國立台東大學 生命科學研究所 碩士學位論文
指導教授 : 李炎 博士
新種洋菜分解菌 Alcaligenes sp. Strain Yen 所產洋菜酶純化與部分特性
Purification and some properties of agarase from a novel agarolytic bacterium,
Alcaligenes sp. Strain Yen
研究生 : 謝育峰 撰
中 華 民 國 97年 1月 24日
新種洋菜分解菌 Alcaligenes sp. Strain Yen 所產洋菜酶純化與部分特性
Purification and some properties of agarase from a novel agarolytic bacterium,
Alcaligenes sp. Strain Yen
研究生 : 謝育峰 Student:Yu-Fong Sie 指導教授 : 李炎 博士 Advisor:Yen Lee﹐Ph.D.
國立台東大學 生命科學研究所 碩士學位論文
Submitted to Department of Institute of Life Science National Taitung University
In Partial Fulfillment of the Requirements for the Degree of Master in Life Science
January 2008
Taitung, Taiwan, Republic of China
中 華 民 國 97年 1月 24日
致謝
首先誠摯的感謝指導教授李炎博士,老師悉心的教導使我得以一窺 碩士以及微生物領域的深奧,不時的討論並指點我正確的方向,使我在 這些年中獲益匪淺。老師對學問的嚴謹更是我輩學習的典範。
本論文的完成另外亦得感謝所辦大美女的惠嵐姐大力協助,及農改 場蘇得銓教授的支持、輔英科大食營系在硬體支援,以及中華醫事科技 大學廖東南教授對我研究上的指教評點。因為有你們的體諒及幫忙,使 得本論文能夠更完整而嚴謹。
兩年半裡的日子,實驗室裡共同的生活點滴,學術上的討論、言不 及義的閒扯、讓人又愛又怕的宵夜、趕作業的革命情感、因為睡太晚而 遮遮掩掩閃進實驗室...,感謝眾位學長姐、同學、學弟妹的共同 砥礪,你/妳們的陪伴讓兩年的研究生活變得絢麗多彩。
感謝尚澄學長、惠春、佳蓮學姐們不厭其煩的指出我研究中的缺 失,且總能在我迷惘時為我解惑,也感謝大胖、智翔、玟玲同學的幫忙,
恭喜我們順利走過這兩年。實驗室的世慶、世峯、年展、仲欣、信樺學 弟、徐于、莉雪學妹們當然也不能忘記,妳的幫忙及搞笑我銘感在心。
親密戰友孟慈在背後的默默支持更是我前進的動力,沒有孟慈的體諒、
包容,相信這兩年的生活將是很不一樣的光景。最後,謹以此文獻給我 摯愛的雙親。
中 文 摘 要
本實驗室於台東市區排水溝中,篩選到一株洋菜分解菌。經實驗發 現此菌株為革蘭氏陰性桿菌,不發酵、具運動性、好氧、catalase、oxidase 為正反應,且不產 H2S 和 Indole。菌株經生化試驗和 16S rDNA 序列 比對,將此洋菜分解菌鑑定為 Alcaligenes sp. Strain Yen (楊,2007)。
將此菌於 C-10 培養液中振盪培養( 35℃、150 rpm ) 48 hr,收集離 心去除菌體後之培養液以 20~80% 硫酸銨劃分及 Sephadex G-100 膠 體過濾層析法進行洋菜酶的分離與純化,獲得洋菜酶。首先,培養液經 硫酸銨劃分透析後之比活性為 157.33 U/mg,純化倍數為 2.19 倍,回 收率為 54.61 %;再以 Sephadex G-100 膠體管柱純化,分劃具有洋菜 能力的蛋白質波峰劃分區域,收集後命名為 Yen-agarase,其比活性分 別為 406.44 U/mg,純化倍數分別為 5.66 倍,回收率分別為7.96 %。
Yen-agarase 其最適作用 pH 值為 7.5-8.5,最適作用溫度為 35℃,但是於 55℃下反應 30 min 活性減少 50 %,其分子量經由 SDS-PAGE 後推估為 110 KDa。以 high-melting-point agarose 為基質時 所表現之基質特異性活性最佳,此洋菜酶不能分解ι-鹿角藻(ι
-arrageenan ) 和κ-鹿角藻(κ-arrageenan ) 但是卻可以分解藻酸鈉 ( sodium alginate )。
使用 agarose 做為酵素反應基質進行水解試驗,並將水解產物以薄 層色層分析( TLC ) 展開,經由判定水解後所得為新洋菜寡醣類,推測 該酵素作用位置為 β-1,4 鍵結,初步推估此酵素為 β 型。
Abstract
The agar-degrading bacterium was isolated from Taitung, Taiwan. After biochemical tests and 16S rDNA sequencing, we found the agar-degrading bacterium was a bacillus, gram-negative, mobile, aerobic, catalase and oxidase positive, didn't produce H2S and Indole.
This bacterium was identified as Alcaligenes sp. strain Yen (AY744384).
Strain Yen was grown at RT for 48 hr in C-10 broth with shaking(150 rpm). After removing the cell debries, crude enzyme was collected, and then proceeded to separation and purification. First step was to concentrate and dialyse the crude enzyme, and to purify it, and yielded a solution with 157.33 U/mg(purified 2.19 fold), and production rate was 54.61%. Then Sephadex G-100 chromatographs was used to final purify Yen-agarase. The results showed its specific activity was 406.44 U/mg(purified 5.66 fold), and 7.96% production rate.
The optimum pH for Yen-agarase Ι was found to be 7.0-8.0. The optimum temperature for Yen-agarase Ι was 35℃, but approximately 50%
of the enzyme activity was inactivated in 30 minutes under 55℃. Estimation of the molecular mass by SDS-PAGE was 101 kDa. The best substrate
specificity activities of Yen-agarase toward high-melting-point agarose were D-galactose. Yen-agarase did not have the capability to degradeι
-carrageenan and κ-carrageenan. The agarose water solution was used to carry on experiments, and solved outcome launched on thin-layer
chromatography( TLC). By judging the dissolved carbohydrates produced by neoagarooligosaccharidase, we could relize the enzyme functional position was β -1,4 band, so, the enzyme as β type.
目 錄
中 文 摘 要· · · i
A b s t r a c t · · · i i 目 錄· · · i i i 圖 目 錄· · · v i i i 表 目 錄· · · x i 壹 、 前 言· · · 1
貳 、 文 獻 整 理 · · · 3
一 、 洋 菜 的 組 成 與 特 性 · · · 3
二 、 洋 菜 酶 生 產 菌 株 及 其 分 離 來 源 · · · 4
三 、 洋 菜 酶 對 洋 菜 水 解 的 作 用 機 制 · · · 4
四 、 洋 菜 酶 作 用 後 之 水 解 產 物 · · · 5
4 . 1 作 用 機 制 與 水 解 產 物 · · · 5
4 . 2 洋 菜 酶 之 受 質 特 異 性 · · · 6
五 、 洋 菜 酶 之 生 化 特 性 · · · 7
5 . 1 溫 度 和 p H 值 · · · 7
5 . 2 分 子 量 · · · 7
5 . 3 金 屬 離 子 · · · 8
六 、 洋 菜 酶 之 純 化 · · · 9
6 . 1 硫 酸 銨 鹽 沉 澱 · · · 9
6.2 離子交換層析法···9
6.3 膠體過濾層析法···10
6.4 親和性管柱層析法···10 七 、 洋 菜 水 解 產 物 之 分 析 · · · 1 1 7.1 洋菜水解產物之獲得···11 7.2 薄層層析法(TLC)···11 7 . 3 高 效 能 液 相 層 析 法 ( H P L C ) · · · 1 1 八 、 洋 菜 水 解 產 物 之 應 用 · · · 1 2 8.1 海藻多醣之生理活性···12 8.2 非消化性碳水化合物之生理活性···12 九 、 洋 菜 酶 的 應 用 · · · 1 3 9 . 1 細 胞 的 分 離 · · · 1 3 9 . 2 D N A 或 R N A 分 離 · · · 1 3 9 . 3 藻 類 原 生 質 體 的 製 作 · · · 1 3 9 . 4 融 合 瘤 細 胞 分 離 · · · 1 4 9 . 5 開 發 海 藻 寡 糖 之 健 康 食 品 · · · 1 5 参 、 實 驗 設 計· · · 1 6 肆 、 實 驗 材 料 與 方 法· · · 1 7 一 、 藥 品 與 耗 材 · · · 1 7 二 、 儀 器 與 設 備 · · · 1 7 三 、 菌 種 鑑 定 · · · 1 8 3 . 1 菌 種 生 化 鑑 定 · · · 1 8 3 . 2 菌 種 1 6 S r D N A 分 析 · · · 1 8 四 、 培 養 基 配 製 與 菌 株 之 保 存 · · · 1 8 4 . 1 培 養 基 配 製 · · · 1 8
