Copyright © 2015. All rights reserved.
1
Saturated fatty acid induces cancer stem cell-like properties in human hepatoma cells
L-W. Chong
1, R-H. Chou
2,3, C-C. Liao
4, T-F. Lee
5, Y. Lin
1, K-C. Yang
1and Y-C. Hsu
61 Division of Hepatology and Gastroenterology, Department of Internal Medicine, Shin Kong Wu Ho-Su Memorial Hospital, Taipei, Taiwan
2 Graduate Institute of Cancer Biology and Center for Molecular Medicine, China Medical University, Taichung, Taiwan
3 Department of Biotechnology, Asia University, Taichung, Taiwan
4 Proteomics Research Center, National Yang-Ming University, Taipei, Taiwan
5 Institute of Clinical Medicine, School of Medicine, National Yang Ming University, Taipei, Taiwan
6 Institute of Biomedical Sciences, Mackay Medical College, New Taipei City, Taiwan
Corresponding author: Yi-Chao Hsu Ph.D., Institute of Biomedical Sciences, Mackay Medical College. No.46, Sec. 3, Zhongzheng Rd., Sanzhi Dist., New Taipei City 252, Taiwan . E-mail: [email protected]
Abstract
Hepatic steatosis has been reported to be a risk factor for the development of liver cancer. The underlying mechanism for carcinogenesis remains to be elucidated.
It has been postulated that cancer stem cells (CSCs) within tumor tissues are a subset of cells with stem cell properties of self-renewal and undifferentiation. The purpose of this study was to investigate the effects of a saturated fatty acid, palmitate (PA), on CSC-like properties of human hepatoma HepG2 cells. We investigated the effects of PA on HepG2 cells and primary rat hepatocytes (PRH) by exposing them to PA to induce lipid accumulation. Significant fat accumulation was obser- ved by Oil Red O staining in cells exposed to PA, and it was accompanied by significant increase in NFκB (p65) nuclear translocation in HepG2 cells. Notably, PA significantly enhanced the sphere forming ability of HepG2 cells, but not PRH. Furthermore, PA significantly increased stemness gene expressions of Sox2 and Oct4, and sonic hedgehog (Shh) production. Notably, NFκB inhibitors, N-Acetyl-L-cysteine and pyrollidine dithiocarbamate, and a NOX inhibitor, diphenyleneiodonium, significantly attenuated PA-induced sphere forming ability of HepG2 cells. Our results suggest that lipid accumulation may not only induce pro-inflammatory res- ponses in hepatocytes but may also activate CSC-like properties of hepatoma cells through NFκB activation.
Key words: Palmitate, cancer stem cells, NFκB.
Introduction
The prevalence of non-alcoholic fatty liver disease (NAFLD) is rising rapidly because of ongoing epide- mics of obesity and type 2 diabetes, which become a public health issue. The spectrum of NAFLD starts from simple steatosis to steatohepatitis, liver fibrosis, cirrhosis and finally progresses to hepatocellular car- cinoma (1). According to the two-hit-hypothesis of non-alcoholic steatohepatitis (NASH), hepatic steatosis (“first hit”) is a prerequisite for subsequent pathologic events (“second hits”), including environmental and genetic factors that result in liver injury (2). Free fatty acids (FFAs) are the major mediators of lipid accumu- lation in the liver. In NAFLD patients, plasma levels of circulating FFAs are found to be correlated with disease severity (3). Hepatocytes will be damaged when excess FFAs in form of triacylglycerols (TGs) is stored in lipid droplets. Lipid overload will further result in oxidative stress, mitochondrial dysfunction and pro-inflammatory responses (“second hits”) (3), leading to inflammation and fibrogenesis in NASH. Recent studies showed that nearly a quarter of NASH patients develop cirrhosis, which is eventually at high risk in developing hepato- cellular carcinoma (HCC).
Initiation of cancer may be the result of mutations occurring in stem cells, which interfere with the diffe- rentiation of these cells. Many common properties of the stem cells and some tumor cells suggest that can- cer stem cells (CSCs) may be responsible for the ini- tiation and progression of cancer. The special property of CSCs is the ability for self-renewal and cell proli-
feration (spherogenesis), which are the major causes of cancer recurrence and metastasis. Sonic Hedgehog (Shh) signaling pathway and stemness-connected trans- cription factors, such as Sox2 and Oct4, are primarily responsible for CSC proliferation (4-6). Although the mechanisms of carcinogenesis remains to be clarified, it has been proposed that CSCs may participate in the initiation, progression and metastasis of HCC, and pos- sibly result in the failure of chemo- and radiotherapy.
Notably, the sphere formation capacity of cancer cells is a functional approach to enrich the potential CSC subpopulations when the specific markers for CSCs are still controversial (7). Sphere culture in liver cancer was established as a functional approach to isolate, identify and characterize liver CSCs (7).
In this study, we investigated the effects of a satura-
ted fatty acid, palmitate (PA), on CSC-like properties
in HepG2 cells. Our results demonstrated that exposure
of hepatocytes to PA resulted in NFκB activation and
inflammatory cytokine expression in HepG2 cells. Fur-
thermore, PA significantly enhanced the sphere forming
ability associated with the significant increase of stem-
ness gene expressions of Sox2 and Oct4, and Shh pro-
duction in HepG2 cells. Notably, NFκB inhibitors, N-
Acetyl-L-cysteine (NAC), and pyrollidine dithiocarba-
mate (PDTC), and a NADPH oxidase (NOX) inhibitor,
diphenyleneiodonium (DPI), could significantly reduce
PA-induced sphere forming ability of HepG2 cells. Our
results suggest that lipid accumulation not only induced
NFκB activation, but also activate CSC-like properties
of HepG2. This study elicits the possibility that satura-
ted fatty acid, such as PA, might activate CSC-like pro-
L-W. Chong et al. / Palmitate induces cancer stem cell properties in hepatoma cells.
perties of hepatoma cells in NAFLD patients through NFκB pathway. Thus, this study provides a promising model to study the mechanisms of carcinogenesis in NAFLD and to develop chemopreventive therapies for NAFLD.
