國立臺灣大學生物資源暨農學院園藝學系 碩士論文
Department of Horticulture College of Bioresources and Agriculture
National Taiwan University Master Thesis
番椒細胞質雄不稔系統穩定性之研究
The Stability for Cytoplasmic Male Sterile in Pepper
鄞妙真 Miao-Jen Yin
指導教授:林宗賢 博士;楊雯如 博士
Advisor: Tzong-Shyan Lin, Ph. D., and Wen-Ju Yang, Ph. D.
中華民國 九十八 年 二 月
Feb., 2009
i
誌謝
首先感謝恩師 林宗賢老師與楊雯如老師於修業期間的督促與鼓勵,讓我順利完成 碩士學位與論文。林老師您雖曾暫時離開園藝系至台中科博館當館長,仍抽空關 心我的試驗狀況並給予我許多的意見,激盪出更多有趣的想法與觀點,而您對農 業的熱忱與堅持、待人處事的方法及態度,為我努力學習的目標。楊老師您對我 的指導照顧,包含生活上的瑣事、每一次的上台演出、及未來的人生,總是用您 獨特的風格談笑一切,卻也都刺中核心,我總是一直放在心上並反覆思考,成為 我能不斷進步的原因。
感謝亞蔬—世界中心Dr. Gniffke 提供我試驗材料與場地,您總是非常有耐心
地與我溝通並給我最大的協助,感謝林世雯小姐與謝雪琴小姐幫助我對試驗材料 的熟悉,給我許多幫忙及意外的美食。感謝鍾美珠老師給我極大的自由與空間運
用您的設備及論文的修改,每每一起看著顯微鏡一起討論data,都讓我滿載而歸。
感謝李勇毅博士於我試驗之初提供試驗方向,並教導切片技術。感謝黃玲瓏老師 與陳香君老師帶我進入且愛上植物解剖,上妳們的課是種很棒的享受。感謝中研 院植微所簡萬能老師與技術人員、及台大生科院生物影像平台(TC5 Bio- image) 提 供設備及切片技術指導,讓我順利完成試驗。
謝謝大學的同窗好友芳瑀、蓓文、愛媜、祖威、紹益及紹謙,即使大學畢業 後就各自在不同地方打拼,偶爾的相聚總能帶給我滿滿的開心。謝謝一蘆學長、
錦興學長、元聰學長在lab meeting 時提供各式各樣的意見,在中研院的芳綺學姊、
桂瑞學姊及紹安學長的照顧與餵食(哈),還有可愛的嵐雁、孟姿、美蘭、李廉、嘉 彬、少雄、資棟、楓俊、禎陽、本忞、芳怡、恩康跟伊婷,與你們聊天及搞笑為 我的研究生生活中帶來更多的歡樂。
最後,謝謝爸媽給我無限的包容與溫暖,讓我順利完成學業且不需煩惱家裡 經濟,謝謝大哥跟二哥對妹妹的照顧跟疼愛。特別謝謝一蘆學長,是你帶我踏入 真正的農業;謝謝李廉,你的支持與陪伴幫我度過許多難過的日子。謝謝。
ii
Content
誌謝 ……….. i
Content ………. ii
Tables list ………... iv
Figures list ………... v
中文摘要 ……….... ix
Abstract ……… xi
Chapter 1 Introduction ………... 1
Chapter 2 Literature Review ………... 5
2.1 Cytoplasmic male sterility in higher plant ……… 5
2.2 The characters of CMS-related genes ………... 6
2.3 The characters of restorer genes and interaction with CMS-related genes ….. 7
2.4 Phenotypes and abnormalities in tapetum of CMS plants ……… 9
Chapter 3 Genetic Background Identification in Hot and Sweet Pepper Lines …... 11
3.1 Introduction ………. 11
3.2 Materials and methods ……… 14
3.3 Results and discussion ……… 18
Chapter 4 The Influences of Temperature and Seasonal Changes on Cytoplasm Male Sterile Pepper Lines ……… 31
iii
4.1 Introduction ……….... 31
4.2 Materials and methods ……… 33
4.3 Results ……… 35
4.4 Discussion ..……… 38
Appendix 4.1 ……… 51
Appendix 4.2 ……… 52
Chapter 5 The Influences of Temperature on Anther Wall Differentiation and Microgametogenesis of CMS Pepper Lines ………... 53
5.1 Introduction ……… 53
5.2 Materials and methods ……… 55
5.3 Results ………... 57
5.4 Discussion ………... 60
Chapter 6 Conclusion ……….. 72
References ………... 75
iv
Tables list
Table 3.1 The pedigree of CMS, maintainer or near-isogenic lines, and restorer pepper lines derived from AVRDC- The World Vegetable Center. ……… 21 Table 3.2 Primer sets used in this study. ……… 23 Table 3.3 Genomic pepper DNA analysis by ATP-1, ATP-2, ATP-3, CMS P1, CMS P2, and CRF-S primer sets. ……….. 24 Table 4.1 The pedigree of CMS lines and maintainers from AVRDC-The World
Vegetable Center. ………... 44
v
Figures list
Fig. 3.1 PCR amplification of ψatp6-2 fragment from pepper leaf genomic DNA by (A) ATP-1, (B) ATP-2, and (C) ATP-3 primer sets. ……….. 26 Fig. 3.2 PCR amplification of orf456 fragment from pepper leaf genomic DNA by (A)
CMS P1, and (B) CMS P2 primer sets. ……….. 27 Fig. 3.3 PCR amplification of Rf fragment from pepper leaf genomic DNA by CRF-S
primer set. ………... 28 Fig. 3.4 PCR amplification of ψatp6-2 fragment from pepper leaf genomic DNA by ATP 1 primer set. ……… 28 Fig. 3.5 PCR amplification of orf456 fragment from pepper leaf genomic DNA by CMS
P1 primer set. ……….. 29 Fig. 3.6 PCR amplification of Rf fragment from pepper leaf genomic DNA by CRF-S
primer set. ………... 29 Fig. 4.1 CMS and near-isogenic hot pepper plants grown in the AVRDC field. (A) unstable CMS line ‘CCA 7243’; (B) near-isogenic line ‘PBC 385’ of ‘CCA 7243’;
(C) stable CMS line ‘CCA 7244’; (D) near-isogenic line ‘9907-9611’ of ‘CCA 7244’. ……….. 45 Fig. 4.2 CMS and near-isogenic hot pepper flowers harvested from plants in the
vi
AVRDC field. (A)sterile flower of unstable CMS line ‘CCA 7243’(B) restored fertile flower of unstable CMS line ‘CCA 7243’; (C) fertile flower of near-isogenic line ‘PBC 385’; (D) sterile flower of stable CMS line ‘CCA 7244’; (E) fertile flower of near-isogenic line ‘9907-9611’. ……….. 45 Fig. 4.3 Sweet pepper plants grown in the AVRDC field. (A) unstable CMS line ‘CCA
7236’; (B) near-isogenic line ‘PBC 84sel’ of ‘CCA 7236’; (C) stable CMS line
‘CCA 7234’; (D) near-isogenic line ‘9847-4754’ of ‘CCA 7234’. ………. 46 Fig. 4.4 CMS and near-isogenic sweet pepper flowers harvested from plants in the
AVRDC field. (A)sterile flower of unstable CMS line ‘CCA 7236’; (B)fertile flower of near-isogenic line ‘PBC 84sel’; (C) sterile flower of stable CMS line
‘CCA 7234’; (D) fertile flower of near-isogenic line ‘9847-4754’ of ‘CCA 7234’. ……….. 46 Fig. 4.5 The trend of pollen viability percentage in hot pepper lines of (A) ‘CCA 7243’
(B) ‘CCA 7244’ (C) ‘PBC 483’(D) ‘9907-9611’ in the greenhouse (GH) and in the phytotron of NTU at 30/25 oC, 25/20 oC, and 20/15oC from Apr. 29 to Jun. 29, 2007. ………... 47 Fig. 4.6 The trend of pollen viability percentage in hot pepper lines in the field of AVRDC–The World Vegetable Center from December 7 to July. ………. 48
vii
Fig. 4.7 The trend of pollen viability percentage in sweet pepper lines (A) ’CCA 7236’, (B) ‘CCA 7234’, (C) ‘PBC 84 selex’, and (D) ‘9847-4754’ in the greenhouse (GH) and in the phytotron of NTU at 30/25 oC, 25/20 oC,and 20/15oC from Mar. 20 to Jun. 29, 2007. ………... 49 Fig. 4.8 The trend of pollen viability percentage in sweet pepper lines in the field of
AVRDC–The World Vegetable Center from December 7 to July 4. ………….. 50 Fig. 5.1 Semi-thin transverse sections of hot pepper maintainer line ‘PBC 483’ anther
under high temperature. Sections stained by 1% TBO. (A) Primary sporogenous cell stage with periclinal division in endothecium and middle layer; (B) Early pollen mother cell stage; (C) Pollen mother cell stage during prophase I; (D) Tetrad stage; (E) Early released free microspore stage; (F) Mid-vacuolated microspore stage; (G) Late vacuolated pollen stage during nucleus mitosis (arrowed); (H) Mature pollen stage before anther wall dehiscence. ……….. 65 Fig. 5.2 Semi-thin transverse sections of CMS hot pepper line ‘CCA 7244’ anther under
high temperature. Sections stained by 1% TBO. (A) Secondary sporogenous cell stage with anther wall differentiating into 4 distinct layers; (B) Pollen mother cell stage during prophase stage; (C) Tetrad stage; (D) Early released free microspore stage; (E) Relative late free microspore stage; (F) Debris in the locule before anther dehiscence. ………. 67
viii
