Corresponding author: Dr. Shih-Chieh Chen, Department of Anatomy, Kaohsiung Medical University, 100 Shih-Chuan 1st Road, Kaohsiung, Taiwan, R.O.C. Tel: 886-7-3121101 ext. 2301, Fax: 886-7-3119849, E-mail: [email protected]
Received: February 1, 2005; Revised: March 8, 2005, 1999; Accepted: March 18, 2005.
Effects of Hyperthermia Pretreatment on
Expression of Heme Oxygenase-1 and Nitric
Oxide Synthase in Rats Subjected to
Experimental Anaphylactic Shock
Rei-Cheng Yang*, Chao-Yuah Chang, Tzong-Shi Lu* and Shih-Chieh Chen#Department of Anatomy and Physiology Kaohsiung Medical University
100 Shih-Chuan 1st Road Kaohsiung, Taiwan
Abstract
Previous studies have shown that hyperthermia pretreatment results in an attenuation of increased vascular leakage in rats subjected to experimental anaphylactic shock. It is known that both nitric oxide synthase (NOS) and heme oxygenase-1 (HO-1) play a role in the maintenance of microvascular integrity. In the study, we investigated the effect of hyperthermia pretreatment on mRNA expression of endothelial NOS (eNOS), inducible NOS (iNOS), and HO-1 in heated or nonheated rats subjected to anaphylactic shock using a semi-quantitative RT-PCR. Protein contents of eNOS and HO-1 in tissue were also assayed. Plasma nitrite and nitrate before and after induction of anaphylactic shock were quantified using a NO analyzer. The heated, anaphylactic rats showed a significant increase of HO-1 mRNA expression in heart as compared to both non-heated, anaphylactic and control rats. HO-1 protein contents in both heart and lung tissues in the heated, anaphylactic rats were significantly higher than both non-heated, anaphylactic and control rats. Protein contents of eNOS in various tissues appeared to be the same among groups. No significant change of iNOS mRNA expression was detected among groups. Plasma nitrite and nitrate before and after anaphylactic treatment appeared to be the same among groups. These data suggest that reduction of anaphylactic hypotension by hyperthermia pretreatment in rats subjected to anaphylactic shock may be resulted from over-expression of HO-1 rather than NOS in various tissues.
Key Words: anaphylactic shock, heme-oxygenase-1, hyperthermia, nitric oxide synthase
Introduction
The increase of vascular permeability induced by vasoactive inflammatory mediators are attributed to the pathophysiologic conditions of anaphylactic shock. The heat shock response by a brief period of hyperthermia provides a protective effect for experi-mental animals against subsequent various forms of stress (1, 5, 9, 26). Our previous studies (4) have shown that hyperthermic pretreatment results in a significant attenuation of anaphylactic hypotension and increased plasma protein leakage in heart and
lung tissues in rats subjected to anaphylactic shock. Heat shock proteins (HSPs) belong to multigene families and are universally induced in all organisms by hyperthermia and various chemical or physical stressors (2, 13, 18, 20). The inducible heme-oxyge-nase-1 (HO-1) belongs to heat shock protein family (2, 18, 20). HO catalyzes oxidative degradation of heme to carbon monoxide (CO), bilirubin, and iron. CO has been implicated in the modulation of blood vessel function (16, 23). It was previously shown that HO-1 has salutary effects in pulmonary, hepatic and cardiovascular diseases (15). Nitric oxide (NO)
pro-duced by NO synthase (NOS) play a role in the maintenance of microvascular integrity under physi-ological and pathophysiphysi-ological conditions (10-12). There are 3 isoforms of NOS: endothelial NOS (eNOS), neuronal NOS (nNOS) and inducible NOS (iNOS). In the present studies, we investigated the effect of hyperthermia pretreatment on mRNA ex-pression of eNOS, iNOS and HO-1 in various tissues in rats subjected to anaphylactic shock, using a semi-quantitative RT-PCR. Protein contents of both HO-1 and eNOS were further assayed in tissue. Plasma nitrite and nitrate contents in femoral veins were also measured before and after anaphylactic shock as an indication of NO production in tissue.