4 . 1 . 1 菌 株 液 體 生 長 培 養 基 · · · 1 8 4 . 1 . 2 菌 株 S 3 P Y 固 體 生 長 培 養 基 · · · 1 9 4 . 2 菌 株 保 存 · · · 1 9 五 、 菌 株 之 活 化 · · · 1 9 六 、 實 驗 方 法 · · · 2 0 6 . 1 洋 菜 酶 之 生 產 · · · 2 0 6 . 2 洋 菜 酶 的 活 性 測 定 · · · 2 0 6 . 2 . 1 還 原 糖 酵 素 活 性 測 定 酵 素 活 性 · · · 2 0 6 . 2 . 2 活 性 單 位 定 義 · · · 2 0 6 . 2 . 3 蛋 白 質 的 定 量 · · · 2 1 6 . 3 洋 菜 酶 之 分 離 純 化 · · · 2 1 6 . 3 . 1 膠 體 過 濾 層 析 ( S e p h a d e x G - 1 0 0 ) · · · 2 1 6 . 4 洋 菜 酶 之 生 化 性 質 測 試 · · · 2 1 6 . 4 . 1 分 子 量 的 鑑 定 · · · 2 1 6 . 4 . 2 S D S - P A G E 的 材 料 製 備 · · · 2 2 6 . 4 . 3 S D S - P A G E 分 析 操 作 步 驟 · · · 2 3 6 . 5 N a t i v e P A G E 分 析 · · · 2 3 6 . 6 洋 菜 酶 活 性 染 色 分 析 · · · 2 3 七 、 基 質 特 異 性 · · · 2 3 八 、 最 適 作 用 p H 值 · · · 2 4 九 、 p H 值 的 安 定 性 · · · 2 4 十 、 最 適 作 用 溫 度 · · · 2 4 十 一 、 溫 度 安 定 性 · · · 2 5 十 二 、 金 屬 離 子 的 影 響 · · · 2 5
1 2 . 1 鈣 離 子 的 影 響 · · · 2 5 1 2 . 2 鈉 離 子 的 影 響 · · · 2 5 1 2 . 3 其 他 金 屬 離 子 耐 受 度 的 影 響 · · · 2 5 十 三 、 酵 素 動 力 學 · · · 2 6 十 四 、 水 解 產 物 分 析 ( T L C ) · · · 2 6 1 4 . 1 T L C 分 析 水 解 產 物 · · · 2 6 伍 、 結 果 與 討 論· · · 2 7 一 、 Y e n 菌 種 鑑 定 · · · 2 7 二 、 酵 素 的 純 化 · · · 2 7 1 . 粗 酵 素 液 的 製 備 · · · 2 7 2 . S e p h a d e x G - 1 0 0 膠 體 過 濾 層 析 · · · 2 8 三 、 洋 菜 酶 的 生 化 特 性 · · · 2 8 1 . 分 子 量 · · · 2 9 2 . 基 質 特 異 性 · · · 2 9 3 . 最 適 作 用 p H 值 · · · 3 0 4 . p H 值 的 安 定 性 · · · 3 1 5 . 最 適 作 用 溫 度 · · · 3 1 6 . 溫 度 安 定 性 · · · 3 2 7 . 鈣 離 子 的 影 響 · · · 3 3 8 . 鈉 離 子 的 影 響 · · · 3 4 9 . 其 他 金 屬 離 子 耐 受 度 的 影 響 · · · 3 5 四 、 酵 素 動 力 學 ( K m 、 V m a x ) · · · 3 5 五 、 洋 菜 酶 之 分 解 產 物 的 探 討 · · · 3 5 陸 、 結 論· · · 3 8
柒 、 參 考 文 獻 · · · 3 9 附 錄 · · · 6 9
圖 目 錄
圖一、Yen 在 C-10 培養基上,在室溫下培養2天之菌落形態···51
圖二、Yen 在 C-10 固體培養基上,在室溫下培養2天後,利用碘 液來偵測洋菜酶活性···52
圖三、Yen 之16S rDNA 序列定序結果···53
圖四、Yen 之洋菜酶 SDS-PAGE CBR 染色染···54
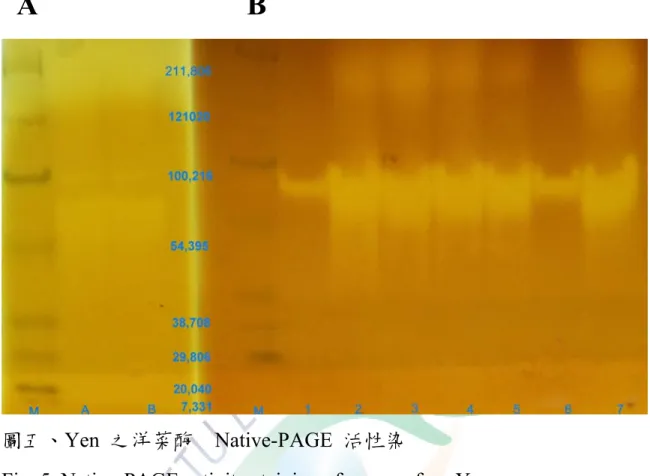
圖五、Yen 之洋菜酶 Native-PAGE 活性染···55
圖六、Yen-agarases 於不同緩衝液中的最適作用 pH 值···56
圖七、pH 值對 Yen-agarases 之安定性···57
圖八、Yen-agarases 的最適反應溫度···58
圖九、Yen-agarases 對溫度穩定性分析···59
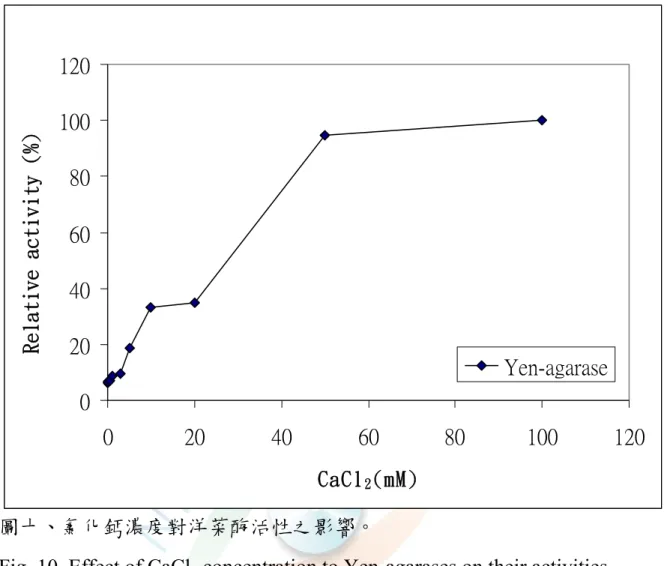
圖十、氯化鈣濃度對各洋菜酶活性之影響···60
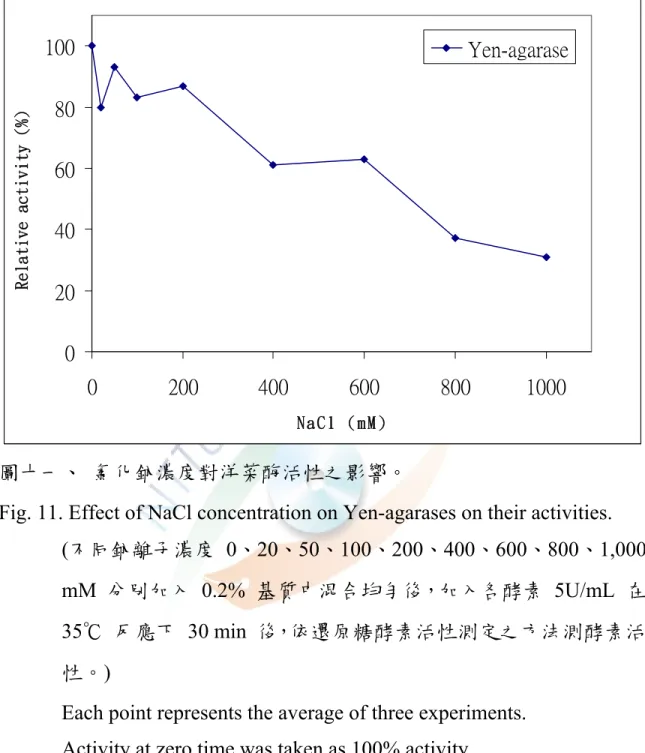
圖十一、氯化鈉濃度對各洋菜酶活性之影響···61
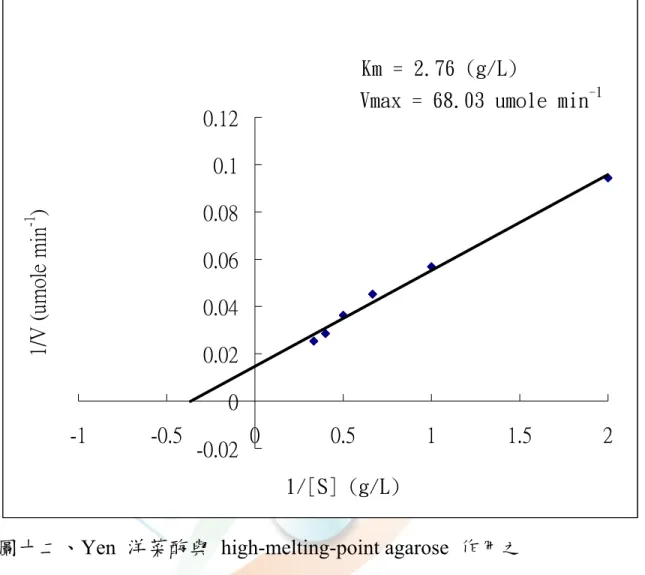
圖十二、Yen 粗洋菜酶與 high-melting-point agarose 作用之 Lineweaver-Burk 圖···62
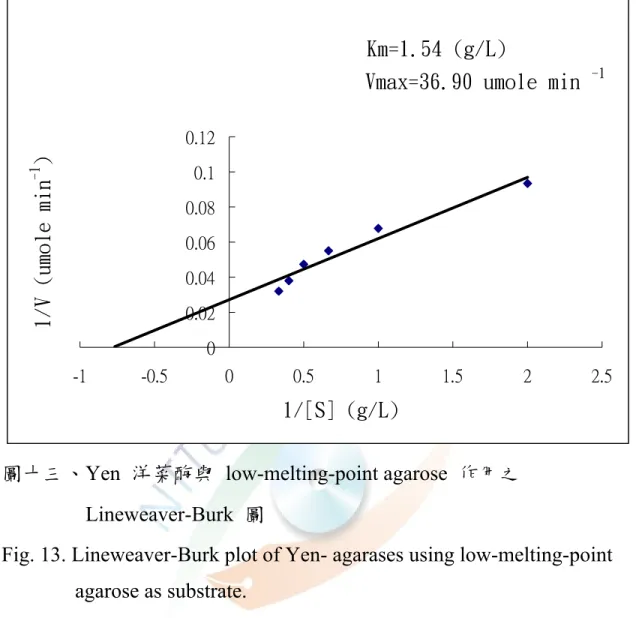
圖十三、Yen 粗洋菜酶與 Low-melting-point agarose 作用之 Lineweaver-Burk 圖···63
圖十四、以 TLC 分析 Yen- agarases 水解洋菜糖之水解產物···64
表 目 錄
表一、 Alcaligenes sp. strain Yen 生化特徵表···65
表二、Yen 洋菜酶之純化表···66
表三、 Yen-agarases 對九種多糖之基質特異性···67
表四、 金屬離子對 Yen-agarases 活性之影響···68
壹、前言
洋菜( agar )又名瓊脂,主要是由紅藻綱( Rhodophyceae )的藻類粹取 出的生物體分子,海藻細胞壁的多醣是構成洋菜的主成分,目前廣泛地 被應用於食品工業與微生物實驗的膠體凝結劑。中性洋菜(agarose)為其 主要的凝結成分,其組成為一連串重複的1,4鍵結的3,6-脫水左旋半乳糖 (4-O-linked 3,6-anhydro-β-L- galactopyranose)與1,3鍵結的右旋半乳糖 (3-O-linkedα-D- galactopyranose)。
早在 1902 年,Gran 首先發現會水解洋菜的海洋細菌(顏,2002),
這些菌所分泌出的酵素通稱洋菜酶( agarase ),目前這些洋菜酶可依照對 agarose 作用模式分成兩大類,一為 α-洋菜酶( α-agarase ),另一為 β- 洋菜酶( β-agarase )等兩類。α-agarase 會作用於中性洋菜之α-(1→3)鍵 結,所產生寡醣產物稱為洋菜寡醣( agarooligosacchardies ),β-agarase 會 作用於中性洋菜之β-(1→4) 鍵結,作用後所產生寡醣產物稱為新洋菜寡 醣( neoagarooligosaccharidase )
( Hosoda et al., 2003; Kang et al., 2003; Sugano et al., 1993b )。
經洋菜酶分解洋菜所得新洋菜寡醣具有低熱量、防止澱粉老化及抑 制細菌生長等特性(德永和日高,1991),且新洋菜二糖 ( neoagarobiose ) 具有吸濕能力以及讓 B16 murine melanoma cells 變白效果( Kobayashi et al., 1997 )。另外,新洋菜寡糖亦具不同生理活性,如條斑紫菜
( Porphyra yezoensis ) 之萃取多醣,經 β-agarase 分解後, 於體外實驗 能刺激巨噬細胞活性( Yoshizawa et al., 1995 )。江蘺( Gracilaria
verrucosa ) 水溶性多醣經 β-agarase 降解後,具刺激小鼠巨噬細胞產生
能力( Yoshizawa et al., 1996 )。使用Arthrobacter sp. S-22 所產 β-agarase 分解紫菜多醣( porphyran )後,所得新洋菜四糖( neoagarotetraose )、硫化 寡糖( sulphated oligosaccharide )、硫化四糖( sulphated tetrasaccharide )、
及硫化二糖( sulphated disaccharide )等寡醣具有降低小鼠總膽固醇程度 的能力( Osumi et al., 1998 )。2001 年 Kong 的報告中認為洋菜寡醣具 疫調節能力,可使老鼠脾臟細胞增生,因此認為添加洋菜寡醣於食品中 可以提高人體的免疫活性。此外洋菜酶的應用很廣,如細胞的分離 ( Carlsson and Malmqvist,1977; Solursh, 1991 )、DNA 的分離( Couto et al., 1989; Kamiunten, 1995; Osoegawa et al., 1998; She, 2003; Shizuya and Kouros-Mehr, 2001; Singh et al., 2003 )、藻類原生質體的製作( Araki et al., 1998a; Chen and Chiang, 1994 )、融合瘤細胞分離( Gray et al., 1995 )等。
本研究室於台東市區水溝中發現一株具有洋菜分解酶基因質體之 洋菜分解菌,且已成功將洋菜分解質體轉殖到 E. coli 中表現,基於洋 菜酶及其產物在生理活性、免疫調節、食品添加和生物科技應用上,有 很大的發展,本研究工作內容為就本實驗室 Alcaligenes sp. Strain Yen 所生產洋菜酶純化分離為單一酵素,並瞭解洋菜酶所具有多各種生化特 性及其作用產物且利用其作用產物來初步推斷洋菜分解酶的分型,藉著 探討其特性進一步了解洋菜酶和產物的發展性。
貳、文獻探討
一、洋菜的組成與特性
藻類是廣泛存在於江、湖、海洋中的水生植物,其含有各種不飽和 脂肪酸、維生素,成分會隨產地、季節、種類而有所不同,是水中生物 之食物鏈的一環,有些藻類的細胞壁中可萃取得到的物質稱為洋菜( Ha et al., 1997 ),海藻根據藻類所含的儲存性及細胞壁多醣可分為三個綱︰