Fig. 5.3 Semi-thin transverse sections of unstable CMS hot pepper line ‘CCA 7243’
anther under high temperature. Sections stained by 1% TBO. (A) Primary sporogenous cell stage with periclinal division in endothecium and middle layer;
(B) Early pollen mother cell stage; (C) Pollen mother cell stage during prophase I;
(D) Tetrad stage; (E) Mid-vacuolated microspore stage with vacuolated tapetum cells; (F) Mid-vacuolated microspore stage with remained dense cytoplasm in tapetum cells; (G) Tetrad stage with dense cytoplasm in tapetum cells; (H) Early released free microspore stage; (I) Early released free microspore stage pressed; (J) Relative late-vacuolated microspore stage with debris and abnormal microspores in the locule; (K) Debris and abnormal microspores in the locule before anther dehiscence. ………... 69 Fig. 5.4 Semi-thin transverse sections of unstable CMS hot pepper line ‘CCA 7243’
anther under cool temperature. Sections stained by 1% TBO. (A) Primary sporogenous cell stage with periclinal division in endothecium and middle layer;
(B) Early pollen mother cell stage; (C) Tetrad stage; (D) Early released free microspore stage; (E) Mid-vacuolated microspore stage; (F) Late-vacuolated microspore stage; (G) Vacuolated pollen stage during nucleus mitosis; (H)Mature pollen stage before anther wall dehiscence. ………... 70
ix
摘要
細胞質雄不稔 (cytoplasmic male sterility, CMS) 由細胞質與細胞核之間的交 互作用所控制,但某些雄不稔番椒品系並非可以一直維持其雄不稔性,植株受低 溫影響會部分跟完全恢復稔性,當溫度升高則植株會再回復雄不稔性。本試驗的 目的為確認所收集的番椒品系之遺傳背景、溫度及季節變化對雄不稔系之影響、
以及穩定與不穩定雄不稔系之解剖之差異。
所有參試雄不稔番椒品系可藉由引子組,分別擴增出預期之(S)atp6-2 片段,
而其辣椒維持系則無法擴增出預期產物;針對分子標記orf456 亦有相似的結果。
上述結果可顯示所有參試雄不稔番椒之細胞質均含有(S)atp6-2 與 orf456,而該
辣椒維持系因具N 型細胞質,故不會帶有(S)atp6-2 與 orf456。然而,不穩定雄
不稔甜椒之維持系‘PBC 84 selex’ 與‘9946-2138’,可由 CMS P1 或 CMS P2 引子
組擴增出orf456 片段,表示在特定的維持系中,其細胞質之遺傳背景可能較複雜
且異於其他維持系。
不穩定雄不稔辣椒系‘CCA 7243’,於溫室與田間之夜溫低於 21oC 可恢復稔性,
且於20/15oC 之人工氣候室內也有恢復稔性之現象;夜溫低於 21 oC 或 17 oC,另
一不穩定雄不稔甜椒系‘CCA 7236’可恢復稔性,但日溫低於 18oC 時,則該植株不
易恢復稔性。此外,不穩定雄不稔系之稔性恢復並非持續表現,而是呈現規律週 期。兩不穩定雄不稔系夜溫升高時均會回復雄不稔。雄不稔系‘CCA 7244’與‘CCA 7234’於一段長時間的低夜溫環境下,仍幾乎維持其雄不稔性。
不穩定雄不稔系‘CCA 7243’於高溫下表現雄不稔性,其絨氈層細胞之異常極 不同於穩定雄不稔系‘CCA 7244’之絨氈層。‘CCA 7243’之花藥腔室於四分子體形 成時會增大,或者不會;相較於‘CCA 7244’,其花藥腔室於該時期不會擴大。隨 著小孢子細胞發育期間,‘CCA 7243’之絨氈層為濃稠的細胞質或具有一液泡於細
x
胞質中,相較於‘CCA 7244’之絨氈層僅呈現液泡化;此外,絨氈層細胞瓦解之時 間點難以辨識,但兩品系之絨氈層細胞最終會消逝。由上述解剖結果顯示,不穩 定雄不稔系之遺傳調控可能較為複雜。雄不稔不穩定系‘CCA7243’之花藥恢復稔 性,表示其發育構造上與維持系‘PBC 385’無異。
關鍵字:花藥分化、atp6、番椒、細胞質雄不稔、小配子發生、orf 456、溫度
xi
Abstract
Cytoplasmic male sterility (CMS) is the result of interaction between cytoplasm and nuclear genes. The sterility of the CMS pepper lines was not always stable. Some of them might restore their fertility partially or completely due to low temperature, and such restoration could be resumed as temperature re-elevate. The objective of this thesis is to study the genetic background of collected pepper lines, the influences of temperature and seasonal changes on CMS pepper lines, and the anatomy difference between stable and unstable CMS line.
All CMS pepper lines could generate predicted a (S)atp6-2 fragment by primer sets, while their maintainer lines couldn’t produce the predicted products, and similar results were obtained for orf456 fragment. These results revealed that the cytoplasms of all the CMS pepper lines contained (S) atp6-2 and orf456, and their maintainers of hot pepper lines, with N cytoplasm, didn’t have such defected atp6-2 and orf456.
Surprisingly, ‘PBC 84 selex’ and ‘9946-2138’, maintainer lines of unstable CMS sweet pepper, could generate orf456 fragment by CMS P1 or CMS P2 primer sets, representing that the genetic background of cytoplasm in specific maintainers might be more complicated and different from other maintainers.
xii
Unstable CMS hot pepper line ‘CCA 7243’ could restore fertility in the greenhouse and in the field while the night temperature was below 21 oC, and also could restore in the 20/15 oC phytotron. Plants of unstable CMS sweet pepper line
‘CCA 7236’ restored fertility while night temperature was below 21oC or 17 oC, but plants hardly restored while day temperature below 18 oC. Besides, the fertility restoration of unstable CMS lines showed as a regular cycle, not always steady expressed. When night temperature went higher, both lines were reverted male sterile.
Both CMS line ‘CCA 7244’ and ‘CCA 7234’ almost expressed male sterile even under a long period of lower night temperature.
As unstable CMS line ‘CCA7243’ expressed sterility under high temperature, abnormalities in tapetum cells were very different from the results of stable CMS line
‘CCA7244’. While tetrads formed, locule in ‘CCA 7243’ may or may not increase in size as compared with locular space did not increase in ‘CCA 7244’. As microspores development, the appearance of tapetum in ‘CCA 7243’ with dense cytoplasm or containing a vacuole was different from vacuolated tapetum in ‘CCA 7244’, and timing of tapetum cells degeneration was hardly identified, but tapetum cells in both lines were gone at the end. These anatomical data revealed that the genetic control might be more complicated in unstable CMS line. As the anthers of ‘CCA7243’ resumed the
xiii
fertility, there was no difference between ‘CCA7243’ and its maintainer ‘PBC 385’
anatomically.
Keywords: anther differentiation, atp6, Capsicum annuum L., cytoplasmic male sterility (CMS), microgametogenesis, orf 456, temperature
1
Chapter 1 Introduction
Capsicum is a genus of plants from Solanaceae family and native to the tropical
and subtropical Americas. Among approximately 30 species of Capsicum, five major domesticated species are used widely: C. annuum, C. baccatum, C. chinense, C.
frutescens, and C. pubescens. C. annuum, which contains both hot and sweet pepper, is
the most cultivated and economically important species worldwide. Pepper fruits are in a tremendous variety of colors, shapes, sizes, and tastes (Bosland, 1992; Bosland and Votava, 2000; Heiser and Pickersgill, 1969; Smith et al, 1987).
C. annuum is a warm-season crop, and the optimum range of temperature for
plant growth is 21-30 oC. High temperature 33 oC exposure during bell pepper floral bud at < 2.5 mm in length does not affect flower production, but reduced pollen viability and fruit set. In addition, fertilization is also sensitive to high temperature 33
oC, which inhibited fruit set (Erickson and Markhart, 2001; Erickson and Markhart,
2002). Besides, high night or low night temperature also decreased fruit production.