Materials and methods
Cell culture (human HepG2 cells and primary rat he- patocyte, PRH)
The human hepatoma cell line, HepG2, was grown in Dulbecco’s modified Eagle’s medium (DMEM) with 10% heat-inactivated fetal calf serum (FBS) at 37°C in a humidified atmosphere with 5% CO . The HepG2 cells were passaged in the T75 flask and maintained in DMEM containing 10% FBS.
O was added with agitation for 7 minutes, followed by washing in 85% propylene glycol. The dishes were then rinsed in distilled water and counterstained with hema- toxylin. For each dish, three images were photographed, and a representative image is shown.
Transiently transfected cells and luciferase assays Cell were seeded at a density of 10
5cells/well on 24‐
well plates the day before transfection. Plasmid NFκB‐
Luc (1 μg/well) (Strategene, La Jolla, CA, USA) and pRL‐SV40 (0.2 μg/well) (Promega, Madison, USA) were transfected into cells by lipofectamine (Invitrogen, California, USA). The pNFκB‐Luc consists of NFκB‐
binding region, followed by the reporter gene firefly luciferase. Plasmid pRL‐SV40 served as an internal control to normalize the transfection efficiency (10, 11).
After treatment with PA for 6 h in a 5% CO incubator Primary hepatocyte isolation and culture
Hepatocytes were isolated from male Wistar rats (275-300 g) using a two-stage non-recirculating perfu- sion method described previously (8, 9). Livers were perfused for 10 min via the portal vein with Ca
2+-, Mg
2+-free Herb’s buffer, containing 1% antibiotics, fol- lowed by buffered with antibiotics and 0.25 mg/ml col- lagenase Type IV (435 U/mg; Sigma; MO, USA) for at least 7 minutes. The minced liver was passed through two layers of cheesecloth and hepatocytes were re- suspended in Herb’s buffer and filtered through nylon (62 mesh). After centrifugation for 5 min at 100 x g, hepatocytes were re-suspended in DMEM and filtered through two layers of nylon. Hepatocytes were added to 6-well plates coated with collagen film (Vitrogen 100;
at 37 °C, the cells were harvested and lysed in 100 μL of lysis reagent. Twenty mircoliter of cell lysate was then mixed with 100 μL of luciferin (the substrate of lucife- rase) right before luminescence detection. The lumines- cence, generated by luciferase activity, was measured with an AutoLumat LB953 (Berthold Technologies, Bad Wildbad, Germany). All reagents for luciferase assays were purchased from Promega (Madison, USA).
Western blotting analyses for Shh, NFκB (p65) nu- clear translocation
Cells (5 × 10
6) were seeded in medium containing 10% FBS. After 24 h, the cells were washed twice with PBS and the medium was replaced by serum‐free medium. PRH or HepG2 cells (in serum‐free medium) Collagen Corp., Palo Alto, CA, USA) at 2 x 10
5cells/ were treated with PA for 6 h in 5% CO incubator at well density. After 4 hours, unattached cells were remo-
ved and fresh growth medium was applied. During 20 h incubation, cells were treated with PA for indicated time and concentration as described.
Chemicals
Palmitic acid, albumin (BSA), N-Acetyl-L-cys- teine (NAC), pyrollidine dithiocarbamate (PTDC) and diphenyleneiodonium (DPI) were purchased from Sig- ma-Aldrich. Palmitate (PA; 102553; Sigma) stock solu- tion was carried out as described previously (10, 11).
Briefly, a 400 mM PA stock solution was prepared in DMSO. A filtered 3% (w/v) FFA-free BSA (9048-46-8;
Sigma) solution was prepared in DMEM and maintained at 55 °C in a water bath. 4 mM FFA/BSA solution was obtained by mixing the appropriate amount of PA stock solution to 3% BSA at 55 °C for another 30 min. FFA/ BSA solution was diluted with DMEM and adjusted to the concentration for treatment.
The above solution was fresh prepared before use. The 62.5 mM NAC (A9165; Sigma) stock solution, 50 mM PDTC (P8765; Sigma) stock solution and 5 mM DPI (D2926; Sigma) stock solution were prepared in ddH O and stored in -20
oC.
Oil Red O Staining
Sub-confluent monolayers of HepG2 cells were exposed to PA or BSA for 12 hours. Cells were stai- ned with Oil-Red-O to examine the amount of fat accu- mulation in the cells. Briefly, dishes were washed with
cold phosphate-buffered saline and fixed in 10%
neutral formalin. After 2 changes of propylene glycol, Oil-Red-
2
2
2
2
L-W. Chong et al. / Palmitate induces cancer stem cell properties in hepatoma cells.
