行政院國家科學委員會專題研究計畫 期中進度報告
新生鼠給予抗憂鬱劑-百憂解可保護新生兒期窒息腦傷及拮
抗青少年期之古柯鹼成癮(2/3)
計畫類別: 個別型計畫 計畫編號: NSC93-2314-B-006-014- 執行期間: 93 年 08 月 01 日至 94 年 07 月 31 日 執行單位: 國立成功大學醫學系小兒科 計畫主持人: 黃朝慶 報告類型: 精簡報告 處理方式: 本計畫可公開查詢中 華 民 國 94 年 6 月 1 日
行政院國家科學委員會補助專題研究計畫
□期中進度報告
新生鼠給予抗憂鬱劑-百憂解可保護新生兒其窒息腦傷及拮抗青少年期之
古柯鹼成癮(2/3)
計畫類別:
□
個別型計畫 □ 整合型計畫
計畫編號:NSC93-2314-B-006 -014
執行期間:
93 年 08 月 01 日至 94 年 07 月 31 日
計畫主持人:黃朝慶
共同主持人:郭余民
計畫參與人員:
成果報告類型(依經費核定清單規定繳交):□精簡報告 □完整報告
本成果報告包括以下應繳交之附件:
□赴國外出差或研習心得報告一份
□赴大陸地區出差或研習心得報告一份
□出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份
處理方式:除產學合作研究計畫、提升產業技術及人才培育研究計畫、列
管計畫及下列情形者外,得立即公開查詢
□涉及專利或其他智慧財產權,□一年□二年後可公開查詢
執行單位:成功大學醫學院小兒科
中 華 民 國 94 年 05 月 25 日
關鍵詞:百憂解、窒息腦傷、水迷宮
外界環境或藥物對發展中的生物可透過其生理狀態之可塑性的改變,進而改變此生物對 環境之適應性。發展中的腦部之可塑性特別敏感,尤其是週產時期更易受到壓力、內毒素、 生長激素或轉錄因子的影響(programming)。cAMP response element-binding protein (CREB) 為一重要之細胞轉錄因子,在神經系統之發展及其可塑性扮演一重要角色,且在學習及記憶 形成,成癮藥上癮過程及對抗窒息缺氧之細胞存活機制均極為重要。細胞受到刺激時,很容 易活化此因子,在Ser133 之位置受磷酸化後,進而促進神經信息傳遞,細胞突觸可塑性, 細胞內信息傳遞等基因的表現。 抗憂鬱劑之使用可促進成鼠腦部海馬回及大腦皮質CREB 之活化,特別是血清素 (serotonin)回收抑制劑-百憂解,特具此功能,也有研究顯示百憂解透過 CREB 之活化來 促進brain-derived nerve growth factor (BDNF)之作用,而百憂解之抗憂鬱作用一般認為與此 機制有關。 新生鼠使用百憂解是否對其腦部之神經可塑性有長期作用,至今未明。我們在新生兒期 使用低劑量百憂解前處理可在行為上看到其對抗新生鼠缺氧缺血腦傷之神經保護作用,但在 高劑量之下卻有惡化神經功能之表現,我們進一步分析發現低劑量之百憂解可促進pCREB 之表現以及增加再生神經細胞之存活能力;這些發現均意味著活化CREB 來促進神經可塑性 及再生能力,可為改善新生兒腦傷之重要機制。
Abstract
Key words: fluoxetine, neural plasticity, hypoxic-ischemia, water maze
The brain is sensitive to perinatal programming by agents known to permanently affect brain development. Neuroplasticity after perinatal programming may allow for neuroprotection against hypoxic-ischemia (HI) at birth. The cAMP response element-binding protein (CREB) is a key mediator of stimulus-induced nuclear responses that underlie cell survival, learning and memory, and plasticity of the nervous system. CREB activation in the cortex and hippocampus are
upregulated by chronic fluoxetine, a selective serotonin reuptake inhibitor, treatment. We therefore examined whether fluoxetine administration before HI may protect against neonatal HI brain injury through CREB-mediated mechanisms. We found that fluoxetine pretreatment in rat pups
significantly decreased the mortality rate during HI. Fluoxetine also reduced functional deficits in adult rats with neonatal HI brain injury in a dose-related manner. The neuroprotective mechanisms were associated with increased CREB phosphorylation. Neurogenesis also increased because of greater precursor cell survival in the hippocampal dentate gyrus. These findings suggest that functional deficits after HI in the developing brain can be reduced by agents that enhance neural plasticity and neurogenesis through CREB activation, which may implicate a new strategy for influencing the cognitive outcome of neonatal HI injury.
