The effect of NaCl on proline accumulation in rice leaves
Chuan Chi Lin, Yi Ting Hsu and Ching Huei Kao*
Department of Agronomy, National Taiwan University, Taipei, Taiwan, Republic of China; *Author for correspondence (e-mail: [email protected]; phone: 886-02-2369-8159; fax: 886-02-2362-0879) Received 27 February 2001; accepted in revised form 17 July 2001
Key words: Ammonium, NaCl, Oryza sativa, Proline accumulation
Abstract
The regulation of proline accumulation in detached leaves of rice (Oryza sativa cv. Taichung Native 1) was in-vestigated. Increasing concentrations of NaCl from 50 to 200 mM progressively increased proline content in detached rice leaves. NaCl induced proline accumulation was mainly due to the effect of both Na+and Cl−ions. Proline accumulation caused by NaCl was related to protein proteolysis, an increase in ornithine-␦ -aminotrans-ferase activity, a decrease in proline dehydrogenase activity, a decrease in proline utilisation, and an increase in the content of the precursors of proline biosynthesis, ornithine and arginine. Results also show that proline ac-cumulation caused by NaCl was associated with ammonium ion acac-cumulation.
Abbreviations: DIDS – 4,4⬘-diisothiocyano-2,2⬘-disulfonic acid, FW – fresh weight, GS – glutamine synthetase,
OAT – ornithine-␦-aminotransferase, P5C –⌬1-pyrroline-5-carboxylate, P5CR –⌬1-pyrroline-5-carboxylate re-ductase, PDH – proline dehydrogenase, RWC – relative water content
Introduction
Plants are exposed to various types of environmental stress. Among these stresses, osmotic stress, in par-ticular that due to drought and salinity, is the most serious problem that limits plant growth and crop pro-ductivity in agriculture (Boyer 1982). Proline accu-mulation in plant cells exposed to salt or water stress is a widespread phenomenon (La Rosa et al. 1991; Lutts et al. 1999; Madan et al. 1995; Mattioni et al. 1997; Moftah and Michel 1987; Sudhakar et al. 1993; Treichel 1986; Yoshiba et al. 1997). However, the ac-tual role of proline accumulation remains unclear (Rhodes et al. 1999), but it has been speculated that it can serve as an osmotic regulator (Pollard and Wyn Jones 1979), a protector of enzyme denaturation (Pa-leg et al. 1984), a stabiliser of some macromolecules or molecular assemblies (Schwab and Gaff 1990), a reservoir of nitrogen and carbon sources (Fukutaku and Yamada 1984) or a hydroxyl radical scavenger (Smirnoff and Cumbes 1989). However, some reports indicate no correlation between proline accumulation
and stress resistance (Bhaskaran et al. 1985; Garcia et al. 1997; Lutts et al. 1996; Moftah and Michel 1987; Tully et al. 1979).
Proline accumulation in plant tissues has been sug-gested to result from (a) a decrease in proline degra-dation, (b) an increase in proline biosynthesis, (c) a decrease in protein synthesis or proline utilization, and (d) hydrolysis of proteins (Charest and Phan 1990; Yoshiba et al. 1997). In plants, proline is syn-thesised from glutamic acid (Glu) via⌬1 -pyrroline-5-carboxylate (P5C) by two enzymes, P5C synthetase and P5C reductase (P5CR, EC 1.5.1.2). It has been shown from labelling experiments that ornithine (Orn) can also serve as a precursor to proline biosyn-thesis in higher plants (Brown and Fowden 1966; Chiang and Dandekar 1995). The isolation of cDNA encoding ornithine-␦-aminotransferase (OAT, EC 2.6.1.13) in higher plants (Delauney and Verma 1993; Roosens et al. 1998) suggests that OAT participates in proline biosynthesis by producing P5C from orni-thine and␣-ketoglutarate. Arginine can also contrib-ute to proline biosynthesis, and the pathway from © 2002 Kluwer Academic Publishers. Printed in the Netherlands.
arginine proceeds via ornithine as a result of catalytic activity of arginase (Brown and Fowden 1966; Ling-nowski and Splittstoesser 1971). Proline is metabo-lised to glutamic acid via P5C by two enzymes, pro-line dehydrogenase (PDH, EC 1.5.1.2) and P5C de-hydrogenase (EC 1.5.1.12) (Yoshiba et al. 1997).
It has been shown that proline accumulation in re-sponse to NaCl could be attributed to an increase in P5CR activity (Madan et al. 1995; Mattioni et al. 1997; Sudhakar et al. 1993), an increase in OAT ac-tivity (Lutts et al. 1996; Madan et al. 1995) or a de-crease in PDH activity (Mattioni et al. 1997; Sudhakar et al. 1993). Recently, we have shown that a decrease in proline utilisation might contribute to dark- and water stress-induced proline accumulation in detached rice leaves (Yang et al. 1999, 2000). However, there is no information concerning the ef-fect of NaCl on proline utilisation. Little is known about whether the contents of three amino acids (glutamic acid, ornithine and arginine) involved in the proline biosynthetic pathway are limiting factors for proline accumulation in plant tissues. In a recent work, Lutts et al. (1999) provided evidence to show that proline accumulation was associated with an in-creased production of glutamic acid. In this paper, we report the results of investigations into the regulation of proline accumulation in detached rice leaves ex-posed to NaCl.
