魚貝類的化學組成與其死後變化
The Chemical Composition in Fish and Shellfish and Their Post- mortem Changes
邱思魁 Tze-Kuei Chiou
國立臺灣海洋大學生命科學院食品科學系
Received 22 January 2018; revised 7 March 2018; accepted 12 March 2018; available online 20 March 2018
摘 要
本文內容涵蓋兩項主題,第一部分:魚貝類的化學組成,重點包括主要的組成 分、脂質、蛋白質、含氮萃取物成分、維生素和礦物質等五項。第二部分:魚類的死 後變化,重點包括感官的變化、自家消化的變化、細菌的變化、脂質氧化和水解。其 中,自家消化的變化項目又分為僵直後肌肉中能量的產生、自家消化和核苷酸分解作 用、蛋白質分解酵素參與的自家消化的變化(分別討論組織蛋白酶、鈣蛋白酶、膠原蛋 白酶等)、凍藏中自家消化的變化等重點。細菌的變化項目又包含:活魚的菌相、微生 物的侵入、貯藏期間微生物菌群的變化和腐敗/特定的腐敗微生物、貯藏與腐敗過程中 細菌生長所誘發的生化變化、氧化三甲胺的還原等重點。
關鍵字:魚貝類、化學組成、死後變化、自家消化、腐敗微生物。
一、前言
水產物資源在提供人類糧食的來源 上,日漸重要。從水產利用的角度來觀 看魚貝類的特性,可歸納出:(1)漁獲的 不 安 定性 、( 2) 魚種 及成分組 成的多樣 性、(3)生理活性物質的存在、(4)血合肉
*通訊作者電子信箱:[email protected]
的存在、(5)容易腐敗變質、(6)有毒種類 的出現、(7)造成或影響加工過程出問題 之萃取物成分等(吳, 1998)。和陸上畜產 品比較,水產品的種類多,棲息的水生 環境又含括淡水、海水甚至半鹹水等,
本質上環境的組成就有所不同之外,環 境因素例如鹽度、水溫等的變化,對於 生長棲息其中的水生生物也帶來莫大的 影響。
在海洋水產院校的(水產)食品相關科 系,「水產化學」一直是重要的必修科 目,透過認識這海洋水產特色的專業,
始能十足地認識水產品的真面目,進而 充 分 掌 握 水 產 物 資 源 的 研 發 與 有 效 利 用。在國內,有關的教科書或專書主要 有吳等(1991)編著的「水產化學上下冊」
(職校用)、吳等(1992)編著的「水產化學 上下冊」(專校用)、吳與邱(1996)譯著的
「水產食品學」、吳(1998)譯的「水產利 用化學」等,這些書都是由國立編譯館 出版,後來職專校專業教科用書都委由 華香園出版社出版之外,另兩本專書已 缺版無貨。聯合國糧農組織(FAO)出版的
「 Quality Changes in Fresh Fish 」 報 告 (Huss, 1995),以介紹生鮮魚類的品質變 化為主題,其內容豐富,討論的範圍層 面廣且敘述又有深度,個人深感值得參 考。由於篇幅,僅摘取和水產化學的最 基本問題部分,加以編寫,最後以本文 作完稿。
二、魚貝類的化學組成 (一) 主要組成分
魚的化學組成(chemical composition) 在種類間或個體間,會因年齡、雄雌、
環境和季節而變動很大。魚和哺乳動物 的主要組成分可分成相同的類別,以及 舉 例說明魚 組成分 的 變 動, 如表 1 所 示,其中亦與牛肉的組成一起比較。從 表中的數據,可看到魚肉的組成分有相 當大的正常變動範圍,所列的最大值與 最小值屬極端而非常態。
魚化學組成的變動與餌料攝取、洄游 和產卵所牽連的性轉變等密切關連。因 自然或生理原因(譬如洄游和產卵)、或由 於外界因素譬如食物不足,魚會有飢餓 期間,通常,產卵期不論有無長時間洄 游都需要較多的能量,以脂質型式貯藏 能量的魚就依賴於此,進行長時間洄游 的 種 類 在 到 達 特 定 的 產 卵 場 或 河 川 之 前,除脂質之外也會利用蛋白質提供能 量。因而同時消耗脂質和蛋白質貯藏,
導致魚的生物狀況(biological condition)變 差,又大多數種類在產卵洄游期間通常 不會消化太多的食物,因而透過飼養也 不能夠提供能量。
表 1. 魚肉和牛肉的主要組成分(%)
組成分 魚肉
最低 正常變動值 最高 牛肉
蛋白質 6 16~21 28 20
脂質 0.1 0.2~25 67 3
碳水化合物 <0.5 1
灰分 0.4 1.2~1.5 1.5 1
水分 28 66~81 96 75
資料來源:Stansby, 1962; Love, 1970。
密集進食的期間,首先肌肉組織的蛋 白質含量會提高至一定的程度,這取決 於有多少量被消耗,例如產卵洄游中。
其次,脂質含量會明顯地快速增加,產 卵之後,回復攝食行為,常洄游尋找食 物來源。攝食浮游生物的種類例如鯡魚 (herring)會自然經歷另一次比產卵更加明 顯的季節性變動,因為浮游生物的產量 會受到季節和海洋中的各種物理因素等 的影響。
脂質是變動最大的組成分,通常,有 些魚種的變動會顯現特別的季節變化曲 線,最低值位於產卵時間。圖 1 顯示北 海(North Sea)產鯡魚(a)和鯖魚(b)的特別 變動情形。
雖然大多數魚種其蛋白質部分的含 量 較 穩 定 , 鮭 魚 在 長 時 間 的 產 卵 洄 游 (Ando et al., 1985b; Ando and Hatano, 1986),以及波羅的海鱈魚從 1 月至 6/7 月的產卵季節(Borresen, 1992),都觀察到 蛋白 質 減少 之變 動, 後者 例 如圖 2 所 示。
有些熱帶魚類的化學組成也會出現 明顯的季節性變動。西非鰣魚(Ethmalosa dorsalis)在一整年間的脂質含量 2~7% (濕 重),最大值在 7 月(Watts, 1957)。巴西海 岸捕撈 的黃 花魚(Micropogon furnieri)和 pescada-foguete (Marodon ancylodon)的脂 質含量分別為 0.2~8.7%及 0.1~5.4% (Ito and Watanabe, 1968),同時這些種類的脂 質含量也因魚體的大小而不同,較大型 者比較小型者高約 1%。Watanabe (1971)
也探討尚比亞產淡水魚,發現包括中上 層(pelagic)與底層(demersals)棲息的四種 魚的脂質含量介於 0.1 至 5.0%。
區分寡脂(lean)與多脂(fatty)魚種的一 種可行方法,即是寡脂魚僅將脂質貯藏 於肝臟,但多脂魚將脂質貯藏分布於其 它體組織的脂肪細胞(fat cells)。典型的寡 脂魚是底棲性魚種,如鱈魚(cod)、綠青 鱈(saithe)和無鬚鱈(hake)。多脂魚的種類 包括棲息中上層水域的魚,如鯡(鰊)魚 ( h e r r i n g ) 、 鯖 魚 ( m a c k e r e l ) 和 小 鯡 魚 (sprat),有些種類的脂質只貯藏於體組織
圖 1. (a)鯡魚(Clupea harengus)與(b)鯖魚(Scomber scombrus)肉化學組成的季節變動。每點代表 8 片魚肉的平均值。
圖 2. 白令海產鱈魚肌肉乾物比率的變動。豎線代 表平均值的標準誤差(Borresen, 1992)。
的限定部位,或者比典型的多脂魚含較 低的貯藏量,因而稱半多脂(semi-fatty)魚 種,例如梭魚(barracuda)、鯔魚(mullet)和 鯊魚(shark)。
寡 脂 魚 肉 片 的 脂 質 含 量 低 且 變 動 小,相對的,多脂魚種肉片的脂質含量 變動幅度大。油脂比率的變化也反映於 水分的比率,由於油脂和水分兩者通常 合 計 占 肉 片 的 80% 左 右 , 依 據 經 驗 法 則,從肉片水分量的分析可估計油脂含 量。實際上,「Torry Fish Fat Meter」之 油脂分析儀器即應用此原理,水分含量 才是真正測定的對象(Kent et al., 1992)。
無論寡脂魚或多脂魚,實際的油脂 含量對死後變化(post mortem)都有所影 響。在生鮮(fresh)寡脂魚所發生的變化,
可從了解其蛋白質區分的生化學變化而 預 測 , 在 多 脂 魚 種 則 尚 包 含 脂 質 的 變 化,此用意是可能由於脂質氧化而縮短 貯藏時間,或者須採取特別的預防措施 來防止。
各種魚類的水分、脂質和蛋白質含 量的變動情形,如表 2所示。
魚 類 肌 肉 中 的 碳 水 化 合 物 含 量 很 低 , 通 常 < 0.5% 。 屬 橫 紋 肌 者 大 都 如 此,碳水化合物是以肝醣(glycogen)及作 為核苷酸類的組成的一部分而存在,後 者 來 自 死 後 自 家 消 化 的 變 化 (autolytic changes)所釋出的核糖(ribose)。
如上所述,不同魚種的化學組成因 季節變化、洄游行為、性成熟、餵食週 期等而變動,這些因素在開放海域與內
表 2. 各種魚類肌肉的化學組成
種類 學名 水分
(%)
脂質 (%)
蛋白質 (%)
熱量 (kJ/100 g) 藍鱈 Blue whiting a) Micromesistius poutassou 79~80 1.9~3.0 13.8~15.9 314~388
鱈魚 Cod a) Gadus morhua 78~83 0.1~0.9 15.0~19.0 295~332
鰻魚 Eel a) Anguilla anguilla 60~71 8.0~31.0 14.4
鲱魚 Herring a) Clupea harengus 60~80 0.4~22.0 16.0~19.0
鰈魚 Plaice a) Pleuronectes platessa 81 1.1~3.6 15.7~17.8 332~452
鮭魚 Salmon a) Salmo salar 67~77 0.3~14.0 21.5
鱒魚 Trout a) Salmo trutta 70~79 1.2~10.8 18.8~19.1
鮪魚 Tuna a) Thunnus spp. 71 4.1 25.2 581
挪威龍蝦 Norway lobster a) Nephrops norvegicus 77 0.6~2.0 19.5 369 雷吉牙漢魚 Pejerrey b) Basilichthys bornariensis 80 0.7~3.6 17.3~17.9
鯉魚 Carp b) Cyprinus carpio 81.6 2.1 16.0
條紋鯪脂鯉 Sabalo c) Prochilodus platensis 67.0 4.3 23.4 淡水白鯧 Pacu c) Colossoma macropomum 67.1 18.0 14.1 大蓋具脂鯉 Tambaqui c) Colossoma brachypomum 69.3 15.6 15.8 虎紋鴨嘴鯰 Chincuiña c) Pseudoplatystoma tigrinum 70.8 8.9 15.8 巴西異鱗石首魚 Corvina c) Plagioscion squamosissimus 67.9 5.9 21.7
巴格海鱨 Bagré c) Ageneiosus spp. 79.0 3.7 14.8
資料來源:a) Murray and Burt, 1969;b) Poulter and Nicolaides, 1985a;c) Poulter and Nicolaides, 1985b。
陸水域的野生、自由生存的魚種都可觀 察到。水產養殖魚也會顯現化學組成上 的變動,此情況下,由於有些因素是受 控制的,故可預估其化學組成。在一定 程度上,養殖魚的化學組成透過選擇的 養殖條件就能夠設定,已知例如飼料組 成 、 環 境 、 魚 體 大 小 和 遺 傳 特 徵 等 因 素 , 都 會 影 響 養 殖 魚 類 的 組 成 和 品 質 (Reinitz, et al., 1979)。
影響化學組成之最大單一因素是飼 料組成。養殖漁民希望盡可能以最少量 的餌料使魚生長愈快,因為餌料是養殖 最主要的成本項目。當餵食提供熱量的 高脂質以及含有胺基酸組成均衡的高量 蛋白質之飼料,此時養殖魚的潛在成長 速率最高。
但是,相較於蛋白質,對於有多少 的脂質能被代謝,在魚類的基礎代謝模 式會設定一些限制,因為蛋白質比起脂 質是更貴的飼料成分,很多實驗都探討 盡可能將愈多的蛋白質改以脂質取代,
可 參 考 下 列 的 文 獻 :Watanabe et al.
