Tackling the Cytotoxic Effect of a Marine Polycyclic
Quinone-Type Metabolite: Halenaquinone Induces Molt 4 Cells Apoptosis via Oxidative Stress Combined with the Inhibition of HDAC and Topoisomerase Activities
Shou-Ping Shih
1,†, Man-Gang Lee
2,3,†, Mohamed El-Shazly
4, Yung-Shun Juan
5,6,7, Zhi-Hong Wen
2, Ying-Chi Du
8, Jui-Hsin Su
1,9, Ping-Jyun Sung
1,9, Yu-Cheng Chen
10, Juan-Cheng Yang
11,12, Yang-Chang Wu
11,12,13,14,* and Mei-Chin Lu
1,9,*
1
Graduate Institute of Marine Biotechnology, National Dong Hwa University, Pingtung 944, Taiwan; E-Mails: m [email protected] (S.-P.S.); x2219@nm m ba.gov.tw (J.-H.S.);
[email protected] (P.-J. S.)
2
Department of Marine Biotechnology and Resources, National Sun Yat-sen University,
Kaohsiung 804, Taiwan; E-Mails: m [email protected] m .t w (M.-G.L.);
wzh@ m ail.nsysu.edu.tw (Z.-H.W.)
3
Division of Urology, Department of Surgery, Zuoying Branch of Kaohsiung Armed Forces General
Hospital, Kaohsiung 813, Taiwan; E-Mail: m [email protected] m .tw
4
Department of Pharmacognosy and Natural Products Chemistry, Faculty of Pharmacy, Ain-Shams
University, Organization of African Unity Street, Abassia, Cairo 11566, Egypt;
E-Mail: elshazly444@google m ail.com
5
Department of Urology, Kaohsiung Municipal Hsiao-Kang Hospital, Kaohsiung 812, Taiwan;
E-Mail: j u a n uro@g m ail.com
6
Department of Urology, College of Medicine, Kaohsiung Medical University, Kaohsiung 807, Taiwan
7
Department of Urology, Kaohsiung Medical University Hospital, Kaohsiung 807, Taiwan
8
Department of Botanicals, Medical and Pharmaceutical Industry Technology and Development
Center , New Taipei City 248, Taiwan; E-Mail: ycdu0626@g m ail.com
9
National Museum of Marine Biology & Aquarium, Pingtung 944, Taiwan
10
The PhD Program of Cancer Biology and Drug discovery, China Medical University,
Taichung 404, Taiwan; E-Mail: j520c@hot m ail.com
11
School of Pharmacy, College of Pharmacy, China Medical University, Taichung 404, Taiwan;
E-Mails: [email protected] m .tw (J.-C.Y.); yachwu@ m ail.c m u.edu.tw (Y.-C.W.)
12
Chinese Medicine Research and Development Center, China Medical University Hospital,
Taichung 404, Taiwan
13
Graduate Institute of Natural Products, Kaohsiung Medical University, Kaohsiung 807, Taiwan
14
Center of Molecular Medicine, China Medical University Hospital, Taichung 404, Taiwan
†
These authors contributed equally to this work.
* Authors to whom correspondence should be addressed;
E-Mails: yachwu@ m ail.c m u.edu.tw (Y.-C.W.); [email protected] (M.-C.L.);
Tel.: +886-4-2205-7153 (Y.-C.W.); +886-8882-5037 (M.-C.L.);
Fax: +886-4-2206-0248 (Y.-C.W.); +886-8882-5087 (M.-C.L.).