綠藻( Chlorophyceae )、褐藻( Phaeophyceae )和紅藻( Rhodophyceae ),依 照 US Pharmacopoeia 對洋菜的定義,洋菜是從 Rhodophyceae 這個綱 的某些海藻中所萃取出的親水性膠體,它的顏色由白色到淡黃色,是一 種無味或是略有少許的特殊香味及黏性物質。洋菜不溶於冷水,但在 80
℃以上之熱水中則會膨潤快速溶解,回復至較低的溫度時,由中性洋菜 所形成的雙螺旋鏈會聚集在一起而形成結構較為鬆散的束群,而此現象 即稱之為 Gel I state。隨後當溫度繼續降低時,這些成束的中性洋菜會 繼續與彼此結合形成我們所見的洋菜膠,而此階段即稱為 Gel II state (李,2005)。
洋菜( agar )又名瓊脂,主要是從藻類中萃取得到,海藻細胞壁的多 醣是構成洋菜的主成分,在 1902 年,Gran 首先發現會水解洋菜的細 菌(顏,2002 ),這些菌所分泌出的酵素通稱洋菜酶( agarase )。洋菜是一 種多醣類,主要成分為聚半乳糖,其含量大約為70%,而聚半乳糖成分 主要為酸性洋菜( agaropectin )和中性洋菜二種構造,中性洋菜形成凝膠 的分枝結構,是由1→3鍵結的 β-D-galactose 和1→4鍵結的
3,6-anhydro-α-L-galactose 交互聚合而成的長鏈聚合物(附錄二),酸性洋
菜為非凝膠部分,它是以中性洋菜做為基本架構,再加上複雜的支鏈如 硫酸鹽、葡萄糖醛酸和丙酮酸醛等所構成,但它的詳細結構還沒有被研 究出來( Duckworth and Yaphe,1971; Ohta, Y. et al.,2004 )(附錄二)。
二、洋菜酶生產菌株及其分離來源
Swartz and Gordon 在 1959 年的文獻中指出,早於 1902年 Gran 已分離出具分解洋菜的海洋細菌,經鑑定為 Bacillus gelaticus。自此之 後,可從文獻中得知越來越多能夠分解洋菜的菌株被發現,如
Achromobacter、Agarbacterium、Alteromonas、Cytophaga、
Pseudoalteromonas、Pseudoalteromonas、Vibrio( Leon et al. 1992; Zhong,Z.
et al. 2001 )。Vera et al. 於 1998 年認為由於大分子的洋菜無法進入細 胞內,所以這些細菌就分泌出洋菜酶,將洋菜分解成洋菜寡醣、雙醣或 單醣做為碳源。這些生產洋菜酶菌株大部分分離自沿岸海水及河口環 境,但也有許多是分離自陸上之淡水、廢水及土壤,或其他環境中。目 前陳( 2006 ) 將已發表洋菜酶生產菌株種類、所屬洋菜酶種類及分離來 源之文獻整理於附錄三。
三、洋菜酶對洋菜水解的作用機制
洋菜酶為可將洋菜分解成寡醣的一種水解酵素,Allouch et al.
2003,認為洋菜酶作用形式類似醣苷水解酶( glycoside hydrolases
,GH )(李,2005)。這些洋菜酶可依照對中性洋菜作用模式分成兩大類,
一為α-洋菜酶( α-agarase ),另一為β-洋菜酶( β-agarase )等兩類。
α-agarase 會作用於中性洋菜之 α-(1→3) 鍵結,所產生寡醣產物稱為洋 菜寡醣,此洋菜寡醣產物在還原端具 3,6-anhydro-L-galactose 殘基
( residue )及非還原端的 D-半乳糖( D-galactose ) ( Hosoda et al., 2003;
Kang et al., 2003; Sugano et al., 1993b; Young et al., 1978 )。β-agarase 會 作用於中性洋菜之 β-(1→4) 鍵結,作用後所產生寡醣產物稱為新洋菜 寡醣,此新洋菜寡醣產物在還原端為 D-galactose 而非還原端為
3,6-anhydro-L- galactose( Hosodaet al., 2003; Kang et al., 2003; Sugano et al., 1993b )。
洋菜酶對洋菜作用具有明顯的特徵,如:使得洋菜膠表面軟化形成 凹陷並且有透明圈( clear zone )產生或使洋菜膠完全水解成液體。
四、洋菜酶作用後之水解產物 4.1 作用機制與水解產物
不同的洋菜酶水解洋菜,產生的水解產物不完全相同。洋菜酶水解 洋菜主要的產物為新洋菜雙糖、新洋菜四糖及新洋菜六糖
( neoagarohexaose )。微生物產生各種洋菜酶是需要被誘導,且洋菜酶對 誘導物具有專一性,不同的誘導物可使同一菌株產生不一樣的洋菜酶 ( Hosoda et al., 2003 )。而目前洋菜酶水解洋菜的作用機制,研究較為詳 細的作用機製是菌株是Pseudoalteromonas atlantica ATCC19292 及 Alteromonas agarlyticus strain GJ1B。
微生物所產洋菜酶以 β-agarase 為主,首先洋菜會誘導
Pseudomonas atlantica ATCC 19292 的胞外 β-agarase 作用於洋菜 β-(1→4) 鍵結,產生一系列新洋菜寡醣,而此醣會誘導新洋菜寡醣水解 酶( neoagarotetraose hydrolase )產生,最後把寡糖切成新洋菜四糖。接著 與細胞連結的第 II 型外切 β-洋菜酶( exo β-agarase II ),可以將新洋菜 四糖中間的 β( 1,4 ) 鍵結水解打斷,而產生新洋菜二糖,最後新洋菜
二糖再誘導細胞間質α型新洋菜雙糖水解酶( α-neoagarobiose hydrolase ) 產生,並作用於新洋菜二糖,最後形成 D-galactose 和
3,6-anhydro-L-galactose 等產物給予細胞間的酵素代謝利用( Hosoda et al., 2003 )。
Alteromonas agarlyticus strain GJ1B 的作用機制方面,Alteromonas agarlyticus strain GJ1B 含有兩種酵素 α-garase 和 β-半乳糖苷酶 ( β-galactosidase ),α-garase 可將洋菜水解成洋菜寡糖( agaro-
oligosaccharide )其最小產物為洋菜四糖,而 β-galactosidase 作用於洋菜 寡糖的非還原端,最終產物洋菜三糖( agarotriose ) ( Vera et al., 1998 )。
微生物會被洋菜誘導產生的各種洋菜酶,陳(2006)將洋菜水解所得知產 物整理如附錄四。
4.2 洋菜酶之受質特異性
Alteromonas sp. E-1 的 β 型洋菜酶無法水解 κ 型鹿角藻
( κ-carrageenan )、藻酸鈉 ( sodium alginate ) ( Kirimura et al., 1999 );
Pseudomonas sp. PT-5 的洋菜酶水解洋菜、鹼處理紫菜( porphyran )、
高熔點的洋菜糖、低熔點的洋菜糖等基質 30 分鐘,產生 32-37 % 還 原糖,而水解紫菜只有產生 20 % 還原糖,無法水解 κ 型鹿角藻、 λ 型鹿角藻、阿拉伯橡膠 ( gum Arabic galactan ) ( Yamaura et al., 1991 );
Pseudomonas vesicularis MA103 的洋菜酶 I 水解 0.2 % 洋菜糖、高熔 點的洋菜糖、低熔點的洋菜糖等基質,所產生的還原糖量分別為 182、
170、 67 mmole min/ mg (陳,2006 );Vibiro sp. JT0107 的 β 型 洋菜酶無法水解 ι 型、 κ 型、 λ 型鹿角藻、藻酸鈣 ( Sugano et al., 1993,1995 ) 。由前人研究發現目前已知洋菜酶都無法水解 ι 型、 κ
型、 λ 型鹿角藻、藻酸鈉,但可以水解洋菜糖、高熔點的洋菜糖、低 熔點的洋菜糖。
五、洋菜酶之生化特性 5.1 溫度和 pH 值
目前已發表文獻之洋菜酶最適作用溫度及 pH 值之範圍已經於陳 (2006) 整理如附錄五。文獻中微生物所產洋菜酶最適作用溫度分布在 25℃到60℃之間,不同的洋菜酶其最適作用 pH 值不太相同,大部分的 洋菜酶之最適 pH 值為 pH 6.5~7.5 之間。
5.2 分子量
Vera et al. (1998) 根據文獻中洋菜酶分子量大小,共分成三類。第 一類為分子量在 50 kDa 以下,如 Pseudoalteromonas altantica
ATCC19292 ( Morrice et al., 1983a ) 和 Pseudomonas sp. PT-5 ( Yamaura et al., 1991 )所產洋菜酶之分子量皆為 31 kDa,而Pseudoalteromonas antarctica N-1 ( Vera et al., 1998 )所產洋菜酶分子量則為 33 kDa。第二 類則為分子量介於 50 kDa 與 100 kDa 之間,如Alteromonas sp. C-1 ( Leon et al., 1992 ) 洋菜酶分子量為 52 kDa,而 Pseudomonas sp. W7 ( Ha et al., 1997 ) 的洋菜酶分子量為59kDa ,Pseudoalteromonas
atlantica T6c ( Belas et al., 1988 ) 則生產分子量為55 kDa 的洋菜酶。第 三類則為分子量大於 100 kDa 的洋菜酶,如Vibrio sp. JT107 ( Sugano et al., 1993a ) 所產洋菜酶,分子量為 105 kDa。1998年 Vera et al. 指出能 生產前兩種洋菜酶分子量之菌株,在固態洋菜培養基上,菌落周圍會呈 現軟化及透明狀,這可能與小分子量洋菜酶能夠經洋菜膠體間的孔徑擴
散而出,進而分解洋菜多醣造成菌株周圍形成軟化現象,至於菌落周圍 發生液化現象者,則可能為菌株產生大量洋菜酶所致。
5.3 金屬離子
由於大多數產洋菜酶的菌株是海洋性細菌,Ha et al. (1997) 認為添 加適量的氯化鈉( NaCl )可以增加洋菜酶活性。另外,如添加其他金屬離 子和化學藥劑也會對微生物所產洋菜酶有不同的影響,如 Alteromonas sp. C-1 所產洋菜酶在鹽濃度低於 0.1 M 時,洋菜酶活性會被抑制,但 是於其他 0.1 M 金屬鹽濃度在時,會有更好的洋菜酶活性( Leon et al., 1992 )。而 Pseudomonas sp. PT-5 所產的洋菜酶被 Zn2+、Cu2+、Co2+、 Fe2+ 和 Al3+ 等金屬離子抑制( Yamaura et al., 1991 ) 。Pseudomonas sp.