(1979)、Watanabe (1982)、Wilson and Halver (1986)和Watanabe et al. (1987)。
經常,不論脂質的含量,大多數魚 種都會利用一些蛋白質來作為能源,當 脂質含量超過能被代謝為能源之最大量 時,剩出的部份仍貯藏於組織,這使得 魚含有很高的油脂量。除了對整體品質 有負面的影響,也使產量減少,因多餘 的脂肪都貯藏在腹腔內的脂肪層中,當 去除內臟與剖取魚肉片(filleting)時就被丟
棄。
在收穫前,降低養殖魚的油脂含量 之常態作法是讓魚飢餓一段期間。一些 研究都指出這可影響多脂魚和少脂魚的 脂 質 含 量 (Reinitz, 1983; Johansson and Kiessling, 1991; Lie and Huse, 1992)。
(二) 脂質
硬骨魚的脂質可分為兩大類:磷脂 質 (phospholipids) 和 三 酸 甘 油 酯 (triglycerides)。磷脂質是構成細胞中細胞 膜 的 整 體 結 構 , 通 常 稱 為 結 構 脂 質 (structural lipids)。三酸甘油酯是將能量貯 存於脂肪蓄積處之脂質,通常貯存於磷 脂質膜和相當弱的膠原蛋白網狀結構所 環繞的特定脂肪細胞(fat cells),三酸甘油 酯一般稱為貯藏油脂(depot fat),少數魚 種含蠟酯(wax esters),是貯藏油脂的一部 分。
典型的寡脂魚如鱈魚(cod)所含的脂 質 低 於 1% , 其 中 的 磷 脂 質 占 約 90%
(Ackman, 1980),寡脂魚肌肉中的磷脂質 區分由大約 69%磷脂醯膽鹼(phosphatidyl choline)、19%磷酯醯乙醇胺(phosphatidyl ethanolamine) 和 5% 磷 酯 醯 絲 胺 酸 (phosphatidyl serine)等組成,另還有其它 微量的磷酯質類。
磷脂質存在於膜結構中,包括細胞 外 膜 (outer cell membrane) 、 內 質 網 (endoplasmic reticulum)和細胞內的小 管系 統(intracellular tubule systems),以及粒線體 (mitochondria) 等 胞 器 (organelles) 的 膜 。 除
了磷脂質,膜也含有膽固醇,貢獻膜的 剛性(rigidity)。寡脂魚的肌肉中,膽固醇 含量可達總脂質(total lipids)量的 6%,類 似於哺乳動物肌肉。
前已說明,依照如何為能量而貯存 脂 質 , 魚 的 種 類 可 分 為 寡 脂 魚 或 多 脂 魚 , 寡 脂 魚 利 用 肝 臟 作 為 能 量 貯 存 場 所,多脂魚將脂質貯存於遍佈全身的脂 肪細胞。
多 脂 魚 的 脂 質 貯 存 所 在 的 脂 肪 細 胞 , 通 常 位 於 皮 下 組 織 (subcutaneous tissue)、腹部皮瓣肌(belly flap muscle)和 主導鰭及尾部動作的肌肉,有些含特別 高 量 油 脂 的 魚 種 也 會 貯 存 在 腹 腔 (belly cavity),根據多元不飽和脂肪酸的含量,
多數魚類的油脂在低溫下或多或少都是 液體。
最後,油脂貯存通常都是遍佈於肌 肉結構,脂肪細胞的濃度大抵在靠近肌 隔 (myocommata) 、 以 及 在 普 通 肉 (light/ordinary muscle) 與 血 合 肉 (dark muscle)之間的區域為最高(Kiessling et al., 1991)。即使是寡脂魚,血合肉在其肌肉 細胞內包含一些的三酸甘油酯,蓋因血 合肉能夠直接代謝脂質為能量。普通肉 細 胞 則 依 賴 肝 醣 作 為 嫌 氣 性 代 謝 (anaerobic metabolism)的能量來源。
在血合肉中,能量儲備最後被完全 代謝為二氧化碳和水,相對的在普通肉 則 生 成 乳 酸 。 普 通 肉 中 的 能 量 調 動 (mobilization of energy)比在血合肉中快速 很多,但乳酸的生成易產生疲勞,使肌
肉 無 法 以 最 大 的 速 率 長 時 間 運 作 。 因 此,血合肉是利用於持續性長時間的游 泳活動,而普通肉利用在瞬間爆發性的 能量,譬如要捕獲獵物或者逃避捕食者 的追捕。
鯖魚(mackerel)和柳葉魚(capelin)油脂 蓄積的季節性變動,如圖 3 所示,可看 出不同組織內的脂質含量變動相當大,
脂質儲藏通常是為了長時間產卵洄游和 建構生殖腺時(Ando et al., 1985a)。當脂 質因這些目的而調動時,三酸甘油酯中 不同的脂肪酸是否選擇性地被利用,仍 不清楚,鮭魚明顯並無該情形,但鱈魚 會選擇性利用 DHA (C22:6) (Takama et
al., 1985)。
持續性洄游時,磷脂質也可被動員 一定的程度(Love, 1970),儘管一般認為 磷脂質比起三酸甘油酯,被保留更多。
在鯊魚等軟骨魚類(elasmobranchs),
相當量的脂質貯存於肝臟,包含二醯基- 烷基-甘油酯類(diacyl-alkyl-glyceryl esters)
圖 3. 挪威產鯖魚(上圖)和柳葉魚(下圖)不同部位 中總油脂的分佈(Lohne, 1976)。
或鯊烯(squalene)等油脂,有些鯊魚肝臟 油 至 少 80% 脂 質 是 不 皂 化 物 (unsaponifiable substance),其中大多是鯊 烯 (squalene) (Buranudeen and Richards- Rajadurai, 1986)。
魚類脂質不同於哺乳動物脂質,主 要的不同在於魚類脂質包括多至 40%長 鏈脂肪酸(14~22 個碳原子),且是高度不 飽和化,哺乳動物脂質每一脂肪酸分子 很少含有超過 2 個雙鍵者,而魚的貯藏 油脂含有 5 或 6 個雙鍵的數種脂肪酸 (Stansby and Hall, 1967)。
含 4、5 或 6 個雙鍵的高度不飽和脂 肪酸的比率,在淡水魚脂質(約 70%)略低 於 在 海 水 魚脂 質 ( 約 88%) (Stansby and Hall, 1967),但是,脂質的組成並非完全 固定,仍隨攝食餌料和季節而變動。
在 人 類 , 營 養 素 脂 肪 酸 如 亞 麻 酸 (linoleic acid)和次亞麻酸(linolenic acid)由 於體內無法合成而視為是必需的。在海 水魚,這些脂肪酸只占總脂質的 2%左 右,和許多蔬菜油比較,比率是低的。
但魚油含有其它的高度不飽和脂肪酸,
如同亞麻酸和次亞麻酸,是預防皮膚病 症所必需的。屬於次亞麻酸家族(第一個 雙鍵位在第 3 位置,從末端甲基算起為 ω-3)的成員,對成長中小孩神經系統具有 好處。這些脂肪酸之一的二十碳五烯酸 (EPA; eicosapentaenoic acid; C20:5ω3)最 近受到重視,丹麥科學家發現在一群格 陵蘭愛斯基摩人的飲食中,此種脂肪酸 含量高;英國和其它地區的調查也指出
血液中的 EPA 是非常強力的抗血栓因子 (antithrombotic factor) (Simopoulos et al., 1991)。
(三) 蛋白質
魚類肌肉組織中的蛋白質,可分為下 列三群:
1. 結構蛋白質(structural proteins):肌 動 蛋 白 (actin) 、 肌 凝 蛋 白 (myosin) 、 原 肌 球 蛋 白 (tropormyosin) 和 肌 動 凝 蛋 白 (actomyosin),構成總蛋白質含量 的 70~80%,這些蛋白質可溶解於 離子強度(ionic strength)相當高(0.5 M)的中性鹽溶液。
2. 肌 漿 蛋 白 質 (sarcoplasmic proteins) : 肌 白 蛋 白 (myoalbumin)、球蛋白(globulin)和 酵 素 , 可 溶 解 於 低 離 子 強 度 ( < 0.15 M)的中性鹽溶液,構成總蛋 白質的 25~30%。
3. 結締組織蛋白質(connective tissue proteins):膠原蛋白(collagen),構 成硬骨魚及軟骨魚的蛋白質各約 3%及 10%,在哺乳動物為 14%。
結構蛋白質建構收縮機構(contractile apparatus)以負責肌肉的運動。雖物理性 質有些差異,胺基酸組成大抵類似於哺 乳 動 物 肌 肉 中 對 等 的 蛋 白 質 。 等 電 點 (isoelectric point; pI)在 pH 4.5~5.5,在此 對應的 pH 值範圍內,蛋白質的溶解度最 低,如圖 4所示。
魚 類 蛋 白 質 的 構 形 結 構 (conformational structure)受到物理環境的 改變而易生變化。在圖 4,顯示肌原纖維 蛋白質(myofibrillar proteins)溶解度特性 在凍結乾燥後的改變,高鹽濃度或加熱 的處理會導致變性(denaturation),在此之 後天然蛋白質(native protein)發生不可逆 的改變。
當蛋白質在控制條件下變性,得到 的性質可應用於加工上的目的。很好的 範例為水產煉製品(surimi-based products) 的製造,就是利用肌原纖維蛋白質的膠 體形成能力(gel forming ability),水洗及 擂潰後的肌肉蛋白質加入鹽、穩定劑,