Materials and methods Plant material
Rice (Oryza sativa L., cv. Taichung Native 1) was cultured as previously described (Lin et al. 1999). Briefly, rice seedlings were planted on a stainless net floating on half-strength Johnson’s modified nutrient solution (Johnson et al. 1957) in a 500 ml beaker. The nutrient solution (pH 4.8) was replaced every three days. Rice plants were grown for 12 days in a green-house, where natural light was provided and the tem-perature was controlled at 30 °C during the day and at 25 °C at night. The apical 3 cm of the third leaves was used for the experiment. A group of 10 segments was floated in a Petri dish containing 10 ml of test solution dissolved in 5 mM sodium phosphate buffer at pH 7.0. Unless otherwise indicated, incubation was carried out at 27 °C for 3 days in the light (40
mol m−2s−1).
RWC, Na+and Cl−measurements
RWC, defined as water content of leaf tissue as a per-centage that of the fully turgid tissue, was determined by the method of Weatherley (1950). For Na+determination, harvested leaf segments were washed three times (with each one minute) with dis-tilled water, dried at 65 °C for 2 days, extracted in 1 N HCl at room temperature (Hunt 1982) and analy-sed with a flame photometer (Evans, Electroselenium LTD, England). Chloride was estimated in a separate extract made according to the method described by Hodson et al. (1985) and estimated using an ion meter (Mittler Delta 350, UK). The contents of Na+
and Cl− are expressed on the basis of fresh weight (FW).
Determinations of proline and other amino acids
Proline was extracted and its concentration deter-mined by the method of Bates et al. (1973). Leaf seg-ments were homogenised with 3% sulfosalicylic acid and the homogenate was centrifuged at 3,000 g for 20 min. The supernatant was treated with acetic acid and acid ninhydrin, boiled for 1 h and then absor-bance at 520 nm was determined. Contents of proline are expressed asmol g−1FW. For determination of glutamic acid, glutamine, arginine, ornithine, and to-tal amino acids, leaf samples were extracted with 2% sulfosalicylic acid and the homogenate was centri-fuged at 15,000 g for 20 min. The supernatant was used directly for amino acid analysis. Amino acid analysis was carried out by an amino acid analyser (Beckman 6300, California, USA) and contents of amino acids are expressed as nmol g−1FW ormol g−1
FW. For protein determination, leaf segments were homogenised in 50 mM sodium phosphate buffer (pH 6.8). The extracts were centrifuged at 17,600 g for 20 min, and the supernatants were used for determination of protein by the method of Brad-ford (1976). Protein content is expressed as mg g−1 FW.
Enzyme assays
A similar extraction procedure was used for P5CR and PDH, which is mainly based on the procedure described by Lutts et al. (1999). Detached rice leaves were homogenised in a prechilled mortar and pestle with 50 mM Tris-HCl buffer (pH 7.4) containing 7 mM MgCl2, 0.6 M KCl and 3 mM EDTA. The ho-mogenate was centrifuged at 15,000 g for 20 min. The
supernatant was desalted by a Sephadex G-25 column before the assay of P5CR and PDH. The solution used for extraction of OAT was 100 mM potassium phos-phate buffer (pH 7.4) containing 1 mM pyridoxal-5⬘-phosphate, 1 mM EDTA and 10 mM 2-mercaptoetha-nol. The extract was centrifuged at 12,000 g. The supernatant was desalted by a Sephadex G-25 column before the assay of OAT. All the extraction procedures were conducted at 4 °C.