Abstract: A marine polycyclic quinone-type metabolite, halenaquinone (HQ), was found to inhibit the proliferation of Molt 4, K562, MDA-MB-231 and DLD-1 cancer cell lines, with IC
50of 0.48, 0.18, 8.0 and 6.76 μg/mL, respectively. It exhibited the most potent activity against leukemia Molt 4 cells. Accumulating evidence showed that HQ may act as a potent protein kinase inhibitor in cancer therapy. To fully understand the mechanism of HQ, we further explored the precise molecular targets in leukemia Molt 4 cells. We found that the use of HQ increased apoptosis by 26.23%–70.27% and caused disruption of mitochondrial membrane potential (MMP) by 17.15%–53.25% in a dose-dependent manner, as demonstrated by Annexin-V/PI and JC-1 staining assays, respectively. Moreover, our findings indicated that the pretreatment of Molt 4 cells with N-acetyl-
L-cysteine (NAC), a reactive oxygen species (ROS) scavenger, diminished MMP disruption and apoptosis induced by HQ, suggesting that ROS overproduction plays a crucial rule in the cytotoxic activity of HQ. The results of a cell-free system assay indicated that HQ could act as an HDAC and topoisomerase catalytic inhibitor through the inhibition of pan-HDAC and topoisomerase IIα expression, respectively. On the protein level, the expression of the anti-apoptotic proteins p-Akt, NFκB, HDAC and Bcl-2, as well as hexokinase II was inhibited by the use of HQ. On the other hand, the expression of the pro-apoptotic protein Bax, PARP cleavage, caspase activation and cytochrome c release were increased after HQ treatment. Taken together, our results suggested that the antileukemic effect of HQ is ROS-mediated mitochondrial apoptosis combined with the inhibitory effect on HDAC and topoisomerase activities.
Keywords: halenaquinone; histone deacetylase (HDAC); mitochondria; reactive oxygen species (ROS); topoisomerase
1. Introduction
Protein acetylation is one of the vital post-translational modifications that regulate protein stability,
function and intracellular compartmentalization. It is involved in epigenetic regulation through
chromatin remodeling. Histone deacetylase inhibitors (HDACi) exert anticancer activity by promoting
acetylation of histones, as well as by promoting acetylation of non-histone protein substrates, including α-tubulin, heat shock protein 90 or p53 [1,2]. Accumulating evidence suggested that HDAC activity plays an important role in tumorigenesis (especially leukemia), and thus, these epigenetic modifications could be potential targets for cancer therapy [3,4]. Suberoylanilide hydroxamic acid (SAHA) and romidepsin (depsipeptide) were approved as HDACi for clinical treatment by the U.S.
Food and Drug Administration for the treatment of cutaneous T-cell lymphoma in 2006 and 2009, respectively [5–7]. There are currently ten HDAC inhibitors in clinical trials, including six hydroxamic acids, two benzamides and two fatty acids [8].
There are several lines of evidence demonstrating that drugs targeting DNA topoisomerase (topo) I and II act as specific DNA-damaging agents and can be used in cancer therapy, because tumor cells could not sustain significant defects in the DNA repair pathway [9]. Topo I poison-targeting drugs used in cancer therapy include camptothecin, irinotecan and topotecan. These agents stabilize the formed cleavable complex via the interfacial inhibition, which induces DNA damage that ultimately leads to cell death. However, de novo or acquired clinical resistance to these drugs is common [10]. To overcome such drawbacks, another line of drugs has emerged targeting topo IIα [11]. It was found that topo IIα-mediated ERBB2 co-amplification could lead to the development of tumors accompanied with an increase in the expression of topo IIα, which may be tight relative to the phenotypes of cancer cells [12]. Two types of topo Iiα-targeting agents were developed, including topo IIα poisons and catalytic inhibitors. Clinical drugs acting as topo IIα poisons are epipodophyllotoxin etoposide and anthracycline doxorubicin. They act by increasing the levels of covalent enzyme-cleaved DNA complexes [12]. Unfortunately, these drugs are linked to the development of acute myeloid leukemia through causing rearrangement at chromosomal band 11q23 [13]. On the other hand, it is believed that catalytic inhibitors of topo IIα do not lead to such side effects and could act as potential anticancer drugs.
Xestoquinone and halenaquinone (HQ), which are polycyclic quinone-type metabolites, were found to exhibit various biological activities, such as antifungal, cardiotonic, cytotoxic and topoisomerase activities, as well as acting as inhibitors for different protein kinases [14–18]. In addition, halenaquinone specifically inhibited the secondary DNA binding of RAD51, leading to the accumulation of chromosomal aberrations induced by unrepaired double-strand breaks [19]. Recently, Tsukamoto et al. suggested that halenaquinone inhibited RANKL-induced osteoclastogenesis by suppressing the NFκB and Akt signaling pathways [20]. HQ, the marine natural product isolated from the Petrosia sponge, is a broad spectrum tyrosine kinase inhibitor and more potent than xestoquinone, with a carbonyl group at the C-3 position. In this study, the cytotoxic and antitumor mechanisms of HQ were further investigated in a human leukemia Molt 4 cellular and xenograft animal model.
2. Results
2.1. Effect of HQ on Cellular and Tumor Growth in Vitro Assay and in Vivo Animal Model
The cytotoxicity of HQ was evaluated using the MTT assay against various human cancer cell lines.
HQ was found to inhibit the proliferation of Molt 4 (human acute lymphoblastic leukemia), K562
(human chronic myelogenous leukemia), MDA-MB-231(human breast adenocarcinoma) and DLD-1
(human colon adenocarcinoma) cancer cells, with IC
50of 0.18, 0.48, 8 and 6.76 μg/mL after 72 h, respectively. It exhibited the most potent activity against leukemia Molt 4 and K 562 cells (the values of IC
50were less than 4 μg/mL). These findings encouraged us to expand our cytotoxic study aiming to reveal the HQ mechanism of action against leukemia cancer cell lines. To pursue this goal, the cytotoxic effect of HQ against Molt 4 cells was determined after 24 h, resulting in IC
50values of 0.61 μg/mL. Furthermore, it was important to determine whether the cytotoxic effect of HQ is specific for cancer cells. The effect of HQ on the viability of rat normal lymphocytes was evaluated. The results indicated that even at the highest dose (2.5 μg/mL), HQ treatment caused only 26.32%
suppression in the viability of lymphocytes, but a significant decrease of about 70.78% and 31.33% in Molt 4 and K562 cells (Figure 1b). Thus, it may be concluded that HQ’s cytotoxic effect is more specific towards Molt 4 cells compared to K562 cells.
The in vivo anti-tumor activity of HQ was determined by evaluating its effect on the tumor growth of a human leukemia Molt 4 xenograft in an animal model. Molt 4 (1 × 10
5) cells were inoculated subcutaneously at the right flank of female immunodeficient athymic mice. After one month of treatment, the tumor growth of Molt 4 cells was significantly suppressed under the influence of HQ (1 μg/g) intraperitoneal injection. The average tumor size on Day 31 in the control group was 570.13 mm
3, whereas the average tumor size in the HQ-treated group was 211.29 mm
3(Figure 1c).
The tumor size was significantly lower in the HQ-treated group as compared to the control group (p = 0.002), with no significant difference in the mice body weights. At the end of the treatment, the tumor tissue was isolated and weighed. The average weights of the tumor were obviously less in the HQ-treated group (0.10 ± 0.04 g) compared to the control group (0.71 ± 0.36 g) (Figure 1d). These results suggested the anti-tumorigenic effect of the HQ in vivo xenograft model.