High night temperature at 24 oC caused considerable blossom drop in bell pepper (Rylski and Spigelman, 1982). Low night temperature 10 oC and 15 oC decreased pollen viability (Mercado et al, 1997). As floral bud development under low night temperature 10±2 oC, starch accumulation in pollen grains at 3 days before anthesis
2
and more than two-fold in total soluble sugars in the mature pollen grains were both decreased (Pressman et al, 2006).
Pepper is also an important vegetable crop in Central and South Taiwan. Chiayi County, Pingtung County, and Kaohsiung County are the mainly produced places in hot pepper production; Nantou County, Yunlin County, and Pingtung County are in sweet pepper production. Pepper plant is typically grown as annual in Taiwan even they can be also grown as perennial in other regions. Although pepper is a warm-season crop, plant is usually grown in fall and winter in Taiwan. The commercial pepper cultivars are often sold as F1 hybrids, which were produced by three-line cytoplasmic male sterility (CMS) system. Using three-line CMS system in pepper F1 hybrid seeds producing has several advantages, as it can reduce labor costs for emasculation, generate purer hybrid seed, and protect all the proprietary while increasing crop vigor and yield through heterosis (Schnable and Wise, 1998; Wise and Pring, 2002; Eckardt, 2006).
According to previous studies, the stability of CMS pepper lines could not always maintain their sterility. Some CMS pepper lines can restore partial or completely fertility caused by low temperature, revert to sterility again when temperature elevated (Kaul, 1988; Peterson, 1958; Shifriss and Guri, 1979; Shifriss, 1997). This
3
phenomenon also observed in specific CMS pepper lines which ware bred by breeders in Taiwan. Two CMS sweet pepper lines of AVRDC-The World Vegetable Center,
‘CCA 7229’ and ‘CCA 7236’ showed fertility in the field in winter from 2005 to 2006 (unpublished data). A CMS hot pepper line ‘ACC. 17’, which was bred by Evergrow Seed CO., LTD, could produce normal pollens and fruit set in the growth chamber at 25/18 °C, 20/18 °C, and 20/12 °C (Hong, 2003).
Up to date, the anther structure and microgametogenesis observations by light microscopy were only compared in CMS pepper and near-isogenic line, for identifing the reasons for the production of no functional pollen by CMS pepper plant (Horner and Rogers, 1974; Luo et al., 2006; Novák and Betlach, 1970). But, few studies have worked on the unstable CMS pepper lines by comparing the anatomical differences between fertile and sterile phenotype under varied temperatures (Hong, 2003).
In present study, both identified CMS and maintainer pepper lines are studied to confirm their genetic background by previously developed markers. Next, the stability of CMS hot and sweet pepper were examined in the phytotron of NTU and in the field of AVRDC–The World Vegetable Center to figure out the critical temperature causing unstable CMS lines restored fertility. Finally, the reason of unstable CMS pepper lines restored fertility were discussed by comparing the anther structure and
4
microgametogenesis in the stable CMS line and the unstable CMS line which expressed fertile and sterile phenotype.
5
Chapter 2 Literature Review
2.1 Cytoplasmic male sterility in higher plant
Cytoplasmic male sterility (CMS) system which had been detected over 150 plant species was a maternally inherited trait (Kaul, 1988). CMS system involves interactions between nuclear and cytoplasm genes. The CMS plants fail to produce functional pollens, which is caused by cytoplasmic dysfunction in plant mitochondrial genomes (Hanson, 1991; Hanson and Bentolila, 2004; Schable and Wise, 1998; Wise and Pring, 2002). The specific nuclear genes, known as fertile restorer genes, restored male fertility of plant carrying a CMS cytoplasm (Bentolila et al., 2002; Hanson and Bentolila, 2004; Schable and Wise, 1998). CMS plants are able to generate next generation by crossing with pollens offered by fertile plants containing fertile restorer genes.
The ‘three-line’ CMS system, which was a breeding system for commercial use, consisted of a male sterile line, a maintainer line, and a restorer line. A male sterile line was maintained by a maintainer line which was propagated by self-pollinating at the same time, and crossed with a restorer line to produce F1 hybrid seeds (Kaul, 1988).
This system played an important role in agronomy due to several advantages, as it
6
could reduce labor costs for emasculation, generate purer hybrid seed, and protect all the proprietary while increasing crop vigor and yield through heterosis (Schnable and Wise, 1998; Wise and Pring, 2002; Eckardt, 2006).
2.2 The characters of CMS-related genes
CMS-related genes, which were expressed as novel chimeric genes, had been already identified in plant mitochondrial genomes (Hanson and Bentolila, 2004;
Schnable and Wise, 1998). Mitochondria were the organelles of respiration, a process of series metabolic pathways of citric acid cycle, mitochondrial electron transport chain and ATP synthesis (Browse et al., 2006). Well-evidenced CMS-related loci were found in standard mitochondrial genes associated to respiration, as in ATP synthase subunit coding sequences, cytochrome oxidase subunit gene coding regions, NADH dehydrogenase subunit, or associated to ribosomal and transfer protein. Besides, more CMS-related genes were often associated with abnormal open reading frames (orfs), which function was not clear, combined with sequences of standard mitochondrial genes or co-transcript with standard mitochondrial genes in some case (Hanson, 1991;
Hanson and Bentolila, 2004; Schable and Wise, 1998; Wise and Pring, 2002).
Several CMS-related genes had been reported in crops. In Brassica napus with
“Polima” (pol) cytoplasm, a rearranged region, orf 224, was in the upstream of atp6 in
7
the CMS phenotype (Handa et al., 1995; Singh and Brown, 1991). In Solanaceae, Petunia was an important model for studying CMS system. The CMS-related gene,
called as pcf (petunia CMS-associated fused gene), consisted of the 5’ region of the atp9 gene which was parts of the first and second exons of the cox II gene, and an
unidentified sequence (urfS) which were co-transcribed (Conley and Hanson, 1995;
Schnable and Wise, 1998). In CMS hot pepper, which was also from Solanaceae family, atp6 and cox II were two candidate CMS-related genes. One copy of atp6 genes lost
one EcoRI site through a C-to-T mutation at 1892bp in the 5’ region and had a 251 bp truncated in the 3’ region. A new ORF, called orf456, was found at the 3’ end of the cox II gene and co-transcribed with the cox II gene upstream (Kim et al, 2001; Kim and Kim, 2005; Kim and Kim, 2006; Kim et al, 2007).
2.3 The characters of restorer genes and interaction with CMS-related genes
The specific dominant nuclear genes, fertility restorer genes (Rf), were able to alter the male sterile phenotype by suppressing or counteracting the expression of the CMS-associated gene (Bentolila et al., 2002; Hanson and Bentolila, 2004; Schable and Wise, 1998). The restoration systems divided into two forms depending of the number of restorer genes. In one form, one or two major restorer loci conferred complete restoration. In another form, full restoration required the concerted action of multiple
8
genes, each one of which provided only small incremental effects (Schable and Wise, 1998).
In molecular identification, pentatricopeptide repeat (PPR) proteins, which contained degenerated repeat units of 35 amino acids, were encoded by the Rf genes, involving the male-fertility restoration to CMS plants. The PPR gene family was a eukaryote-specific protein family particularly expended in higher plants (Small and Peeters, 2000; Andrés et al., 2007; Bentolila et al., 2004; Chase, 2006). The typical structure of PPR family consisted of 3 parts: untargeting protein or plastids- or mitochondria-targeting protein at N-terminus, repeat structure, and additional C-terminal extensions. The character of repeat structure containing 2 to 26 repeat proteins (also called as repeat motifs), with an average of 12 proteins in plants, are as tandem arrays. Depending on the type of repeat proteins, the PPR gene family divided into P subfamily, which comprised all the same repeat motifs with 35 amino acids, and PLS subfamily, which comprised an organized pattern of triple motifs (P-L-S) repeated.
The PLS subfamily of proteins was distinguished by C-terminal extensions, which contained three types of domain, E, E+, and DYW and were unrelated to the repeat motifs. These C-terminal extensions were present as E alone, E combined with E+, or E combined with E+ and DYW, but the roles of these domains were not clear (Andrés
9
et al., 2007; Chase, 2006; Lurin et al., 2004).