Introduction
Hypoxic-ischemia (HI) encephalopathy is a major cause of neonatal mortality and neurological disabilities in childhood.1-3 Our understanding of the pathogenesis of neonatal HI brain injury has increased considerably over the last decades1,3; there is, however, no effective clinical treatment. Perinatal plasticity of physiological systems may allow environmental factors or drugs to alter the “set-point” or to “hard-wire” the developmental plasticity of the physiological system by which organisms can adapt their physiological responses to better meet environmental demands.4 The brain is a key target for such effects and is sensitive to perinatal programming by agents known to permanently affect brain development, such as stress, enrichment or handling, and steroids.4,5 Neuroplasticity after perinatal programming may allow for neuroprotection against HI at birth.
The cAMP response element-binding protein (CREB), a transcription factor, is a key mediator of stimulus-induced nuclear responses that underlie cell survival, regeneration, learning and
memory, and the plasticity of the nervous system.6 CREB is phosphorylated on Ser133 (pCREB) and consequently binds to the cAMP response element (CRE) of target genes, which controls CREB-dependent gene expressions, including the survival and plasticity growth factor, brain-derived neurotrophic factor (BDNF).6,7 Our previous study showed that carotid-artery ligation preconditioning against HI in the neonatal brain is associated with robust and sustained phosphorylation of CREB.8 Furthermore, pharmacological activation of the cAMP-CREB signaling pathway protected the neonatal rat brain through a sustained increase in pCREB expression.
Molecular studies have identified the important role of CREB in the mechanism of action of chronically administered antidepressants against depression.9,10 CREB activation in the cortex and hippocampus of both rodents and humans is upregulated by chronic treatment with fluoxetine, a selective serotonin reuptake inhibitor (SSRI).9-11 Temporal and regional upregulation of BDNF mRNA and its receptor, trkB, parallels CREB activation after chronic antidepressant
raises the possibility that chronic fluoxetine treatment may protect against HI injury in the neonatal brain through CREB-mediated mechanisms.
Structural plasticity, including increased neurogenesis, dendritic arborization, and
synaptogenesis, provides the cellular mechanism underlying the long-term effect of antidepressants in the adult brain.14-16 Furthermore, chronic antidepressant treatment influences the proliferation as well as the maturation, differentiation, and survival of newborn neurons in adult rats.16,17 Recently, the newborn neurons in the rodent brain after stroke injury or learning were shown to be actively integrated into the existing circuitry and to contribute to ameliorating neurological deficits or forming hippocampal-dependent memory.18,19 Whether a chronic regimen of antidepressants can protect against HI damage in rat pups by enhancing neurogenesis and synaptogenesis has not been examined.
Despite the strong evidence for the neuroplasticity benefits of the SSRIs in adult rats, the neuroprotective effects of fluoxetine against neonatal HI brain injury have not been investigated. Fluoxetine facilitates motor recovery in stroke patients undergoing rehabilitation,20,21 and improves cognition after traumatic brain injury.22 Furthermore, it is the only SSRI approved for pediatric use. Therefore, we used an established rat pup model of HI brain injury to determine the effects and underlying mechanisms of early-life exposure to fluoxetine on neural plasticity after HI brain injury in neonatal rats. In this study, we examined the following hypotheses: 1) fluoxetine
pretreatment is neuroprotective against HI damage in the neonatal brain, and 2) the neuroprotective mechanism of fluoxetine pretreatment involves a) increased phosphorylation of CREB, b)
Materials and Methods
This study was approved by our university's Animal Care Committee. Ten to twelve
Sprague-Dawley pups per dam were used and housed with a 12/12-hour light/dark schedule in a temperature- and humidity-controlled colony room. The pups were housed with their dams until weaning on postnatal (P) day 21, and then housed in groups of 4-5 per cage.