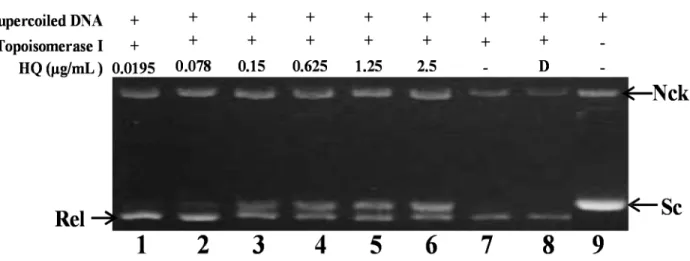
2.2. Effect of HQ on Topoisomerase I and II Activity
As previously described, the marine polyketide analog, xestoquinone, was identified as a topo I
inhibitor utilizing calf thymus proteins, which was used to determine the relaxation of ColE1
DNA [21]. To further confirm whether the other polyketide analog, HQ, could inhibit topo I or II
activity in the cell-free system, the purified human DNA topoisomerase protein was used to examine
the relaxation of supercoiled plasmid DNA [22]. Initially, the effect of HQ on topo I activity was
studied. A cell-free DNA cleavage assay using an enzyme-mediated negatively-supercoiled pHOT1
plasmid DNA was applied to study this effect. As shown in Figure 2, HQ at the lowest dose
(0.0195 μg) induced DNA relaxation in the presence of topo I (Lane 1); but at higher doses (0.078,
0.15, 0.625, 1.25 and 2.5 μg/mL), it inhibited the ability of topo I to convert supercoiled DNA to the
relaxed form (Lanes 2–6). HQ at doses of 0.0195, 0.078, 0.15, 0.625, 1.25 and 2.5 μg/mL significantly
decreased the relaxation of supercoiled DNA induced by topo I by 4%, 9%, 30%, 40%, 48% and 64%,
respectively, compared to the control supercoiled DNA. As expected, the relaxation of the supercoiled
DNA was inhibited by HQ in a dose-dependent manner, with an IC
50value of 1.19 μg/mL.
Doses of HQ (μg/mL, 24h)
(a) (b)
(c) (d)
Figure 1. Effect of halenaquinone (HQ) on cellular viability in vitro and tumor growth in vivo animal model. (a) Chemical structure of marine polycyclic quinone-type metabolites, HQ, isolated from Petrosia sp. Sponge; (b) Human leukemia Molt 4 and K562 cells, as well as normal rat lymphocytes were treated with HQ at different doses for 24 h.
The viability was determined by the MTT assay; (c) In vivo inhibition of tumor growth
with human leukemia Molt 4 xenograft by HQ. Female nude mice bearing leukemia Molt 4
tumors were treated with the solvent (negative control, n = 8) or HQ (1 μg/g, n = 8) for one
month. Tumor volumes were measured every other day, and the results are expressed as
the mean ± SD. * Significantly different from control groups at p = 0.015; (d) Histogram
of the tumor weight from the control group and HQ-treated group. Values are expressed as
the mean ± SD. ** Significantly different from control groups at p = 0.002. Representative
photos of the subcutaneous tumors, which were collected after treatment with the solvent
only (upper) or with HQ (lower) for 31 days.
Figure 2. Effect of HQ on topo I activity. Lanes 1–6: HQ (0.0195, 0.078, 0.15, 0.625, 1.25 and 2.5 μg); Lane 7: plasmid DNA + topo I; Lane 8: plasmid DNA + topo I + solvent control (induction of DNA relaxation); Lane 9: negative control plasmid DNA (supercoiled DNA). Nck, nicked DNA; Rel, relaxed DNA; Sc, supercoiled DNA.