Up to date, Rf genes, which encoded mitochondria-targeting proteins with PPR repeat proteins, had been cloned in Petunia, radish, and rice (Hanson and Bentolila, 2004). PPR proteins seemed to operate the maturation of plant mitochondrial (and chloroplast) mRNAs at the post-transcriptional level, including transcript editing and transcript processing. Therefore, the decreased abundance of CMS-related transcripts, or the occurrence of internal processing events that truncate theses transcripts, often accompanied protein products of the CMS-related genes loss in the presence of restorer genes which encoded PPR proteins (Bentolila et al., 2002; Chase, 2006; Mackenzie and Mclntosh, 1999; Small and Peeters, 2000). Such as Rf gene encoding Rf-PPR592 in Petunia, which was able to decrease pcf (petunia CMS-associated fused gene) mRNA expression and reduce the protein products to nearly undetectable levels, restored fertility when transferred to rf/rf CMS plants (Bentolila et al., 2002; Gillman et al., 2007).
2.4 Phenotypes and abnormalities in tapetum of CMS plants
The CMS plants of maize, petunia, and sunflower, which are no distinct morphological differences from male fertile plants but with the degenerated stamens, could not produce viable, functional pollens due to the dysfunction in tapetum (Hanson
10
and Bentolila, 2004; Chase, 2006). The tapetum, an inner layer of anther wall around the sporogenous tissue, played an important role in pollen development. Several functions had been attributed to the tapetum, including the supply of sporopollenin and a role in exine formation, breakdown of callose wall around tetrads, supply of nutrients to developing pollen and reserve metabolites of mature pollen; and supply of exine proteins and surface coat substances (pollenkitt/tryphine). Hence, the abnormalities in the structure and/or function of the tapetum resulted in pollen development arrested (Shivanna et al., 2005).
In some CMS plants, pollen formation is disrupted at the meiotic or post meiotic stage. Such as CMS P. hybrid and P. parodii, the sporogenous tissue of sterile anther stopped developing during early meiosis I, showing that pollen mother cells rarely completed anaphase I (Conley and Hanson, 1995). In CMS sunflower (Helianthus annuus), irregular tapetum cells enlarged radially and crushed tetrads, thereafter
tapetum degenerated. In CMS sweet pepper lines, pollen mother cells could complete meiosis, but the tetrads failed to release microspores and were crushed by highly vacuolated tapetum cells (Horner and Rogers, 1974). In another CMS sweet pepper lines, tetrads of were able to release microspores, which failed to develop into mature pollens due to crushed by the swollen tapetum cells (Luo et al., 2006).
11
Chapter 3
Genetic Background Identification in Hot and Sweet Pepper Lines
3.1 Introduction
CMS system is an interaction between nuclear and cytoplasm genes. The male sterility was often associated with abnormal open reading frames (ORFs) in plant mitochondrial genomes, causing cytoplamic dysfunction (Hanson, 1991; Hanson and Bentolila, 2004; Schable and Wise, 1998; Wise and Pring, 2002). The fertility could be restored by nuclear genes known as fertile restorer genes (Rf) which suppressed the male-sterility phenotype to lead functional pollens produced by plants with CMS cytoplasm (Bentolila et al., 2002; Hanson and Bentolila, 2004; Schable and Wise, 1998).
Structural differences of CMS-related genes in the mitochondrial genomes between male sterility (S type) and male fertility (N type) cytoplasms were observed (Hanson, 1991; Hanson and Bentolila, 2004; Schable and Wise, 1998; Wise and Pring, 2002). In hot pepper, the mitochondrial genes that involved in CMS being identified includes atp6 (ATP synthase subunit 6) and cox II (cytochrome oxidase subunit 2) (Kim et al, 2001; Kim and Kim, 2005; Kim and Kim, 2006; Kim et al, 2007), which participate in ATP synthesis and electron transport chain, respectively. In the
12
mitochondrial genome of pepper, two copies of atp6 genes were identified, designed as atp6-1 and atp6-2. Comparing the sequences of the (N)atp6-2 and (S)atp6-2, the later
one lost one EcoRI site through a C-to-T mutation at 1892bp in the 5’ region and had a 251 bp truncated region at 3’, and (S)atp6-2 was thus renamed as (S)ψatp6-2. Besides, both (N)atp6-2 and (S)ψatp6-2 could be normally transcribed in pepper mitochondria (Kim and Kim, 2005; Kim and Kim, 2006). Another major difference was located at the downstream region of the stop codon of cox II. The sequences of the CMS pepper and the respective maintainer lines diverted from 41bp downstream of the stop codon, and an orf456 adjacent cox II was identified in the CMS line which encoded for a 17-kDa protein and putatively interact with the fertility restore genes (Rf) (Kim and Kim, 2005; Kim et al, 2007). Five primer sets were designed according to sequence of (S)ψatp6-2 and orf456 for comparison the cytoplasmic genetic background of CMS pepper lines developed in TSIPS (Taiwan Seed Improvement and Propagation Station, COA) with hot pepper line ‘Milyang’ in our previous studies (Kim and Kim, 2005;
Kim and Kim, 2006; Kim et al, 2007), and the result confirmed that the TSIPS lines contain (S)ψatp6-2 and orf456 (Lee et al., 2007).
Fertility restorer genes (Rf) of nuclear had the ability to suppress the deleterious effects causing by CMS-related genes. In the Petunia, transformation of the Rf gene,
13
encoded for Rf-PPR 592, could restore the fertility of the CMS Petunia due to decrease mRNA expression of pcf (petunia CMS-associated fused) (Bemtolila et al., 2002). In hot pepper, Rf gene could affect the expression of atp6 mRNA from anther and thus produce functional pollens (Kim and Kim, 2006). Several markers for detecting Rf genes in hot pepper lines have been developed. Two RAPD markers, OP131400 and OW19800, were also able to detect the major Rf gene in hot pepper especially OP131400
which wastightly linked to Rf gene (Zhang et al, 2000); however, these markers could not detect Rf gene in sweet pepper lines even if they were male fertile (Zhang et al, 2000; Kumar et al, 2007). The restorer marker CRF-S reported by Guylas et al (2006) was specific to a major Rf gene in hot pepper. Moreover, a co-dominant CAPS marker conversed from AFLP marker, AFRF8CAPS, was also linked to Rf gene in pepper lines and could differentiate Rf and rf genes (Kim et al, 2006).
The sterility of the CMS pepper lines was not always stable. Some of them might restore fertility partially or completely due to low temperature, and such restoration could be resumed as temperature re-elevating (Hong, 2003; Kaul, 1988; Peterson, 1958;
Shifriss and Guri, 1979; Shifriss, 1997). The genetic background of the unstable CMS lines was not clear yet. In the present study, all the collected CMS pepper lines and the respective maintainers would be examined by the five primer sets that we designed
14
previously for the identification of (S)ψatp6-2 and orf456 (Lee et al., 2007). Besides, both stable and unstable sweet pepper lines developed by AVRDC – The World Vegetable Center would be examined as well. All the samples were also checked for the Rf gene.
3.2 Materials and methods
Plant materials
The CMS lines and the respective maintainer and restorer lines of hot peppers and sweet peppers used in this study were listed in Table 3.1. Seeds of hot and sweet pepper were sown in flats in growth chamber at day/night temperature of 30/25 oC with a 14h photoperiod. Seedlings were be transplanted to 3’ pots until they reached six leaves, and then transplanted to 5’ pots. Plants were maintained in the greenhouse of NTU. Other CMS, maintainer, and restorer pepper lines from AVRDC - The World Vegetable Center examined in this study were also listed in Table 3.1. The leaves of each line were harvested and placed into sealed plastic bags and sent to Taipei under refrigeration and then frozen immediately by liquid nitrogen. The leaf samples were stored at – 80 oC until used.
Reagents and chemicals
Extraction buffer consists of 3% CTAB (w/v), 1.4 M NaCl, 20 mM EDTA, 100
15
mM Tris-HCl (pH 8), and 0.2% β-mercaptoethanol (v/v) added just before use. TNE buffer consists 10 mM Tris-HCl (pH 8), 100 mMNaCl, and 0.5 mM EDTA (pH 8). TE buffer consists 100 mM Tris-HCl (pH 8) and 1 mM EDTA (pH 8). In addition, chloroform : isoamyl alcohol (24:1), 75% ethanol, and 7.5 M ammonium acetate were also prepared.
Extraction of pepper plant’s genomic DNA
0.2 g young green leaves of each pepper line were harvested and ground into fine powder with a mortar and pestle in liquid nitrogen. Powder was transferred to a clean autoclaved 1.5mL eppendorf tube and 700 μl extraction buffer was added. The tube was incubated in 60oC for 30-40 min with twice swirling. 700 μl chloroform: isoamyl alcohol was added into the same tube, and mixed by gentle inversion for 10 times. The tube was centrifuged at 14000 rpm for 10 min at 4 oC and supernatant was transferred to another tube. Repeat these two steps once. The supernatant was precipitated with 450 μl ice-cold isopropenol and centrifuged at 14000 rpm for 10 min at 4 oC. The supernatant was discarded and the pellet was washed twice with 700 μl 75% ethanol.
The pellet was air dried at RT and dissolve in 400 μl TNE buffer. 2 μl RNaseA (10 μg/ml) was added and placed at 4 oC overnight. 400 μl chloroform: isoamyl alcohol was added to the tube, and mixed by gentle inversion for 10 times. The tube was
16
centrifuged at 14000 rpm for 10 min at RT or 4 oC and supernatant was transferred to another tube. 800 μl ice-cold 95% ethanol and 40 μl ice-cold ammonium acetate was added and mixed by gentle inversion. The tube was centrifuged at 14000 rpm for 10 min at RT. The supernatant was discarded and the pellet was washed with 1 mL 70%
and 95% ethanol. The pellet was air dried at RT and dissolve in 200 μl TE buffer. And the DNA solution was stored at – 20 oC until used.
Genomic DNA quality and quantity analysis
The quality of extracted DNA was checked by agarose gel electrophoresis. 5 μl DNA solution and 2 μl loading dye were mixed and loaded in a 0.7% agarose gel. The gel were run at 50 mA for 20 min, and stained by ethidium bromide for 10 min and rinsed by water for 30 min. The quality was estimated by UV light (UVi-Silver GAS7308, UVi-tec, Violet Bioscience INC.).
The quantity of extracted DNA was estimated by spectrophotometry (U-2001 Spectrophotometer, HITACHI Instrument, INC.) by calculating A260/280 ratio, and the DNA concentration was estimated by measuring absorbance at 260 nm. DNA concentration (ng/μl) = O.D. 260 x 50 μg/ml x 100 (dilution factor). The concentration of each sample was adjusted to10 ng/μl, and stored the DNA solution at -20 oC until used.
17
PCR amplification to genetic background identification
In order to identify genetic background of each pepper line, primer sets listed in table 3.2 for the identification of CMS-related genes and fertile restorer gene were used in this study. PCR amplification was performed in RoboCycler GRADIENT 96 (STRATAGENE INC.) Each PCR reaction mixture of 25 μl consisted of 60ng genomic DNA, 2.5 μl 10X reaction buffer, 3 μl of 25 mM MgCl2, 2 μl of 2.5 mM dNTPs, 0.5μl of 10 μM primer set, and 0.5 μl Taq DNA polymerase. For identification of CMS-related genes, the PCR reaction mixtures were heated at an initial step of 92
oC for 2 min, and then subjected to 35 cycles of following program: 92 oC for 1min, 52
oC for 45 s, 72 oC for 1 min. After the last cycle, temperature was maintained at 72 oC
for 10 min, and then cooled at 6 oC for 10 min. For identification of fertile restorer gene (Gulyas et al., 2006), The PCR reaction mixtures were heated at an initial step of 94 oC for 3 min, and then subjected to 35 cycles of following program: 94 oC for 30s, 58 oC for 1 min, 72 oC for 1 min. After the last cycle, temperature was maintained at 72
oC for 5 min, and then cooled at 6 oC for 10 min. The amplified DNA were stored – 4
oC until used. The amplified DNA was electrophorised in a 1.2% agarose gel
containing 0.5 mg/ml ethidium bromide at 50 mA for 25-30 min. The results were recorded by UV light (UVi-Silver GAS7308, UVi-tec, Violet Bioscience INC.).
18
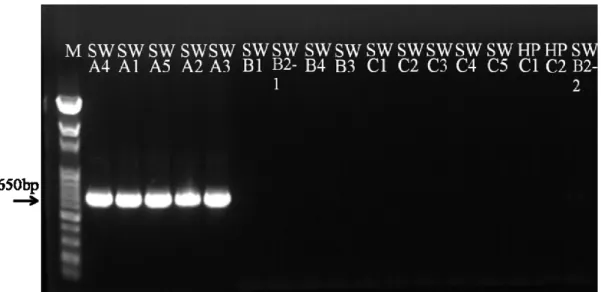
3.3 Results and discussion
All the CMS pepper lines could generate predicted (S)ψatp6-2 fragments by each primer set, and their maintainer lines didn’t produced the predicted products (Fig. 3.1, Fig. 3.3 and Table 3.3). For orf456, similar results were obtained (Fig. 3.2 and Fig. 3.4) and summarized in Table 3.3. These results revealed that the cytoplasms of all the CMS pepper lines including hot and sweet pepper lines contained (S)ψatp6-2 and orf456, and their maintainers of hot pepper lines, with N cytoplasm, didn’t have such
defected ψatp6-2 and orf456 as the maintainer of CMS hot pepper line ‘Milyang’ (Kim and Kim, 2005; Kim and Kim, 2006). Surprisingly, ‘PBC 84 selex’ and ‘9946-2138’, maintainer lines of unstable CMS sweet pepper ‘CCA 7236’ and ‘CCA 7231’
respectively, could generate orf456 fragment by CMS P1 or CMS P2 primer set (Fig.
3.2, Fig. 3.4 and Table 3.3). For ‘9847-4754’, the other maintainer line of stable CMS line ‘CCA 7234’, did not generate orf456 fragment. (Fig. 3.5 and Table 3.3). These results revealed that the genetic background of cytoplasm in maintainers of unstable CMS sweet pepper lines might be more complicated and different from it in maintainers of CMS hot pepper lines and of stable CMS sweet pepper line. In previous studies, CMS-related genes, (S)ψatp6-2 and orf456, were cloned only in hot pepper line ‘Milyang’ (Kim et al, 2001; Kim and Kim, 2006; Kim et al, 2007), and it was not
19
clear that which one of CMS-related genes mainly caused male sterile phenotype (Kim et al, 2007). All of the restore lines including hot and sweet peppers did not generateψatp6-2 fragment by ATP 1 primer set, but only two restorer sweet pepper line ‘C03804’ and ‘PBC1419-Y’, as an unstable restorer and a stable restore respectively, generated orf456 fragment by CMS P1 primer set (Fig. 3.3, Fig. 3.4 and Table 3.3). These results showed that cytoplasm of restorer lines containing orf456 fragment was not related to the stability of fertile expression.
For Rf gene, all the CMS lines and maintainers did not generate predicted Rf fragment by CRF-S, and a control ‘A5’, a restorer hot pepper line with identified genetic background Rf Rf from TRIS, generated predicted product (Fig. 3.5, Fig. 3.6 and Table 3.3). These results revealed that the nuclei of all the CMS and maintainers including hot and sweet pepper lines did not have Rf as the results of a CMS line (Gulyas et al., 2006). Surprisingly, only restorer hot pepper line ‘PBC 142’ and restorer sweet pepper line ‘9852-131’ could generate predicted product (Fig. 3.6 and Table 3.3).
The result reveals that this primer set might not be tightly linked to Rf gene. Another two RAPD markers, OP131400 and OW19800, were also able to detect the major Rf gene in hot pepper, especially OP131400 which wastightly linked to Rf gene (Zhang et al, 2000); however, the markers could not detect Rf gene in some restorer lines with
20
identified genetic background Rf Rf or Rf rf (Zhang et al, 2000; Kumar et al, 2007).
The markers for detecting CMS-related genes were necessary for early genetic background identification, and reducing costs for waiting plants grown up to examine floral characteristics. Besides, the analysis results could be convinced even if the unstable sterility expression of CMS plants influenced by environment (Kim and Kim, 2005). According to the data in this study, the S cytoplasm of CMS lines including hot and sweet peppers were able to examine by each primer sets while plants was young, but N cytoplasm of maintainer sweet pepper lines need identify by fertility phenotype expression.
21
Table 3.1 The pedigree of CMS, maintainer or near-isogenic lines, and restorer pepper lines derived from AVRDC- The World Vegetable Center.
No. CCA Code Pedigree hot/sweet pepper line of CMS system Stability of sterility
CCA7243 Seungchon(cms)/8*Arunalu hot CMS Unstable
= CCA4758/PBC 483
CCA7244 Seungchon(cms)/8*9907-9611 hot CMS Stable
=CCA4759/9907-9611
PBC 483 Arunalu hot maintainer of CCA 7243
9907-9611 PBC 385 sle. hot maintainer of CCA 7244
CCA7236 PBC385-Aline/9*Jin's Sweetie selex sweet CMS Unstable CCA7234 PBC385-Aline/9*Mito Lee Selex sweet CMS Stable PBC 84 selex Jin's Sweetie sweet maintainer of CCA 7236
9847-4754 Mito Lee selex sweet maintainer of CCA 7234
SW-A1 CCA7234 PBC385-Aline/9*Mito Lee Selex sweet CMS Stable SW-A2 CCA7236 PBC385-Aline/9*Jin's Sweetie selex sweet CMS Unstable SW-A3 CCA7231 Suwon cms//9*Maor/Perennial sweet CMS
SW-A4 CCA7229 PBC385-Aline/8*White King sel. sweet CMS Unstable SW-A5 CCA7235 PBC385-Aline//9*HDA120/MI-Gold sweet CMS Unstable SW-B1 9847-4754 F1 Mito Lee selex sweet maintainer of CCA 7234
SW-B2-1 PBC 84 selex* Jin's Sweetie sweet maintainer of CCA 7236 SW-B2-2 PBC 84 selex* Jin's Sweetie sweet maintainer of CCA 7236
*Same line from different individual plant.