W7 的 β-agarase 在 0.9 M 氯化鈉下活性會增加,Pseudomonas atlantic 的 β-neoagarotetraose hydrolase、neoagarobiose hydrolase 分別在
0.05-0.20 M、0.10 M 鹽濃度下,洋菜酶活性會增加( Ha et al., 1997 )。
添加 0.002 M 的 CaCl2 和 0.005 M 的 MgCl2 可提高活性1.2倍
( Morrice et al., 1983a );1 mM Hg2+ 可以完全抑制 Vibrio sp.PO-303 的 agarase-a、agarase-b、agarase-c 的活性,1 mM Al3+ 及 EDTA 對 Vibrio sp. PO-303 的 agarase-c 有強的抑制效果,但對 agarase-a、agarase-b 的 活性表現卻無影響( Araki et al., 1998 )。添加 2 mM Mn2+ 、Cu2+ 、 Zn2+ 、Fe2+ 和 Hg2+ 可完全抑制 Alteromonas sp. E-1 的 β-agarase 的 活性( Kirimura et al., 1999)。
六、洋菜酶之純化 6.1 硫酸銨鹽沉澱
鹽析沉澱法可分離出培養液中的蛋白質。蛋白質在水溶液中的溶解 度,會因溶液中鹽類濃度的改變而增減,可利用來沉澱蛋白質。硫酸銨 的兩價離子,在溶液中搶走附在蛋白質表面的水分子,使得非極性部分 相互吸引而聚集沉澱。各種蛋白質,因其表面的非極性區域分佈不同,
各在其特定的硫酸銨飽和濃度下沉澱,常用於蛋白質純化步驟。其硫銨 沈澱所使用的濃度有 40 ~ 60 % ( Yamaura et al., 1991 ) 、70 % ( Potin et al., 1993;Sugano et al., 1994 )、75 % ( Aoki et al., 1990; Leon et al., 1992;Vera et al., 1998 )、80 % ( Duckworth and Turvey, 1969;Araki et al., 1998 )、60 ~ 80 % ( Kirimura et al., 1999 )、 90 % ( Morrice et al., 1983a;
Sugano et al., 1993 )。
6.2 離子交換層析法
離子交換層析法的固定相是擔體上的電荷基團,可為正或負電荷;
若擔體帶有正電荷,可吸附著負電荷分子,則為陰離子交換法;不帶淨 電荷的分子,及陽離子則直接流出,不會被吸附上去。蛋白質通常以離 子鍵吸附,因此要流洗出蛋白質時,以提高鹽濃度來達成,大多使用 NaCl ;也可改變緩衝液的 pH 值,使得蛋白質的電荷改變而溶離出來。
帶有較高電荷比的蛋白質,與擔體的結合就愈緊密,溶離所需使用的鹽 濃度就較高。分離洋菜酶常用的離子交換層膠體如下:DEAE-Toyopearl 650M ( Araki et al.,1998;Kirimura et al., 1999 )、Mono Q ( Araki et al., 1998;Kirimura et al.,1999;Potin et al., 1993;Sugano et al., 1994 )、
QAE-Toyopearl column ( Sugano et al., 1994 )、DEAE-cellulose( Leon et al.,
1992;Vera et al., 1998 )、DEAE–Sepharose CL-6B( Morrice et al., 1983a;
Kim et al., 1999 )、 DEAE-Fractogel 650 M( Aoki et al., 1990 )。
6.3 膠體過濾層析法
膠體過濾層析法,以蛋白質分子量大小來分離蛋白質,如樣本的分 子量小,則容易進入膠體中,因此會被滯留在膠體中,較慢被溶離出來,
可以與分子量較大的樣本分離。分離洋菜酶常用的膠體過濾膠體如下:
Sephacryl S-100HR (陳,2006 )、Superose 12 ( Sugano et al., 1994 )、
Superdax 200 ( Sugano et al., 1994 )、Sephadex G-75( Leon et al., 1992 ; Vera et al., 1998 )、Sephadex G-100 ( Morrice et al., 1983a )、 Sephadex G-200 ( Duckworth and Turvey, 1969;Malmqvist et al., 1978; Sugano et al., 1994;Kirimura et al.,1999 )、Fractogel HW-55 column ( Duckworth and Turvey, 1969 )、Toyopearl-HW 50S ( Aoki et al., 1990、Yamaura et al., 1991 )
6.4 親和性管柱層析法
Sepharose 膠體以洋菜糖為主要材料,所以在分離洋菜酶上,適合 作為親和性管柱膠體,分離洋菜酶常用的親和性管柱膠體如下:
Sepharose CL-6B (陳,2006、Aoki et al., 1990;Potin et al., 1993 )、divinyl sulphone ( DVS )-Sepharose 4B ( Malmqvist et al., 1978 )、
A-8010G(Hydroxyapatite) ( Malmqvist et al., 1978;Morrice et al., 1983a.;
Sugano et al., 1994;Yamaura et al., 1991 )
七、洋菜水解產物之分析 7.1 洋菜水解產物之獲得
洋菜水解產物之獲得,可藉由酸水解、鹼水解、酵素水解而獲得。
酸、鹼水解主要是打斷 α ( 1, 3 ) 鍵結( Araki, 1956 ),所以水解產物主 要是以 agarobiose 為基本結構的寡糖;酵素水解主要是打斷 β ( 1, 4 ) 鍵結,少部分可以打斷 α ( 1, 3 ) 鍵結,所以酵素的水解產物主要是以 neoagarobiose 為基本結構的糖,但因為洋菜酶種類繁多,作用機制也不 完全相同,所以酵素水解產物比酸和鹼水解產物更為豐富多變化。且因 為酵素專一性,可利用不同的洋菜酶生產不同的水解產物,前人研究也 發現,有些洋菜酶會切醣類的特定端點,產生特定的產物( Suzuki, H. et al., 2003 )。
7.2 薄層層析法( Thin-layer chromatography, TLC ) ( Ohta et al., 2004 ) 最常被用來分析洋菜水解產物的方法,此法可以快速方便的分析洋 菜水解產生的寡糖類。常見的步驟是把洋菜水解產物,點在薄層層析片 上,再放入展開液中,洋菜水解產物會隨著展開液展開,不同的寡多糖 因為其本身對液相及固相的親和力不同,因此展開的速度不同,和標準 品新洋菜六糖、四糖、雙糖的 Rf 值比較可得知水解產物。
7.3 高效能液相層析法( High performance Liquid Chromatography,
HPLC )
HPLC 比 TLC 更靈敏,可偵測到比較微量的水解產物 ( Vera et al., 1998 )。且HPLC 也可利用波峰的面積,計算出較精準之水解產物量。
八、洋菜水解產物之應用
經洋菜酶分解洋菜所得新洋菜寡醣具有低熱量、防止澱粉老化及抑 制細菌生長等特性(德永和日高,1991),且 neoagarobiose 具有吸濕能 力以及讓 B16 murine melanoma cells 變白效果( Kobayashi et al.,
1997 )。
8.1 海藻多醣之生理活性
由於海藻中富含硫酸根多醣類與肝素( heparin )類,具有抗血液凝 固、血液淨化及降低血液中的三甘油脂作用( Noda et al., 1989 )。褐藻 Hijikia fusiforme 的熱水萃取物中多醣類的部分被證實可增進老鼠脾臟 細胞的增生反應,特別是 B cell ( Osumi et al., 1998 )。研究發現以 Pseudomonas atlantica 產生之 β 型洋菜酶水解條斑紫菜得到之寡醣水 溶液,在生物體外進行巨噬細胞培養實驗,結果發現以 β 型洋菜酶處 理之寡醣水溶液具有提高巨噬細胞之免疫能力( Yoshizawa et al.,
1995 )。紫菜( Porphra )經洋菜酶水解所得之水解產物經純化後,發現具 有新洋菜四糖、硫酸寡醣、硫酸四醣、硫酸醣等部分,經餵食老鼠後發 現都具有降低總血清膽固醇之作用 ( Osumi et al., 1998 )。
8.2 非消化性碳水化合物之生理活性
海藻經由洋菜酶水解後之海藻多醣,屬於水溶性膳食纖維,因為人 類並無法消化利用,所以可以加速腸道蠕動、增加糞便體積及排便頻率 以及降低血膽固醇 ( Vahouny , 1987;Roberfroid , 1993 )。
九、洋菜酶的應用 9.1 細胞的分離
在組織培養當中,常用洋菜或中性洋菜膠來培養細胞,但是由於培 養在中性洋菜膠上的細胞,不易用無菌吸管將活細胞與中性洋菜膠分 離,所以需要活細胞來進行下一次的培養,並且不會影響到細胞表面的 結構,是非常重要的。Carlsson and Malmqvist (1977) 發展出一種有效的 方法,即自中性洋菜膠上分離活細胞,利用經過適當純化及濃縮後的微 生物洋菜酶,加入正在培養神經膠質瘤細胞 ( glioma cells )的中性洋菜 膠中,純化後的洋菜酶僅會分解中性洋菜膠,故神經膠質瘤細胞即可被 完整的分離出來。利用掃描電子顯微鏡 ( scanning electron microscopy, SEM )來觀察二種不同處理組之表面結構,發現經過洋菜酶的處理後與 未經洋菜酶處理者之兩者的表面並無明顯的差異存在。Solursh (1991) 時亦曾利用洋菜酶來分離培養在膠體上的軟骨細胞( chondrocytes )。
9.2 DNA 或 RNA 分離
利用洋菜酶來回收洋菜糖膠體上的 DNA 或 RNA 片段(Potier et al., 1992;Gnirke et al., 1993;Osoegawa et al., 1998 )。
9.3 藻類原生質體的製作
高等植物原生質體的生成與被利用在生理學與細胞學的研究工作 上,已經被發展許久了,特別是在穀類基因操作實驗中,但是藻類原生 質體的製作則是一個新的領域且遠落後於高等植物的相關研究,故在海 藻的細胞融合與基因操作應用上,分離藻類原生質體的製作方法之發展 是很重要的( Araki et al., 1998a)。Chen et al. 在1994年時,利用產自
Pseudomonas atlantica 的 β-agarase 與其他的酵素如纖維素酶
( Cellulase ) R-10、Macerozyme 與 Abalone enzyme 等,對中性洋菜、
纖維素( cellulose )、木聚醣 (xylan)、紫菜多醣及藻類細胞壁間質等物質 進行分解實驗。發現 Pseudomonas atlantica 所產β-agarase 對紫菜多醣 與中性洋菜遠較實驗中其他酵素有較高的酵素活性,而經由
Pseudomonas atlantica 所產 β-agarase 處理後所產生的藻類原生質體,