P5CR was assayed by a NADH dependent P5CR reaction (Madan et al. 1995). The assay mixture con-tained 0.06 mM NADH, 0.15 mM P5C, 120 mM po-tassium phosphate buffer, 2 mM dithiothreitol, and the enzyme extract. The reaction was started by the addition of P5C and the decrease in absorbance was followed at 340 nm. P5CR is expressed as units g−1 FW (one unit is defined as a decrease in 1 A340 per min) or units mg−1protein (one unit is defined as a decrease in 0.01 A340per min). OAT activity was sayed according to Vogel and Kopac (1960). The as-say mixture contained 0.2 ml enzyme extract and 0.8 ml 100 mM potassium phosphate buffer (pH 8.0) con-taining 50 mM L-ornithine, 20 mM ␣-ketoglutarate and 1 mM pyridoxal-5⬘-phosphate. The reaction me-dium was incubated at 37 °C for 30 min. The reac-tion was stopped by adding 0.5 ml trichloroacetic acid (10%) and the colour was developed by incubating the reaction mixture with 0.5 ml o-aminobenzalde-hyde (0.5%) in ethanol (95%) for 1 h. After centri-fugation at 12,000 g for 10 min, the clear supernatant fraction was collected to measure the absorbance at 440 nm. OAT is expressed as units g−1FW (one unit is defined as an increase in 1 A440per h) or units mg
−1
protein (one unit is defined as an increase in 0.01 A440 per h). PDH was assayed by following the NAD+
re-duction at 340 nm in a 0.15 M Na2CO3-HCl buffer (pH 10.3) containing 13 mM L-proline and 1.5 mM NAD+
(Lutts et al. 1999). PDH is expressed as units g−1
FW (one unit is defined as an increase in 1 A340 per min) or units mg−1protein (one unit is defined as an increase in 0.01 A340per min).
For extraction of GS (EC 6.3.1.2), leaf segments were homogenised with 10 mM Tris-HCl buffer (pH 7.6, containing 1 mM MgCl2, 1 mM EDTA and 1 mM 2-mercaptoethanol) using a prechilled pestle and mor-tar. The homogenate was centrifuged at 15,000 g for 30 min and the resulting supernatant was used for de-termination in GS activity. GS was assayed by the method of Oaks et al. (1980). The reaction mixture contained in a final volume of 1 ml was 80 mol Tris-HCl buffer, 40 mol L-glutamic acid, 8 mol
ATP, 24mol MgSO4, and 16mol NH2OH; the fi-nal pH was 8.0. The reaction was started by addition of the enzyme extract and, after incubation for 30 min at 30 °C, was stopped by adding 2 ml 2.5% (w/v) FeCl3 and 5% (w/v) trichloroacetic acid in 1.5 M HCl. After centrifugation the absorbance of the super-natant was read at 540 nm. GS is expressed as units per g−1FW (1 unit is defined as 1mol L-glutamate °C-monohydroxamate formed per min).
Determination of proline utilisation
For proline utilisation, detached rice leaves were pre-treated with 50 mM ornithine for 3 h [since addition of ornithine has been observed to be more effective than that of glutamic acid or arginine in increasing proline content in rice leaves (Yang et al. 1999)] to increase the endogenous proline content and then transferred to distilled water and 100 mM NaCl for 8 h in the light. Proline content was then determined. A decline in proline content is considered to imply that proline is utilised (Yang et al. 1999).
Determination of ammonium ions
Ammonium ions were extracted by homogenising leaf segments in 0.3 mM sulphuric acid (pH 3.5). The homogenate was centrifuged for 10 min at 39,000 g and the supernatant was used for determination of ammonium as described previously (Lin and Kao 1998).
Experimental design
For all measurements, each treatment was repeated four times. All experiments described here were re-peated at least four times. Similar results and identi-cal trends were obtained each time. The data reported here are from a single experiment.
Results and discussion
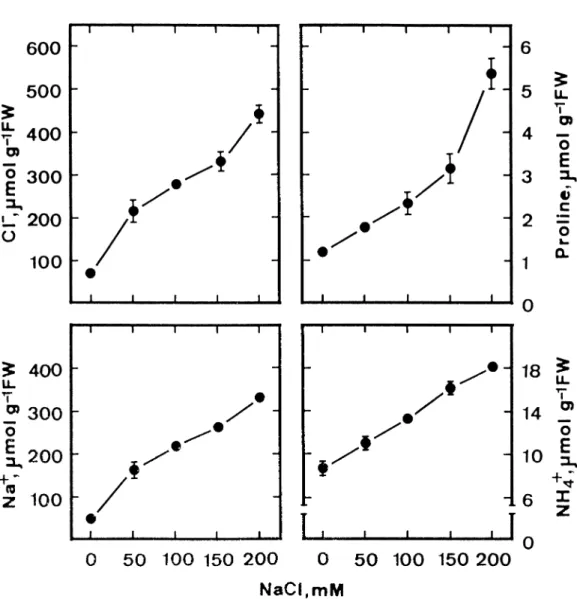
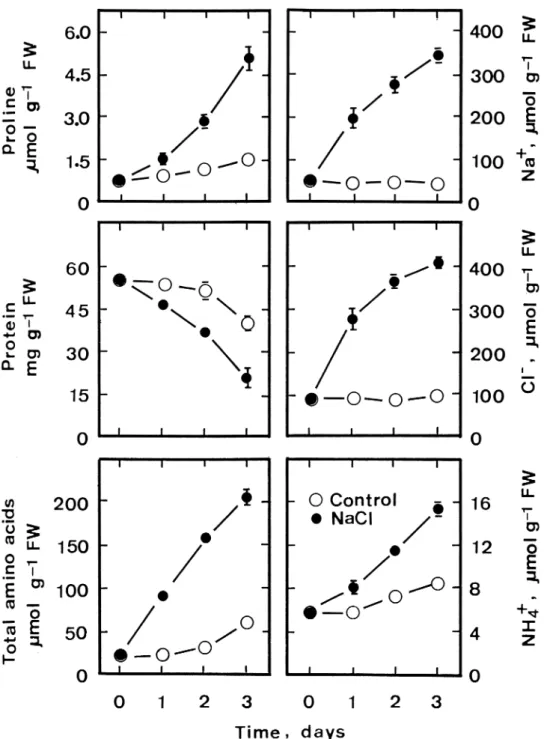
Proline content in detached rice leaves increased with the increase of NaCl concentrations (Figure 1). Pro-line content increased about 4-fold in detached rice leaves treated with 200 mM NaCl for 3 days in the light (Figures 1, 2 and 3), but there was only a slight increase in proline contents in control leaves (Fig-ure 2). Proline contents in detached leaves treated with 200 mM NaCl increased significantly with the
increase of incubation time (Figure 2). To be sure that the described proline accumulation was related to leaf Na+and Cl− contents, both Na+ and Cl− concentra-tions were determined in detached rice leaves treated with various concentrations of NaCl. Both Na+ and Cl−contents in NaCl-treated detached rice leaves in-creased with increasing concentrations of NaCl (Fig-ure 1) but Na+and Cl−contents remained unchanged in control leaves (Figure 2). However, Na+ and Cl− contents in detached rice leaves treated with 200 mM NaCl significantly increased with the increase of in-cubation time (Figure 2).
The effect of NaCl on proline accumulation in de-tached rice leaves could be attributed to Na+
, Cl− or both. Previously, we have reported that NaCl
treat-ment resulted in an accumulation of proline in roots of etiolated rice seedlings (Lin and Kao 1996). We also observed that NaCl-induced proline accumula-tion in rice roots of etiolated seedlings was mainly due to Na+, rather than Cl−(unpublished data). Thus, it is of great interest to know whether proline accu-mulation caused by NaCl in detached rice leaves is also due to Na+, rather than Cl−. To test this possibil-ity, we determined the effect of 4,4⬘-diisothiocyano-2,2⬘-disulfonic acid (DIDS), a nonpermeating amino-reactive disulfonic acid known to inhibit the uptake of Cl−
(Lin 1991), on NaCl-induced proline accumu-lation in detached rice leaves. If Cl−
plays no role in proline accumulation in detached rice leaves treated with NaCl, then addition of DIDS is expected to have
Figure 1. Effect of NaCl on Na+, Cl−, proline, and NH 4
+contents in detached rice leaves. Detached rice leaves were incubated in 5 mM sodium phosphate buffer (pH 7.0) in the presence of NaCl (0−200 mM). Measurements were made after 3 days of treatment in the light. Vertical bars represent standard errors (n = 4).
no effect on proline accumulation. Results (Figure 3) show that DIDS decreased Cl−
content without affect-ing Na+
content, and also decreased proline content in NaCl-treated detached rice leaves. Thus, it seems that both Na+and Cl−are involved in the increase in proline content induced by NaCl in detached rice
leaves. No or only slight difference in RWC was ob-served between NaCl-treated leaves and control leaves (Table 1), suggesting that the osmotic effect is unlikely to be a major factor contributing to the ac-cumulation of proline in detached rice leaves treated with NaCl. This suggestion is supported further by the
Figure 2. Time courses of the NaCl effect on Na+, Cl−, protein, proline, total amino acid, and NH 4
+contents in detached rice leaves. De-tached rice leaves were incubated in 5 mM sodium phosphate buffer (pH 7.0) in the presence or absence of NaCl (200 mM) in the light. Vertical bars represent standard errors (n = 4).
observations that detached rice leaves treated with sorbitol at the concentration iso-osmotic with 200
mM NaCl had much lower proline contents than those treated with 200 mM NaCl and significant proline ac-cumulation was mainly due to the presence of NaCl (Table 2). The fact that NaCl has no effect on RWC is most likely due to a certain amount of osmotic ad-justment caused by the higher contents of Na+ and Cl− in NaCl-treated detached rice leaves (Figure 2).
The ammonium ions are a central intermediate in the metabolism of nitrogen in plants. Ammonium ions have been shown to accumulate in detached rice leaves subjected to water stress (Lin and Kao 1998). Proline accumulation is often considered to be in-volved in stress resistance mechanisms, although its precise role continues to be controversial (Hare et al. 1999; Rhodes et al. 1999; Yoshiba et al. 1997). Based on data in the literature, Rabe (1990) postulated that any stress condition causing reduced growth or im-paired plant health will result in ammonium ion ac-cumulation early in the stress period and suggested that the detoxification process in which excess ammo-nium ions in the cells is removed results in the accu-mulation of nitrogen-containing compounds, such as putrescine and proline. In recent studies, we found that exogenous NH4Cl and methionine sulfoximine,
Figure 3. Effect of NaCl and 4,4⬘-diisothiocyano-2,2⬘-disulfonic
acid (DIDS) on Na+, Cl-,and proline contents in detached rice leaves. Detached rice leaves were incubated in 5 mM sodium phos-phate buffer (pH 7.0) in the presence or absence of NaCl (200 mM). Measurements were made after 3 days of treatment in the light. Vertical bars represent standard errors (n = 4).