Materials and Methods Experimental Animals
Male Wistar rats were purchased from the National Animal Center (Nan-kang, Taipei, Taiwan) and housed in the animal center in Kaohsiung Medical University. Experiments described in these studies were approved by the Animal Committee of the Kaohsiung Medical University.
Antigen Sensitization, Induction of Anaphylactic Shock and Plasma NO Measurement
Rats were randomized into 3 groups (anaphylactic shock, heat and anaphylactic shock, and control with-out any treatment) of 6 rats each. The procedure of induction of anaphylactic shock was performed as in the previous studies (4). In brief, rats (300g-400g) were sensitized by subcutaneous injection of bovine serum albumin (BSA) in complete Freund’s adjuvant. After 14 days, anesthetized sensitized rats in both heat and non-heat group were boosted with BSA via the femoral vein and blood pressure (BP) was re-corded from the femoral artery with a BP transducer connected to a polygraph recorder. Non-heated, non-sensitized control rats received saline injection only. The changes of BP appeared to be the same as in the previous studies (4) and plasma nitrite and nitrate contents were assayed in femoral venous blood. In the same rats received anaphylactic treatments as described above, the other femoral vein was also exposed. Before BSA injection and 10 min after, blood samples (1 ml) were collected by inserting a 24 gauge needle attached with a syringe and equal vol-ume of saline was replenished. Plasma nitrite and nitrate contents were measured by a NO analyzer (ENO-20, Eicom Cor.; Tokyo, Japan) using an auto-mated procedure based on the Griess reaction. The NO analyzer was also interfaced with a commercial personal computer software (Powerchrom, AD
Instruments; Castle Hill, Australia) to quantify nitrite a n d n i t r a t e c o n t e n t s b a s e d o n a c a l i b r a t i o n chromatogram. Plasma levels of NO were presented by the sum of both nitrite and nitrate.
Heat Shock or Hyperthermia Pretreatment
Twenty-four hours before BSA challenge to induce anaphylactic shock, rats of the heated group were treated with whole-body hyperthermia (4) by placing rats on the automatic heat-pads; the rat rectal temperature was maintained at 41.5°C (±0.5°C) for 15 minutes. Non-heated or control rats were anesthetized, but not treated with hyperthermia and placed back in cage.
Isolation of RNA in Tissue and Reverse Transcription After monitoring BP changes, the vascular system of rats was exsanguinated with 0.9% NaCl solution injected through the left ventricle. Heart, lung and liver were removed and stored in RNAlater (Ambion Inc.; Austin, TX, USA). Then the tissues were frozen in liquid nitrogen and pulverized with a pestle in a mortar. Total RNA was isolated using a SV Total RNA Isolation System (Promega Co.; Madison, WI, USA). The integrity of isolated RNA was confirmed by agarose gel electrophoresis and RNA concentration was determined by spectrophotometry. Samples of 1 µg total RNA were reverse transcribed into cDNA as in the previous studies (28). The resulting cDNA was frozen at -20°C .
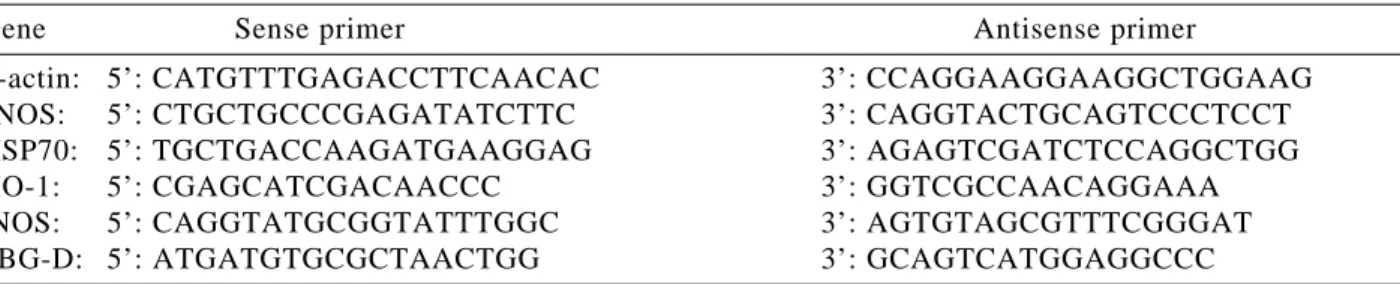
Detecting of eNOS, iNOS and HO-1 mRNA by Duplex PCR Target genes were amplified using a Taq DNA polymerase kit (FastStart Taq DNA polymerase, Roche Diagnostics GmbH; Mannheim, Germany) as in the previous studies (28). In brief, a volume of 1.5 µl of cDNA from RT was replicated in PCR reactions in a volume of 25 µl including Taq DNA polymerase, buffer supplied with enzymes, MgCl2, dNTP and primers (in Table 1). Duplex PCR was performed using primers of either porphobilinogen deaminase (PBGD) or β-actin and one of target genes: iNOS, HO-1, eNOS. PCR condition was 94°C for 4 min followed by 25 ~ 28 cycles of 94°C for 30 sec, 60°C for 30 sec, 72°C for 45 sec and finally 72°C for 7 min. Primer sequences (β-actin and eNOS) were adopted from the previous studies (3, 25), whereas those of iNOS (accession # D14051), PBG-D (accession # X06827) and HO-1 (accession # J02722) were designed via a commercial personal computer software (Lightcycler probe design; Roche Diagnos-tics GmbH; Mannheim, Germany). All primers were made by a commercial company (GIBCO BRL;
Rockville, ML, USA). The expected size for the DNA fragments of eNOS, β-actin, iNOS, PBGD and HO-1 was 230, 440, 241, 186 and 335 bp respectively.
After electrophoresis, the PCR products were quantified by a densitometer interfaced with Bio-profil image analysis software (Vilber Lourmat, Marne La Vallee Cedex 1, France) as in the previous studies (28). And the results were expressed as ratios relative to PBGD or β-actin. Primer sequences of HSP 70 was adopted from the previous studies (6). PCR condition was similar to the procedure mentioned above except that 33 cycles of 94°C for 30 sec, 59.2°C for 30 sec, 72°C for 45 sec were used. The expected size for the DNA fragment of HSP 70 gene was 492 bp.
HO-1 Protein Detection by Western Blot and Immunostaining
In another series of anaphylactic experiments performed as above, rats were randomized into 4 groups (anaphylactic shock, heat and anaphylactic shock, heat only and control without any treatment) of 5 rats each. After exsanguination with 0.9% NaCl solution, heart, lungs and liver were excised. Protein isolation, Western blot and immunostaining were performed as in the previous studies (4). In brief, the proteins on the gel after electrophoresis were transblotted to a nitrocellulose membrane. The membranes were incubated with monoclonal anti-HO1 antibody (Stressgen Biotech., British Columbia, Canada) and followed by incubation with goat anti-mouse IgG conjugated with peroxidase (Amersham Pharmacia, Uppsala, Sweden). Target protein bands were visualized by enhanced chemilluminedcence (ECL kit. NEN life science products, Boston, MA, USA). Beta-tubulin was also extracted and tested by Western immunoblotting using anti-tubulin antibody (Chemicon International, Temecula, USA) and used for comparison among different groups.
Endothelial NOS (eNOS) Protein Assay
The protein contents in tissue extracts from above was determined by a Bradford assay (Bio-Rad Lab.;
Hercules, CA, USA) and used for comparison among different groups. The protein contents of eNOS in tissue extracts from above were analyzed using the ELISA technique. The procedure of performing ELISA was designed according to the manufacturer’s proto-col (Quantikine eNOS Immunoassay, R&D Systems, Inc.; Minneapolis, MN, USA).
Statistical Analysis
Values are presented as means±SD. The non-parametric Kruskal-Wallis test was used to compare the means of 3 or 4 groups of data. If the Kruskal-Wallis test indicated an overall significant difference (P< 0.05), the pairwise multiple comparison test (Duncan test) was used to determine the significance of difference in the mean between any two groups. The Student t test was used to compare the means of two groups of data.
Results Plasma Nitrite and Nitrate
Systemic BP of heated rats subjected to anaphy-lactic treatment promptly returned to baseline levels and hypotension was sustained for the duration of the experiment in non-heated rats as in the previous studies (4). Plasma nitrite and nitrate before and 10 min after BSA challenge of anaphylactic treatment were 53.9±10.6; 56.3±11.2 in non-heated rats (n=6) and 62.5±21.2; 53.4±12.9 in heated rats (n=6) respectively. Plasma nitrite and nitrate before and 10 min after saline injection were 57.6±15.3; 59.8±13.7 in control rats (n=6). The results showed that plasma nitrite and nitrate in blood collected after BSA chal-lenge in either heated or non-heated rats appeared to be the same as basal levels.
Gene Expression of eNOS, iNOS, HO-1 and HSP70 in Various Tissues by RT-PCR
Fig. 1 depicts a procedure for semi-quantitative assessment of gene expression for HO-1 and PBG-D
Table 1. Oligonucleotide primers used for PCR.
Gene Sense primer Antisense primer
β-actin: 5’: CATGTTTGAGACCTTCAACAC 3’: CCAGGAAGGAAGGCTGGAAG eNOS: 5’: CTGCTGCCCGAGATATCTTC 3’: CAGGTACTGCAGTCCCTCCT HSP70: 5’: TGCTGACCAAGATGAAGGAG 3’: AGAGTCGATCTCCAGGCTGG
HO-1: 5’: CGAGCATCGACAACCC 3’: GGTCGCCAACAGGAAA
iNOS: 5’: CAGGTATGCGGTATTTGGC 3’: AGTGTAGCGTTTCGGGAT
using the techniques of RT combined with duplex PCR. Initially duplex PCR experiments for both HO-1 and PBG-D were performed with various amplifica-tion cycles using cDNA isolated from heart tissue (Fig. 1A). Upon analysis of the density of PCR products for both HO-1 and PBG-D amplified for 20-35 cycles, an amplification cycle was chosen to en-sure the cycles were not subjected to plateau effects (Fig. 1B). In the present study, the intensities ob-tained at an amplification cycle of 28 were used for group comparison. The heated rats subjected to
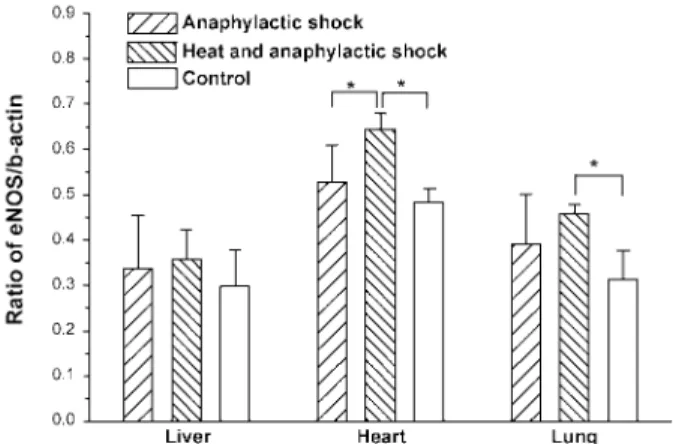
ana-phylactic treatment showed a significant increase in the levels of HO-1 mRNA expression in heart tissue as compared to both non-heated, anaphylactic and control rats (Fig. 1C). In contrast, no obvious change in the levels of HO-1 mRNA expression in both liver and lung tissues were detected among 3 groups of rats. Expression of eNOS mRNA in the examined tissue in non-heated rats subjected to anaphylactic treatment appeared to be the same as compared with control rats (Fig. 2). However, the heated rats sub-jected to anaphylactic treatment showed a significant increase in the levels of eNOS mRNA expression in heart and lung tissues as compared to control rats. In liver, no obvious change in the levels of eNOS mRNA expression was noted among 3 groups of rats. No obvious change of iNOS mRNA expression was de-tected in the examined tissue among 3 groups of rats (Fig. 3). Inducible HSP70 appeared overexpressed in heart tissue of heated rats as compared to both non-heated and control rats (data not shown).