與其他酵素處理過後的藻類原生質體相較之下呈現較好的存活率。亦有 利用洋菜酶與不同的藻類,如 Gracilaria ( Araki et al.,1998a )、
Grateloupia ( Chen and Chiang, 1994 )等藻屬作用,產生原生質體以利後 續基因操作。
9.4 融合瘤細胞分離
Gray et al. (1995) 指出由於單株抗體的產生與製造是非常依賴專一 性,並且在分離一株具有價值的抗體時,是不能存在一些不適當的細 胞,而流式細胞儀( flow cytometers ) 可用來偵測適當的細胞,但是僅能 在其與微粒( microdroplets )結合時才能偵測,而且在融合瘤活細胞所分 泌的產物會快速分解,故並無較好的方法偵測融合瘤活細胞所分泌的產 物。所以 Gray et al. 在1995年時利用一種技巧,能夠在微環境下捕捉分 泌的產物,進一步鑑定與分類單一的融合瘤活細胞,即是使用 secretion capture andreporter web (SCRW) 結合來自活的融合瘤細胞所分泌之免 疫球蛋白,接著可並偵測出,藉由中性洋菜微粒與融合瘤細胞形成微膠 囊,如此可以引起螢光抗體特異性三明治分析方法。所以使用 flow cytometers 來鑑定及分類融合瘤細胞時,最後可使用洋菜酶分解中性洋 菜微粒以獲得所需融合瘤細胞並供培養用。
9.5 開發海藻寡糖之健康食品
研究指出以洋菜酶水解海藻所得之水解產物,發現具有新洋菜四 糖、sulfated oligosaccharide、sulfated tetrasaccharide、sulfated
disaccharide 等部分,經餵食老鼠後發現都具有降低總血清膽固醇之作 用 ( Osumi et al., 1998 )。且海藻經由洋菜酶水解後之海藻多醣,屬於 水溶性膳食纖維,因為人類並無法消化利用,所以可以加速腸道蠕動、
增加糞便體積及排便頻率以及降低血膽固醇 ( Vahouny , 1987;
Roberfroid , 1993 )。以不同之洋菜酶水解海藻,可能可以獲得對人體有 益的海藻寡糖,有機會開發成健康食品。
参、實驗設計
Sephadex G-100 膠體管柱過濾純化測 酵素活性,收集具酵素活性劃分部
份並濃縮。
粗酵素液萃取 (在 4℃ 以 10000 rpm 離心30 min 取上清液),20~80 %硫酸銨
劃分,透析過夜 16S rDNA 生化試驗
序列比對 Alcaligenes sp. Strain Yen
於含 700 mL C-10 培養基接種 1%
已活化之菌液,於 RT、150 rpm 振 盪培養 48 hr。
生化性質測試 1. 分子量的鑑定 2. 基質特異性 3. 最適作用 pH 值 4. pH 值的安定性 5. 最適作用溫度 6. 熱安定性
7. 金屬離子的影響
1.活性染色 2.酵素動力學 3.分解產物之探討
肆、實驗材料與方法
一、藥品與耗材
Bradford reagent、Protein marker (Bio-Rad Laboratories) SDS-PAGE(Pierce)
Millipore Amicon Ultra-30(millipore) Agarose (Bio-Rad Laboratories, Sigma) DifcoTM agar,Granulated (BD,Difco) Agar A(Bio basic INC.)
Agar power (Osaka)
Standard methods agar (PCA-D,Biomerieux) Coomassie brilliant blue R-250 , CBR (Sigma) Bromphenol blue sultone, BPB (Sigma)
Calcium chloride, CaCl2 (Sigma)
3,5-Dinitrosalicylic acid (DNSA) (Sigma) 1.3-Dihydroxynaphthalene(Sigma)
D-galactose (Sigma)
Seaniless Cellulose Tubing(Wako,Small size 24)
二、儀器與設備
-80℃凍藏櫃 (freezer) (Leguci, Revco,U.S.A)
0.2 µm syringe filters (25 mm; Pall Corp., Ann Arrbor, MI, U.S.A.;Pall) 分光光度計 (spectrophotometer) (Heλios-β, Lnicam, Tokyo,Japan;
Shimadzu)
恆溫水浴槽 (general purpose bath) (Firstek,Model-B403L)
恆溫振盪培養箱 (orbital shaking incubator) (S300R, Firstek) 低溫高速離心機 (high-speed refrigerated centrifuge) (Himac CR 229,Hitachi, Tokyo, Japan; Hitachi)
蠕動泵 (peristaltic pump)
凝膠 (gel) (Sephadex G-100, Bio-Rad)
電泳槽 (electrophpretic unit) (Bio-Rad Laboratories) 電源供應器 (power supply) (Bio-Rad Laboratories)
三、菌種鑑定(楊,2007) 3.1 菌種生化鑑定
使用台東署立醫院檢驗科 MicroScan-AS4 微生物自動分析儀器,
分析菌種。
3.2 菌種 16S rDNA 分析(楊,2007)
委託新竹食品工業研究所抽取菌株 Yen 之 DNA,進行 PCR 放大 菌株 Yen 之 16S rDNA並進行定序,定序序列結果與美國國家生物科 技資訊中心( NCBI ) 資料庫進行比對,找出序列最接近的已知菌種。
四、培養基配製與菌株之保存 4.1 培養基配製
4.1.1 菌株 Yen 液體生長培養基 ( C-10 broth ) pH 7.5
HEPES(N-2-hydroxyethyl- piperazine-N-2-ethanesulfonic acid) 1.2 g/L
MgSO4.7H2O 0.25 g/L
K2HPO4 0.05 g/L
Ca(NO3)2.4H2O 0.025 g/L KNO3 1.0 g/L Na2-EDTA (Na2-ethylenediaminetetraacetic acid) 0.01 g/L Fe2(SO4)3.6H2O 0.004 g/L
agar (Dfico) 2 g/L
4.1.2 菌株 S3PY 固體生長培養基
添加 1.5% 的 agar 於 C-10 broth 並調整其 pH 至 7.5,於 121
℃ 中滅菌 15 分鐘。
4.2 菌株保存
採用本實驗室分離之能產洋菜酶的菌株 Alcaligenes sp. Strain Yen,菌株 Yen 於固態洋菜培養基上菌落周圍有液化和軟化現象。
將 Alcaligenes sp. strain Yen 於含 0.2% agar 之100 mL C-10 broth,在室 溫下以 150 rpm 振盪培養,當培養菌液之OD 600 nm 達1.0-1.2時,分 別適當培養菌液注入含已滅菌甘油( glycerol ) 保存管中,將菌液與甘油 1:7混合均勻後,於 4 ℃ 預冷2小時後,再置於-20 ℃進行預冷2小時,
最後置於 -80 ℃ 再進行保存。
五、菌株之活化
由-80 ℃冰箱取出保存菌種兩管,以37℃回溫,使用100 mL C-10 broth 在室溫下、150 rpm 振盪培養,培養48小時後,再取2 mL 菌液接 種至新的 C-10 培養24小時,做為種菌。
六、實驗方法 6.1 洋菜酶之生產
於 500mL 之液體培養基,好氧培養菌株 Yen (室溫,150 rpm ),
培養48小時。菌液離心( 4 ℃,10000 rpm,30 min )收集上清液。上清液 以 20~80 % 硫酸銨沉澱,沉澱之蛋白質再溶於50 mM Tris-HCl buffer ( pH 7.0 )中。在以 50 mM Tris-HCl buffer ( pH 7.0 )透析24小時後,使用 0.45 µm syringe filters 過濾不融蛋白質,所得之溶液即為 Yen 粗洋菜酶 溶液。
6.2 洋菜酶的活性測定
6.2.1 依還原糖酵素活性測定酵素活性( Miller,1959 )
洋菜酶反應基質組成為分別取 0.9 mL 含有0.2 % agarose Sodium phosphate buffer ( pH 7.5 ),然後置於35℃水浴槽中 5 min,加入稀釋適 當量的 100 µL 粗酵素液振盪混合均勻,立即放入 35℃水浴槽反應30 min,取出立即加入 1.0 mL 的 DNS 溶液 (秤取 10 g 的
3,5-dinitrosalicylic acid、16 g NaOH 及 300 g K-Na tarutrate 加熱溶解 後,定體積至 1.0 L,儲於不透光的塑膠瓶),接著在 100 ℃ 沸水浴中 靜置 5 min 終止酵素反應,取出冷卻至室溫後,使用分光光度計於546 nm 波長下測定吸光值。以不同濃度的 D-galactose 進行 DNS 呈色反 應,以建立 D-galactose 之標準曲線,並將酵素呈色與 D-galactose 標 準曲線比較,以量化試驗酵素之活性。
6.2.2 活性單位定義( Leon et al.,1992 )
活性單位:每一單位( 1 unit )的酵素活性為:每分鐘能產生相當
1 µmole 半乳糖的酵素量 1 unit = 1 µmole galactose equivalent/min
6.2.3 蛋白質的定量( Bradford, 1976 ) (適用範圍為 0.05-50 µg / mL) 取適當量之樣品,以二次去離子水定容至 200 µL,再加入1000 µL 的 Bio-Rad 染劑( dye reagent ),混合均勻反應 5 min,1 hr內測量波長 595 nm 的吸光值,並以牛血清蛋白( bovine seriumalbumin, BSA ) 做為 標準蛋白質,求得標準曲線,經由標準曲線內插換算出樣品之蛋白質量。
6.3 洋菜酶之分離純化
6.3.1 膠體過濾層析( Sephadex G-100 )
修改楊( 2007 )的膠體過濾層析方法,裝填 1..0 x 60 cm 管柱,以 500 mL 緩衝溶液 Sodium phosphate buffer ( pH 7.5 )於4oC 冰箱流洗過 夜後。接著以 Sodium phosphate buffer ( pH 7.5 )流洗,流速每分鐘 0.2 mL,每劃分收1.5 mL,收集酵素活性劃分部分,以具有分子量 30 kDa MWCO 之超過濾濃縮離心管進行濃縮及透析。
6.4 洋菜酶之生化性質測試(蘇,2005) 6.4.1 分子量的鑑定
參考楊( 2007 )的方法,本實驗採用4~20 % SDS 聚丙烯胺膠體電泳 ( SDS-polyacrylamide gel electrophoresis ) 之方法分析。利用 bio-rad SDS-PAGE 標準品進行分子量估算。
6.4.2 SDS-PAGE 的材料製備