37 °C. After treatments, nuclear extracts containing the NFκB active protein were prepared from cells using a nuclear extraction kit (Chemicon, Temecula, CA, USA) according to the manufacturer’s instructions and our published methods (10, 11). Twenty microgram of pro- teins in cytoplasmic fraction or nuclear fraction was separated on a 10% SDS-PAGE and transferred onto Immobilon‐PVDF (Millipore, Bedford, MA, USA) in a transfer buffer (6.2 mM boric acid, pH8.0). The blots were incubated initially with blocking buffer (5% BSA) for 1 h at room temperature, and then with specific pri- mary antibodies against α‐actin (GTX100095, Gene- Tex, Taiwan), α‐tubulin (sc-8035, Santa Cruz), PCNA (sc-56, Santa Cruz) or NFκB (p65) (MAB3026, Santa Cruz) and Shh (MABD175, Millipore). Primary anti- bodies had been diluted (1:10000) with Tris‐buffered saline‐Tween 20 (TBS‐T) containing 1% BSA. After antibody incubation, the blots were washed with TBS‐T and incubated with HRP conjugated secondary antibo- dies (1:2000) (Santa Cruz) for 1 h at room temperature.
After washing the secondary antibodies with TBS‐T, immune-detection was performed by Luminescence Imaging System (FUJI las-4000mini, Japan). Film ex- posure ranged from a few seconds to 5 minutes. Bands were quantified using Kodak Image Analysis Software (Rochester, NY, USA).
Semi-quantitative and quantitative reverse transcrip- tase polymerase chain reaction (RT-PCR)
Semi-quantitative (for ICAM1, IL8, TNF-α, Shh)
and quantitative RT-PCR (for Sox2, Oct4, Nanog) as-
says were employed to assess the relative levels of the mRNA expression of PA-stimulated HepG2 cells and PRH (10-12). Total RNA was isolated from HepG2 cells or PRH by the method of Chomczynski and Sacchi (13). For cDNA synthesis, 1 mg of total RNA was reverse- transcribed in a 30 ml of reaction mixture containing
10 mM dNTP mix, 500 µg/ml oligo(dT), 0.2 mM di- thiothreitol, 40 units of RNase inhibitor, 200 units of M-MLV reverse transcriptase, and 5x buffer (with 1.5 mM MgCl ) (Invitrogen, USA). The reaction mixture was incubated at 37 °C for 60 minutes and then denatu- red at 65 °C for 10 minutes. Quantitative PCR analysis was performed using an ABI prism 7900 HT Sequence Detection System (Applied Biosystems). The specific primers used in this study were described in Table 1.
Sphere formation assay
PRH or HepG2 cells were maintained as a mono- layer in high glucose DMEM with 10% FBS, 100-IU/
ml penicillin G and 100-μg/ml streptomycin at 37 °C
sphere may be due to the aggregation of several small spheres. Therefore, we investigated the effects of PA on the sphere formation of HepG2 cells by counting the numbers of spheres in two different sizes.
Data analysis
Data are expressed as the mean ± SEM. One‐way analysis of variance (ANOVA) was used for comparison of biochemical and molecular parameters.
A non‐para- metric method (the Dunn procedure under the Kruskal– Wallis test) was used for multiple pairwise comparisons between groups for the histological grades of fibrosis. Statistical significance was accepted at p <
0.05.
Results
Effects of PA on NFκB (p65) nuclear translocation and NFκB luciferase activity in HepG2 cells
We have previously demonstrated that lipid accu- mulation in the liver led to hepatic inflammation and in a humidified 5% CO incubator. Cells were collec- fibrogenesis through NFκB pathway (12). To clarify ted and washed to remove serum, then suspended in
serum-free DMEM/F12 supplemented with 100 IU/ml penicillin, 100 μg/ml streptomycin, Dulbecco’s modi- fied Eagle’s medium HG/F-12 supplemented with B27 (Invitrogen), 50 ng/ml epidermal growth factor, 20 ng/
ml FGF2, and 5 μg/ml heparin, 2% B27, 1% N2 supple- ment (Invitrogen, Carlsbad, CA, USA). The cells were subsequently cultured in ultra low attachment 6 cm dis- hes (Corning Inc., Corning, NY, USA) at a density of no more than 1x 10
4cells/well in 5 ml sphere medium.
if NFκB-mediated hepatic inflammation contributes to any effect on cancerous cells in NASH patients, we in- vestigated the effects of saturated fatty acid PA on NFκB (p65) nuclear translocation and NFκB luciferase activi- ty in HepG2 cells. The results showed that PA (200 μM) induced NFκB (p65) nuclear translocation and NFκB luciferase activity in HepG2 cells significantly (Figures
1a, *p<0.05). Interestingly, PA (200 μM)-induced NFκB (p65) nuclear translocation was also regulated in a time- dependent manner, reaching a maximum at 6 h after PA
Cells were cultured in 5% CO in a 37 °C incubator. The treatment (Figure 1b, **p<0.01). The functional effect spheres (diameter between 70-150 μm and larger than
150 μm) were both counted directly under microscope after 7 days in vitro (7). Due to the fact that cell aggre- gation may affect the numbers of spheres. One large
Table 1.