Heme Oxygenase-1 (HO-1) and eNOS Protein Expression HO-1 protein contents in heart and lung tissues in the heated rats subjected to anaphylactic treatment were significantly higher than both non-heated, anaphylactic rats and control rats (Fig. 4). Likewise, significant increase of HO-1 protein levels were also detected in examined tissue in rats pretreated with hyperthermia only. In contrast, HO-1 protein in e x a m i n e d t i s s u e i s o l a t e d f r o m 5 d a y s a f t e r hyperthermia pretreatment obviously decreased to basal control levels (Fig. 5). In liver there was no significant difference for HO-1 protein levels among groups. Basal eNOS protein level was detected in all examined tissues (Table 2). No obvious change of Fig. 1. Semi-quantitative assessment of gene expression for
heme oxygenase-1 (HO-1) and porphobilinogen deami-nase (PBGD) using the techniques of RT combined with duplex PCR. A. An initial duplex PCR experiment was performed to determine the cycles required to measure gene expression for both HO-1 and PBG-D using cDNA isolated from heart. Lane M: 1 kb DNA ladder; lane 1: 20 cycles; lane 2: 25 cycles; lane 3: 30 cycles; lane 4: 35 cycles. B. Relative density of PCR products of both HO-1 and PBG-D obtained upon amplification for 20-35 cycles were plotted. C. Expression of HO-1 mRNA relative to PBG-D in various tissues in rats received different treatment (n = 6) and control rats (no treatment; n = 6). *: P < 0.05.
Fig. 2. Expression of endothelial nitric oxide synthase (eNOS) mRNA relative to β-actin mRNA among different groups of rats using the similar techniques described in Fig. 1. Rats (n = 6)/group; *: P < 0.05.
eNOS protein expression was noted in the examined tissue among groups.
Discussion
We have used a semi-quantitative assessment of duplex PCR in order to investigate expression levels of mRNA of specific genes among different groups of rats: anaphylactic and non-heated, anaphylactic and heated, and control. The specific genes were syn-thetic enzymes of both NO and CO. Protein contents of both HO-1 and eNOS were further assayed in tissue. The present studies showed that overproduc-tion of iNOS was not detected in rats subjected to anaphylactic treatment. Heat shock response was indicated by overexpression of HSP70 mRNA and HO-1 protein in the present studies. Hyperthermic pretreatment apparently increased HO-1 protein lev-els in both heart and lung in the heated rats subjected to anaphylactic treatment as compared to both non-heated, anaphylactic and control rats. In contrast, HO-1 protein obviously decreased in rats 5 days after hyperthermic pretreatment. These results indicate that attenuation of microvascular leakage and ana-phylactic hypotension is highly related with presence of heat shock proteins induced by hyperthermia.
The physiological role of heat shock proteins and their protective effects against various form of stress have been extensively studied (2, 20). Among HSPs, the possible mechanisms underlying the pro-tective effect of HSP70 include their function as molecular chaperons by refolding and renaturing the damaged proteins (2, 18, 20, 27). Inducible HO-1 also belongs to the heat shock protein family (HSP32). Overexpressing of HO-1 has a principal role in many cell types including endothelial cell adaptive response to oxidative damage (2, 18). HO-1 also has protective
effects in pulmonary and cardiovascular diseases (15). It is known that CO is one of the products of heme degradation by HO and results in vasorelaxation (15). Bilirubin, another product of heme degradation by HO, desensitizes mast cells and works as an anti-inflammatory substance (22). The present studies showed that significant increase of HO-1 protein was observed in heart and lung tissues in the heated rats Fig. 3. Expression of iNOS mRNA relative to PBGD mRNA
among different groups of rats using the same techniques described in Fig. 1. Rats (n = 6)/group.