Table 1. Effect of NaCl on relative water content (RWC) in
de-tached rice leaves
Time (days) RWC (%) Control NaCl 0 99.8 ± 4.0 1 98.2 ± 2.7 96.0 ± 2.1 2 98.4 ± 2.7 94.4 ± 2.1 3 96.6 ± 2.9 91.2 ± 3.2
Detached rice leaves were incubated in sodium phosphate buffer (5 mM, pH 7.0) in the presence or absence of NaCl (200 mM). Means ± S.E. (n = 4).
Table 2. Effect of the concentration of NaCl and sorbitol on the
proline content in detached rice leaves
Treatment Proline,mol g−1FW
Control 1.06 ± 0.03
Sorbitol, 400 mM 1.53 ± 015 Sorbitol, 300 mM + NaCl, 50 mM 1.98 ± 0.05 Sorbitol, 200 mM + NaCl, 100mM 3.40 ± 0.64 NaCl, 200 mM 4.88 ± 0.56
The osmotic potential was kept equivalent to that of medium with 200 mM NaCl by replacing NaCl by sorbitol. Sorbitol and NaCl wer dissolved in sodium phosphate buffer (5 mM, pH 7.0). Proline was determined after 3 days of treatment in the light. Means ± S.E. (n = 4).
which caused an accumulation of ammonium ions in detached rice leaves, increased proline content (Yang and Kao 1999). In the present investigation, we also found that NaCl treatment resulted in an ammonium ion accumulation (Figures 1 and 2).
The decrease in protein content was faster in NaCl-stressed detached rice leaves than in control leaves (Figure 2). Therefore, protein degradation might contribute to NaCl-induced proline accumula-tion in detached rice leaves. This suggesaccumula-tion is sup-ported further by the observation that total amino ac-ids were higher in detached rice leaves exposed to NaCl than control leaves (Figure 2). However, Lutts et al. (1999) found that proline accumulation in rice (cv. I Kong Pao) plants was not related to proteoly-sis. It is generally considered that glutamic acid, or-nithine and arginine can all contribute to the accumu-lation of proline (Chiang and Dandekar 1995). The results of Figure 4 show that NaCl treatment resulted in an increase in glutamic acid, ornithine and argin-ine contents in detached rice leaves. The increase in glutamic acid, ornithine and arginine contents in NaCl-treated rice leaves is most likely to be a result of protein degradation.
Glutamine synthetase (GS) catalyses the conver-sion of ammonium ion and glutamic acid to glutamine (Ireland and Lea 1999). Figure 5 shows that NaCl markedly decreased GS activity in detached rice leaves. Thus, the possibility that the increase in am-monium ion and glutamic acid contents (Figures 1, 2 and 4) in detached rice leaves treated with NaCl is due to NaCl-inhibited GS activity cannot be excluded. To determine the role of the biosynthetic pathways for proline accumulation caused by NaCl, the effect of NaCl on OAT and P5CR activities was examined. The results are shown in Figure 6. OAT activity, ex-pressed either on per g FW or per mg protein basis, was higher in NaCl-treated leaves than in control leaves. However, P5CR activity in NaCl-treated leaves was lower than that in control leaves (Fig-ure 6). Thus, the increase in OAT activity may have, to some extent, contributed to the elevated content of proline by NaCl. An increase in OAT activity along with an increase in the content of proline in plants under salt stress has been reported (Lutts et al. 1999; Madan et al. 1995). The results of the present inves-tigation are consistent with those of La Rosa et al. (1991); Lutts et al. (1999), who reported that elevated accumulation of proline under NaCl stress is not due to increase in P5CR. P5C synthetase is also involved in proline biosynthesis (Yoshiba et al. 1997). Thus,
NaCl could also have an effect on this enzyme. How-ever, for unknown reasons, we failed to detect any P5C synthetase activity in crude extracts. Thus, this enzyme was not considered in the present investiga-tion. The enzyme PDH is reported to catalyze proline oxidation (Yoshiba et al. 1997). In the present
inves-Figure 4. Time courses of the NaCl effect on glutamic acid (Glu),
ornithine (Orn), and arginine (Arg) contents in detached rice leaves. Detached rice leaves were incubated in 5 mM sodium phosphate buffer (pH 7.0) in the presence or absence of NaCl (200 mM) in the light. Vertical bars represent standard errors (n = 4).
tigation, NaCl treatment resulted in a decrease in PDH activity in detached rice leaves (Figure 6). This result suggests that proline oxidation (or degradation) contributes to proline accumulation in detached rice leaves exposed to NaCl. Sudhakar et al. (1993) also reported that PDH was inhibited to a greater extent in green gram seedlings under salt stress.
Recently, we demonstrated that proline in detached rice leaves under dark conditions and exposed to wa-ter stress is utilised less than in the control treatment (Yang et al. 1999, 2000). It is possible that less utili-sation of proline may also contribute to the accumu-lation of proline in detached rice leaves caused by NaCl. To test this possibility, detached rice leaves were pretreated with ornithine for 3 h in the light to increase endogenous proline content and then trans-ferred to phosphate buffer in the presence or absence of 200 mM NaCl for 8 h in the light; proline contents were then determined. As indicated in Table 3, the proline content was lower in the absence of NaCl than in the presence of NaCl, suggesting that proline in detached rice leaves treated with NaCl is utilised less than in the absence of NaCl.
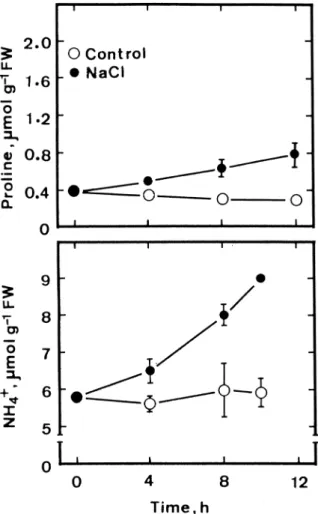
Of particular interest in the reported investigation is the finding that both proline and ammonium ion accumulations were observed in detached rice leaves treated with NaCl (Figures 1 and 2). To test the causal relationship between ammonium ion accumulation
and proline accumulation in detached rice leaves caused by NaCl, a short term experiment was con-ducted. The results are shown in Figure 7. The accu-mulation of ammonium ions caused by NaCl was ev-ident at 4 h after treatment. Proline content decreased slightly in control leaves during the first 10 h of in-cubation in the light. Significant proline accumulation in detached rice leaves was observed at 8 h after NaCl treatment. It is obvious that ammonium ion accumu-lation preceded proline accumuaccumu-lation in NaCl-treated detached rice leaves, suggesting that ammonium ion accumulation is involved in regulating proline accu-mulation during NaCl stress. This suggestion is sup-ported further by the observations that (a) NH4Cl, similar to NaCl treatment, resulted in an increase in OAT activity and an inhibition in PDH activity in de-tached rice leaves (Lin and Kao 2001) and (b) pro-line in NH4Cl- or methionine sulfoximine-treated rice leaves is less utilised than control leaves (Yang et al. 1999), a finding in the same way that NaCl did (Ta-ble 3).
In conclusion, our current results suggest that pro-line accumulation in detached rice leaves caused by NaCl is related to protein hydrolysis, an increase in OAT activity, a decrease in PDH activity, and a de-crease in proline utilisation. Evidence is also provided to show that ammonium accumulation is involved in
Figure 5. (A)Effect of NaCl concentrations on glutamine synthetase (GS) activity in detached rice leaves; (B) Changes in GS activity in
detached rice leaves treated with 200 mM NaCl. GS activity in (A) was determined after 3 days of treatment in the light. Detached rice leaves were incubated in 5 mM sodium phosphate buffer (pH 7.0) in the presence of absence of NaCl. Vertical bars represent standard errors (n = 4).
Figure 6. Time courses of the NaCl effect on the activities or specific activities of⌬1-pyrroline-5-carboxylate reductase (P5CR), ornithine-␦-aminotransferase (OAT), and proline dehydrogenase (PDH) in detached rice leaves. Detached rice leaves were incubated in 50 mM sodium phosphate buffer (pH 7.0) in the presence or absence of NaCl (200 mM) in the light. Vertical bars represent standard errors (n = 4).
proline accumulation in detached rice leaves exposed to NaCl.
It has been shown that NaCl-inhibited root growth of etiolated rice seedlings is associated with proline accumulation and exogenously applied proline
inhib-its root growth (Lin and Kao 1996). The physiologi-cal significance of proline accumulation in intact rice leaves is not fully understood. Studying the effect of NaCl stress on enzyme activities involved in proline metabolism in intact rice leaves could provide valu-able information on the physiological significance of its accumulation. Regulation of proline accumulation in detached rice leaves under NaCl stress as we re-ported here is not necessarily similar to that in intact rice leaves. However, the results of the present work do provide some basic information which should be valuable for our future studies.
Acknowledgements
This work was supported by the National Science Council of the Republic of China. Amino acid analy-ser was kindly provided by NTU Instrumentation Center, Republic of China.