of PA on NFκB activity was further examined by the lu-
Figure 1. Effects of PA on NFκB (p65) nuclear translocation and promoter activity of HepG2 cells. (a) PA [10-200 μM] concen- tration-dependently increased NFκB (p65) nuclear translocation in HepG2 cells. PA [200 μM] significantly increased NFκB (p65) Target genes Primer Sequences for Human Genes
GAPDH Forward: 5’-GAAGGTGAAGGTCGGAGTC-3’
Reverse: 5’-GAAGATGGTGATGGGATTTC3’
IL-8 Forward: 5’-ATGACTTCCAAGCTGGCCGTGGCT-3’
Reverse: 5’-TCTCAGCCCTCTTCAAAAACTTCTC-3’
ICAM-1 Forward: 5’-GGCTGGAGCTGTTTGAGAAC-3’
Reverse: 5’-TCACACTGACTGAGGCCTTG-3’
Shh Forward: 5’-GGCTGGAGCTGTTTGAGAAC-3’
Reverse: 5’-TCACACTGACTGAGGCCTTG-3’
TNF-α Forward: 5’-CCTGTGAGGAGGACGAACAT-3’
Reverse: 5’-GAGGAAGGCCTAAGGTCCAC-3’
hSox2 Forward: 5’- CGCAGACCTACATGAACG-3’
Reverse: 5’- CCCTGGAGTGGGAGGAA-3’
hOct4 Forward: 5’- AAAGCAGAAACCCTCGT-3’
Reverse: 5’- TCCAGGTTGCCTCTCAC-3’
hNanog Forward: 5’- GAGACAGAAATACCTCAGCC-3’
Reverse: 5’- TCTGCGTCACACCATTG-3’
Target genes Primer Sequences for Rat Genes rGAPDH Forward: 5’-AGCCCAGAACATCATCCCTG-3’
Reverse: 5’-CACCACCTTCTTGATGTCATC-3’
rIL-8 Forward: 5’-AGGGATTCACTTCAAGAACA-3’
Reverse: 5’-GCATCTTTTGGACAATCTTC-3’
rICAM-1 Forward: 5’-CACTAGAGGAGTGAGCAGGTTAACAT-3’
Reverse: 5’-TATGACTCGTGAAAGAAATCAGCTCTT- rTNF-α 3’Forward: 5’-GCCAATGGCATGGATCTCAAAG-
3’ Reverse: 5’-CAGAGCAATGACTCCAAAGT-3’
2
2
2
nuclear translocation in HepG2 cells (n=3). **p<0.01 vs. control.
(b) PA [200 μM] time-dependently increased NFκB (p65) nuclear trans- location in HepG2 cells. (c) PA [200 μM] significantly
increased NFκB promoter activity in HepG2 cells (n=3). *p<0.05 vs. control.
Figure 2. Effects of palmitate on mRNA expressions of proin- flammatory genes (ICAM-1, IL-8, TNF-α) in HepG2 cells. (a, b) PA treatment significantly induced the mRNA expressions of ICAM-
1, IL-8, TNF-α genes in HepG2 cells (n=3). *p<0.05 vs. control;
**p<0.01 vs. control.
ciferase activity assay. PA induced the NFκB-mediated promoter activity in a dose-dependent manner (Figure 1c). Taken together, the results suggest that PA could in- duce translocation of NFκB (p65) into nucleus, leading to transcriptional activation of the downstream genes of NFκB in regulation of cellular functions.
Effects of PA on mRNA expression of pro-inflamma- tory genes (ICAM‐1, IL-8 and TNF-α) in HepG2 cells To clarify if NFκB-mediated pro-inflammatory cyto-
kines contributes to any effect on cancerous cells pa- tients, we investigated the effects of saturated fatty acid PA on mRNA expression of pro-inflammatory genes (ICAM‐1, IL-8 and TNF-α) in HepG2 cells. PA (200 μM) significantly stimulated the mRNA expression le- vels of ICAM‐1, IL-8 and TNF-α genes in HepG2 cells (Figures 2a and 2b), consistently suggesting that lipid accumulation might induce inflammatory responses.
Effects of PA on the expression of stemness genes (Oct4, Sox2, Nanog and Shh) in HepG2 cells
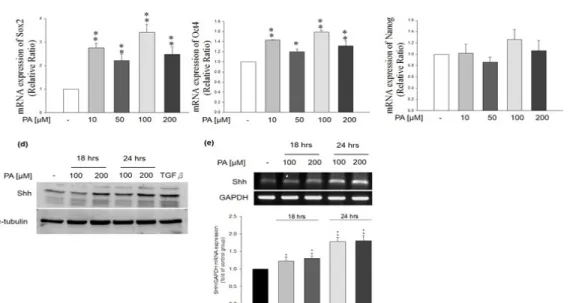
To clarify if PA contributes to any effect on cancerous cells, we further investigated the effects of PA on the expression of stemness genes (Oct4, Sox2, Nanog and Shh) in HepG2 cells. PA treatment significantly induced the mRNA expression of stemness genes, such as Sox2 (Figure 3a, n=3, **p<0.01) and Oct4 (Figure 3b, n=3,
*p<0.05, **p<0.01). Protein and mRNA expressions of Shh were also both up-regulated in PA-treated HepG2 cells (Figures 3d and 3e, n=3, *p<0.05, **p<0.01, res- pectively). In contrast, PA treatment did not alter the mRNA expression of Nanog (Figure 3c, n=3). The data suggested that PA might potentially induced self-re- newal and tumor-initiation CSC-like properties of HCC cells.