並經一定條件的加熱與冷卻步驟之後,
蛋 白 質 就 形 成 高 彈 性 的 膠 體 (Suzuki, 1981)。
肌漿蛋白質的大部分是參與細胞代 謝的酵素,譬如從肝醣至 ATP 之厭氧性 能量轉換(anaerobic energy conversion)。
如果肌肉細胞內的胞器受損,該蛋白質 區 分 也 會 含 有 原 存 在 於 內 質 網 (e n d o p l a s m a t i c r e t i c u l u m) 、 粒 線 體
圖 4. 在 pH 2-12 範圍下凍結乾燥前後的肌原纖維 蛋白質的溶解性(Spinelli et al.,1972)。
(mitochondria) 和 溶 菌 酶 (lysosomes) 等 內 部 的代謝酵素。
當胞器破損而造成肌漿蛋白質區分 組成的改變,被應用於辨識生鮮和冷凍 魚的方法,前提是冷凍前的胞器都是完 整的(Rehbein et al., 1978; Rehbein, 1979;
Salfi et al., 1985)。但後來的研究指出,
使用這方法要很小心,因魚冰藏期間有 些酵素也會從胞器釋出(Rehbein, 1992)。
肌漿區分中的蛋白質非常適用於辨 識不同的魚種,當採用等電點聚焦電泳 (isoelectric focusing)的分離方法,所有不 同 的 種 類 都 顯 示 特 徵 的 帶 圖 紋 (band pattern),此方法由 Lundstrom (1980)開發 成功,許多實驗室也採用並應用於很多 的魚種,這方面的文獻整理參照 Rehbein (1990)。
膠原蛋白的化學與物理性質,因組 織如皮、浮鰾(swim bladder)和肌肉內的 肌隔(myocommata)而不同,通常,膠原 蛋白纖維形成微妙的網狀構造,不同的 結締組織中其複雜性也不同,如同在哺 乳動物的情形;但是,魚類膠原蛋白的 熱穩定性更差,比起溫血脊椎動物的膠 原蛋白,所含的交聯(cross-links)程度較 少且更不穩定的。雖然魚類膠原蛋白中 的羥脯胺酸(hydroxyproline)含量變動介於 4.7%至 10%之間,但一般而言魚類的羥 脯 胺 酸 含 量 低 於 哺 乳 類 (Sato et al., 1989)。
各種魚類的體組織各含不同量的膠 原蛋白,這導致一項理論即膠原蛋白的
分佈反映出魚種的游泳行為(Yoshinaka et
at, 1988)。又,魚類的不同含量與不同類
型的膠原蛋白也影響魚類肌肉的質地性 質 (Montero and Borderias, 1989) , Borresen (1976)開發一種方法,即分離圍 繞在每一個別肌肉細胞的膠原蛋白網狀 構造;鱈魚中這些的構造的結構和組成 已深入探討(Almaas, 1982)。魚 類 膠 原 蛋 白 的 角 色 已 有 綜 述 (Sikorski et al., 1984),Bremner (1992)發 表更完整的綜述,發現自魚類的各種型 式膠原蛋白的有關近期文獻,大多有列 入。
魚類蛋白質含有全部的必需胺基酸 (essential amino acids),如同牛奶、蛋類 及 哺 乳 動 物 肉 類 蛋 白 質 , 生 物 價 (biological value) 很高 (表 3)。
穀類蛋白質通常其離胺酸(lysine)及/
或含硫胺基酸(甲硫胺酸 methionine 與半 胱胺酸 cysteine)的含量低,相對的,魚類 蛋白質是這些胺基酸的優良來源。穀物 為主食,若再補充魚,可顯著提升生物 價。
除了提及的魚類蛋白質,自副產物 (特別是內臟)回收特定的蛋白質部分,這 問題也受重視。其中一例為雄性魚的鹼 性 蛋 白 質 (basic protein) 或 魚 精 蛋 白 (protamines),分子量大致在 10000 kD 以 下,等電點大於 10,極端的胺基酸組成 例中,精胺酸比率可高達 65%。很早就 知道鹼性蛋白質的存在,且並非所有魚 類中都有(Kossel, 1928),最佳的來源為鮭 科(salmonids)和鯡魚(herring),相對的,
底棲魚類如鱈魚並不含魚精蛋白。
魚精蛋白的極偏鹼性的特性因有些 原因而受注目。可吸附至大多數較低鹼 性的其它蛋白質,因而具有促進其它食 品蛋白質的功能性質之作用(Poole et al., 1987; Phillips et al., 1989),但是,蛋白質 製 備 時 要 除 去 精 巢 中 的 脂 質 , 問 題 仍 大,使用在食品中的濃度,會有不良風 味的問題。鹼性蛋白質另一受注目的特 點為防止微生物生長之能力(Braekkan and Boge, 1964; Kamal et al., 1986),有可能是 這些鹼性蛋白質未來最有希望的應用。
表 3. 各種蛋白質中的必需胺基酸(%)
胺基酸 魚 牛奶 牛肉 蛋
離胺酸 Lysine 8.8 8.1 9.3 6.8
色胺酸 Tryptophan 1.0 1.6 1.1 1.9
組胺酸 Histidine 2.0 2.6 3.8 2.2
苯丙胺酸 Phenylalanine 3.9 5.3 4.5 5.4
白胺酸 Leucine 8.4 10.2 8.2 8.4
異白胺酸 Isoleucine 6.0 7.2 5.2 7.1
蘇胺酸 Threonine 4.6 4.4 4.2 5.5
甲硫胺酸-胱胺酸 Methionine-Cystine 4.0 4.3 2.9 3.3
纈胺酸 Valine 6.0 7.6 5.0 8.1
資料來源:Braekkan, 1976; Moustgard, 1957。
(四) 含氮萃取物成分
含 氮 萃 取 物 成 分 (N-containing extractives) 可定義為 非 蛋白質 本 質 (non- protein nature)的水溶性、低分子量的含氮 化合物,非蛋白態氮(non-protein nitrogen) 區分構成硬骨魚類的 9~18%總氮量(total nitogen)。
非蛋白態氮區分的主要組成分為:
揮 發 性 鹽 基 類 (volatile bases) 譬 如 氨 (ammonia) 及 氧 化 三 甲 胺 (trimethylamine oxide; TMAO)、肌酸(creatine)、游離胺基 酸(free amino acids)、核苷酸與其嘌呤鹽 基類(nucleotides and purine bases)、及在 軟骨魚(cartilaginous fish)的尿素(urea)。
表 4中,列出各種魚類、禽肉與哺 乳動物肉的非蛋白態氮區分中的一些組 成分。
圖 5. 兩種硬骨魚(A: 嘉鱲;B: 日本竹筴魚)、一 種軟骨魚(C: 鯊魚)和一種淡水魚(D: 香魚)肌 肉 中 的 非 蛋 白 態 氮 分 佈 (Konosu and Yamaguchi, 1982; Suyama et al., 1977)。
淡水魚與海水魚的非蛋白態氮區分 中各種化合物的分佈樣式如圖 5 所示。
須注意:組成不僅隨種類的不同,即使 同一種類也因體型大小、季節、肌肉樣 品等而變動。
表 4. 肌肉萃取物成分中的主要差異
化合物 (mg/100 g 濕重) 1 魚類
鱈魚
魚類 鯡魚
魚類 鯊魚類
甲殼類 龍蝦
禽類 腿肉
哺乳類 肌肉
1) 萃取物成分總量 1200 1200 3000 5500 1200 3500
2) 游離胺基酸總量: 75 300 100 3000 440 350
精胺酸 Arginine <10 <10 <10 750 <20 <10
甘胺酸 Glycine 20 20 20 100~1000 <20 <10
麩胺酸 Glutamic acid <10 <10 <10 270 55 36
組胺酸 Histidine <1.0 86 <1.0 - <10 <10
脯胺酸 Proline <1.0 <1.0 <1.0 750 <10 <10
3) 肌酸 Creatine 400 400 300 0 - 550
4) 甜菜鹼 Betaine 0 0 150 100 - -
5) 氧化三甲胺 Trimethylamine oxide 350 250 500~1000 100 0 0
6) 甲肌肽 Anserine 150 0 0 0 280 150
7) 肌肽 Carnosine 0 0 0 0 180 200
8) 尿素 Urea 0 0 2000 - - 35
1 表中的單位是指化合物的總分子量 (Shewan, 1974)。
氧化三甲胺構成海水魚非蛋白態氮 區分中的一項特徵且是重要的部分,此 成分在所有海水魚都存在,含量從肌肉 組織(乾重)的 1%至 5%,但淡水魚和陸生 生物中幾乎不存在(Anderson and Fellers, 1952; Hebard et al., 1982)。例外發現維多 利 亞 湖 產 鱸 魚 (Nile perch) 與 吳 郭 魚 (tilapia),TMAO 量達 150~200 mg/100 g 肉(Gram et al., 1989)。
雖然研究不少,TMAO 的來源與扮 演 的 作 用 不 明 處 仍 多 。
Stroem et al.