22
Table 3.1 (cont.) The pedigree of CMS, maintainer or near-isogenic lines, and restorer pepper lines derived from AVRDC- The World Vegetable Center.
No. CCA Code Pedigree hot/sweet pepper line of CMS system Stability of sterility SW-B3 9946-2138 Maor/Perennial-1 sweet maintainer of CCA 7231
SW-B4 9852-174-3 F1 BlueStar/ECW-30R sweet near-isogenic line
HP-C1 9955-15 9955-15 hot restorer
HP-C2 PBC 142 Pant C-1 hot restorer
SW-C1 PP0037-7645 Yellow#1 sweet restorer (unstable)
SW-C2 9852-131 CCA 215A sweet restorer
SW-C3 0407-7094 0407-7094 sweet restorer (unstable)
SW-C4 C03804 403 7499 sweet restorer (unstable)
SW-C5 PBC1419-Y C01757 SELEX sweet restorer
23
Table 3.2 Primer sets used in this study.
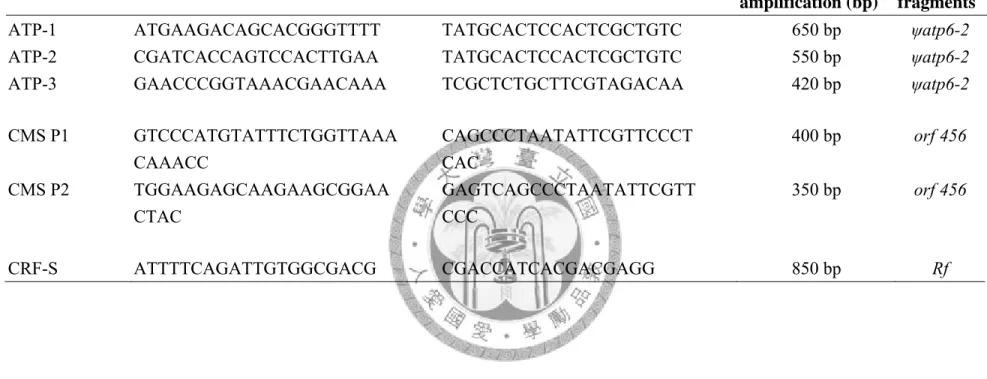
Primer set 5` forward primer 3` reverse primer size of predicted amplification (bp)
name of fragments ATP-1 ATGAAGACAGCACGGGTTTT TATGCACTCCACTCGCTGTC 650 bp ψatp6-2 ATP-2 CGATCACCAGTCCACTTGAA TATGCACTCCACTCGCTGTC 550 bp ψatp6-2 ATP-3 GAACCCGGTAAACGAACAAA TCGCTCTGCTTCGTAGACAA 420 bp ψatp6-2
CMS P1 GTCCCATGTATTTCTGGTTAAA CAAACC
CAGCCCTAATATTCGTTCCCT CAC
400 bp orf 456
CMS P2 TGGAAGAGCAAGAAGCGGAA CTAC
GAGTCAGCCCTAATATTCGTT CCC
350 bp orf 456
CRF-S ATTTTCAGATTGTGGCGACG CGACCATCACGACGAGG 850 bp Rf
24
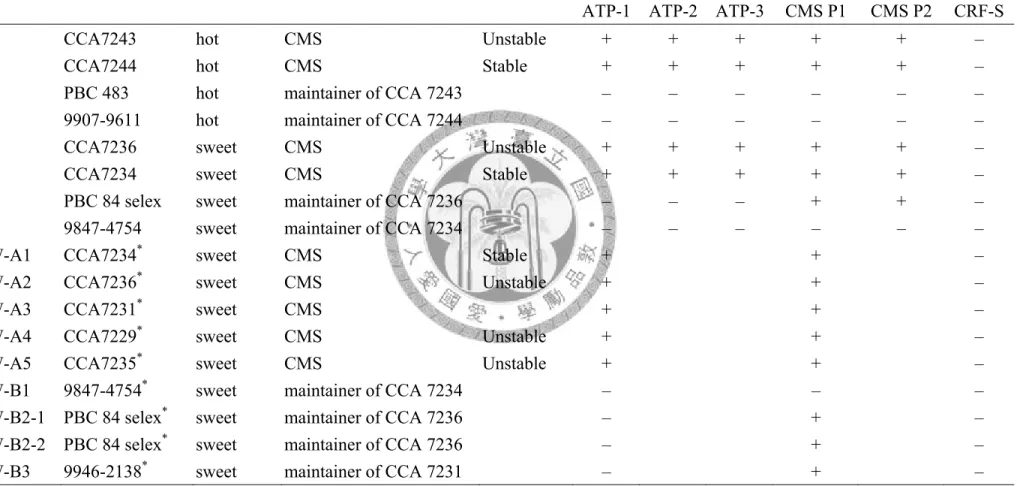
Table 3.3 Genomic pepper DNA analysis by ATP-1, ATP-2, ATP-3, CMS P1, CMS P2, and CRF-S primer sets.
No. CCA Code hot/sweet pepper
line of CMS system Stability of sterility
primer seta
ATP-1 ATP-2 ATP-3 CMS P1 CMS P2 CRF-S
CCA7243 hot CMS Unstable + + + + + –
CCA7244 hot CMS Stable + + + + + –
PBC 483 hot maintainer of CCA 7243 – – – – – –
9907-9611 hot maintainer of CCA 7244 – – – – – –
CCA7236 sweet CMS Unstable + + + + + –
CCA7234 sweet CMS Stable + + + + + –
PBC 84 selex sweet maintainer of CCA 7236 – – – + + – 9847-4754 sweet maintainer of CCA 7234 – – – – – –
SW-A1 CCA7234* sweet CMS Stable + + –
SW-A2 CCA7236* sweet CMS Unstable + + –
SW-A3 CCA7231* sweet CMS + + –
SW-A4 CCA7229* sweet CMS Unstable + + –
SW-A5 CCA7235* sweet CMS Unstable + + –
SW-B1 9847-4754* sweet maintainer of CCA 7234 – – –
SW-B2-1 PBC 84 selex* sweet maintainer of CCA 7236 – + – SW-B2-2 PBC 84 selex* sweet maintainer of CCA 7236 – + –
SW-B3 9946-2138* sweet maintainer of CCA 7231 – + –
a (+) Presence and (–) absence of bands.
*CMS-related genes analysis by primer set ATP6 1 and CMS P1.
25
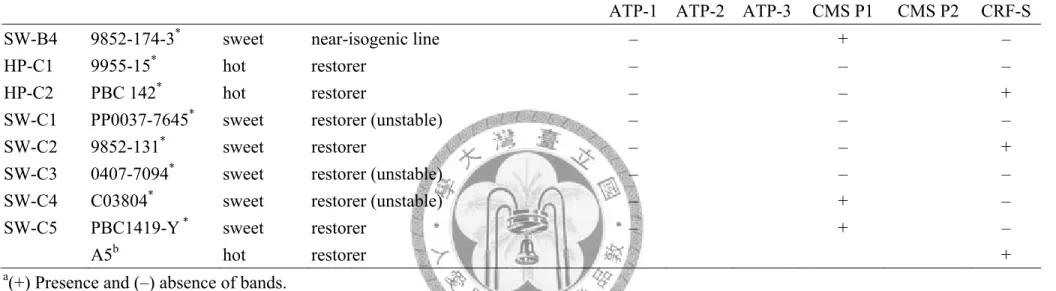
Table 3.3 (cont.) Genomic pepper DNA analysis by ATP-1, ATP-2, ATP-3, CMS P1, CMS P2, CRF-S primer sets.
No. CCA Code
hot/sweet
pepper line of CMS system
Stability of sterility
primer seta
ATP-1 ATP-2 ATP-3 CMS P1 CMS P2 CRF-S
SW-B4 9852-174-3* sweet near-isogenic line – + –
HP-C1 9955-15* hot restorer – – –
HP-C2 PBC 142* hot restorer – – +
SW-C1 PP0037-7645* sweet restorer (unstable) – – –
SW-C2 9852-131* sweet restorer – – +
SW-C3 0407-7094* sweet restorer (unstable) – – –
SW-C4 C03804* sweet restorer (unstable) – + –
SW-C5 PBC1419-Y * sweet restorer – + –
A5b hot restorer +
a(+) Presence and (–) absence of bands.
bGenomic DNA of hot pepper plant with identified genomic background Rf Rf from TRIS.