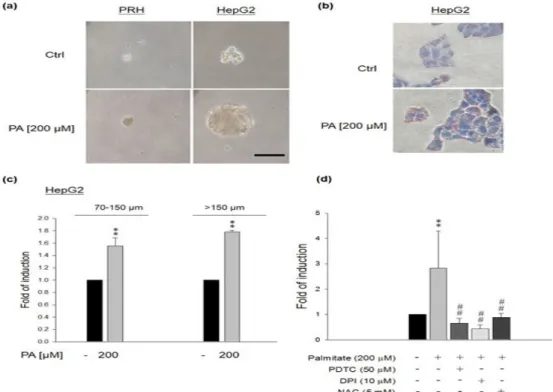
Effects of PA on sphere formation abilities of HepG2 cells, but not normal PRH
We hypothesized that PA could enhance CSC-like
property in cancerous cells (HepG2) rather than nor-
mal cells (PRH). Therefore, we compared the sphere
formation ability between these two cell lines. The re-
sults showed that PRH did not have sphere formation
after PA (200 μM) treatment (Figure 4a). However, PA
(200 μM) significantly enhanced the sphere formation
of HepG2 cells (Figure 4a). Lipid accumulation was
observed in HepG2 cells by exposing them to PA (200
μM) to simulate the excessive influx of fatty acids into
cells. Significant fat accumulation was observed by Oil
Red O staining in cells exposed to PA (Figure 4b). PA
could induce significantly more spheres of HepG2 cells
(Figure 4c, n=4, **p<0.01). Furthermore, NAC, PDTC
Figure 3. Effects of PA on stemness gene expression in HepG2 cells. PA treatment significantly induced the mRNA expression of (a) Sox2 (n=3).
**p<0.01, (b) Oct4 (n=3). *p<0.05, **p<0.01. (c) PA treatment did not alter the mRNA expression of Nanog. (d) PA treatment significantly induced the protein production of Shh. (e) PA treatment significantly induced the mRNA expression level of Shh (n=3, *p<0.05, **p<0.01).
Figure 4. Effects of PA on sphere formation in PRH and HepG2 cells. (a) PA (200 μM) significantly enhanced the sphere formation of HepG2 cells, but not PRH. (b) Lipid accumulation was observed in HepG2 cells by exposing them to PA (200 μM) to stimulate the excessive influx of fatty acids into cells. Significant fat accumulation was observed by Oil Red O staining in cells exposed to PA. (c) PA could induce significantly more spheres in HepG2 cells (n=4, **p<0.01). (d) NAC, PDTC and DPI could attenuate palmitate-induced sphere formation in HepG2 cells (n=4,
**p<0.01 vs. control, ##p<0.01 vs. PA treatment).
and DPI could attenuate PA-induced sphere formation in HepG2 cells (Figure 4d), indicating that PA could induce the sphere formation of HepG2 cells through NFκB and NOX activation.
Discussion
In this study, we demonstrated a potential mechanism for the pathophysiological role of hepatic lipid accumu- lation during liver carcinogenesis. Our previous study demonstrated that PRH and HepG2 cells incubated with PA showed increased ROS production, NOX expression (12). Conditioned media from PRH treated with PA led to the activation of HSC-T6 cells and fibrogenesis (12). In this study, we showed that PA significantly induced the activation of NFκB signaling pathway, such as NFκB (p65) nuclear translocation and promoter activity (Figure 1), and mRNA expression of proinflammatory genes (Figure 2) in HepG2 cells. In particular, PA signi- ficantly enhanced stemness gene expression and Shh production in HepG2 cells (Figures 3). Moreover, PA also significantly increased sphere formation in HepG2 cells, which could be inhibited by NFκB and NOX inhi- bitors (Figure 4). In our previous study, increased ROS formation was observed in PA-treated PRH and HepG2 cells (12). Oxidative stress (OS) causes lipid peroxida- tion in cell membranes. Lipid peroxidation products can induce inflammatory responses with up-regulation of pro-inflammatory cytokines (14). Our results demons- trated increased ICAM-1, IL-8 and TNF-α gene trans- cripts in HepG2 cells after PA treatment (Figure 2). The elevated pro-inflammatory gene expression could be a result of OS induced by lipid accumulation (15). PA has
also been demonstrated to induce IL-8 production from hepatocytes and HepG2 cells (16).
Saturated free fatty acids have been suggested to
play important roles in many biological processes such as hormone production and initiation of signaling pa- thways downstream the cellular membrane (17). Palmi- toylation is one of the most important posttranslational modifications in cellular proteins that is mediated by a membrane-bound palmitoyl acyl transferase (17). Pal- mitoylation has been shown to enhance the hydrophobi- city and membrane affinity of the target protein. It also involves in regulating cytosolic proteins’ trafficking, stability and sorting. Dysregulation of palmitoylation process has been shown in different cancer types such as colon cancer (18), prostate cancer (19) and breast cancer (20, 21). The Wnt and Hedgehog (Hh) pathways have been suggested their importance in embryonic develop- ment and the maintenance of CSCs (22). Hh ligand ove- rexpression is correlated with HCC (23), pancreatic (24) and breast cancers (25). Notably, palmitoylation of Wnt and Hh determines their signaling activity (22). Further- more, palmitoylation of Wnt is critical for the binding capacity to its Frizzled receptors in cell surface and induce downstream stabilization of β-catenin (26). In addition, palmitoylation of Hh facilitates its binding to the cell surface, which mediates its interaction with the receptor Patched to activate the downstream Gli trans- cription factors (27). In this study, our data showed that PA treatment induced the mRNA and protein expression of Shh in HepG2 cells (Figure 3). Future efforts in iden- tifying the palmitoylated proteins in PA-treated HepG2 cells will help elucidate the mechanism of PA-induced CSC properties.
Recently, the sphere formation capacity of cancer
cells has been demonstrated to enrich the potential CSC
populations. Sphere culture in liver cancer was also
established as an functional approach to isolate, iden-
tify and characterize liver CSCs (7). In this study, we
observed that PA significantly increased the numbers
of spheres in HepG2 cells, but not in PRH (Figure 4a).