(1979)指出:TMAO 是透過某些動物性浮 游生物中的生合成而形成,這些生物具 有 一 種 酵 素 三 甲 ( 基 ) 胺 單 氧 合 酶 (TMA mono-oxygenase; trimethylamine; TMA 三 甲胺)可將 TMA 氧化成為 TMAO。TMA 普遍存在於海洋生物,如同許多其它甲 基化胺類(methylated amines)如甲(基)胺 (monomethylamine) 和 二 甲 ( 基 ) 胺 (dimethylamine),當攝食浮游生物的魚類 (plankton-eating fish)捕食這些動物性浮游 生 物 後 , 就 獲 取 了 TMAO 。Belinski (1964)、Agustsson and Stroem (1981)也指 出:某些魚類能夠從 TMA 合成 TMAO,
但這樣合成的重要性小。
TMA-氧化酶(TMA-oxidase)系統存在 於細胞的微粒體(microsomes),其活性取 決 於 菸 草 醯 胺 腺 嘌 呤 二 核 苷 酸 磷 酸 鹽 (nicotinamide adenine dinucleotide phosphate; NADPH)的存在:
(CH3)3N + NADPH + H+ + O2 → (CH3)3NO + NADP+ + H2O
令人費解的是,這單氧合酶(mono- oxygenase)卻發現廣泛存在於哺乳動物(認 為作解毒劑功能),大多數魚類的該酵素 活性低或無法測出。Kawabata (1953)指出 某些表層魚類(pelagic fishes)的血合肉中 存 在 TMAO 還 原 系 統 (TMAO-reducing system)。
肌肉組織中的 TMAO 含量取決於種 類、季節、漁場等。通常,最高含量存 在於軟骨魚和魷魚(75~250 mg N/100 g);
鱈魚的含量稍低(60~120 mg N/100 g),而 比目魚和表層魚類的含量最低;更廣泛 的數據描述可參考 Hebard et al. (1982)的 綜述。根據 Tokunaga (1970),表層魚類 如 沙 丁 魚 (sardines) 、 鮪 魚 (tuna) 、 鯖 魚 (mackerel),以血合肉中的 TMAO 濃度最 高,但底棲的白肉魚其普通肉中的含量 更高。
在軟骨魚類,TMAO 似在滲透壓調 節中扮演作用,有報告顯示,將小魟魚 (small rays) 移 至淡 水 和海水 (1 : 1) 混合 中,細胞內的 TMAO 濃度降低 50%。硬 骨魚類中 TMAO 的作用更加不清楚。
關 於 TMAO 的角色有幾種假設提 出:
TMAO 完 全 是 一 種 廢 棄 的 產 物,TMA 的解毒化型式
TMAO 是 一 種 滲 透 壓 調 節 物 (osmoregulator)
TMAO 作為一種抗凍結 ("anti- freeze") 的功能
TMAO 不具有意義的功能,魚 餵食含 TMAO 的餌料,就蓄積 肌肉
根據 Stroem (1984)的報告,目前普 遍認為 TMAO 的滲透壓調節作用。在 Gram et al. (1989)發表以前,TMAO 都僅 限 於 海 洋 物 種 中 存 在 ; 也 有 報 告 推 測 TMAO 連同高含量的牛磺酸可能有額外 的作用,至少針對淡水魚(Anthoni et al., 1990)而言。
定量上,非蛋白態氮區分的主要組 成分為肌酸(creatine),在靜止態(resting) 的魚,大部分肌酸都磷酸化(以肌酸磷酸 creatine phosphate 型式),提供肌肉收縮 所需的能量。非蛋白態氮區分含相當量 的游離胺基酸,鯖魚(Scomber scombrus) 的普通肉中達 630 mg/100 g,在大西洋鯡 魚 (Clupea harengus) 為 350~420 mg/100 g,在毛麟魚(Mallotus villosus)為 310~370 mg/100 g。不同胺基酸的相對重要性因種 類而異,大部分魚類中,牛磺酸、丙胺 酸 、 甘 胺 酸 與 含 咪 唑 (imidazole- containing)胺基酸等的含量大抵較高,含 咪唑胺基酸之中,組胺酸(histidine)最受 注目,因被微生物的脫羧基化之後就生 成組織胺(histamine)。活動性強、帶血合
肉魚種譬如鮪魚和鯖魚都含有高量的組 胺酸。
魚死後核苷酸(nucleotides)與其裂解 物的含量取決於魚的狀態,在後面章節 介紹。
(五) 維生素和礦物質
維生素和礦物質的含量是種屬特異 的 (species-specific) , 也 會 因 季 節 而 變 動。通常,魚肉是維生素 B 的好來源,
多脂魚也是維生素 A 與 D 的好來源。一 些淡水魚如鯉魚含有的硫胺酶(thiaminase) 活性高,因而這些魚種的硫胺(thiamine;
B1)含量通常都低。至於礦物質,魚肉被 認 為 是 鈣 和 磷 的 很 好 來 源 , 特 別 也 是 鐵、銅和硒。鹹水魚的碘含量高,表 5 和表 6 分別列出一些維生素與礦物質的 含量。由於這些組成分的自然變動,給 準確的數字是有所困難。
除了多脂魚的魚肉含多量的維生素 A 與 D,以及 譬如鱈 魚(cod)與比目魚
表 6. 魚肌肉中的礦物質組成分
元素 平均值 (mg/100 g) 範圍(mg/100 g)
鈉 72 30~134
鉀 278 19~502
鈣 79 19~881
鎂 38 4.5~452
磷 190 68~550
資料來源:Murray and Burt, 1969。
表 5. 魚的維生素含量
魚類 A (IU/g) D (IU/g) B1 (thiamine) (µ/g)
B2 (riboflavin) (µ/g)
Niacin (µ/g)
Pantothenic acid (µ/g)
B6
(µ/g)
鱈魚肉片 0~50 0 0.7 0.8 20 1.7 1.7
鯡魚肉片 20~400 300~1000 0.4 3.0 40 10 4.5
鱈魚肝油 200~10.000 20~300 - 1) 3.4 1) 15 1) 4.3 -
1) 整個肝臟 ;資料來源:Murray and Burt (1969)。
(halibut)等種類的肝臟也含量豐富之外,
魚類的維生素含量相似於哺乳動物。但 須 注 意 的 是 , 魚 肉 的 鈉 含 量 相 對 地 較 低,適用於低鈉膳食。
在養殖魚類,維生素和礦物質的含 量 會 反 映 出 魚 飼 料 中 對 應 組 成 分 的 組 成,雖然數據的解釋上須小心。為保護 對於魚與人健康都重要的 n-3 高度不飽和 脂肪酸,飼料中會添加維生素 E 作為抗 氧化劑,報告指出在魚組織中的維生素 E 濃度相當於在飼料中的濃度(Waagboe et
al., 1991)。
三 、 魚 類 的 死 後 變 化 (Postmortem
changes)
(一) 感官的變化
感官的變化(Sensory changes)是指感 官 上 所 感 受 得 到 的 變 化 , 譬 如 外 觀 (appearance) 、氣味 (odor) 、 質 地 (texture) 和滋味(taste)。
1. 生鮮魚(fresh fish)的變化
魚類貯藏過程中,首先引人注意的 感官的變化是外觀和質地。魚類的特徵 滋 味 通 常 在 冰 藏 的 前 幾 天 期 間 仍 舊 保 留。
最 激 烈 的 變 化 為 死 後 僵 直 (rigor mortis)的發生。剛死去的魚肌肉仍處於鬆 弛 狀 態 , 具 有 彈 性 的 質 地 常 持 續 數 小 時,之後肌肉才會收縮。當成為硬直之 後,整個魚體變硬,魚即處於死後僵直 的狀態,這狀況延續一天或以上,然後 僵直解除。死後僵直的解除使肌肉再度
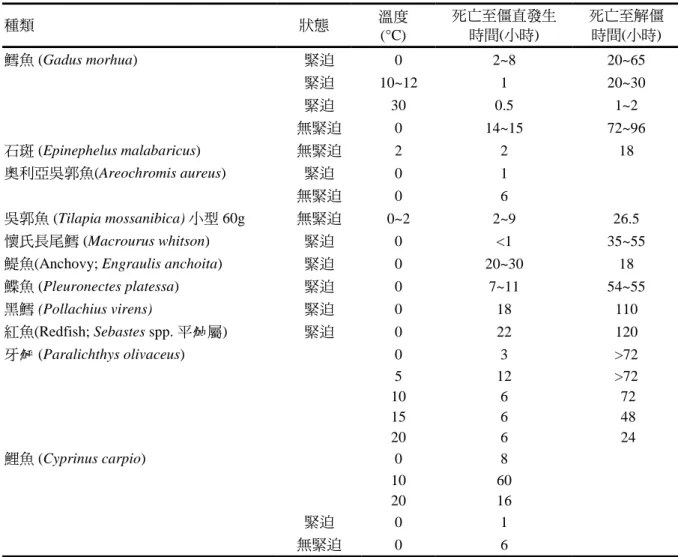
鬆弛並軟化,但彈性觸感已不如僵直之 前。死後僵直的發生與消失的速率因魚 種而異,溫度、處置方式、大小與魚的 物理狀態等也有影響(表 7)。
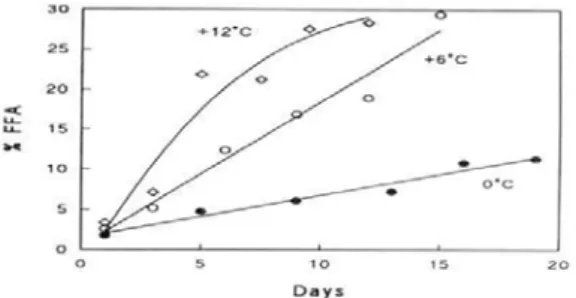
溫度對僵直的影響並不一致。鱈魚 為例,高溫下死後僵直發生快速且很強 烈,必須避免這樣的情況,因死後緊縮 強烈會導致肌肉的龜裂(gapping),即弱化 結締組織而使魚片破裂。
一般認為死後僵直的發生與時間是 在高溫下較快速,但另特別觀察熱帶魚 種,卻顯示溫度對於僵直發生的影響剛 好相反,這些魚種在 0℃比在 10℃更快 速發生僵直,且和在 0℃的生化學變化有 關連(Poulter et al., 1982; Iwamoto et al., 1987)。Abe and Okuma (1991)解釋:鯉魚 (Cyprinus carpio)死後僵直的發生取決於 原棲息的水域溫度和貯藏溫度,當兩者 的溫度差大時從死亡至僵直發生之時間 就 短 , 反 之 溫 度 差 小 時 發 生 的 時 間 就 長。
如果魚飢餓或者肝醣儲存已耗盡、
或者魚類受緊迫,死後僵直於死後立即 或很快地開始。用來迷昏與猝死魚的方 法也影響僵直的發生,以低溫(將魚置冰 水中使之失溫而死亡)使之昏迷與猝死僵 直發生最快,而敲擊頭部而致死則可延 遲 至 多 達 18 小 時 (Azam et al., 1990;
Proctor et al., 1992)。
當僵直前或僵直中切割魚片,死後 僵 直 在 加 工 技 術 上 的 意 義 就 顯 得 很 重 要。僵直中的魚體是完全僵硬的,魚片
表 7. 不同魚種死後僵直的發生時間與期間
種類 狀態 溫度
(°C)
死亡至僵直發生 時間(小時)
死亡至解僵 時間(小時)
鱈魚 (Gadus morhua) 緊迫 0 2~8 20~65
緊迫 10~12 1 20~30
緊迫 30 0.5 1~2
無緊迫 0 14~15 72~96
石斑 (Epinephelus malabaricus) 無緊迫 2 2 18
奧利亞吳郭魚(Areochromis aureus) 緊迫 0 1
無緊迫 0 6
吳郭魚 (Tilapia mossanibica) 小型 60g 無緊迫 0~2 2~9 26.5
懷氏長尾鱈 (Macrourus whitson) 緊迫 0 <1 35~55
鯷魚(Anchovy; Engraulis anchoita) 緊迫 0 20~30 18
鰈魚 (Pleuronectes platessa) 緊迫 0 7~11 54~55
黑鱈 (Pollachius virens) 緊迫 0 18 110