Experimental Groups
Hypoxic-ischemia. From P1 to P7, rat pups of either sex received daily intraperitoneal injection of fluoxetine (Sigma, St. Louis, MO) or normal saline. Animals received a 100-µl solution containing either saline (Ns-HI group) or fluoxetine: 5 mg/kg (5-Flu-HI group) or 15 mg/kg (15-Flu-HI group) dissolved in saline. On P7, three hours after the last injection, rats pups were subjected to HI (right common carotid artery ligation followed by 8% oxygen hypoxia for 2.5 hours), as described
previously.8 Another group of rat pups that received a daily normal saline (Ns group) injection from P1 to P7 without further exposure to HI served as controls.
On P7, the Ns-HI-, 5-Flu-HI-, and 15-Flu-HI-group animals were anesthetized with 2.5% halothane (balance, room air), and the right common carotid artery was surgically exposed and permanently ligated with 5-0 surgical silk. After surgery, the pups were returned to the dam for a 1-hour recuperation period before HI. The animals were then placed in airtight 500-ml containers partially submerged in a 37°C water bath through which humidified 8% oxygen (balance, nitrogen) was maintained at a flow rate of 3 L/min for 2.5 hours. After completion of HI, the rat pups were returned to their cages. Behavioral assessment by Morris water maze (MWM) was performed from P35 to P40, and pathological examination by hemispheric weight reduction determined at P42. Outcome Measure- Behavioral Assessment
Morris water maze (MWM). Rats undertook the MWM as described previously, but with some modifications.8,23,24 In brief, the pool was illuminated by room lights, four points on the perimeter of the pool were designated, and the pool was divided into four quadrants. An 8 cm × 8 cm
one of the quadrants, 1 cm below the water surface. Escape distance, escape latency, swimming speed, and swimming patterns of the rats were monitored by a camera mounted above the pool and connected to a computer program (EthoVision, Wageningen, The Netherlands). The MWM task was divided into five phases. Training and testing was conducted between 9:00 and 17:00. Phase I: Acquisition phase (Days 1-2). Phase I consisted of 2 days of training with 2 sessions per day. Each session consisted of 4 trials with 4 different starting positions. After swimming from each of the starting positions and mounting the submerged platform, the animals were allowed to remain there for 60 seconds. If an animal did not find the platform within 120 seconds, it was placed directly on the platform and allowed a 60-second rest period.
Phase II: Probe test 1 (Day 3). On day 3, the platform was removed from the pool, and the rats were placed in the pool in the quadrant opposite the previous platform position. The rats were allowed 60 seconds of free swimming. Swimming time in each quadrant was recorded. The
percentage of time spent in the target quadrant where the platform had been previously located was calculated.
Phase III: Reversal phase (Days 4-5). Phase III was performed by placing the platform at a new position in the quadrant opposite the one used in Phase I. This reversal training was performed using the same procedure as described in Phase I.
Phase IV: Probe test 2 (Day 6). This new probe test was performed using the same procedure as described in Phase II. The percentage of time spent in the new target quadrant was calculated for each animal.
Phase V: Cued version (Visible platform test) (Day 6). Two hours after the probe test on day 6, the visible platform test was performed to determine whether the experimental manipulation resulted in a crude change in visual motor performance. The animals were released from a specific location and required to locate a green-colored escape platform 2 cm above the water surface in a position not used during the former sessions. If the animal did not reach the platform within 120 seconds, it was recorded as 120 seconds.
Outcome Measure-Pathological Assessment
Hemispheric weight reduction at P42. Our previous study showed that, in P7-rats with cerebral HI, changes in hemispheric weight reduction were highly correlated not only with changes in the brain areas, and but also with hemispheric volume changes measured on P36.8 Therefore, only the hemispheric weight data are shown in the present study. After they had been anesthetized with pentobarbital (50 mg/kg, i.p.), rats were decapitated and the brain was removed. The brainstem and cerebellum were removed, the forebrain was sectioned at the midline, and left and right hemispheric weights were determined. The percentage of hemispheric weight reduction, measured as [(left hemisphere weight − right hemisphere weight)/left hemisphere weight], was used as the measure of cerebral injury.