References
Bates L.S., Waldren S.P. and Teare I.D. 1973. Rapid determination of free proline for water-stress studies. Plant Soil. 39: 205–207. Bhaskaran S., Smith R.H. and Newton R.J. 1985. Physiological changes in cultured sorghum cells in response to induced water stress I. Free proline. Plant Physiol. 79: 266–269.
Boyer J.S. 1982. Plant productivity and environment. Science 218: 443–448.
Bradford M.M. 1976. A rapid and sensitive method for the quan-titation of microgram quantities of protein utilizing the princi-ple of protein-dye binding. Anal. Biochem. 72: 248–254. Brown D.H. and Fowden L. 1966. Metabolism of␦-acetylornithine
in two Leguminous species. Phytochemistry 5: 887–892. Charest C. and Phan C.T. 1990. Cold acclimation of wheat
(Triti-cum aestivum): Properties of enzymes involved in proline me-tabolism. Physiol. Plant. 80: 159–168.
Chiang H.-H. and Dandekar A.M. 1995. Regulation of proline ac-cumulation in Arabidopsis thaliana (L.) Heynh during devel-opment and in response to desiccation. Plant Cell Environ. 18: 1280–1290.
Delauney A.J. and Verma D.P.S. 1993. Proline biosynthesis and osmoregulation in lants. Plant J. 4: 215–223.
Fukutaku Y. and Yamada Y. 1984. Sources of proline nitrogen in water-stressed soybean (Glycine max). II. Fate of15N-labelled protein. Physiol. Plant. 61: 622–628.
Garcia A.B., de Almeida J., Iyer S., Gerats T., Van Montagu M. and Caplan A.B. 1997. Effect of osmoprotectants upon NaCl stress in rice. Plant Physiol. 115: 159–169.
Hare P.D., Cress W.A. and Van Stadan J. 1999. Proline synthesis and degradation: a model system for eludicating stress-related signal transduction. J. Exp. Bot. 50: 413–434.
Table 3. Proline content in ornithine-pretreated detached rice leaves incubated in sodium phosphate buffer in the presence or ab-sence of NaCl
Treatment Proline,mol g−1FW
H2O, 3 h 0.77 ± 0.17
Ornithine, 3 h 27.13 ± 0.68
Ornithine, 3 h→ Control, 8 h 14.14 ± 1.35 Ornithine, 3 h → NaCl, 8 h 18.27 ± 1.12
Detached rice leaves were pretreated with 50 mM ornnithine for 3 h in the light and then incubated in sodium phosphate buffer (5 mM, pH 7.0) in the presence or absence of NaCl (200 mM) for 8 h in the light. Means ±S.E. (n = 4).
Figure 7. Effect of NaCl on proline and NH4+contents in detached rice leaves. Detached rice leaves were incubated in 5 mM sodium phosphate buffer (pH 7.0) in the presence or absence of NaCl (200 mM) in the light. Vertical bars represent standard errors (n = 4).
Hodson M.J., Opik H. and Wainwright S.J. 1985. Changes in ion and water content of individual shoot organs in a salt-tolerant and a salt-sensitive clone of Agrostis stoloniferra L. during and subsequent to treatment with sodium chloride. Plant Cell Envi-ron. 8: 657–658.
Hunt J. 1982. Dilute hydrochloric acid extraction of plant material for routine cation analysis. Commun. Soil Sci. Plant Analysis 13: 49–55.
Ireland A.J. and Lea P.J. 1999. The enzymes of glutamine, glutamate, aspargine, and aspartate metabolism. In: Singh B.K. (ed.), Plant Amino Acids. Mercel Dekker Inc., New York, pp. 49–109.
Johnson C.M., Stout P.R., Broyer T.C. and Carlton A.B. 1957. Comparative choline requirements of different plant species. Plant Soil. 8: 337–353.
La Rosa P.C., Rhodes D., Rhodes J.C., Bressan R. and Csonka L.N. 1991. Elevated accumulation of proline in NaCl-adapted to-bacco cells is not due to altered⌬1-pyrroline-5-carboxylate re-ductase. Plant Physiol. 96: 245–250.
Lin C.C. and Kao C.H. 1996. Proline accumulation is associated with inhibition of rice seedling root growth caused by NaCl. Plant Sci. 114: 121–128.
Lin C.C. and Kao C.H. 2001. Regulation of ammonium-induced proline accumulation in detached rice leaves. Plant Growth Regul. 35: 69–74.
Lin J.-N. and Kao C.H. 1998. Water stress, ammonium, and leaf senescence in detached rice leaves. Plant Growth Regul. 26: 165–169.
Lin J.-N., Wang J.-W. and Kao C.H. 1999. Effect of abscisic acid and water stress on the senescence of detached rice leaves. Biol. Plant. 42: 313–316.