As previously reported, the marine polyketide analogs, xestoquinone and halenaquinone, did not
inhibit the activity of calf thymus topo II [21]. Nevertheless, other xestoquinone analogs inhibited
topo I and II activity, and they did not intercalate with DNA [23]. In order to confirm HQ’s effect on
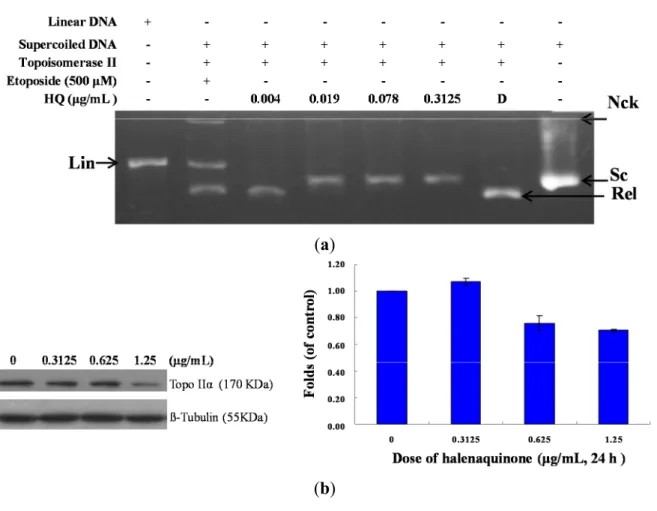
topo II activity and to understand its actual cytotoxic mechanism of action, a cell-free DNA cleavage
assay using an enzyme-mediated negatively-supercoiled pHOT1 plasmid DNA was applied. A linear
DNA strand was observed upon treating the supercoiled pHOT1 plasmid DNA with etoposide, a
standard topo II poison (Lane 5) [24]. The use of HQ in increasing concentrations (0.004, 0.019, 0.078
and 0.3125 μg/mL) completely inhibited DNA relaxation by 43%, 81%, 90% and 98%, respectively,
compared to the control supercoiled DNA and resulted in the formation of supercoiled DNA products
in the presence of topo IIα (Lanes 2–6) (Figure 3a). HQ showed comparable activity to adociaquinones
A and B [23], resulting in the inhibition of supercoiled DNA relaxation in a dose-dependent manner. In
addition, Western blotting indicated that the use of HQ (1.25 μg/mL) significantly diminished topo IIα
protein expression (Figure 3b). Furthermore, HQ inhibited topo I and II activities with IC
50of 1.19 and
0.0055 μg/mL, respectively as demonstrated by the cell-free system. These results suggested that HQ
could act as a dual inhibitor of topo I and II. To further precisely identify the effect of topo I and II, we
used the purified human protein, not calf, to examine the inhibitory effect of HQ.
(a)
(b)
Figure 3. Effect of HQ on topo II activity. (a) The effect of HQ on topo Iiα mediated supercoiled pHOT1 plasmid DNA relaxation with the cell-free system. Lane 1: linear DNA; Lane 2: positive control, etoposide (500 μM), as the topo II poison (induction of linear DNA); Lanes 3–6: HQ (0.004, 0.019, 0.078 and 0.3125 μg/mL); Lane 7: plasmid DNA + topo IIα (induction of DNA relaxation); Lane 8: negative control plasmid DNA (supercoiled DNA). Nck, nicked DNA; Rel, relaxed DNA; Sc, supercoiled DNA; Lin, linear DNA; (b) HQ decreased the expression of topo IIα protein in Molt 4 cells. Molt 4 cells were treated with HQ (0, 0.3125, 0.625 and 1.25 μg/mL) for 24 h. The protein expression of topo IIα was analyzed with Western blotting. The bands were quantified via densitometry and normalized relative to the β-tubulin levels.
2.3. Effect of HQ on Histone Deacetylase Activity
Our results demonstrated a significant inhibitory effect of HQ on the proliferation and
tumorigenesis of cancer cells (Figure 1). To evaluate whether HQ can inhibit epigenetic expression, a
cell-free HDAC colorimetric acetylated lysine side chain assay using an enzyme-mediated
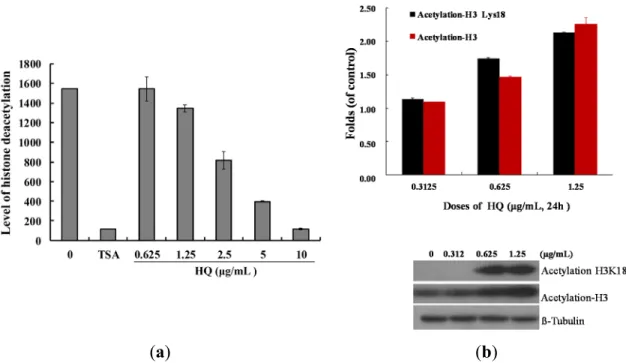
deacetylation was used. As shown in Figure 4a, HQ inhibited deacetylation of HDAC activity
compared to the solvent control with an IC
50value of 2.95 μg/mL, and the effect was in a
concentration-dependent manner. Next, the effect of HQ on the level of histone acetylation of Molt 4
cells was examined with Western blotting analysis. A significant acetylation of H3 was observed at
doses of 0.625 and 1.25 μg/mL after 24 h with an increase of 1.14- and 2.28-fold (acetyl-H3), as well
as 1.74- and 2.13-fold (acetyl-H3K18) compared to the control group, respectively (Figure 4b).
(a) (b)
Figure 4. Effect of HQ on histone deacetylase activity. (a) The effect of HQ on HDAC mediated deacetylation of lysine side chain with the cell-free system. Trichostatin A (TSA) is a potent reversible inhibitor of HDAC and was used as the positive control. Relative deacetylation of histone was progressively deceased by the treatment of HQ (0.625, 1.25, 2.5, 5 and 10 μg/mL); (b) HQ increased the expression of acetyl-H3 and acetyl-H3K18 in Molt 4 cells. Molt 4 cells were treated with HQ (0, 0.3125, 0.625 and 1.25 μg/mL) for 24 h. The protein expression was analyzed with Western blotting. The bands were quantified via densitometry and normalized relative to the β-tubulin levels.
2.4. Effect of HQ on Apoptosis Induction Involved Mitochondrial Dysfunction in Molt 4 Cells
Previous studies demonstrated a significant cytotoxicity of HQ against several cancer cells [14,17,21]. To examine whether the cytotoxic effect of HQ on K562 and Molt 4 cells involves apoptotic induction, the cells were treated with HQ, and the apoptotic-related proteins (PARP, caspases 3 and 7) were analyzed with Western blotting. As indicated in Figure 5a, the treatment with HQ caused cleavage of PARP at low doses (0.3125 and 0.625 μg/mL) in both types of cells after 24 h.
In Molt 4 cells, HQ resulted in an activation of caspases 3 and 7 in a dose-dependent manner. On the other hand, caspases 3 and 7 in K562 cells were activated only after the treatment with 1.25 μg/mL of HQ. To better comprehend the antitumor mechanism of HQ, the apoptotic cells population was determined using Annexin V/PI and DAPI staining assay in Molt 4 cells. After 24 h of treatment, the percentage of apoptotic cell population was significantly increased by 26.23%, 39.93% and 70.27%
compared to the solvent control (Figure 5b). To assess the nuclear morphological change induced by
HQ, the Molt 4 cells were further examined with DAPI staining and observed under a fluorescence
microscope. The results showed that HQ treatment increased the number of condensed nuclei
compared to the control, which exhibited intact and normal nuclei (Figure 5c). To address whether the
apoptotic induction of HQ was related to the mitochondrial pathway, JC-1 fluorescent dye was used to
determine the mitochondrial membrane potential. Molt 4 cells were treated with different doses of HQ
for 24 h and then stained with JC-1. As shown in Figure 5d, the use of HQ (0.3125 μg/mL) increased the population of Molt 4 cells with disrupted membrane potential from 6.03% up to 16.36%. This effect was dramatically increased upon the treatment with HQ at 0.625 and 1.25 μg/mL, which resulted in 32.96% and 53.06% cells with disturbed MMP, respectively. To further explore the mechanism of HQ-induced apoptosis, the effect of HQ on apoptotic- and mitochondrial metabolism-related proteins was evaluated.
(a) (b)
(c)
Figure 5. Cont.
(d)
(e)
(f) Figure 5.
Cont.
3
2.5
Caspase 8 Caspase 9
*
*
2
1.5
1
0.5
0 0.3125 0.625 1.25
Doses of halenaquinone (μg/mL, 24 h)
(g)
Figure 5. Apoptotic effect of HQ on Molt 4 cells. Cells were treated with the indicated concentration of HQ for 24 h. (a) HQ treatment enhanced the cleavage of apoptosis-related proteins in K562 and Molt 4 cells; (b) The change of nuclear morphology was determined with DAPI staining and observed using fluorescent microscope; (c) Apoptosis induction and (d) mitochondrial membrane potential were assessed with annexin V/PI and JC-1 staining using flow cytometric analysis; (e) Proapoptosis-and (f) Survival-related proteins were determined with Western blotting. The results are presented as means ± SD of three independent experiments (* p < 0.05; ** p < 0.01); (g) After treatment, cytoplasmic lysates (50 μg) were prepared and the enzymatic activity of caspases 8 and 9 like protease was determined by the incubation with colorigenic substrates, for 2 h at 37 °C. The release of chromophore pNA was monitored spectrophotometrically (405 nm).
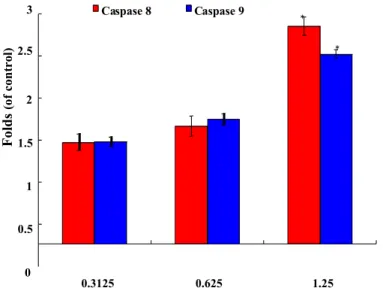
As shown in Figure 5e, HQ treatment eliminated the expression of the anti-apoptotic proteins Bcl-2 and Bid. Concomitantly, HQ treatment induced the expression of cytochrome c and the pro-apoptotic protein, Bax. It increased the expression of cytochrome c 1.73- and 2.19-fold, as well as Bax expression 1.8- and 2.03-fold at doses of 0.625 and 1.25 μg/mL, respectively. Moreover, the treatment of leukemia Molt 4 cells with different concentrations of HQ diminished the expression of p-Akt (Ser473), p-PTEN (Ser380), p-GSK3β (Ser9), p-PDK1 (Ser241) and cytosolic hexokinase II, but enhanced the expression of cytosolic hexokinase I about 1.34-fold compared to the control group at the high dose of 1.25 μg/mL (Figure 5f). Caspase-3 is a key executioner of apoptosis, which is activated by an initiator caspase, such as caspases 8 or 9. These activated caspases cleaved many cellular substrates, ultimately leading to cell apoptosis [25]. To explore whether the mechanism of the apoptotic effect of HQ involved the activation of caspase initiator, the cleavage of caspases 8 and 9 was tested using the calorigenic assay. The treatment of Molt 4 cells with the 0.3125, 0.625 and 1.25 μg/mL of HQ led to an increase of the active forms of caspase 8 by about 1.2-, 1.4- and 2.6-fold, as well as the active forms of caspase 9 by about 1.2-, 1.5- and 2.3-fold, respectively (Figure 5g).
Folds (of control)
2.5. HQ Induced Apoptosis Is Mediated by Excessive ROS Generation
One previous study demonstrated that N-acetyl-
L-cysteine, an antioxidant agent, could completely attenuate apoptosis induced by HQ in prostate PC12 cancer cells. The structure of this compound suggests its potential as an antioxidant agent [17]. To examine whether the HQ-induced apoptosis in Molt 4 cells involves the overproduction of ROS, the levels of ROS at different times following HQ treatment were determined. A time-dependent increase in ROS generation was monitored using the carboxy derivative of fluorescein, carboxy-H
2DCFDA dye. As shown in Figure 6a, HQ treatment (1.25 μg/mL) for 10, 30, 40 and 50 min resulted in 2.67-, 2.87-, 2.27- and 1.05-fold increases in the ROS levels, respectively, as compared with the mean fluorescence index (MFI) of the control. To clarify whether ROS generation is the major regulatory factor in HQ-induced apoptosis, Molt 4 cells were pretreated with 1.5 mM N-acetyl-