Immunohistochemical Stain for Neurogenesis
Bromodeoxyuridine (BrdU) labeling. BrdU is a thymidine analog incorporated into the DNA during the synthesis (S) phase of the cell cycle. Because BrdU is available for 30 minutes, only a
proportion of dividing cells in the S phase of the cell cycle will be labeled by a single injection.26 To investigate the effects of fluoxetine treatment specifically on neurogenesis, male rat pups with daily administration of saline or 5-mg/kg fluoxetine from P1 to P7 were given a single intraperitoneal injection of BrdU (100 mg/kg; Sigma, St. Louis, MO) 30 minutes after the last dose of saline or fluoxetine on P7. For the proliferation study, the animals (Ns and 5-Flu groups) were killed 3 hours after BrdU injection to study the number of newly-formed cells in the brain. To determine the survival and cell phenotypes of the newly born cells after HI injury, Ns-HI- and 5-Flu-HI-group rats were not killed until 14 days after the BrdU injection and HI. After perfusion, brains were
post-fixed overnight in paraformaldehyde at 4°C and stored at 4°C in 30% sucrose. Serial sections of the brains (30 µm thick) were cut on a freezing microtome.
BrdU immunostaining. BrdU staining was described previously.27 In brief, the sections were heated (37°C for 1 hour), incubated in 50% formaldehyde (65°C for 1 hour), 2 M HCl (37°C for 30
minutes, and blocked in 3% normal rabbit serum/0.3% Triton X-100 in PBS (room temperature for 1 hour), followed by incubation with goat anti-BrdU (1:200; Accurate Chemical & Scientific Corporation, Westbury, NY) at 4°C overnight. Subsequently, the sections were incubated with anti-goat secondary antibody (1:200; Santa Cruz Biotechnology, Inc, Santa Cruz, CA) and developed with an ABC kit (Vectastain; Vector Laboratories, Burlingame, CA). For fluorescence imaging, the sections were incubated with anti-goat-FITC secondary antibody (1:200; Santa Cruz). The sections were then incubated with rabbit anti-glial fibrillary acidic protein (GFAP, 1:1000; Sigma) or mouse anti-neuron-specific nuclear protein (NeuN, 1:500; Chemicon International, Inc., Temecula, CA). The sections were then incubated with goat anti-rabbit-rhodamine (1:200;
Chemicon) or goat anti-mouse-rhodamine (1:200; Chemicon), respectively. Sections were rinsed, dried, and coverslipped using fluorescence mounting medium (DAKO Corporation, Carpinteria, CA). Control sections were processed with the primary antisera omitted. Labeling was imaged with a fluorescent microscope (Eclipse E800; Nikon Instech Co., Ltd., Kawasaki, Japan).
Quantification. Stereological analysis of BrdU(+) cells in every sixth section in a series of 30-µm coronal sections of dorsal hippocampus was performed by a blinded examiner. Labeled cells were counted in the area encompassing the entire granule cell layer (GCL) and extending approximately one to two cell layers deep into the hilus and the hilus area. For cellular counting, images were magnified 100× and captured digitally using an image analysis system (Image-Pro Plus; Media Cybernetics, Inc., Silver Spring, MD). Area measurements of both the dentate GCL and hilus were made from each slide used for the cell counts. The density of BrdU(+) cells in each section was calculated by dividing the numbers of BrdU(+) nuclei by the area. Density for the five to seven sections was averaged to obtain a mean density value for each animal. The volume of the
hippocampal dentate gyrus was determined by summing the traced areas for each section multiplied by the distance between sample sections. We also examined the BrdU(+) cells in the subventricular zone (SVZ) present on every 10th coronal section. To determine phenotype, we counted 100 BrdU(+) cells in the hippocampal GCL per animal to obtain the percentage of BrdU(+) cells
co-localized with NeuN. