Am J Clin Nutr 1995;62:960 – 8.
[26] Parry-Billings M, Evans J, Calder PC, Newsholme EA. Does glutamine contribute to immunosuppression after major burn? Lancet 1990;338:523–5.
[27] Stinnett JD, Alexander JW, Watanabe C, MacMillan BG, Fischer JE, Morris MJ, Trocki O, Miskell P, Edwards L, James H. Plasma and skeletal muscle amino acids following severe burn in patients and experimental animals. Ann Surg 1982;195:75– 89.
[28] Yoshida S, Lanza-Jacoby S, Stein P. Leucine and glutamine metabolism in septic rats. Biochem J 1991;276:405–9.
[29] Dipiro JT. Cytokine networks with infection: mycobacterial infections, leishmaniasis, human immunode-ficiency virus infection, and sepsis. Pharmacotherapy 1997;17:205–23.
[30] Chen TY, Shang HF, Chen TL, Lin CP, Hwang J. Recombinant protein composed of pseudomonas exotoxin A, outer membrane protein I and F as vaccine against P. aeruginosa infection. Appl Microbiol Biotechnol 1999;52:524 –33.