Fig. 4. Heme oxygenase-1 (HO-1) and beta-tubulin in the heart, lung and liver tissues isolated from rats received various treatments by Western blotting and immunochemical staining. The ratio of HO-1 over beta-tubulin was used for comparison among different groups. Rats (n = 5)/group; *: P < 0.05.
subjected to anaphylactic treatment. Tamion et al. (23, 24) showed that induction of HO-1 reduces lung edema and prevents the systemic responses to hemor-rhagic shock. Recent studies also showed that HO-1 decrease the release of histamine from mast cells (16, 22). Therefore, HO-1 could decrease histamine re-lease through the same mechanism to prevent plasma leakage in the microvasculature of the heated rats in the present studies.
No significant change in the levels of iNOS (NOS type II) mRNA expression was detected in examined tissues in rats subjected to anaphylactic treatment as compared to control rats. The results are consist with previous observations that a lack of induction of iNOS in anaphylactic shock (14, 17, 21). However, increased production of NO via activation of the eNOS was found during anaphylactic hypoten-sion (14, 17). The present studies also showed that plasma nitrite and nitrate in the femoral vein in non-heated rats subjected to anaphylactic shock appeared to be the same as basal levels before anaphylactic shock. This discrepancy of enhanced NO production via activation of the eNOS may be due to enhanced NO production detected only at 3-4 min early periods (14) during anaphylactic hypotension rather than at 10 min for collecting plasma in the present studies. Another explanation was possibly due to detection of NO in peripheral tissues with a NO electrode (14) versus the plasma nitrite and nitrate assay based on the Griess reaction in the present studies. Other unknown mediators may also contribute anaphylactic hypotension since anaphylactic hypotension is not totally returned to normal levels by pretreatment of NOS inhibitors (17). Together, whether massive amount of NO is produced and the role of NO during anaphylactic shock needs further studies.
Basal levels of NO produced by constitutive eNOS play a role in the maintenance of microvascular integrity under physiological and pathophysiological conditions (10-12, 19). Increased eNOS mRNA ex-pression is induced by a variety of stimuli such as shear stress and exercise training (7, 8). Although the heated, anaphylactic rats showed a significant in-crease in the level of eNOS mRNA in heart and lung, eNOS protein levels appeared to be the same among groups. The present studies also showed that plasma nitrite and nitrate of heated rats subjected to anaphy-lactic treatment appeared to be the same as basal levels before anaphylactic treatment. Therefore, a direct and sensitive method is required to measure NO production in various tissues in rats treated with hyperthermia in further studies.
Table 2 Protein contents of eNOS in tissue isolated from rats received various treatments.
eNOS/protein * in various tissues
Rats received various treatments heart lung liver
Anaphylactic shock 62.3±17.0 32.2± 5.3 3.0±1.4
Heat and anaphylactic shock 68.1±19.3 46.7±21.9 7.2±2.3
Control 64.2±22.0 45.1±20.2 8.3±5.0
Heat only 57.9±11.1 32.3±10.7 4.2±3.8
*: eNOS/protein: pg/µµµµµg, mean±SD, n = 5
Fig. 5. Heme oxygenase-1 (HO-1) and beta-tubulin in tissue isolated from rats received various treatments by Western blotting and immunochemical staining in typical experiments. C: control without any treatment; H: heated, anaphylactic treatment; HC: one day after hyperthermia (only) treatment; H5: 5 days after hyperthermia (only) treatment; NH: non-heated, anaphylactic treatment.