紅魚(Redfish; Sebastes spp. 平鮋屬) 緊迫 0 22 120
牙鮃 (Paralichthys olivaceus) 0 3 >72
5 12 >72
10 6 72
15 6 48
20 6 24
鯉魚 (Cyprinus carpio) 0 8
10 60
20 16
緊迫 0 1
無緊迫 0 6
資料來源:Hwang et al., 1991; Iwamoto et al., 1987; Korhonen et al., 1990; Nakayama et al., 1992; Nazir and Magar, 1963; Partmann, 1965; Pawar and Magar, 1965; Stroud, 1969; Trucco et al., 1982。
的採肉率會很差,且處理粗糙可能另引 起龜裂。如果僵直前就切除骨架的魚片 就能自行收縮,因而隨著僵直的發生魚 片會變短,血合肉(dark muscle)收縮可達 52%,而普通肉(white muscle)達原長度的
15%
(Buttkus, 1963)
。 如 僵 直 之 前 就 煮魚,質地變成很軟、有些糊狀般,反之 如煮僵直中的魚,其質地硬但不乾澀,
而僵直後的魚會變得緊實、多汁且有彈 性。
凍結僵直前的全魚和魚片,如細心 地在低溫下解凍,為使仍凍結的肌肉通 過死後僵直的時間進程,也能得到好的 產品。
在 市 場 和 卸 貨 地 點 , 生 鮮 魚 (raw fish)的感官品評乃評定外觀、質地和氣 味。魚類的感官屬性(sensory attributes)列 於表 8, 多 數 的 評 分 制 是 植 基 於 融 冰 (melting ice)貯藏期間所發生的變化,必 須 記 得 特 徵 的 變 化 會 因 貯 藏 方 法 而 改
變。貯藏於不加冰塊的冷藏條件下,魚 類外觀的變化不若冰藏魚那樣的複雜,
但魚腐敗更快速,故也需要品評煮熟風 味(cooked flavor)。因而,漁船卸貨後,
瞭解魚的時間/溫度歷程是不可欠缺的。
魚僵直後的特徵感官變化,因種類 與貯藏方法的不同而差別大。一般的說 明載列在歐洲經濟共同體(EEC)提出的魚 類品質評定指引中,如表 8。所建議的尺 度從 0 至 3 號碼,3 為最佳的品質。西歐 魚類技師協會也編寫氣味和風味有關的 多種語言語彙,當作魚類鮮度的感官品 評而尋找描述用語時幫助很大(Howgate et al., 1992)。
2. 食用品質(eating quality)的變化
如 需 要 冷 藏 魚 在 貯 藏 中 的 品 質 標 準,可進行煮熟魚的感官評定,針對煮 熟魚介類的一些屬性列於表 8。魚類冰藏 中,品質變差的特徵樣式,分為下列 4 個階段:
A. 第 一 階 段 : 魚 很 新 鮮 , 帶 有 甜 的、海藻般與好吃般的滋味,滋 味 中 可 能 有 非 常 弱 的 金 屬 般 味 道 。 鱈 魚 (cod) 、 黑 線 鱈 (haddock)、牙鱈(whiting)和比目魚 (flounder)在捕獲 2-3 天後甜味達最 高。
B. 第二階段:特徵氣味與滋味有些 減弱,魚肉變為中性的(neutral),
但仍無不良氣味,質地也令人喜 歡的。
C. 第三階段:出現腐敗的跡象,依 魚種與腐敗型式(好氧的 aerobic、
厭氧的 anaerobic)的不同而產生一 系列揮發性不愉快的氣味物質,
揮 發 性 化 合 物 之 一 的 三 甲 胺 (trimethylamine; TMA)源自氧化三 甲胺(trimethylamine oxide; TMAO) 的細菌性還原,三甲胺具有非常 特別的魚腥味(fishy smell),在第 三階段初期的不良風味可能略有 酸味(sour)、果香(fruity)與微苦味 (bitter),尤其在多脂魚。至後期期 間,死甜般(sickly sweet)、類似白 菜 (cabbage-like) 、 氨 味 (ammoniacal)、硫味(sulphurous)與 油燒(rancid)等氣味產生,質地變 得軟而多水般的或者乾硬的。
D. 第四階段:魚的特徵為已變壞的 (spoiled)和腐敗的(putrid)。
號碼尺度(numbered scale)可用於煮 熟魚的感官品評,如圖 6所示。尺度從 0 至 10 編 號 , 10 為 絕 對 新 鮮 (absolute freshness),8 為品質佳(good quality),6 為中性無味的魚(a neutral tasteless fish),
拒絕的級別為 4,使用這種方式的尺度其 圖形變成 S-型,顯示魚的品質在第一階 段快速下降,在第二階段速率較緩,在 第三與最後的階段當魚腐敗時速率又變 快。
其它尺度也能夠適用而變更圖形的 形狀,但重要的是要知道必需從感官分
表 8. 鮮度評級(Freshness ratings):理事會法規(歐洲經濟共同體)第 103/76 OJ No. L20 (1976 年 1 月 28 日) (EEC, 1976)
標準(Criteria)
標識(Marks)
檢查部位 3 2 1 0
外觀
皮 明亮、特有色彩、
無褪色 水漾般透明黏液
有色彩但無光澤 黏液稍混濁
逐漸褪色與暗沉 黏液白濁
1色澤暗沉 黏液混濁
眼 凸出 (鼓起)
眼睛透明 瞳孔黑且明亮
稍下陷但仍凸出 眼睛稍不透明 瞳孔黑但暗沉
平坦 眼睛不透明 瞳孔不透明
中間凹陷 眼睛白濁 瞳孔白灰色
鰓 顏色明亮
無黏液
顏色略淡 有些清澈黏液
已變色 不透明黏液
1偏黃
白濁黏液
肉(腹部) 帶藍色、透明、光
滑、光亮 原有顏色
絲絨般、蠟質、暗沉 顏色稍改變
有些不透明 1不透明
顏色(順沿脊椎) 本色的 略粉紅 粉紅 1紅色
臟器 腎及其餘臟器應是
亮紅、大動脈內部 有血
腎及其餘臟器應是暗紅 的;血液退色
腎及其餘臟器以及 血液為但紅色
腎及其餘臟器以及血 液為褐色
狀態
肉 緊實有彈性
表面平滑
彈性略失 稍軟 (鬆弛)、失彈性
表面蠟質般暗沉
變軟 (鬆弛) 鱗片易脫落、體表皺
縮、蒼白
脊椎 斷裂而不是分離 黏附 稍黏附 1不黏附
腹膜 完全黏附於肉 黏附 稍黏附 1不黏附
嗅聞 鰓、皮、腹
腔
海藻味 聞不出海藻味或任何不
好的氣味
些微酸味 1酸味
1 或處於更加衰退的狀態。
圖 6. 冰 藏 (0 ℃ ) 鱈 魚 的 食 用 品 質 變 化 (Huss, 1976)。
析取得怎樣類型的結果,以便提出正確 的問題給感官評定者。
(二) 自家消化的變化(Autolytic changes) 自 家 消 化 (autolysis) 意 指 自 行 的 消 化,很早以前即瞭解魚類的腐敗至少有 兩種型式:細菌性和酵素性。Uchiyama and Ehira (1974)指出鱈魚(cod)和黃鰭鮪 (yellowtail tuna)的鮮度受酵素性的變化所
主導,較少關連於微生物的品質變化。
在有些魚種(魷魚 squid、鯡魚 herring),
酵素性的變化優先主導冷藏魚的腐敗。
在 其 它 魚 種 , 除 了 微 生 物 介 導 的 過 程 (microbially-mediated processes),自 家消 化對整體品質的下降,貢獻程度不同。
1. 僵直後肌肉中能量的產生
剛死當時,由於心臟不再幫浦血液 與透過腮的循環,氧氣供應至肌肉組織 即中斷。因得不到氧氣作正常呼吸,從 所消化的營養素轉化為能量的產生就大 受限制。圖 7說明大多數活硬骨魚(teleost fish; bony finfish)中肌肉產生能量的正常 途徑,肝醣(儲藏的碳水化合物)或脂肪被 組織內酵素氧化或者燃燒,經由一系列 反應而最後產生二氧化碳、水和高能量 的 腺 苷 三 磷 酸 (adenosine triphosphate;
ATP),這類型的呼吸包含二個階段:厭 氧的(anaerobic)和好氧的(aerobic)階段,
後者取決於一直有氧氣的存在,而氧氣 僅能由循環系統取得,大多數甲殼類動 物在水生環境的外界也能夠呼吸,即在 有限時間內吸收大氣中的氧氣。
圖 7 也說明在厭氧的條件下,ATP 可透過其它兩種重要的途徑從肌酸磷酸 (creatine phosphate) 或 從 精 胺 酸 磷 酸 (arginine phosphate)來合成,前者能量來 源僅限於脊椎動物肌肉(硬骨魚類),後者 是一些無脊椎動物如頭足類(cephalopods) (鎖管 squid 與章魚 octopus)的特徵。在這 兩情況下,當肌酸磷酸或精胺酸磷酸耗
盡時,ATP 的生成就停止;值得注意的 是,頭足類動物的厭氧性代謝最終產物 之章魚鹼(octopine),並非酸性的(不同於 乳酸),因而在這些動物,僵直後 pH 的 任 何 改 變 和 從 肝 糖 產 生 的 乳 酸 都 無 關 連。
對大多數硬骨魚類而言,一旦心臟 停止跳動,醣解(glycolysis)是能量產生的 唯一可能途徑,這效率更低的過程其最 終產物以乳酸與丙酮酸為主,又經由醣 解而產生 ATP,每莫耳葡萄糖的氧化僅 產生 2 莫耳 ATP,相對於在活體動物的 粒腺體中,如醣解的最終產物進行好氣 性 氧 化 , 每 莫 耳 葡 萄 糖 產 生 3 6 莫 耳 ATP。因而死後,厭氧態肌肉無法維持正 常的 ATP 濃度,且當細胞內的含量從 7~10 µmoles/g 降至<1.0 µmoles/g,肌肉 進入死後僵直。死後的醣解會導致乳酸 的蓄積並降低肌肉的 pH。鱈魚(cod)的 pH 從 6.8 降為最終 pH 的 6.1~6.5,一些 魚 種 的 最 後 p H 值 更 低 : 大 型 鯖 魚 ( m a c k e r e l ) 的 最 終 僵 直 p H 可 降 至 5.8~6.0,鮪魚(tuna)和比目魚(halibut)更低 至 5.4~5.6,但這樣低的 pH 值在海水硬 骨魚類並不常見,這些 pH 值鮮少如僵直
圖 7. 魚類肌肉中肝醣的好氧性與厭氧性裂解。
後哺乳動物肌肉所觀察到的那樣低,例 如,牛肌肉在死後僵直中常降至 pH 5.1 附近,所產生的乳酸量和活組織中貯藏 碳水化合物(肝醣)的含量有關,通常,相 較於哺乳動物,魚類肌肉所含的肝糖量 較低,因而死後產生的乳酸量遠不及哺 乳動物,而且,魚類的營養狀態以及死 亡前遭遇的緊迫與運動對於肝醣儲存量 的 影 響 巨 大 , 因 而 最 終 影 響 至 僵 直 後 pH。一般通則,充分休息與餵食的魚比 疲 勞 魚 含 有 更 多 的 肝 醣 。Chiba et al.