5 倍電泳緩衝液( 5 X Running buffer )(稀釋時加入 0.1 % SDS) Tris base 54.5 g/L
EDTA·2Na 144 g/L Boric acid 6 g/L
加水 800 mL 溶解,以 NaOH 調 pH 至 8.4 後,加水至 1000 mL,
室溫保存。使用前要以蒸餾水稀釋五倍。
2 倍樣品緩衝液( 2 X Sample buffer ) ( SDS reducing buffer ) 0.5 M Tris-HCl ( pH 6.8 ) 1.0 mL
Glycerol 2.0 mL 10 % SDS 4.0 mL 2-β-mercatoethanol 0.4 mL 0.1 % ( w / v ) Bromophenol blue 1.0 mL Distilled water 1.6 mL
儲存於4 ℃,使用時與蛋白質樣本 1:1混合使用
CBR 染液
將400 mL 的 methanol 加入400 mL 的水,再加入 100 mL acetic acid 混合均勻後,加入 10 g Coomassie blue R250,補水至體積 1L。
退染液
將 dH2O、ethyl alcohol、acetic acid 以 7:2:1 比例配置。
6.4.3 SDS-PAGE 分析操作步驟
將洋菜酶與 sample buffer 以1:1 混合後,於 100℃加熱 5 min,
冷卻後取 30 µL 注入樣品槽。以 Tris-Boric acid SDS buffer 做為
running buffer。先以 60 volt 電壓預泳 30 min,再以120 volt 進行電泳 90 min。以 Coomassie btilliant blue R-250 染劑染色1天,再以褪染液褪 染 4 hr 並與蛋白質標準品(注入量為10 µL )比對,以求得其分子量。
6.5 Native PAGE 分析
Native-PAGE 大致與 SDS-PAGE 相同但樣本不經由 SDS 處理,
且膠體中也不加入 SDS,除此之外處理蛋白質樣本通常加入
ß-mercatoethanol 處理,但是 Native-PAGE 中不加入此藥劑,也不加熱 處理。
6.6 洋菜酶活性染色分析
樣本使用不加 ß-mercaptoethanol 與不加熱處理,將跑完SDS-PAGE 或 Native PAGE,使用 Tris-HCl buffer (pH 7.0) 清洗5次,放置平台式 振盪器,轉速30 rpm,每次30分鐘。之後將 PAGE 取出倒上 Soft agar ( 0.5% agarose in Tris-HCl buffer ),放置35℃,培養3小時,取出使用 Iodine reagent (0.05 M I2 in 0.12 M KI) 染色,約10分鐘後觀察結果。
七、基質特異性( Allouch et al., 2004 )
分別將市售實驗級之 high-melting-point agarose、low-melting-point agarose、agar(Agar power,Difco,PCH-D,Agar A )、sodium alginate、ι- 鹿角藻和κ-鹿角藻等物質,比較各洋菜酶 0.1 mL(10 U/mL) 對不同多 醣的基質特異性 0.9 mL,依還原糖酵素活性測定之方法測酵素活性。
八、最適作用 pH 值( 修飾 楊,2007 )
測定各酵素之最適作用 pH,所使用的緩衝溶液如下表。測定方法,
分別取 0.1 mL(10 U/mL) 之酵素加入含不同 pH 値緩衝溶液之基質中 0.9 mL,接下來依據酵素活性測定方法測定活性。
Buffer (50 mM) pH Value Citrate-Na2HPO4 buffer 3.0 4.0 5.0 Tris-HCl buffer 6.0 6.5 7.0 7.5 Sodium phosphate buffer 7.0 7.5 8.5 Phosphate buffer 8.0 8.5 Glycine-NaOH 9.0 10.0
九、pH 值的安定性 (修飾 楊,2007 )
分別取 0.1 mL(10 U/mL) 之各酵素與 50 mM 不同 pH 值(3.0、
4.0、5.0、6.0、7.0、7.5、8.0、8.5、9.0、10.0) 0.9 mL 的緩衝溶液混合 均勻,於 4℃ 下儲存 24 hr 後,取各 pH 值的酵素液,依還原糖酵素 活性測定之方法測酵素活性。
十、最適作用溫度( Suzuki et al., 2003 )
將 0.9 mL 0.2% 的 low-melting-point agarose 及 0.2%
high-melting-point agarose,分別置於各反應溫度 20℃、25℃、30℃、
35℃、40℃、45℃、50℃、55℃、60℃中預熱 5 min,使其與該溫度平 衡後,再加入 0.1 mL(10 U/mL) 之各酵素混合均勻,依還原糖酵素活性 測定之方法測酵素活性。
十一、溫度安定性( 蘇,2005 )
測 Yen 粗洋菜酶在不同溫度下處理 30分鐘之酵素殘存活性,了解 洋菜酶受溫度影響之穩定性。
把 Yen 之粗洋菜酶濃度調整成約 10 U/mL 。粗洋菜酶分別在溫度 20、25、30、35、40、45、50、60 下處理 30 分鐘,然後依還原糖酵 素活性測定之方法測酵素殘存活性。
十二、金屬離子的影響
12.1 鈣離子的影響( Li et al., 2003 )
不同鈣離子濃度 0、0.05、0.1、0.2、0.8、1.0、3.0、5.0、10.0、20.0、
100.0 mM 分別加入基質中混合均勻後,加入各酵素 10 U/mL,依活性 測定法測活性。
12.2、鈉離子的影響( Li et al., 2003 )
不同鈉離子濃度 0、20、50、100、200、400、600、800、1,000 mM 分別加入基質中混合均勻後,加入各酵素10 U/mL,依活性測定法測活 性。
12.3 其他金屬離子耐受度的影響(陳,2006)
金屬離子濃度以其在海水中的濃度為基準 (陳,2006),再比較其濃 度降低到 1/10 倍和提高到 10 倍的差異,各離子的基準濃度分別如附 錄六,將此不同濃度之金屬離子分別加入 0.2% 基質中 0.9 mL,依活 性測定其相對活性。由於以上金屬離子濃度差異大,故以 1 mM 的濃
度下做為比較各金屬離子對洋菜酶的影響,加入各酵素10 U/mL,依活 性測定法測活性。
十三、酵素動力學( Michaelis-Menten )(Sugano et al., 1994 )
取洋菜酶 0.1 mL(10 U/mL)分別在 0.9 mL 0.2% low-melting-point agarose 及 high-melting-point agarose (基質)濃度為 0.5、1、1.5、2、2.5、
3.0 g/L 下反應 30分鐘,然後依吸光度法測定基質濃度與酵素活性的關 係,並利用酵素反應速率與基質濃度做成雙倒數圖形,然後由
Michaelis-Menten 方程式求 Km 及 Vmax。
十四、水解產物分析( TLC )
製備的 0.5 mL 0.2 % agarose 與 0.5 mL 10 U/mL 粗洋菜酶或部分 純化的洋菜酶反應,分別 30 min、60 min、5 hr、24 hr 測產生還原糖量。
14.1、TLC 分析水解產物( Kirimura et. al., 1999 ) 材料:
plate 為 silica gel 60
展開劑為 n-butanol-acetic acid- water ( 2: 1: 1 ) 呈色劑為 naphthoresorcinol
( 0.2g naphthoresorcinol+ 100 mL ethanol+10 mL H3PO4 ) 方法:
將 silica gel 60 裁切成要使用的大小,用鉛筆畫上起始線,點出樣 本的位置並在上方劃上終止線,用 TLC 管點上樣本,再進行展開,終 止後拿起風乾,使用呈色劑呈色,在拿至110 ℃烘箱烘乾10分鐘。
五、結果與討論
一、 Yen 菌種鑑定
本實驗室於台東市區水溝中,篩選到一株洋菜分解菌。其在固體培 養基上,培養一天會產生針孔狀凹陷菌落,隨著培養時間增加,菌落的 凹陷也越來越明顯 ( 圖一 ),滴上碘液染色菌落週圍可見清楚透明圈 ( 圖二 ) 。
經傳統生理生化測試發現 此洋菜分解菌為革蘭氏陰性桿菌,不發 酵、具運動性、好氧、catalase、oxidase 為正反應且不產硫化氫( H2S ) 和 Indole。初步鑑定此洋菜分解菌與 Alcaligenes sp. 有97.4% 相似性 ( 表一 )。
以廣效性引子 ,進行 PCR 放大菌株 Yen 之 16S rDNA 序列,
PCR 產物委託新竹食品工業研究所定序,定序序列結果為圖四,與美 國國家生物科技資訊中心( National Center for Biotechnology
Information,NCBI )資料庫進行 16S rDNA 序列比對,結果發現跟 Alcaligenes sp.ON5 有 100 % 的相似度,NCBI 建議將此菌株命名為 Alcaligenes sp. Strain Yen( 圖三 )。
二、 酵素的純化 1. 粗酵素液的製備
將儲存在 -80 ℃ 的菌株( Alcaligenes sp. Strain Yen )於含100 mL C-10 培養基,接種 0.5 % 已活化菌液,接著在室溫下以 150 rpm 的條 件振盪培養 48 hr 後收集發酵菌液,並以 4 ℃ 下 10000 rpm 條件離心
30 min 後,收集上層液,在經 20~80% 硫酸銨鹽沉澱後並以適量 50 mM Tris-HCl buffer ( pH 7.0 )回溶沉澱物在以 50 mM Tris-HCl buffer ( pH 7.0 )透析24小時後,使用 0.45 µm syringe filters 過濾不溶蛋白質稱 之為粗酵素液( Yen-agarases ),其倍率純化表如表二所示,其所得體積 為 600 mL,總蛋白量為 19.20 mg,總 agarase 活性為 1377.12 U/mL,
定為回收率 100%,比活性為 71.73 U/mg,定為純化倍數 1。
2. Sephadex G-100 膠體過濾層析
將粗酵素液濃縮後,進行 Sephadex G-100 膠體過濾層析,以緩衝 溶液流洗,流速每分鐘 0.1 mL,每劃分收 0.2 mL,收集有酵素活性之 管子。其所得到總體積為 5 mL,總蛋白量為 0.27 mg,總 agarase 活 性為 109.74 U/mL,比活性為 406.44 U /mg,純化倍數為 5.66 倍,回 收率為 7.96 % (如表二)。
陳 (2006) 利用濃縮、DE-52 離子樹脂交換層析 Aeromonas
salmonicida MAEF108,其得到之 agarase 比活性為 1069.46 unit/mg、
純化倍率為 1.11 倍、及回收率為 13.32 %。Suzuki et al.(2003)利用硫銨 沉澱法、DEAE-Toyopearl 650S 離子樹脂交換層析( 0.13-0.40 M
NaCl )、Sephadex G-150 及 Superdex 200 膠體過濾層析純化 Bacillus sp.