It has been known that anaphylaxis includes the activation of mast cells or basophils and release of inflammatory mediators, like histamine, serotonin and bradykinin. The inflammatory mediators are thought to mediate body wide vasodilatation, increased permeability of post-capillary venules and protein leakage. Our previous studies (4) have shown that increased protein leakage in heart and lung tissues in rats subjected to anaphylactic shock is reduced by whole body hyperthermic pretreatment. The present studies showed that hyperthermic pretreatment in-creased production of HO-1 enzyme in various tis-sues of the heated rats subjected to anaphylactic shock as compared to both non-heated, anaphylactic rats and control rats. In summary, reduction of en-hanced permeability and attenuation of hypotension may be the result of overproduction of HO-1 rather than NOS in tissue in rats pretreated with whole body hyperthermia. Heat shock proteins like HO-1 could decrease histamine release to prevent plasma leakage in the microvasculature of heated rats.
Acknowledgments
These studies were supported by a grant from the National Science Council, Taiwan, R.O.C. (NSC-90-2314-B-037-094).
References
1. Barbe, M.F., Tytell, M., Gower, D.J. and Welch, W.J. Hyperther-mia protects against light damage in the rat retina. Science 241: 1817-1820, 1988.
2. Benjamin, I.J. and McMillan, D.R. Stress (heat shock) proteins: molecular chaperones in cardiovascular biology and disease. Circ. Res. 83: 117-132, 1998.
3. Chan, J.Y., Wang, S.H. and Chan, S.H. Differential roles of iNOS and nNOS at rostral ventrolateral medulla during experimental endotoxemia in the rat. Shock 15: 65-72, 2001.
4. Chen, S.C., Lu, T.S., Lee, H.L., Lue, S.I. and Yang, R.C. Hyperther-mic pretreatment decreases Hyperther-microvascular protein leakage and attenuates hypotension in anaphylactic shock in rats. Microvasc. Res. 61: 152-159, 2001.
5. Donnelly, T.J., Sievers, R.E., Vissern, F.L., Welch, W.J. and Wolfe, C.L. Heat shock protein induction in rat hearts. A role for improved myocardial salvage after ischemia and reperfusion? Circulation 85: 769-778, 1992.
6. Flohe, S., Dominguez Fernandez, E., Ackermann, M., Hirsch, T., Borgermann, J. and Schade, F.U. Endotoxin tolerance in rats: expression of TNF-alpha, IL-6, IL-10, VCAM-1 and HSP 70 in lung and liver during endotoxin shock. Cytokine 11: 796-804, 1999. 7. Forstermann, U., Boissel, J.P. and Kleinert, H. Expressional control
of the ‘constitutive’ isoforms of nitric oxide synthase (NOS I and NOS III). FASEB J. 12: 773-790, 1998.
8. Govers, R. and Rabelink, T.J. Cellular regulation of endothelial nitric oxide synthase. Am. J. Physiol. 280: F193-F206, 2001. 9. Hotchkiss, R., Nunnally, I., Lindquist, S., Taulien, J., Perdrizet, G.
and Karl, I. Hyperthermia protects mice against the lethal effects of endotoxin. Am. J. Physiol. 265: R1447-R1457, 1993.
10. Kubes, P. and Granger, D.N. Nitric oxide modulates microvascular permeability. Am. J. Physiol. 262: H611-H615, 1992.
11. Kurose, I., Kubes, P., Wolf, R., Anderson, D.C., Paulson, J., Miyasaka, M. and Granger, D.N. Inhibition of nitric oxide production. Mechanisms of vascular albumin leakage. Circ. Res. 73: 164-171, 1993.
12. Laszlo, F., Whittle, B.J., Evans, S.M. and Moncada, S. Association of microvascular leakage with induction of nitric oxide synthase: effects of nitric oxide synthase inhibitors in various organs. Eur. J. Pharmacol. 283: 47-53, 1995.