*CMS-related genes analysis by primer set ATP6 1 and CMS P1.
26
Fig. 3.1 PCR amplification of ψatp6-2 fragment from pepper leaf genomic DNA by (A) ATP-1, (B) ATP-2, and (C) ATP-3 primer sets.
27
Fig. 3.2 PCR amplification of orf456 fragment from pepper leaf genomic DNA by (A) CMS P1, and (B) CMS P2 primer sets.
28
Fig. 3.3 PCR amplification of ψatp6-2 fragment from pepper leaf genomic DNA by ATP-1 primer set.
Fig. 3.4 PCR amplification of orf456 fragment from pepper leaf genomic DNA by CMS P1 primer set.
29
Fig. 3.5 PCR amplification of Rf fragment from pepper leaf genomic DNA by CRF-S primer set.
Fig. 3.6 PCR amplification of Rf fragment from pepper leaf genomic DNA by CRF-S primer set.
30
Fig. 3.6 (cont.) PCR amplification of Rf fragment from pepper leaf genomic DNA by primer set CRF-S.
31
Chapter 4
The Influences of Temperature and Seasonal Changes on Cytoplasm Male Sterile Pepper Lines
4.1 Introduction
The sterility of cytoplasm male sterile (CMS) pepper lines were not always expressed, some CMS lines could be affected by some environmental factors and restored their fertility partial or completely and thus produce functional pollens leading to self-pollination. And the sterility could resume as the temperature raise up again (Kaul, 1988; Peterson, 1958; Shifriss and Guri, 1979; Shifriss, 1997). The environmental factors that might affect the restoration include temperature, photoperiod, light intensity, soil micronutrients and soil pH, and of which temperature and photoperiod were two major environmental factors (Kaul, 1988). In Brassica napus L., the Polima cytoplasmic male sterility (Pol CMS) was highly dependent on temperature and thus could be divided into three types: high temperature CMS lines, low temperature CMS lines and stable CMS lines. Photoperiod did not affect the stability of Pol CMS Brassica napus L. lines (Fu et al. 1990). However, photoperiod-sensitive cytoplasmic male sterility (PCMS) of wheat was caused by the interaction between Aegilops crassa cytoplasm and Triticum aestivum cv. Norin 26 nucleus. It expressed almost complete male sterility under long-day conditions of 15 h or longer, but restored
32
fertility under short-day conditions of 14.5 h or less. The male sterility of PCMS wheat was not sensitive to temperature (Murai and Tsunewaki, 1993). Peterson (1958) observed that CMS sweet pepper would not produce pollens in summer, but produced 20% to 30% pollens in late October. The fertility of these set of plants was completely restored during the whole winter in the greenhouse. However, male sterility would be resumed when temperature went higher. In CMS sweet pepper line ‘Bikura’ and ‘Zohar’, plants restored fertility as night temperature dropped to 15°C in winter, but resumed their male sterility about 6 days after plants moving to growth chamber at 32/25°C (Shifriss and Guri, 1979). The CMS sweet pepper lines of AVRDC–The World Vegetable Center, ‘CCA 7229’ and ‘CCA 7236’, showed fertility restoration in the field in winter from 2005 to 2006, which suggested that these two CMS sweet pepper line were unstable lines which could produce functional pollens when the temperature below 18 °C. Besides, ‘CCA 7229’, ‘CCA 7235’, and ‘CCA 7236’ could restore fertility 17 days after transferring form the summer field to the growth chamber at 23/20 °C (unreported data). In CMS hot pepper line, ‘ACC. 17’ could produce viable pollens and set fruit in the growth chamber at 25/18 °C, 20/18 °C, and 20/12 °C, and the viable pollen numbers produced by ‘ACC. 17’ showed no significant difference with those produced by fertile lines (Hong, 2003). Besides, Hong (2003) also suggested that 20
33
days low temperature period before anthesis was required for fertility restoration.
In the present study, we would confirm the stability of cytoplasmic male sterility of hot and sweet pepper lines both in the phytotron and in the field of AVRDC–The World Vegetable Center.
4.2 Materials and methods
Plant materials
The CMS lines and maintainers of hot peppers and sweet peppers (Table 4.1) were used in this study. Plants were grown in the phytotrons and greenhouse at National Taiwan University from January 2007 to July 2007 and in the field of AVRDC – The World Vegetable Center from December 2007 to July 2008.
Seedlings of hot pepper being grown at NTU were sown on Jan. 3 to Feb. 9, 2007 in flats in growth chamber at day/night temperature of 30/25 oC with a 14h photoperiod.
Seedlings were transplanted into 3’ pots as they reached six leaves and moved to the greenhouse of NTU for 1 month then they were transplanted into 5’ pots. One to three plants of each line were moved to the phytotron of NTU at 30/25 oC, 25/20 oC and 20/15
oC under natural photoperiod on May 15 for hot pepper and on Mar. 18 for sweet pepper.
Remaining plants were maintained in the greenhouse. During the growth period, lateral
34
leaf buds and fruit need to be thinned. Temperature in the greenhouse was recorded by HOBO every day (Appendix 1).
Seedlings that grown at AVRDC – The World Vegetable Center were sown in flats on August 15, October 1, and January 1, in order to maintain plant vigor during the observation. Seedlings were transplanted to the field at AVRDC - The World Vegetable Center as they reached six leaves. During the growth period, fruits need to be thinned.
Temperature in the field was recorded by LogTag Humidity and Temperature Records (LogTag Records Ltd) every day (Appendix 2).
Percentage of pollen viability
Pollen viability investigation was started from the first flower bloomed. In NTU, pollen viability of each line was recorded once per week, one anther of a pre-bloomed bud per line was sampled, and final pollen viability recorded was the average of counts from two light fields. In the AVRDC - The World Vegetable Center, pollen viability was recorded once every two weeks, 6 plants per line and of which one pre-bloomed bud per plant were sampled for pollen viability, every anther was inspected and pollen viability recorded was the average of every plant.
Pollen viability was determined by the percentage of stained pollen grains from
35
one anther with 0.5% acetocarmine under 100X light microscope. The percentage of pollen viability was calculated with “No. of fertile pollens / No. of fertile plus sterile pollens.”
4.3 Results
The phenotype of CMS lines and maintainers in the field of AVRDC – The World Vegetable Center
The morphological phenotypes of CMS pepper plants were similar to that of maintainer lines (Fig. 4.1 and Fig. 4.3) except their anthers. Comparing the phenotype of anthers during anthesis, the anthers of CMS line were smaller than those of the maintainers (Fig. 4.2 and Fig. 4.4). Although the anther of CMS line could dehisce, no light yellow pollen was observed on the surface of anther. When unstable CMS line
‘CCA 7243’ restored fertility, the phenotype of anthers was similar to that of the maintainer ‘PBC 385 (Fig. 4.2 B and C).
The influences of temperature and seasonal changes on CMS hot pepper lines and the maintainers
To determine the critical range of temperature that caused unstable CMS pepper lines restore fertility, plants were grown in the greenhouse of NTU and the phytotron of NTU at 30/25 oC, 25/20 oC and 20/15 oC under natural photoperiod. Since all the plants
36
were grown in the greenhouse before moved to the phytotrons at May 15, 2007, most of CMS hot pepper line ‘CCA 7243’ expressed fertility restoration from April 29 to May 8, 2007 (Fig.4.5A). In the present study, the night temperature in the greenhouse was below 20 oC before Apr. 21 and there was a front at Apr. 5 lowering both day and night temperature to below 15 oC while the day temperature maintained mostly above 25 oC.
Such long period of lower temperature might explain the fertility restoration in the greenhouse. The highest restoration of ‘CCA 7243’ was appeared on May 8 and went down thereafter and finally reverted their male sterility in the greenhouse. ‘CCA 7243’
plant moved to 20/15 oC phytotron kept expressing the fertility while those moved to 30/25 oC and 25/20 oC phytotrons resumed to male sterile (Fig.4.5A). In the stable CMS line ‘CCA 7244’, all plants remained male sterility in every location (Fig. 4.5B). Both maintainer line `PBC 483’ and ‘9907-9611’ expressed fertile in the greenhouse and in the phytotron, especially in the phytotron at 25/20 oC and 20/15 oC, the percentage of pollen viability maintained more than 70% (Fig. 4.5 C and D). Due to server aphid problem, plants in the phytotron could not flower continuously and resulted in no data (Fig. 4.5).