MK03,得到之 β-agarase 其總蛋白量為 0.069 mg、總活性為 0.98 unit、
比活性為 14.2 unit/mg、純化倍率為 129 倍、及回收率為 7.0 %。由於 本實驗在純化過程以及粗萃取液較少,造成活性及蛋白量不夠純和不夠 多,因此酵素活性和純化倍率與回收率與文獻相比均較差。以下就經粗 萃取液之 Yen-agarases 探討其生化特性及作用產物之探討。
三、洋菜酶的生化特性 1. 分子量
將 Yen-agarase 4~20 % SDS-PAGE 電泳後經 CBR 染色、及脫色 以及活性染色,與標準蛋白質組比對,結果如圖四所示,其分子量初步 判定約 101 kDa。Native-PAGE 活性染可以知道酵素液中的酵素組成,
由圖五可以看出原態分子量也是約 101 kDa,也可以看出硫銨劃分取 20~80 % 最適當。使用還原態條件的 SDS-PAGE 來決定分子量 (Vera et al., 1998) 用已知分子量大小的蛋白質進行膠體過濾法,計算未知蛋白 質的分子量大小,但是使用膠體過濾法來計算蛋白質的分子量大小有時 會誤判,因為洋菜酶可能會與膠體相互作用,使洋菜酶延遲洗出而造成 分子量的誤判 (Aoki et al.,1990b;陳,2006)。
Vera et al. (1998) 根據文獻中洋菜酶分子量大小,共分成三 類,第一類為分子量在 50 kDa 以下,第二類則為分子量介於 50 與 100 kDa 之間,第三類則為分子量大於 100 kDa 的洋菜酶。
Vibrio sp. JT0107分子量107 KDa ( Sugano et al., 1993a );Vibrio sp.
PO-303 enzyme B 分子量115 KDa ( Araki et al., 1998 );Pseudomonas sp.
enzyme I 分子量 210 KDa ( Malmqvist,1978 );Alteromonas agarlyticus GJ1B 分子量 360 KDa ( Potin et al.,1993 )。均和本實驗所得之洋菜酶屬 同類別。
2. 基質特異性
分別以不同的基質測定 Yen-agarases 的基質特異性,其活性結果 如表二所示,當基質為 high-melting-point agarose、low-melting-point agarose、aga(Difco、Agar power、Agar A、PCH-D agar)、藻酸鈉時其分
別可產生相當於 51.45、42.60、30.72、24.84、32.68、24.78 及 9.28 μmole D-galactose ,而當基質為 ι-carrageenan 和 κ-carrageenan 時均未檢 出,顯示 Yen-agarases 不能分解或是分解這些物質的能力低到未能檢 出。
Vibiro sp. JT0107 的 β-agarase 0107、β-agarase 0072 無法水解 κ-、
ι-、λ-鹿角藻、藻酸鈉( sodium alginate ) ( Sugano et al.,1993a );
Alteromonas sp. E-1 的 β-agarase 無法水解 κ-鹿角藻、藻酸鈉 (Kirimura et al., 1999)。
由文獻發現目前已知洋菜酶都無法水解 κ-鹿角藻、ι-鹿角藻和藻酸 鈉,但可水解 agar、high-melting-point agarose,本實驗與所得之結果略 為不同,由於目前文獻所探討的洋菜酶都無法水解藻酸鈉,但是
Yen-agarase 卻有此作用,可以近一步探討他的作用機制以及水解產物。
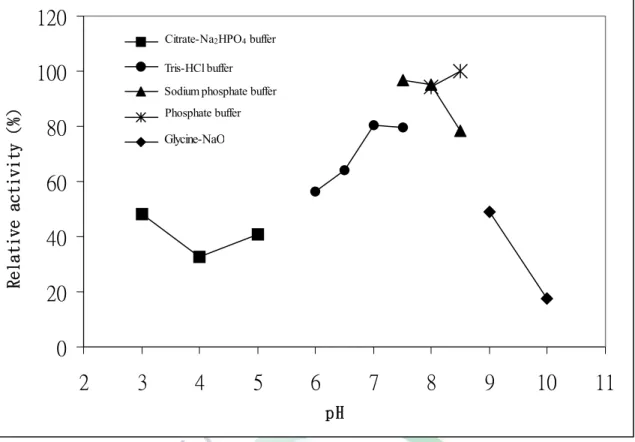
3. 最適作用 pH 值
分別測定各洋菜酶在不同 pH 值的 50 mM 緩衝液之反應活性大 小(citrate-Na2HPO4 buffer pH 3.0-5.0;Tris-HCl buffer pH 6.0-7.5;Sodium phosphate buffer 7.5-8.5;phosphate buffer pH 8.0-8.5;glycine-NaOH pH 9.0-10.0 ),結果如圖六所示,Yen-agarases 其最適作用條件分別為 Sodium phosphate buffer 7.5-8.5 或 phosphate buffer pH 8.0-8.5。由於各 緩衝溶液本身對於洋菜酶會有不同之影響,因此造成最適作用 pH 值未 呈現連續值,因此造成活性降低的情況,例如 Sodium phosphate buffer 在 pH 8.5時呈現下降趨勢。
大部分的洋菜酶之最適 pH 值為 pH 6.5~7.5 之間。
Alteromonas agarlyticus GJ1B 的 α-agarase 其最適 pH 值為 7.2 (Potin
et al., 1993);Pseudomonas sp. W7 的β-agarase 其最適 pH 值為 7.8 (Ha et al., 1997);Alteromonas sp. E-1 的 β-agarase 其最適 pH 值為 7.5 (Kirimura et al., 1999);Bacillus cereus ASK202 的 β-agarase 其最適 pH 值為 7.0 (Kim et al., 1999);Bacillus sp. MK03 的 β-agarase 其最適 pH 值為7.6 (Suzuki et al.,2003) 等,本研究結果顯示 Yen-agarase 在略呈鹼 性環境下有最適 pH 值,與目前文獻比較最適 pH 值呈現偏鹼情形,
所以推論此酵素再偏弱鹼狀態下可存活,可以做進一步的探討。
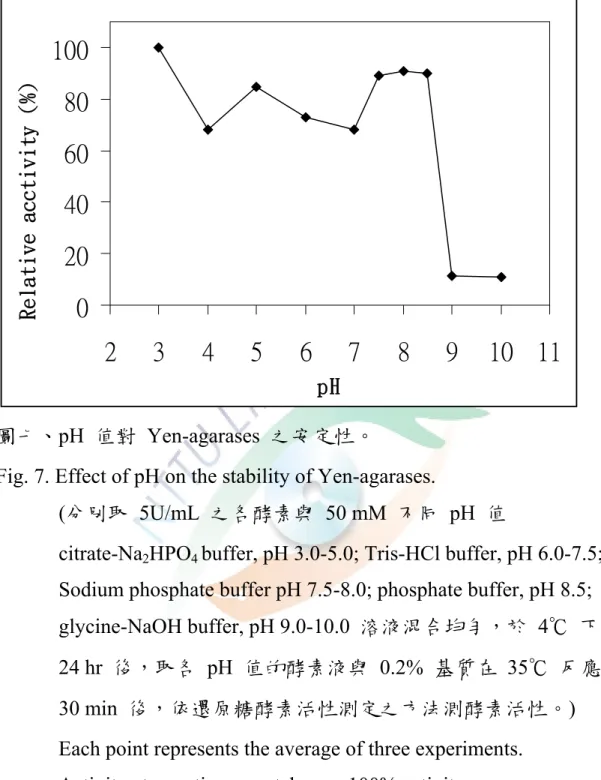
4. pH 值的安定性
分別取一定量之酵素液與 50 mM 不同 pH 值( 3.0-10.0 )的緩衝溶 液,分別在 Ependoff tube 中混合均勻,於 4 ℃ 下儲存 24 hr 後,取各 pH 值的酵素液測量其活性,比較殘存活性之大小,其结果如圖七所示。
Yen-agarase 在 pH 3.0 下殘存活性最高,於 pH 7.5-8.5,都還具有80 % 以上之相對活性,當 pH 值大於 9.0 以上活性幾乎喪失只剩下約20 %。
在 pH 大於3時,活性有下降情形,檢討原因可能 Yen-agarase 純化不 足,尚有其他 agarase 存在,以至於最適作用 pH 值不相符合以及pH 值的安定性圖形有不連續的情形出現。
Vibiro sp. AP-2 在 4℃ 儲存 20 hr,其最安定的 pH 值範圍為 4.0-9.0 (Aoki et al., 1990b) ; Vibrio sp.PO-303 的agarase-a、agarase-b、
agarase-c 在 4℃儲存 24 hr 後,最安定的 pH 值範圍分別為 5.0-8.0、
4.0-10 、及 5-9 (Araki et al.,1998b);P. vesicularis MA103 的 agarase I 在 4℃ 儲存 24 hr 後,其 pH 值小於 7 時,活性逐漸喪失,在 pH 值 低於 4.0 時,活性完全喪失。
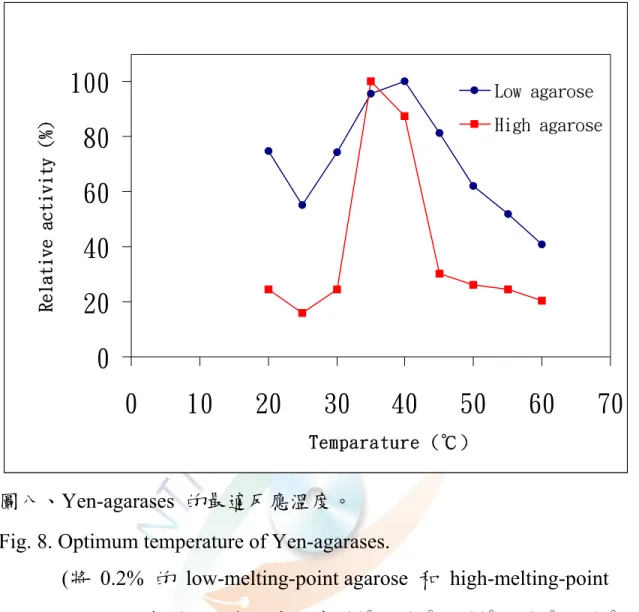
5. 最適作用溫度
將 pH 值為 7.2 之 0.2% 的 low-melting-point agarose 及 0.2%
high-melting-point agarose,分別置於各反應溫度 20℃、25℃、30℃、
37℃、45℃、50℃、55℃、60℃中預熱 5 min,使其與該溫度平衡後,
再加入適量酵素液混合均勻,測量其活性,如圖八所示,Yen-agarases 以 high-melting-point agarose 為基質時之最適作用溫度為 35℃,而以 low-melting-point agarose 為基質時之最適作用溫度為 40℃。
目前所發現的洋菜酶最適作用溫度為 25 ~ 60 ℃ 之間,不同菌株 產生的洋菜酶最適作用溫度不太相同。Vibrio sp. JT0107 最適作用溫度 為30℃ ( Sugano et al., 1993a );Cytophaga flevensis 最適作用溫度為 35
℃ ( van der Meulen and Harder, 1975 ) ; Vibrio sp. PO-303 enzyme C ( Araki et al., 1998 )、Pseudomonas sp. enzyme I 最適作用溫度為 38℃
( Malmqvist, 1978 );Pseudomonas atlantica ( Morrice et al.,1983a )、
Alteromonas sp. E-1 ( Kirimura et al., 1999 ) 、 Bacillus cereus ASK202 ( Kim et al., 1999 )、Vibrio harveyi FLB-17 最適作用溫度為 40℃
( Fukasawa and Kobayashi, 1987 ),其结果均與本實驗结果類似。
6. 溫度安定性
將酵素置於各溫度 20℃、25℃、30℃、35℃、40℃、45℃、50℃、
55℃、60 ℃ 中,分別於 30 min 分鐘後,測定其活性,如圖九所示,
Yen-agarases 於 4 ℃ 下其酵素活性不變,而在 35℃以上作用 30 min 會快速喪失 50% 的活性,檢討溫度安定性圖形與最適作用溫度不相符 合,原因可能 Yen-agarase 純化不足,尚有其他 agarase 存在,以至於
圖形有不連續的情形出現。
Vibiro sp.AP-2 在 45 ℃ 作用 15 min 仍維持原來的活性,在 60℃作用 15 min 會喪失 50% 的活性 (Aoki et al., 1990b);
Alteromonas sp. C-1 的 β-agarase 在 30℃ 以下的溫度作用 1 hr其酵 素活性不變,而在 35 ℃ 作用 20 min 會快速喪失 50% 的活性 (Leon et al., 1992);Vibiro sp. JT0107 的 β-agarase 0107 在40℃ 作用 120 min 的酵素殘存活性為 90%,在 60℃作用 15 min,完全喪失活性 (Sugano et al., 1993a);Vibrio sp. JT0107 的α-NASO hydrolase 在 30 ℃ 作用 120 min 不會喪失活性,超過 35 ℃ 作用會逐漸喪失活性,超過 40 ℃ 作用會快速喪失活性(Sugano et al., 1994);Vibrio sp.PO-303 的 agarase-a、agarase-b、agarase-c 在 60 ℃ 作用 10 min,其相對活性分別 為 25、80、3%(Araki et al., 1998b);Alteromonas sp. E-1 的 β-agarase 在 40℃以下的溫度作用 30 min 酵素比較安定 (Kirimura et al., 1999);P.