13. Lindquist, S. and Craig, E.A. The heat-shock proteins. Annu. Rev. Genet. 22: 631-677, 1988.
14. Mitsuhata, H., Shimizu, R. and Yokoyama, M.M. Role of nitric oxide in anaphylactic shock. J. Clin. Immunol. 15: 277-283, 1995. 15. Morse, D. and Choi, A.M. Heme oxygenase-1: the “emerging molecule” has arrived. Am. J. Respir. Cell. Mol. Biol. 27: 8-16, 2002.
16. Ndisang, J.F., Wang, R., Vannacci, A., Marzocca, C., Fantappie, O., Mazzanti, R., Mannaioni, P.F. and Masini, E. Haeme oxygenase-1 and cardiac anaphylaxis. Br. J. Pharmacol. 134: 1689-1696, 2001. 17. Osada, S., Ichiki, H., Oku, H., Ishiguro, K., Kunitomo, M. and Semma, M. Participation of nitric oxide in mouse anaphylactic hypotension. Eur. J. Pharmacol. 252: 347-350, 1994.
18. Pohlman, T.H. and Harlan, J.M. Adaptive responses of the endot-helium to stress. J. Surg. Res. 89: 85-119, 2000.
19. Rumbaut, R.E. and Huxley, V.H. Similar permeability responses to nitric oxide synthase inhibitors of venules from three animal species. Microvasc. Res. 64: 21-31, 2002.
20. Snoeckx, L.H., Cornelussen, R.N., Van Nieuwenhoven, F.A., Reneman, R.S. and Van Der Vusse, G.J. Heat shock proteins and cardiovascular pathophysiology. Physiol. Rev. 81: 1461-1497, 2001. 21. Szabo, C. Nitric oxide, perioxynitrite and poly(ADP-ribose)
syn-thetase in anaphylactic shock. Shock 8: 153-154, 1997.
22. Takamiya, R., Murakami, M., Kajimura, M., Goda, N., Makino, N., Takamiya, Y., Yamaguchi, T., Ishimura, Y., Hozumi, N. and Suematsu, M. Stabilization of mast cells by heme oxygenase-1: an anti-inflammatory role. Am. J. Physiol. 283: H861-H870, 2002. 23. Tamion, F., Richard, V., Bonmarchand, G., Leroy, J., Lebreton, J.
P. and Thuillez, C. Induction of heme-oxygenase-1 prevents the systemic responses to hemorrhagic shock. Am. J. Respir. Crit. Care Med. 164: 1933-1938, 2001.
24. Tamion, F., Richard, V., Lacoume, Y. and Thuillez, C. Intestinal preconditioning prevents systemic inflammatory response in hem-orrhagic shock. Role of HO-1. Am. J. Physiol. 283: G408-G414, 2002.
25. Thum, T., Haverich, A. and Borlak, J. Cellular dedifferentiation of endothelium is linked to activation and silencing of certain nuclear transcription factors: implications for endothelial dysfunction and vascular biology. FASEB J. 14: 740-751, 2000.
26. Villar, J., Ribeiro, S.P., Mullen, J.B., Kuliszewski, M., Post, M. and Slutsky, A.S. Induction of the heat shock response reduces mortal-ity rate and organ damage in a sepsis-induced acute lung injury model. Crit. Care Med. 22: 914-921, 1994.
27. Wynn, R.M., Davie, J.R., Cox, R.P. and Chuang, D.T. Molecular chaperones: heat-shock proteins, foldases, and matchmakers. J. Lab. Clin. Med. 124: 31-36, 1994.
28. Yen, C.P., Chen, S.C., Lin, T.K., Wu, S.C., Chang, C.Y., Lue, S.I., Jeng, A.Y., Kassell, N.F. and Kwan, A.L. CGS 26303 upregulates mRNA expression of heme oxygenase-1 in brain tissue of rats subjected to experimental subarachnoid hemorrhage. J. Cardiovasc. Pharmacol. 44 (suppl 1): S474-S478, 2004.