For the field observation in AVRDC – The World Vegetable Center, CMS line
‘CCA 7243’ had restored 5 times of fertility from Dec. 7, 2007 to May 8, 2008, and the
37
fertility could restore about 50% as compared to the maintainer line, ‘PBC 385’. The percentage of pollen viability in maintainers ‘PBC 385’ and ‘9907-9611’ was about 60%
or more, except for the last observation in ‘9907-9611’ which that only 18% (Fig. 4.6).
Surprisingly, ‘CCA 7244’ observed some fertility twice in this field experiment. During the experimental period, the night temperature in the field was lower than 21 oC before Apr. 8 while day temperature maintained mostly higher than 26 oC, and long period of lower temperature might also explain the fertility restoration in the field.
The influences of temperature and seasonal changes on CMS sweet pepper lines and the maintainers
In sweet pepper lines, since all the plants of CMS line ‘CCA 7236’ were grown in the greenhouse before transferred into phytotron at May 18, 2007, most plants restored fertility from Apr, 6 to May 8, 2007 (Fig 4.7 A). The temperature changes in the greenhouse of NTU showed in Appendix 1, and long period of lower temperature might also caused the fertility restoration of unstable CMS sweet pepper line ‘CCA 7236’ in the greenhouse. The highest rate of the restoration of ‘CCA 7236’ was appeared on Apr.
22 and which went down thereafter and finally male sterility was reverted in the greenhouse. ‘CCA 7236’ plants which were moved to 20/15 oC phytotron kept their fertility while those were moved to 30/25 oC and 25/20 oC phytotrons resumed to male
38
sterile (Fig.4.7 A). In another CMS sweet pepper line ‘CCA 7234’, all plants remained male sterility in every location (Fig. 4.7 B). Both maintainer lines `PBC 84 selex’ and
`9847-4754’ expressed fertility in each location, but plants could hardly develop flowers in the 30/25 oC phytotron (Fig. 4.7 C and D). Due to server aphid problem, plants in the phytotron could not flower continuously and resulted in no data (Fig. 4.7).
For the field observation in AVRDC in CMS ‘CCA 7236’, plants restored fertility three periods from Dec. 7, 2007 to Apr. 10, 2008, and each restoration period could maintain 2 weeks or more, and the fertility could restore about 35% to 60% as compared to the maintainer line, `PBC 84 selex’. Maintainers `PBC 84 selex’ and
`9847-4755’ maintained fertile but pollen viability was not steady, which was about 20% to 95%. Another CMS line ‘CCA 7234’ remained male sterility in the field (Fig.
4.8). During the experimental period, the night temperature in the field was lower than 17 oC before Mar. 10 while daily temperature maintained mostly higher than 26 oC.
There was a front on Jan. 30 causing day temperature to drop to 18 oC or even lower.
Several sweet pepper plants suddenly wilted due to the infection of Sclerotium rolfsii from Apr. 2008.
4.4 Discussion
39
Peterson (1958) indicated that the floral organ of CMS pepper was similar to that of fertile pepper, but its stamens were approximate one-half of the size of fertile anthers and just produced few viable pollen grains occasionally. The color and size of anthers between CMS line while plants expressed male sterility and their maintainers was distinctly different. When unstable CMS hot pepper line ‘CCA 7243’ restored fertility, the phenotype of anthers in unstable CMS plants was as same as it in maintainer ‘PBC 385’ (Fig. 4.2 B and C), but sometimes no pollen grains were obtained in anthers of unstable CMS line and the fertility restoration needed investigate by light microscope.
The unstable CMS sweet pepper lines of AVRDC – The World Vegetable Center were recorded to restore their fertility approximately after 17 d incubated in 23/20oC growth chamber (unreported). Hong (2003) reported that the temperature at 20 d before anthesis could cause CMS hot pepper fertility restoration. According to the greenhouse and the field observations in this study, the process of pepper floral bud development stages from the visual bud to anthesis needed in almost 30 d, we suggested that the temperature at 30 d before unstable CMS plants showed fertility was related to plant fertility restoration.
Previous reports did not show the critical range of temperature that caused unstable CMS lines restore fertility. Shifriss (1997) suggested that CMS pepper lines restored
40
fertility when the temperature dropped below the optimal for pepper production (day/night temperature was 25/17oC). The other unstable CMS hot pepper line ‘AAC.
17’ could restore fertility at 25/18oC, 20/18 oC, and 20/12 oC growth chamber (Hong, 2003). The unstable CMS sweet pepper lines of AVRDC restored fertility when minimum temperature less than 18 oC in the field, and also restored at 23/20 oC or at 23/15 oC growth chamber (unreported).
In this study, our data showed that night temperature is the critical factor caused unstable CMS lines restore fertility. CMS hot pepper line ‘CCA 7243’ could restore fertility in the greenhouse and in the field while the night temperature was below 21 oC, and also could restore in the 20/15 oC phytotron (Fig. 4.5 A and Fig. 4.7). Plants of CMS sweet pepper line ‘CCA 7236’ restored fertility while night temperature was below 21oC or 17 oC, but plants hardly restored while day temperature below 18 oC (Fig.
4.6 A and Fig. 8). Besides, the fertility restoration of unstable CMS lines showed as a regular cycle, not always steady expressed. When night temperature went higher, both lines were reverted male sterile. Therefore, ‘CCA 7243’ and ‘CCA 7236’ both belonged to unstable CMS line.
In other CMS hot pepper line ‘CCA 7244’, plants had ever restored two times of fertility in the field of AVRDC (Fig. 4.6), but remained as male sterile in the greenhouse
41
and each phytotron (Fig. 4.5 B), and these results representing that ‘CCA 7244’ were less sensitive to temperature changes than ‘CCA 7243’. CMS sweet pepper line ‘CCA 7234’ always expressed male sterile even under a long period of lower night temperature. Therefore, ‘CCA 7244’ and ‘CCA 7234’ are stable CMS lines.
The reasons why unstable CMS plants could restore fertility as environment changes could be classified into two categories. One, the genetic background of CMS plants might be related to fertility restoration expressed under specific conditions. The level of male sterility expression of PCMS wheat lines was determined by the genotype of the nuclear donor (Murai, 2001a). Certain temperature could suppress or delay the recessive restorer gene action in some CMS plants, such as onion, alfalfa, pearl millet and rye (Kaul, 1988). The variation among CMS pepper lines in the expression of sterility presumably was caused by differences in number and nature of male-sterility modifying genes (Peterson, 1958; Shifriss, 1997). In cytological aspects, the PCMS wheat at floret development stage was sensitive to photoperiod, such as pistillate stamens under long-day condition (17h) transferring into normal stamens under short-day condition (13h) and producing functional pollens to self-pollination (Murai and Tunewaki, 1993). On the other hand, the breakdown during the process of microsporogenesis or microgametogenesis in CMS plants might block under the
42
specific conditions. Because malfunction might occur in early microsporogenesis in CMS pepper (Peterson, 1958), meiotic breakdown in unstable CMS pepper lines is either stopped or delayed when the temperature drop below the optimal temperature for pepper production and plants could produce viable pollens (Shifriss, 1997).
The fertile expression in maintainers was also affected by temperature. In hot pepper maintainers ‘PBC 483’ and ‘9907-9611’, the pollen viability maintained 60%
and higher in the 20/15 oC and 25/20 oC phytotron, but fewer flowers bloomed and pollen viability decreased in the 30/25 oC phytotron at (Fig. 4.5 C and D). However,
‘PBC 483’ and ‘9907-9611’ in the field still maintained higher fertility even if the day temperature was higher more than 30 oC but night temperature was lowerthan 25 oC (Fig.
4.6). The same results were in maintainer sweet pepper line ‘PBC 84 selex’ and
`9847-4755’. (Fig. 4.7 C and D and Fig. 4.8). According to previous reports, the highest night temperature (24 oC ± 1) caused sweet pepper ‘Ma'or’ considerable blossom drop, but the highest tested day temperature (28 oC) did not cause increased blossom drop (Rylski and Spigelman, 1982). It could explain that the pollen viability of maintainers decreased in the 30/25 oC phytron at caused by high night temperature.
In tradition three-line CMS system, CMS plant was crossed by maintainer line to maintain their progeny, and crossed by restorer line to produce F1 seeds. The unstable
43
CMS plant which was sensitive to environment was also useful. The unstable CMS plants maintained their progeny by self-pollination while plants restored fertility, and produced hybrid seeds by cross-pollination with restorer while CMS plants remained sterile. It is so-called ‘two-line system’ (Murai and Tsunewaki, 1993; Murai et al, 2008;
Shifriss, 1997; Murai, 2001; Yang et al, 2006). Therefore, unstable CMS pepper lines used in this study also could develop into ‘two-line system’, and plants maintained their progeny from Dec. to Jun while night temperature was below 21 oC and produced hybrid seeds during Jul. to Nov. while night temperature was above 21 oC.