Lin W. 1991. Inhibition of anion transport in corn root protoplast. Plant Physiol. 68: 435–438.
Lingnowski E.M. and Splittstoesser W.E. 1971. The change in argi-nine levels and metabolism of urea and ornithine in Cucurbita moschatta seedlings. Physiol. Plant. 25: 225–229.
Lutts S., Kinet J.M. and Bouharmont J. 1996. Effects of salt stress on growth, mineral nutrition and proline accumulation in rela-tion to osmotic adjustment in rice (Oryza sativa L.) cultivars differing in salinity resistance. Plant Growth Regul. 91: 207– 218.
Lutts S., Majerus V. and Kinet J.M. 1999. NaCl effects on proline metabolism in rice (Oryza sativa L.) seedlings. Physiol. Plant. 105: 450–458.
Madan S., Nainawatee H.S., Jain R.K. and Chowdhury J.B. 1995. Proline and proline metabolising enzymes in in-vitro selected NaCl-tolerant Brassica juncea L. under salt stress. Ann. Bot. 76: 51–57.
Mattioni C., Lacerenza N.G., Troccoli A., De Leonardis A.M. and Di Fonzo N. 1997. Water and salt stress-induced alterations in proline metabolism of Triticum durum seedlings. Physiol. Plant. 101: 787–792.
Moftah A.F. and Michel B.E. 1987. The effect of sodium chloride on solute potential and proline accumulation in soybean leaves. Plant Physiol. 83: 238–240.
Oaks A., Stulen J., Jones K., Winspear M.J., Misra S. and Boesel I.L. 1980. Enzymes of nitrogen assimilation in maize roots. Planta 148: 477–484.
Paleg L.G., Stewart G.R. and Bradbeer J.W. 1984. Proline and gly-cine betaine influence protein solvation. Plant Physiol. 75: 974– 978.
Pollard A. and Wyn Jones R.G. 1979. Enzyme activities in concen-trated solutions of glycinebetaine and other solutes. Planta 144: 291–298.
Rabe E. 1990. Stress physiology: the functional significance of the accumulation of nitrogen-containing compounds. J. Hort. Sci. 65: 231–243.
Rhodes D., Verslues P.E. and Sharp R.E. 1999. Role of amino ac-ids in abiotic stress resistance. In: Singh B.K. (ed.), Plant Amino Acids. Mercel Dekker Inc., New York, pp. 319–356. Roosens N.H.C.J., Thu T.T., Iskandar H.M. and Jacobs M. 1998.
Isolation of the ornithine-␦-aminotransferase cDNA and effect of salt stress on its expression in Arabidopsis thaliana. Plant Physiol. 117: 263–271.
Schwab K.B. and Gaff D.F. 1990. Influence of compatible solutes on soluble enzymes from desiccation-tolerant Sporobolus stap-fianus and desiccation-sensitive Sporobolus pyramidalis. J. Plant Physiol. 137: 208–215.
Smirnoff N. and Cumbes Q.J. 1989. Hydroxyl radical scavenging activity of compatible solutes. Phytochemistry 28: 1057–1060. Sudhakar C., Reddy P.S. and Veeranjaneyulu K. 1993. Effect of salt stress on the enzymes of proline synthesis and oxidation in greengram (Phaseolus aureus Roxb.) seedlings. J, Plant Phys-iol. 141: 621–623.
Treichel S. 1986. The influence on⌬1-pyrroline-5-carboxylate re-ductase in proline-accumulating cell expansion cultures of Mesembryanthemum nodiflorum and other halophytes. Physiol. Plant. 67: 173–181.
Tully R.E., Hanson A.D. and Nelsen C.H.E. 1979. Proline accu-mulation in water-stressed barley leaves in relation to translo-cation and the nitrogen-budget. Plant Physiol. 63: 518–523. Vogel R.H. and Kopac M.J. 1960. Some properties of ornithine-␦
-transaminase from Neurospora. Biochem. Biphys. Acta 37: 539–540.
Weatherley P.F. 1950. Studies in the water relation of cotton plant I. The field measurement of water deficits in leaves. New Phy-tol. 49: 81–97.
Yang C.-W. and Kao C.H. 1999. Ammonium in relation to proline accumulation in detached rice leaves. Plant Growth Regul. 30: 139–144.
Yang C.-W., Lin C.C. and Kao C.H. 1999. Endogenous ornithine and arginine contents and dark-induced proline accumulation in detached rice leaves. J. Plant Physiol. 155: 665–668. Yang C.-W., Wang J.-W. and Kao C.H. 2000. The relation between
accumulation of abscisic acid and proline in detached rice leaves. Biol. Plant. 43: 301–304.
Yoshiba Y., Kiyosue T., Nakashima K., Yamaguchi-Shinozaki K. and Shinozaki K. 1997. Regulation of levels of proline as an osmolyte in plants under water stress. Plant Cell Physiol. 38: 1095–1102.