vesicularis MA103 agarase I 在 40℃ 作用 10 分鐘會喪失 55%的活 性,超過 50 ℃ 作用會完全喪失的活性 (陳,2006)。由文獻發現大部份 的洋菜酶在高於最適溫度一定時間內會快速喪失酵素活性,本實驗結果 與此相符合。
7. 鈣離子的影響
不同鈣離子濃度 0-100 mM 對 Yen-agarases 活性的影響其结果如 圖十所示,添加鈣離子濃度越高時其活性隨著增加,當鈣離子超過20 mM 時,活性會快速增加(6.10%、6.46%、7.00%、8.61%、9.74%、
18.67% 、33.02% 、34.9% 、94.78 及 100.00%) , 顯示 Yen-agarases 在鈣離子含量較高的情況下活性越佳。由此可知,鈣離子可以刺激
Yen-agarases 活性。
0.002 M CaCl2 會增加 Pseudomonas atlantica 的 β-agarase II 20%
活性 (Morrice et al., 1983a);當缺乏 100 mM NaCl、5 mM CaCl2 的金 屬離子下,會使 Alteromonas agarlyticus GJ1B 的 α-agarase 僅存1/2活 性 (Potin et al., 1993 )。
8. 鈉離子的影響
不同鈉離子濃度 0-1000 mM 對 MAEF108-agarases 活性的影響其 结果如圖十一所示,當添加鈉離子濃度越高時其活性隨著降低,分別為 100.00%、80.00%、93.35%、83.46%、87.21% 、61.02% 、63.83% 、 37.64%、及 31.23%,顯示Yen-agarases 在鈉離子含量越高的情況下活 性越差,且有抑制的現象。
由於大多數產洋菜酶的菌株是海洋性細菌,Ha et al. (1997)
認為添加適量的氯化鈉可以增加洋菜酶活性。Pseudomonas sp. W7 的 β-agarase 在 0.9 M NaCl 下活性會增加,Pseudomonas atlantica 的 β-neoagarotetraose hydrolase、neoagarobiose hydrolase 分別在0.05- 0.20 M、0.10 M 鹽濃度下,洋菜酶活性會增加 (Ha et al., 1997);在 2 mM KCl、NaCl、EDTA(Ethylenediamine tetraacetic acid) 會使 Alteromonas sp.
E-1 的 β-agarase 增加一些活性,CaCl2、MgCl2、MnSO4、CuCl2、FeSO4、 ZnCl2、HgCl2 會抑制其活性,AgNO3 抑制少部份活性 (Kirimura et al., 1999)與本實驗不太符合,原因可能是本菌株不來自於海洋,以致於鹽類 部會提高其活性反而會抑制其活性。
9. 其他金屬離子耐受度的影響
將金屬離子濃度以其在海水中的濃度為基準,再比較其濃度降低到 1/10 倍和提高到 10 倍的差異,將此不同濃度之金屬離子分別加入作用 的基質中,測量其活性的變化以推測其對酵素為加強或是抑制效果,由 於各金屬離子濃度差異大,故以 1 mM 的濃度下做為比較各金屬離子 對洋菜酶的影響,其结果如表四,各離子對 Yen-agarases 都有不同程 度的加強或是抑制效果,以同一濃度相比較,含 1 mM MnCl2 及ZnCl2 有較佳增加活性的效果分別為 127.77% 及 111.01%,而高濃度 CoCl2 會抑制其活性使活性減低為 61.67% 。
添加其他金屬離子和化學藥劑會對微生物所產洋菜酶有不同的影 響,0.005 M MgCl2 會增加 Pseudomonas atlantica 的 β-agarase II 20%
活性 (Morrice et al., 1983a);0.1 mM 的Cu2+、Zn2+、Co2+、Fe2+ 和 Al3+
等離子會完全抑制 Pseudomonas sp. PT-5 agarase活性 (Yamaura et al.,1991);Na+、K+、Mg2+、Fe2+、Sr2+、Zn2+、Cs2+、Co2+、Cu2+、Hg2+ 等 離子對 P. vesicularis MA103 的洋菜 agarase I 的活性具有不同程度的 抑制作用,但 agarase I 需要有5 mM 的 Ca2+ 存在時才具有活性(陳,
2006)。
四、酵素動力學(Km、Vmax)
取 Yen-agarase 分別以 high and low-melting-point agarose 為基 質,與催化速率做成雙倒數圖(Lineweaver-Burk plot ),經回歸,由 Michalis-Menten 方程式分別求得以 high-melting-point agarose 為基質 時,Km 為 2.76 g/L,Vmax 為 68.03 μmole min-1(圖十二);以
low-melting-point agarose 為基質時,Km 為 1.54 g/L,Vmax 為 36.9
μmole min-1(圖十三),顯示 Yen-agarase 對 low-melting-point agarose 較 high-melting-point agarose 的基質親和力高。
五、洋菜酶之分解產物的探討
以薄層層析法 TLC 來分析 Yen- agarases 之水解產物(圖十四)。調 整各洋菜酶酵素活性為 10 U / mL ,與 0.2 % agarose 於35℃ 下,分別 作用 0.5、1、2、5、24 小時之水解產物。洋菜酶處理後之多糖液使 用毛細管點在 silica gel 60 上,然後進行展開,終止後拿起風乾。使用 呈色劑呈色,再拿至 110 ℃ 烘箱烘乾 10 分鐘。
洋菜酶水解洋菜糖之水解產物,皆是隨著作用時間延長,水解產物 逐漸趨向較小的寡糖。從結果中發現,Yen –agarase 粗洋菜酶與 agarose 作用後,產生之水解產物主要為新洋菜二糖、新洋菜四糖和半乳糖,其 他寡醣次之,由於洋菜酶具有基質特異性,因此不同洋菜酶水解洋菜之 水解產物也不完全相同。目前所發現之洋菜酶依其洋菜酶之水解產物來 區分。Arthrobacter sp. S-22 ( Osumi et al., 1998 )、Pseudomonas
atlantica ( Groleau and Yaphe, 1977 )、Cytophaga flevensis ( van der Meulen and Harder, 1976 )、Vibrio sp. AP-2 enzyme ( Aoki et al.,
1990b )、Vibrio sp. AP-2 enzyme B ( Aoki et al., 1990a )、Alteromonas sp. E-1 ( Kirimura et al., 1999 )之菌株洋菜酶水解洋菜主要產物為新洋 菜雙糖。 Cytophaga flevensis ( van der Meulen and Harder, 1975 )、
Vibrio sp. PO-303 enzyme B ( Araki et al., 1998 )、Vibrio sp. JT0107 ( Sugano et al., 1993a )之菌株洋菜酶水解洋菜主要產物為新洋菜四糖 及新洋菜雙糖。 Pseudoalteromonas atlantica N-1 ( Vera et al., 1998 )、
Vibrio sp. PO-303 enzyme A ( Araki et al., 1998 ) 之菌株洋菜酶水解洋菜
主要產物為新洋菜六糖及新洋菜四糖。Alteromonas agarlyticus GJ1B ( Potin et al., 1993 ) 之菌株洋菜酶水解洋菜主要產物為 agarotetraose 及洋菜三糖( agarotriose )。Bacillus cereus ASK202 ( Kim et al., 1999 ) 之菌株洋菜酶水解洋菜主要產物為新洋菜六糖、新洋菜四糖及新洋菜雙 糖。Vibrio sp. JT0107 ( Sugano et al., 1994 ) 之菌株洋菜酶水解洋菜主要 產物為洋菜五糖( agaropentaose )、agarotriose、洋菜雙糖及 D -galactose。
Vibrio sp.PO-303 enzyme C ( Araki et al., 1998 ) 之菌株洋菜酶水解 洋菜主要產物為新洋菜八糖( neoagarooctaose ) 及新洋菜十糖
( neoagarodecaose )。Cytophaga flevensis ( van der Meulen and Harder, 1975 ) 、 Pseudomonas atlantica ( Day and Yaphe, 1975 )之菌株洋菜酶水 解洋菜主要產物為 D –galactose 及 3,6-脫水-L型半乳糖
( 3,6-anhydro-L-galactose )。
由於Potin et al. ( 1993 ) 指出如果洋菜酶為切 α-form,並不會產生 還原端,所以無法以測還原糖的方法測到活性,但如果含有 β-半乳糖苷 酶,會將非還原端切掉,露出還原端使可以偵測到的還原糖量上升。
本研究利用這個特點,在酵素中加入 β-半乳糖苷酶偵測到的還原糖 量,如果有上升表示洋菜酶為切α-form 的洋菜酶,如果沒有上升表示 洋菜酶為切 β-form 的洋菜酶,結果發現可能含有大部份切 β-form 的 洋菜酶。由此可以推測菌株 Yen 可能為 β-agarase。若要更精確知道水 解產物及其分型,則需要利用其他儀器如 NMR、HPLC 分析才能確定 產物為何,並利用產物判斷其為那種類型的洋菜酶作用模式。