(1991)指出日本泥鰍(Japanese loach)捕抓 前僅緊迫數分鐘,就導致 3 小時內 pH 降 低 0.50 單位,在未掙扎組僅下降 0.10 單 位,另也指出魚類放血(bleeding)會明顯 減少僵直後乳酸的產生。
僵直後魚類肌肉 pH 的下降會影響肌 肉的物理性質。當 pH 降低,肌肉蛋白質 表面上的淨電荷(net charge)減少,導致蛋 白質部分變性而損失一些保水性(water- holding capacity)。死後僵直狀態中的肌 肉組織,煮後會失去水分,故特別不適 合於包括加熱的後續加工,此乃熱變性 會促進水分的流失。水分損失不利於魚 類肌肉的質地,Love (1975)指出肌肉韌 度(toughness)和 pH 之間呈負相關性,不 能接受的韌度程度(及煮時的水分損失)發 生在較低的 pH 值(圖 8)。
2. 自家消化和核苷酸分解作用
如前所述,當肌肉中的 ATP 濃度降 至<1.0 µmoles/g,死後僵直開始發生。
圖 8. 鱈魚肌肉質地與 pH 的相關性(Love, 1975)。
黑心圓代表大西洋聖基爾達(St. Kilda)捕獲,
三角代表戴維斯海峽 Fells Bank 捕獲。
在活的動物,不僅是肌肉收縮所需的高 能 量 來 源 , ATP 也 作 為 肌 肉 增 塑 劑 (muscle plasticizer)。肌肉收縮本身受到鈣 與存在每個肌肉細胞中的酵素 ATP-ase 的控制,當細胞內 Ca2+濃度為 1 µM,
Ca2+-活化的 ATP-ase 降低了導致主要收 縮蛋白質之間[肌動蛋白(actin)和肌球蛋 白 (myosin)] 的 交 互 作 用 之 肌 肉 游 離 態 ATP 含 量 , 最 後 使 得 肌 肉 縮 短 (shortening),變得僵硬與無法伸展。死後 僵直中的魚不能正常地切剖魚片(filleted) 或加工,因處理時魚體過於僵硬,且經 常扭曲變形,不適合機械處理。
僵 直 的 解 除 仍 是 未 完 全 瞭 解 的 過 程,但總是導致肌肉組織在隨後軟化(鬆 弛),且被認為和自然存在的一種或更多 種肌肉酵素有關,即消化分解死後僵直 複合物(rigor mortis complex)的某些組成 分。僵直解除過程中,肌肉的軟化和自 家消化的變化是平行發生,這些的變化 之中,最早被確認之一的是 ATP 相關化 合物的降解,圖 9 表示 ATP 降解而依序 生 成 腺 苷 二 磷 酸 (adenosine diphosphate;
ADP) 、 腺 苷 單 磷 酸 (adenosine
monophosphate; AMP) 、 肌 苷 單 磷 酸 (inosine monophosphate; IMP) 、 肌 苷 (inosine; Ino) 與 次 黃 嘌 呤 (hypoxanthine;
Hx)。在大多數魚類,ATP 分解代謝物的 降解都以同樣的方式進行,惟每個單獨 反應(從一分解物至另一分解物)的速率因 魚種的不同而變動很大,然其進展程度 常和訓練型分析員所測得感知的腐敗程 度(perceived level of spoilage)一致。Saito
et al. (1959)最早觀察到此一模式,並根據
這些的自家消化變化,針對魚類鮮度建 立下列的公式:K%= [Ino]+[Hx]
[ATP]+[ADP]+[AMP]+[IMP]+[Ino]+[Hx] × 100
其 中 , [ATP] 、 [ADP] 、 [AMP] 、 [IMP]、[Ino]和[Hx]分別表示冰藏過程中 在不同時間測得的魚肉中這些化合物的 相對濃度值。
這 K 或“鮮度”指標,最初是根據肌 肉死後僵直貯藏過程中發生的自家消化 變化,而提出一個相對的鮮度評級,因 此,K 值愈高代表鮮度等級愈低。但不 幸 的 ,一 些魚 種如 大西 洋 鱈 魚 (Atlantic
cod)在訓練型評審員所測得的貯藏期限之 前,就已出現最大的 K 值,因而認為 K 值對所有海洋魚類並不全是可信賴的鮮 度指標。而且,核苷酸分解物的降解只 有和鮮度的感知變化(perceived changes in freshness) 一 致 , 不 一 定 與 鮮 度 劣 敗 (freshness deterioration)的起因有關連,因 為 只 有 次 黃 嘌 呤 被 認 為 對 變 壞 的 魚 (spoiled fish)所感覺的苦的不良風味有直 接的影響(Hughes and Jones, 1966)。一般 認為腺苷單磷酸(IMP)負責貢獻生鮮魚的 風味,而腺苷單磷酸僅高品質的水產品 中存在,自家消化過程中除 ATP 的降解 當然關連於死後僵直,但並無一種核苷 酸分解物被認為與質地的感知變化有關 連。
Surette et al. (1988)分析比較滅菌和 無滅菌鱈魚的 ATP 分解代謝物,鱈魚組 織的滅菌和無滅菌樣品中,IMP 的生成 與分解速率都相同(圖 10a 與圖 10b),顯 示經由肌苷(inosine)的 ATP 降解之異化代 謝途徑(catabolic pathway)完全來自於自家
圖 9. 僵直後魚肉中腺苷三磷酸(ATP)的降解。酵素包括:1 為腺苷三磷酸酶(ATP-ase);2 為肌激酶 (myokinase) ; 3 為 腺 苷 單 磷 酸 脫 氨 酶 (AMP deaminase) ; 4 為 肌 苷 單 磷 酸 磷 酸 水 解 酶 (IMP phosphohydrolase) ; 5a 為 核 苷 磷 酸 酶 (nucleoside phosphorylase) ; 5b 為 肌 苷 核 苷 酶 (inosine nucleosidase);6、7 為黃嘌呤氧化酶(xanthine oxidase)。(Gill, 1992)。
消化酵素。無滅菌樣品的肌苷轉化成次 黃嘌呤快約 2 天,表明細菌性的核苷磷 酸酶(圖 9中酵素 5a)在冷藏鱈魚僵直後的 次黃嘌呤產生扮演重要的角色。值得注 意的是,剛宰殺的鱈魚也無法回收取得 核 苷 磷 酸 酶 (Surette et al., 1988) , 後 來 Surette et al. (1990)從 腐 敗 的 鱈 魚 片 (spoiled cod fillet)分離出變形桿菌(Proteus bacterium) , 並 取 得 核 苷 磷 酸 酶 。 如 前 述,不同魚種的核苷酸降解樣式變動很 大,在圖 11,顯示不同類型魚之間的次 黃嘌呤變動,很清楚的,次黃嘌呤測定 並 不 適 用 於 旗 魚 (swordfish) 和 紅 魚 (redfish)等魚種。
毋庸置疑,物理處理會加速冷藏魚 的自家消化變化。Surette et al. (1988)指 出核苷酸分解為代謝物的降解速率,在 滅菌魚片比在無滅菌去腮的全魚更快,
這或許無需懷疑,因為許多的自家消化 酵素是被分隔存在於離散的膜結合組包 (membrane-bound packages)中,當受到物 理性傷害就破裂,導致酵素與基質充分 混 合 。 冰 塊 或 其 它 魚 所 造 成 魚 體 的 破 損,嚴重影響食用率與魚片切割產率,
即使魚體的微生物含量不高,這突顯自 家消化過程的重要性。嚴禁冰藏魚貯放 盒中的深度超過 30 公分,也要確認貯存 盒不可上下堆疊過密,如果自家消化要 以最低程度進行。輸送魚貨與從漁船卸 魚 貨 也 須 設 計 , 以 期 避 免 損 傷 魚 體 組 織。
一些快速測定個別或全部核苷酸分
(a)
(b)
圖 10. (a)滅 菌鱈 魚 肉片 於貯 藏 3 ℃的 肌 苷酸 (IMP) 、 肌 苷 (inosine) 和 次 黃 嘌 呤 (hypoxanthine) 變 化 (Gill, 1990); (b)無滅菌 鱈魚肉片於貯藏 3℃的肌苷酸(IMP)、肌苷 (inosine) 和 次 黃 嘌 呤 (hypoxanthine) (Gill, 1990)。
圖 11. 一 些 魚 種 於 冰 藏 期 間 次 黃 嘌 呤 (hypoxanthine)蓄積速率的差異(Fraser et al., 1967)。
解 物 並 包 括 鮮 度 指 標 的 方 法 已 開 發 出 來,可參考綜述文獻(Gill, 1990; 1992)。
3. 蛋白質分解酵素參與的自家消化的變 化
從魚類肌肉已分離出多種的蛋白酶 (proteases),這些蛋白質水解方式的分解 (proteolytic breakdown)其作用往往關連魚 肉組織的過度軟化,或許自家消化方式 的蛋白質水解(autolytic proteolysis)最引人 注目的例子之一,為表層魚種(多脂魚)如 鯡魚(herring)與柳葉魚(capelin)發生腹部 破裂,這樣的組織軟化最主要發生在夏 季月份,當表層魚大量進食特別是包含 橈 角 類 (copepods) 和 磷 蝦 (euphausiids) 等
“紅色餌料 red feed”,蛋白質的自家消化 所產生的低分子量胜肽和游離胺基酸,
不僅降低表層魚的商業接受度,且在大 宗儲存的柳葉魚,自家消化會透過提供 更好的成長環境而加速腐敗微生物的生 長 (Aksnes and Brekken, 1988) 。 在 柳 葉 魚,自家消化所導致的細菌性腐敗,也 促使胺基酸的脫羧基(decarboxylation)而 產生生物胺(biogenic amines),並顯著降 低營養價值,這是特別重要的,因自家 消化與細菌生長嚴重降低用於製造魚粉 之表層魚的商業價值。
同樣在使用大宗儲存的鯡魚製作魚 粉 , 也 發 現 含 有 羧 基 肽 酶 A 與 B (carboxypeptidases A and B)、胰凝乳蛋白 酶(chymotrypsin)和胰蛋白酶(trypsin)等活 性;初期實驗顯示添加馬鈴薯萃取物可 抑制蛋白質水解,不僅減緩蛋白質水解 及微生物成長,也保存魚粉的營養價值 (Aksnes, 1989)。後來,Botta et al. (1992) 發現比起生物因素譬如魚體大小、腮中 的紅色餌料數量或卵含量,鯡魚的內臟
腔(腹部破裂)的自家消化和物理處理的關 連性更高。特別對鯡魚的凍結/解凍,在 15℃的解凍時間與冰藏時間對腹部破裂 之影響遠大於生物因素。
(1) 組織蛋白酶
雖然魚組織中已發現數種蛋白質分 解酵素(proteolytic enzymes),最常被提的 可能就是組織蛋白酶(cathepsins)。組織蛋 白酶屬酸性蛋白酶,通常存在於溶菌體 (lysozomes),在活的組織中,認為溶菌體 蛋 白 酶 是 負 責 受 傷 部 位 處 的 蛋 白 質 分 解,因而組織蛋白酶在活的組織的大多 數部位都是無活性的,當僵直後肌肉受 到物理損傷或被凍結與解凍,酵素就被 釋出進入細胞液中。
在魚類組織中的自家消化的降解,
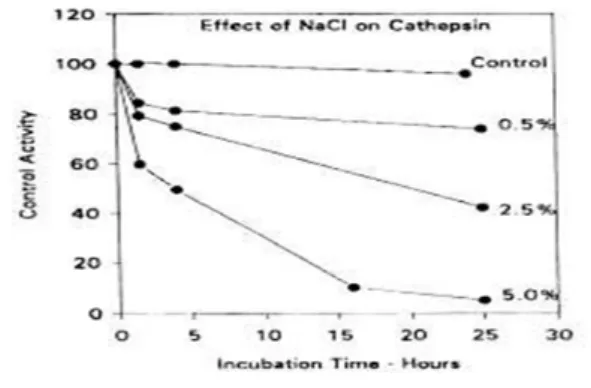
認為組織蛋白酶 D 與 L 扮演主要的作 用,由於其餘多數的組織蛋白酶其活性 的 pH 範圍較窄且很低,而無生理上的意 義。Reddi et al. (1972)指出來自冬季比目 魚(flounder)的一種被認為組織蛋白酶 D 的酵素,在 pH 3~8 範圍具有活性,最大 活性靠近 pH 4.0,雖然並未使用合成基 質或特定抑制劑來確認其酵素種類。然 而,在 ATP 存在下該酵素的活性變很 低,表明此酵素只在僵直後的魚類肌肉 才有活性,且此酵素活性受到鹽的強力 抑制(圖 12),和 5%氯化鈉一起培養 25 小時後幾乎無活性,因而該酵素在鹽藏 魚產品中不太可能表現出活性。