This phenomenon suggested that lipid accumulation might enhance the CSC-like properties, such as Sox2 (Figure 3a), Oct4 (Figure 3b), Shh expressions (Figure 3d and 3e) and sphere forming ability (Figures 4a and 4c) in HepG2 cells. Not surprisingly, PRH has no CSC properties and could not form spheres under sphere culture conditions (Figure 4a). On the other hand, the human hepatoma HepG2 cells showed enhanced CSC- like properties in the presence of PA. In particular, the positive correlation between NFκB (p65) and Shh expression has been demonstrated in the dissected spe- cimens of chronic pancreatitis and pancreatic adeno- carcinoma (24). In pancreatic cancer cells, suppression of NFκB could downregulate the mRNA expression of Shh (24). Further activation of NFκB by inflamma- tory stimuli (IL-1β, TNF-α, and lipopolysaccharide) can induce the upregulation of Shh, further resulting in activation of the Hh pathway. Notably, overexpression of Shh induced by these stimuli can also be suppressed by blockade of NFκB. Therefore, NFκB-mediated Shh expression results in the activation of the Hh pathway and enhanced cell proliferation in cancer cells (24).
Sox2 is the key transcription factor essential to plu- ripotency and is demonstrated its crucial role in many types of cancers. Metastasis and low survival rate in HCC have been correlated with expression of Sox2.
Therefore, Sox2 plays an important role in HCC pro- gnosis and treatment (5). In addition, co-expression of Sox2 and Oct4 in HCC has been suggested the aggres- sive tumor characteristics and a worse clinical outcome (6). Notably, the in situ expression of Sox2 and Oct4 mRNA were both found in hepatoma cell lines and tu- mor tissues. Therefore, the expression of Sox2 and Oct4 can be utilized as novel predictors of poor prognosis for patients undergoing resection of HCC (4). Knockdown of Sox2 and Oct4 gene expression in HCC cells can reduce the sphere formation and increase cellular sen- sitivity to chemotherapy, indicating that Sox2 and/or Oct4 may be a promising target in clinical cancer treat- ment (16). To our knowledge, we are the first to show the effects of PA on CSC-like properties (spherogenesis and stemness marker expression) of hepatoma cells, we also demonstrate that NFκB not only played an impor- tant role in PA-mediated pro-inflammatory (17) and profibrogenic effects (9), but also regulated the CSC- like properties in HepG2 cells. Using choline-deficient diet to induce the accumulation of lipid droplets in rat liver, we also observed both pro-inflammatory and pro- fibrogenic responses by lipid accumulation. Notably, NFκB activation and GSTP expression, which is a pre- neoplastic marker, were both elevated in the liver (data not shown), suggesting the in vivo effects of lipid accu- mulation on the progression of liver cancer.
It has been shown that the epithelial ovarian CSCs also have NFκB-mediated stemness properties and re- present the chemo-resistant population (28). Our results are in agreement with their findings and suggest a pos- sible application of NFκB inhibitors in the treatment of NASH-mediated carcinogenesis (29). As a proof of principle, curcumin has recently been shown to induce the cell death, down-regulation of CSC markers and
suppressed tumorigenicity of CSCs in HCC cell lines
through NFκB inhibition (30). In summary, our results
suggested that lipid accumulation might not only induce ROS production and pro-inflammatory responses in he- patocytes (12) but also activated CSC-like properties of hepatoma cells. This study suggests the possibility that saturated fatty acid might activate CSC-like properties of hepatoma cells in NAFLD patients. Thus, this study provides an attractive model to study the mechanisms of carcinogenesis in NAFLD and to identify and test novel therapies for chemopreventive therapies in NAFLD.
Acknowledgements
This study was supported by grants from the Ministry of Science and Technology of Taiwan (MOST103- 2314- B-715-001-MY2, MOST104-2314-B-715-003 -MY3,
104-2320-B-039-031), Mackay Medical College (MMC RD1010061, RD1020038, RD1020047, RD1030053, RD1030076, RD1040109), and Mackay Memorial Hos- pital (MMH-MM-10304, MMH-MM-10405), and the grant from China Medical University (CMU104-S-02), and the grant from Shin Kong Wu Ho-Su Memorial Hospital, Taipei, Taiwan.
Other articles in this theme issue include references (31-
42).
References
1. Angulo, P., Lindor, K.D. Treatment of non-alcoholic steatohe- patitis. Best Pract Res Clin Gastroenterol. 2002, 16: 797-810. doi:
10.1053/bega.2002.0327
2. Malaguarnera, M., Di Rosa, M., Nicoletti, F., Malaguarnera, L.
Molecular mechanisms involved in NAFLD progression. J Mol Med. 2009, 87: 679-695. doi: 10.1007/s00109-009-0464-1
3. Leamy, A.K., Egnatchik, R.A., Young, J.D. Molecular mecha- nisms and the role of saturated fatty acids in the progression of non- alcoholic fatty liver disease. Prog Lipid Res. 2013, 52: 165-174. doi:
10.1016/j.plipres.2012.10.004
4. Huang, P., Qiu, J., Li, B., Hong, J., Lu, C., Wang, L., et al. Role of Sox2 and Oct4 in predicting survival of hepatocellular carcinoma patients after hepatectomy. Clin Biochem. 2011, 44: 582-589. doi:
10.1016/j.clinbiochem.2011.02.012
5. Sun, C., Sun, L., Li, Y., Kang, X., Zhang, S., Liu, Y. Sox2 ex- pression predicts poor survival of hepatocellular carcinoma patients and it promotes liver cancer cell invasion by activating Slug. Med Oncol. 2013, 30: 503. doi: 10.1007/s12032-013-0503-1
6. Yin, X., Li, Y.W., Zhang, B.H., Ren, Z.G., Qiu, S.J., et al.
Coex- pression of stemness factors Oct4 and Nanog predict liver resection. Ann Surg Oncol. 2012, 19: 2877-2887. doi:
10.1245/s10434-012- 2314-6
7. Cao, L., Zhou, Y., Zhai, B., Liao, J., Xu, W., Zhang, R., et al.
Sphere-forming cell subpopulations with cancer stem cell properties in human hepatoma cell lines. BMC Gastroenterol. 2011, 11: 71-82.