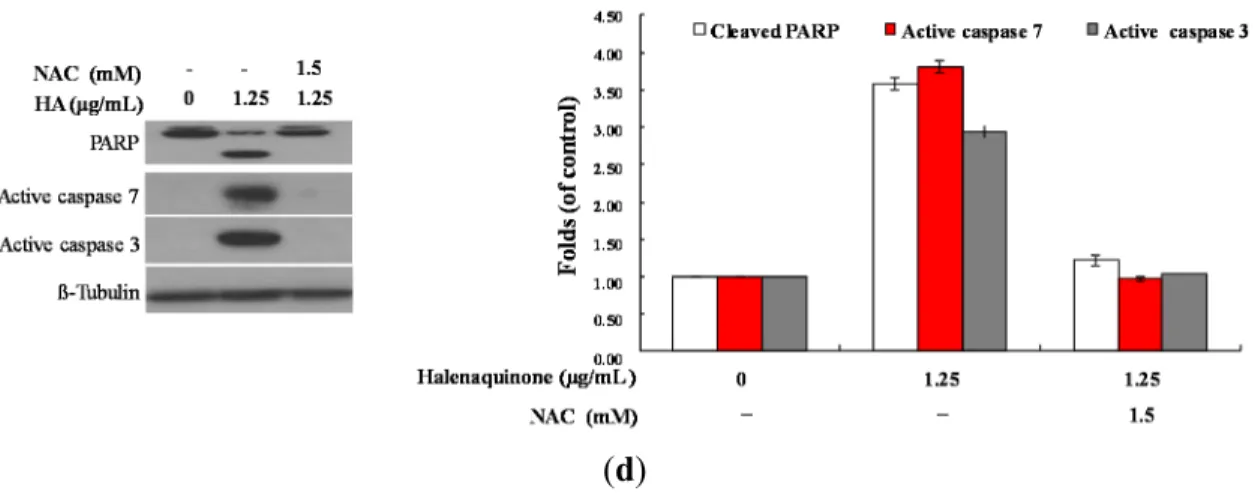
L-cysteine (NAC), an ROS scavenging agent, aiming to suppress the intracellular oxidative stress. The apoptotic population was measured via Annexin V/PI staining after treatment. As shown in Figure 6b, the result of NAC treatment is similar to the negative control group, showing less than 5% of the apoptotic population (data not shown). In addition, the pretreatment with 1.5 mM NAC decreased the apoptotic cell population from 38.63% and 69.6% to 11.45% and 11.33% in response to the use of 0.625 and 1.25 μg/mL of HQ, respectively. These results indicated that blocking the oxidative stress by NAC resulted in saving Molt 4 cells from apoptosis induced by HQ. To further confirm if the MMP disruption induced by HQ is initiated by ROS overproduction, the population of the cells with disturbed MMP in response to HQ treatment with or without NAC pretreatment was determined. The determination of the cell population with disturbed MMP was achieved utilizing a cationic dye, JC-1 (Figure 6c). Cells were divided into four groups, in which two groups were only treated with 0.625 and 1.25 μg/mL of HQ, whereas the other two groups were treated with NAC (1.5 mM) followed by 0.625 or 1.25 μg/mL of HQ. After 24 h, the change in the cell population with disturbed MMP was evaluated. The NAC pretreatment diminished the population of cells with disturbed MMP from 34.3% and 53.02% to 11.9% and 12.4% in response to the treatment with 0.625 and 1.25 μg/mL of HQ, respectively. Furthermore, the effect of NAC pretreatment on the expression of the apoptosis-related proteins was determined. As shown in Figure 6d, NAC pretreatment abrogated PARP cleavage, as well as the activation of caspases 3 and 7.
In agreement with the preceding results of Annexin V/PI staining, these findings indicated that the
cytotoxic effect of HQ in Molt 4 cells is mediated through apoptotic induction, as well as
mitochondrial dysfunction involved in ROS production.
3.5 3 2.5 2 1.5 1 0.5
0 0 10 30 40 50
Time of treatment, mins
(a)
90 55.30%
**
80 * p < 0.05 70 ** p < 0.01 60
50 40 30 20 10 0
27.23%
*
Halenaquinone (μg/mL) 0 0.625 0.625 1.25 1.25
NAC (mM) - - 1.5 - 1.5
(b)
7060 ** p < 0.01 50
40 30 20 10
**
22.38%
**
40.55%
Halenaquinone (μg/mL0 )
NAC (mM)
(c)
Figure 6. Cont.
0 0.625 0.625 1.25 1.25
- - 1.5 - 1.5
Folds (of control) Population of MMP disruption (%) Apoptotic population (%)