組織蛋白酶 L 已知和鮭魚在產卵洄
游期間的肌肉軟化有關,該酵素比起組 織蛋白酶 D 可能對魚類肌肉的自家消化 貢獻更大,因在中性 pH 的活性更大,而 且可消化肌原纖維蛋白質(actomyosin 肌 動 球 蛋 白)及結締組 織兩者。Yamashita and Konogaya (1990)證實鮭魚產卵期間,
是組織蛋白酶 L 甚於其它組織蛋白酶造 成 肉 質 的 軟 化 ; 將 純 化 的 肌 原 纖 維 (myofibrils)以組織蛋白酶 L 處理後再膠體 電泳,結果顯示電白質帶的樣式與產卵 中鮭魚肌肉蛋白質的樣式幾乎相同,甚 且,組織蛋白酶 L 的自家消化活性和儀 器 測 得 的 肌 肉 物 性 ( 破 斷 強 度 breaking strength)之間的相關性很高,在生鮮與冷 凍/解凍組織其線性相關性分別達 r = 0.86 及−0.95。有趣的是在所有舉例,以測定 組織蛋 白酶 L 活性代表自家消化能力 (autolytic ability)時,在冷凍/解凍組織所 測得的數值都高於生鮮組織。凍結與解 凍常會打破細胞膜,而使自家消化的膜 結合型酵素可和其天然的基質反應,作 者並繼續探討這酵素與其天然存在的抑 制劑(Yamashita and Konogawa, 1992)。組
圖 12. 氯化鈉對組織蛋白酶活性的影響(Reddi et al., 1972)。
織蛋白酶 L 也和比目魚(flounder)的膠狀 樣 (jelly-like) 軟 化 (Toyohara et al., 1993b)、粘孢子蟲 (Myxosporidia)寄生大 西洋鱈魚(Pacific hake)肌肉的不可控制的 軟化(Toyohara et al., 1993a)等有關。這樣 被感染魚的組織商業價值低,但目前仍 不清楚到底是寄生蟲或宿主分泌蛋白質 分解酵素讓肌肉進行自家消化。
除了有害質地,組織蛋白酵素可誘 導發酵魚產品進行所要的自家消化的變 化,例如在鹽醃製日本魷(Japanese squid) 與鯽魚(Crucian carp)的發酵過程中,組織 蛋 白 酶 被 認 為 是 負 責 主 要 的 質 地 變 化 (Makinodan et al., 1991; 1993)。
(2) 鈣蛋白酶
細胞內蛋白酶的第二群稱為鈣蛋白 酶 (calpains) 或 鈣 活 化 因 子 (calcium activated factor; CAF),近年來也瞭解和 魚類肌肉的自家消化有關連,存在於肉 類 、 魚 類 和 甲 殼 類 。 已 瞭 解 近 一 個 世 紀 , 紅 肉 (red meat) 的 死 後 熟 成 (post mortem aging) 導 致 肉 質 的 嫩 化 (tenderization)。已知鈣蛋白酶是主要負責 肉類的死後自家消化,透過消化肌原纖 維(myofibril)的 Z-線蛋白質,雖然韌性 (toughness)鮮少是未冷凍魚肉的問題,透 過自家消化的軟化卻是限制商業價值的 一項嚴重課題。鈣蛋白酶屬於細胞內的 內切胜肽酶(intracellular endopeptidases),
需要半胱胺酸(cysteine)與鈣;μ-鈣蛋白酶 需要 5~50 μM Ca2+,而 m-鈣蛋白酶需要
150~1000 μM Ca2+,多數鈣蛋白酶在生理 pH 下都具活性,這合理懷疑它們在冷藏 過程中的魚肉軟化的重要性。
研究顯示在甲殼類肌肉中,鈣蛋白 酶與脫殼誘導的(molt-induced)肌肉質地 變化有關,即進行肌原纖維蛋白質的非 特異的一般性消化。但是,已知脊椎動 物肌肉鈣蛋白酶是高專一性的,主要消 化 肌 鈣 蛋 白 -T (tropinin-T) 、 結 蛋 白 (desmin)、肌聯蛋白 (titin)與伴肌動蛋白 (nebulin),不會攻擊脊椎動物的肌動蛋白 (actin) 與 肌 球 蛋 白 (myosin) (Koohmaraie, 1992)。相反的,魚類鈣蛋白酶消化肌球 蛋白(尤其是肌球蛋白重鏈 myosin heavy chain),形成分子量約 150000 Da (Dalton;
道 爾 頓 ) 的 初 始 片 段 (Muramoto et al., 1989)。同作者也提及較之哺乳動物鈣蛋 白酶,魚類鈣蛋白酶在低溫下的活性更 高,斷裂速率會因種類而異,對熱穩定 性最低的肌球蛋白之活性最高。因而比 起熱帶水域的魚種,順應較低環境溫度 的 魚 種 更 容 易 進 行 鈣 蛋 白 酶 的 自 家 消 化,雖然包括鯉魚(carp) (Toyohara et al., 1985)、吳郭魚(tilapia)與蝦(shrimp) (Wang
et al., 1993) 、 以 及 鮪 魚 (tuna) 、 黃 花 魚
(croaker) 、 嘉鱲 (seabream) 與 鱒 魚 (trout) (Muramoto et al., 1989)等一些魚種都鑑定 出鈣蛋白酶,但鈣蛋白酶活性和質地的 儀器測定之間的「因果」關係鮮少被提 及。(3) 膠原蛋白酶(Collagenases)
在這點,所有提到的死後自家消化 的變化都牽涉於肌肉細胞本身之內的變 化 , 但 硬 骨 魚 的 肌 肉 被 稱 為 肌 隔 (myocommata)之結締組織所分隔,而分 成「肉片 flakes」或肌節(myotomes)的肌 肉細胞塊(blocks of muscle cells),每個肌 肉細胞或纖維被結締組織包圍,結締組 織藉著細膠原蛋白纖維(thin collagenous fibrils)連接細胞尾端的肌隔。冷藏期間,
這 些 纖 維 變 質 (Bremner and Hallett, 1985) , 後 來 研 究 又 指 出 , 當 冷 藏 鱒 魚 (trout)肌肉的 V 型膠原蛋白被溶化,質地 儀器測定值即下降,推測是由於自家消 化的膠原蛋白酶的酵素作用(Sato et al., 1991)。當冰藏長時間或高溫下的短時間 貯藏,大抵是這些酵素引起肌節的裂開 (gapping)或崩解。大西洋鱈(Atlantic cod) 若貯存溫度升至 17°C,就不可避免地裂 開,推測是結締組織的降解和由於高溫 僵直而使肌肉快速縮短之緣故。
冷 藏 大 蝦 (prawn) 由 於 組 織 的 軟 化 (softening)而貯藏壽命較短,也是由於膠 原蛋白酶酵素的存在(Nip et al., 1985),大 蝦 的 膠 原 蛋 白 酶 酵 素 存 在 頭 胸 部 (hepatopancreas) (消化器官)。
4. 凍藏中自家消化的變化
氧 化 三 甲 胺 (trimethylamine oxide;
TMAO)在許多海洋硬骨魚(teleost fish)是 作為一種滲透壓調節化合物,通常受到 微 生 物 的 作 用 而 降 解 成 為 三 甲 胺 (trimethylamine; TMA),但有些魚種的肌
肉也存在一種酵素能將 TMAO 降解為二 甲 胺 (dimethylamine; DMA) 和 甲 醛 (formaldehyde):
(CH3)3 NO → (CH3)2NH + HCHO
注意甲醛和 DMA 兩者的生成量是等 莫耳的關係當然是重要,但在商業上的 意義更大。甲醛誘導肌肉蛋白質的交聯 (cross-linking),使得肌肉變硬且很快失去 保水力(water-holding capacity)。擔負這甲 醛誘導的韌化(toughening)之酵素稱為氧 化三甲胺酶(TMAO-ase)或氧化三甲胺脫 甲基酶(TMAO demethylase),最常見於鱈 科 魚 類 (gadoid fishes) ( 鱈 魚 家 族 cod family),至今報導的大多數氧化三甲胺 脫 甲 基 酶 都 屬 膜 結 合 型 (membrane- bound),當組織膜被凍結或人工使用界面 劑的溶化而破壞時,變得最具活性。血 合(紅色)肉中的活性大於白色肉,其它的 組 織 如 腎 、 脾 和 膽 囊 中 的 酵 素 量 特 別 多,因而如從鱈科魚類製造魚漿並予凍 藏時,要避免肉質的韌化,就得完全去 除腎等器官組織。機械去骨之前,要確 定腎移除有否通常是有困難,因為這特 別的器官沿著整個骨架而黏附。氧化三 甲胺酶已分離自狗鱈(hake)肌肉的粒腺體 區 分 (microsomal fraction) (Parkin and Hultin, 1986) 和 腎 組 織 的 溶 菌 體 膜 (lysosomal membrane) (Gill et al., 1992),
並顯示冷凍狗鱈肌肉的韌化和甲醛的生 成量有關,凍藏溫度高則甲醛的生成速 率愈大(Gill et al., 1979)。另外,甲醛誘
導的韌化也受到凍結前的不當處理漁獲 與凍藏期間的溫度跳動而促進,防止自 家消化產生甲醛的最實際做法,是將魚 凍藏−30°C 以下且避免溫度變動,以及避 免不當處理或魚體凍結前受到擠壓等。
自家消化的變化對生鮮與冷凍魚食用性 的影響,彙整於表 9。通常,影響自家消 化之最重要單一因素是肌肉細胞的物理 性破壞,在此處並不討論鹼性蛋白酶所 關連煮熟煉製品的軟化問題,Kinoshida
et al. (1990)
曾 探 討 熱 活 化 型 (heat- activated) 鹼性蛋白 酶 (alkaline proteases) 和魚漿產品的軟化之關聯。(三) 細菌的變化(Bacteriological changes) 1. 活魚的菌相
活魚與剛捕獲魚的外表全部(皮與腮) 和腸道都有微生物存在,但總數量變動 很大。Liston (1980)指出正常範圍是 102- 107 cfu (菌落形成單位)/cm2皮膚表面,在 腮與腸道兩者含 103至 109 cfu/g (Shewan, 1962)。
剛捕獲魚的菌群取決於捕獲地方的 環境而非魚種(Shewan, 1977)。在很冷、
清淨的水域捕獲魚的菌量較少,溫暖水 域捕獲者較多,若菌量非常高而達到 107 cfu/cm2,這常是來自汙染水域的魚。魚 體表面存在許多不同的細菌種類,在溫 帶水域魚的細菌根據其生長溫度範圍,
分 類 為 耐 冷 菌 (psychrotrophs) 或 嗜 冷 菌 (psychrophiles) 。 耐 冷 菌 能 夠 在 0°C 成 長,但 25°C 左右最適合,嗜冷菌其最高
表 9、冷藏魚中自家消化的變化整理
酵素 基質 遭遇的變化 預防/抑制
醣解酵素 (glycolytic
enzymes) 肝醣
乳酸產生,組織 pH 降低, 肌肉的 保水力降低;
溫度高時,硬直會造成龜裂(gaping)
實際上盡可能將魚在 0°C 附近的溫度通過硬直期;
須避免硬直前的緊迫 自家消化酵素(autolytic
enzymes),涉及核苷酸的 降解)
ATP ADP AMP IMP
新鮮魚風味消失,次黃嘌呤的苦味逐 漸產生(後期階段)
同上
處置不當或擠壓會加速裂 解
組織蛋白酶 (cathepsins) 蛋白質
胜肽 組織軟化而無法或不易加工處理 在貯藏期間與卸貨時處置
粗暴 胰蛋白酶(trypsin)、胰凝乳
蛋白酶(chymotrypsin)、羧 基肽酶(carboxypeptidases)
蛋白質 胜肽
中上層魚內臟腔的自家消化(腹部破 裂)
凍結/解凍或長時間冷藏 會加重
鈣蛋白酶(calpain) 肌原纖維蛋
白質 軟化、甲殼類之脫殼誘導的軟化 移除鈣可避免酵素活化?