doi: 10.1186/1471-230X-11-71
8. Parkes, J.G., Templeton, D.M. Effects of retinol and hepatocyte- conditioned medium on cultured rat hepatic stellate cells. Ann Clin Lab Sci. 2003, 33: 295-305.
9. Wobser, H., Dorn, C., Weiss, T.S., Amann, T., Bollheimer, C., Buttner, R., et al. Lipid accumulation in hepatocytes induces fibroge- nic activation of hepatic stellate cells. Cell Res. 2009, 19:
996-1005. doi: 10.1038/cr.2009.73
10. Chong, L.W., Hsu, Y.C., Chiu, Y.T., Yang, K.C., Huang, Y.T. Anti-fibrotic effects of thalidomide on hepatic stellate cells
and dimethylnitrosamine-intoxicated rats. J Biomed Sci. 2006, 13:
403-
418. doi: 10.1007/s11373-006-9079-5
11. Chong, L.W., Hsu, Y.C., Chiu, Y.T., Yang, K.C., Huang, Y.T.
An- tifibrotic effects of triptolide on hepatic stellate cells and dimethyl- nitrosamine-intoxicated rats. Phytother Res. 2011, 25:
990-999. doi:
10.1002/ptr.3381
12. Chong, L.W., Hsu, Y.C., Lee, T.F., Lin, Y., Chiu, Y.T., Yang, K.C., et al. Fluvastatin attenuates hepatic steatosis-induced fibroge- nesis in rats through inhibiting paracrine effect of hepatocyte on hepatic stellate cells. BMC Gastroenterol. 2015 15: 22-34. doi:
10.1186/s12876-015-0248-8
13. Chomczynski, P., Sacchi, N. The single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform ex- traction: twenty-something years on. Nat Protoc. 2006, 1: 581-585.
doi: dx.doi.org/10.1038/nprot.2006.83
14. Rocco, M., Antonelli, M., Letizia, V., Alampi, D., Spadetta, G., Passariello, M., et al. Lipid peroxidation, circulating cytokine and endothelin 1 levels in healthy volunteers undergoing hyperbaric oxygenation. Minerva Anestesiol. 2001, 67: 393-400.
15. Tobar, N., Villar, V., Santibanez, J.F. ROS-NFkappaB mediates TGF-beta1-induced expression of urokinase-type plasminogen acti- vator, matrix metalloproteinase-9 and cell invasion. Mol Cell Bio- chem. 2010, 340: 195-202. doi: 10.1007/s11010-010-0418-5.
16. Joshi-Barve, S., Barve, S.S., Amancherla, K., Gobejishvili, L., Hill, D., Cave, M., et al. Palmitic acid induces production of proin- flammatory cytokine interleukin-8 from hepatocytes. Hepatology 2007, 46: 823-830. doi: 10.1002/hep.21752
17. Kim, C.S., Ross, I.A. Regulatory role of free fatty acids (FFAs)- Palmitoylation and Myristoylation. Food Nutri Sci., 2013, 4; 202- 211. doi: 10.4236/fns.2013.49A1028
18. Galluzzo, P., Caiazza, F., Moreno, S., Marino, M. Role of ERbe- ta palmitoylation in the inhibition of human colon cancer cell proli- feration. Endocr Relat Cancer 2007, 14:153-167.
19. Fiorentino, M., Zadra, G., Palescandolo, E., Fedele, G., Bailey, D., Fiore, C., et al. Overexpression of fatty acid synthase is asso- ciated with palmitoylation of Wnt1 and cytoplasmic stabilization of beta-catenin in prostate cancer. Lab Invest 2008, 88:1340-1348. doi:
10.1038/labinvest.2008.97
20. Coleman, D.T., Soung, Y.H., Surh, Y.J., Cardelli, J.A., Chung, J. Curcumin Prevents Palmitoylation of Integrin beta4 in Breast Can- cer Cells. PLoS One 2015, 10: e0125399.
21. Takahashi, N., Iwahori, A., Breitman, T.R., Fukui, T. Tunica- mycin in combination with retinoic acid synergistically inhibits cell growth while decreasing palmitoylation and enhancing retinoylation of proteins in the human breast cancer cell line MCF-7. Oncol Res 1997, 9: 527-533.
22. Gao, X., Arenas-Ramirez, N., Scales, S.J., Hannoush, R.N.
Membrane targeting of palmitoylated Wnt and Hedgehog revea- led by chemical probes. FEBS Lett 2011, 585: 2501-2506. doi:
10.1016/j.febslet.2011.06.033
23. Sicklick, J.K., Li, Y.X., Jayaraman, A., Kannangai, R., Qi, Y., Vivekanandan, P., Ludlow, J.W., et al. Dysregulation of the Hedge- hog pathway in human hepatocarcinogenesis. Carcinogenesis 2006, 27: 748-757. doi: 10.1093/carcin/bgi292
24. Nakashima, H., Nakamura, M., Yamaguchi, H., Yamanaka, N., Akiyoshi, T., Koga, .K, Yet al. Nuclear factor-kappaB contributes to hedgehog signaling pathway activation through sonic hedgehog induction in pancreatic cancer. Cancer Res 2006, 66: 7041-7049.