膠原蛋白酶(collagenases) 結締組織 肉片龜裂(gaping) 軟化
結締組織降解和冷藏時間 及溫度有關
氧化三甲胺脫甲基酶 (TMAO demethylase)
氧化三甲胺 (TMAO)
鱈科魚類的甲醛誘導的韌化 (toughening)
魚貯藏溫度≦ -30°C 物理性損傷與冷凍/解凍 促進甲醛誘導的軟化
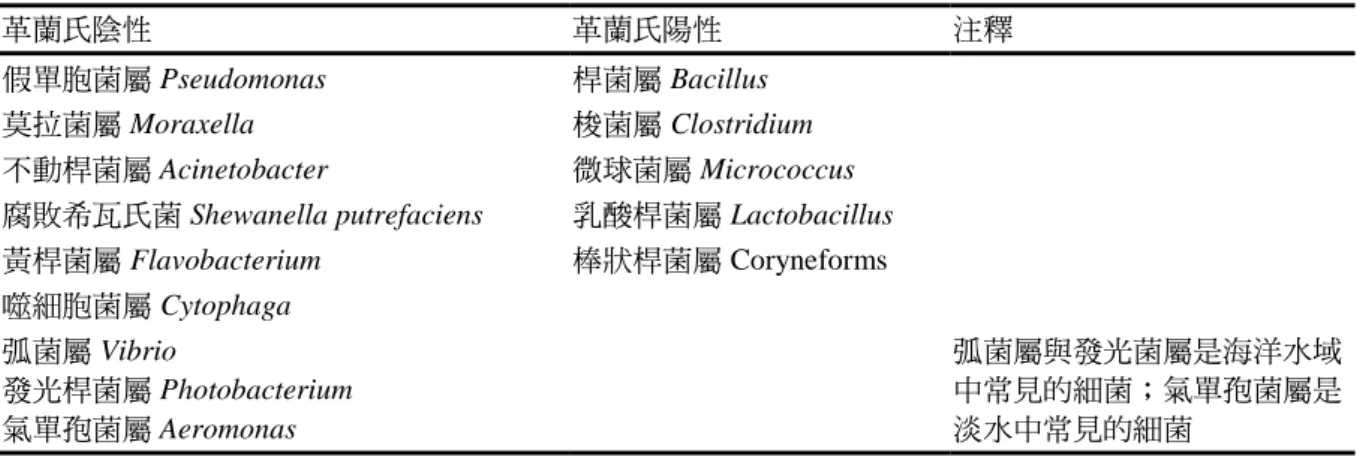
成長溫度 20°C 附近,最適溫度為 15°C (Morita, 1975)。在較溫暖的水域,可分離 出高數量的嗜中溫菌(mesophiles),在溫 帶水域魚類的細菌相以耐冷性、革蘭氏 陰 性 的 桿 狀 細 菌 (rodshaped bacteria) 為 主,包括假單胞菌屬(Pseudomonas)、莫 拉 菌 屬 (Moraxella) 、 不 動 桿 菌 屬 (Acinetobacter)、希瓦氏菌屬(Shewanella) 和 黃 桿 菌 屬 (Flavobacterium) 。 弧 菌 科 (Vibrionaceae)成員的 弧菌屬(Vibrio)與發 光桿菌屬(Photobacterium)、氣單胞菌科 (Aeromonadaceae) ( 氣 單 孢 菌 屬 Aeromonas spp.)也是常見的水生細菌和典 型的魚類的菌群(表 10)。革蘭氏陽性菌如 桿 菌 屬 (Bacillus) 、 微 球 菌 屬 (Micrococcus) 、梭菌 屬(Clostridium)、乳
酸 桿 菌 屬 (Lactobacillus) 和 棒 狀 桿 菌 (Coryneforms)也可能 以 不同比率 存在 , 但 通 常 菌 相 仍 是 革 蘭 氏 陰 性 菌 為 主 。
Shewan (1977)結論:革蘭氏陽性桿菌屬
與微球菌屬是熱帶水域魚類的優勢菌,
但此結論受到質疑。後來的研究指出:
熱帶魚種和溫帶種類的菌相很類似(Acuff
et al., 1984;
Gram et al., 1990; Lima dos Santos 1978; Surendran et al., 1989)。印度 學者的研究指出,剛捕獲魚的菌相包含 假單胞菌屬、不動桿菌屬、莫拉菌屬和 弧菌屬,這和 Liston (1980)的結論相同,熱帶魚種的菌相常含較多的革蘭氏陰性 與腸道細菌(enteric bacteria),但其餘的則 相似於溫帶魚種的菌相。
表 10. 乾淨無汙染水域捕獲魚類的細菌相
革蘭氏陰性 革蘭氏陽性 注釋
假單胞菌屬 Pseudomonas 桿菌屬 Bacillus
莫拉菌屬 Moraxella 梭菌屬 Clostridium
不動桿菌屬 Acinetobacter 微球菌屬 Micrococcus
腐敗希瓦氏菌 Shewanella putrefaciens 乳酸桿菌屬 Lactobacillus
黃桿菌屬 Flavobacterium 棒狀桿菌屬 Coryneforms
噬細胞菌屬 Cytophaga 弧菌屬 Vibrio
發光桿菌屬 Photobacterium 氣單孢菌屬 Aeromonas
弧菌屬與發光菌屬是海洋水域 中常見的細菌;氣單孢菌屬是 淡水中常見的細菌
氣單孢菌屬(Aeromonas spp.)為淡水 魚的典型細菌,海洋水域的典型細菌是 成長需要鈉的一些細菌,包括弧菌屬、
發光桿菌屬和希瓦氏菌屬,但是腐敗希 瓦氏菌(Shewanella putrefaciens)的特性雖 也是鈉需求型,其菌株從淡水環境也分 離出(DiChristina and DeLong, 1993; Gram
et al., 1990; Spanggaard et al., 1993)。雖熱
帶海域也可分離出腐敗希瓦氏菌,對淡 水 魚 的 腐 敗 , 其 重 要 性 低 (Lima dos Santos, 1978; Gram, 1990)。汙染水域中可發現高數量的腸道細 菌科(Enterobacteriaceae),這些生物在清 淨的溫帶水域很快消失,但有報告指出 大 腸 桿 菌 (Escherichia coli) 和 沙 門 氏 菌 (Salmonella)能夠在熱帶水域存活很長的 時 間 , 一 旦 導 入 後 幾 乎 變 為 土 生 種 (Fujioka et al., 1988)。
腐敗希瓦氏菌(S. putrefaciens)的分類 相 當 混 淆 。 最 初 和 無 色 桿 菌 屬 群 (Achromobacter group)有關,後來列在舒 旺假單孢菌屬群 IV (Shewan Pseudomonas group IV)。根據鳥嘌呤(guanine)+胞嘧啶
(cytosine)比率(GC%),又被轉入交替單胞 菌屬(Alteromonas),但基於 5SRNA 同源 性,又再分類成為一個新屬,即希瓦氏 菌 屬 (Shewanella) (MacDonnell and Colwell, 1985) 。 新 近 又 建 議 弧 菌 科 (Vibrionaceae)家族的 一員氣單孢菌屬轉 入 氣 單 胞 菌 科 (Aeromonadaceae) 的 家 族 (Colwell et al., 1986)。
日本學者的研究,顯示魚腸胃道中 含有很高量的微生物,比周圍水域高出 甚多,這表明腸胃道存在著對微生物有 利的生態區位(niche)。Larsen et al. (1978) 報 告 鱈 魚 (cod) 的 腸 道 中 , 含 高 達 107 cfu/g 的類似弧菌(vibrio-like)的微生物,
Westerdahl et al. (1991)也從大菱鮃(turbot) 的 腸 道 分 離 出 高 量 的 類 似 弧 菌 的 微 生 物。從魚表面分離得到的磷光發光菌(P.
phosphoreum)也能從一些魚種的腸道中分 離出高菌量(Dalgaard, 1993),相反的,一 些研究者相信腸胃道的菌群只是環境和 食物攝食的一種反映。
2. 微生物的侵入