25. O’Toole, S.A., Machalek, D.A., Shearer, R.F., Millar, E.K., Nair, R., Schofield, P., et al. Hedgehog overexpression is associated with stromal interactions and predicts for poor outcome in breast cancer. Cancer Res 2011, 71: 4002-4014. doi:
10.1158/0008-5472.CAN-10- 3738
26. Komekado, H., Yamamoto, H., Chiba, T., Kikuchi, A. Glycosy- lation and palmitoylation of Wnt-3a are coupled to produce an active
form of Wnt-3a. Genes Cells 2007, 12: 521-534. doi:
10.1111/j.1365- 2443.2007.01068.x
27. Song, L., Li, Z.Y., Liu, W.P., Zhao, M.R. Crosstalk between Wnt/beta-catenin and Hedgehog/Gli signaling pathways in colon cancer and implications for therapy. Cancer Biol Ther 2015, 16: 1-7.
doi: 10.4161/15384047.2014.972215.
28. Chefetz, I., Holmberg, J.C., Alvero, A.B., Visintin, I., Mor, G.
Inhibition of Aurora-A kinase induces cell cycle arrest in epithelial ovarian cancer stem cells by affecting NFkB pathway. Cell cycle 2011, 10: 2206-2214.
29. Seki, E., Brenner, D.A. The role of NF-kappaB in hepatocarci- nogenesis: promoter or suppressor? J Hepatol. 2007, 47: 307-309.
30. Marquardt, J.U., Gomez-Quiroz, L., Arreguin Camacho, L.O., Pinna, F., Lee, Y.H., Kitade, M., et al. Curcumin effectively inhi- bits oncogenic NF-kappaB signaling and restrains stemness fea- tures in liver cancer. J Hepatol. 2015, 63: 661-669. doi: 10.1016/j.
jhep.2015.04.018
31. Li, Y., Ahmad, A., Sarkar and F. H., ASPP and iASPP: Implica- tion in cancer development and progression. Cell. Mol. Biol. 2015, 61(6): 2-8.
32. Masood, N.,, Qureshi, M. Z. and Yasmin, A., Association of NOTCH with different microRNAs in head and neck cancer. Cell.
Mol. Biol. 2015, 61(6): 9-16.
33. Amirkhah, R., Farazmand, A., Irfan-Maqsood, M., Wolkenhau- erand, O. and Schmitz, U., The role of microRNAs in the resistance to colorectal cancer treatments. Cell. Mol. Biol. 2015, 61(6): 17-23.
34. Wang, Z., Chen, J. and Capobianco, A. J., The Notch signal- ing pathway in esophageal adenocarcinoma. Cell. Mol. Biol. 2015, 61(6): 24-32.
35. Limami, Y., Pinon, A., Riaz, A. and Simon, A., TRAIL and tar- geting cancer cells: between promises and obstacles. Cell. Mol. Biol.
2015, 61(6): 33-38.
36. Silva Galbiatti-Dias, A. L., Pavarino, É. C., Kawasaki-Oyama, R. S., Maniglia, J. V., Maniglia, E. J. V. and Goloni Bertollo, E. M., Cancer stem cells in head and neck cancer: A Mini Review. Cell.
Mol. Biol. 2015, 61(6): 39-43.
37. Musella, A., Marchetti, C., Gasparri, M. L., Salerno, L., Caso- relli, A., Domenici, L., Imperiale, L., Ruscito, I., Abdul Halim, T., Palaia, I., Di Donato, V., Pecorini, F., Monti, M., Muzii, L. and Pani- ci, P. B., PARP inhibition: A promising therapeutic target in ovarian cancer. Cell. Mol. Biol. 2015, 61(6): 44-61.
38. Attar, R., Tabassum, S., Fayyaz, S., Ahmad, M. S., Nogueira, D. R., Yaylim, I., Timirci-Kahraman, O., Kucukhuseyin, O., Cacina, C., Farooqi, A. A. and Ismail, M., Natural products are the future of anticancer therapy: Preclinical and clinical advancements of Viscum album phytometabolites. Cell. Mol. Biol. 2015, 61(6): 62-68.
39. Hsu, Y-C., Hsieh, Y-H., Liao, C-C., Chong, L-W., Lee, C-Y., Yu, Y-L. and Chou, R-H., Targeting post-translational modifications of histones for cancer therapy. Cell. Mol. Biol. 2015, 61(6): 69-84.
40. Smina, T. P., Mohan, A., Ayyappa, K. A., Sethuraman, S. and Krishnan, U. M., Hesperetin exerts apoptotic effect on A431 skin carcinoma cells by regulating mitogen activated protein kinases and cyclins. Cell. Mol. Biol. 2015, 61(6): 92-99.
41. Ahmadi, M., Jafari, R., Marashi, S. A. and Farazmand, A., In- direct role of microRNAs and transcription factors in the regulation of important cancer genes: A network biology approach. Cell. Mol.
Biol. 2015, 61(6): 100-107.
42. Zahoor, A., Mansoor, Q., Farooqi, A. A., Fayyaz, S., Naz, G.
and Ismail, M., Genetic variants in the tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) and death receptor (DR4) genes contribute to susceptibility to colorectal cancer in pakistani popula- tion. Cell. Mol. Biol. 2015, 61(6): 108-112.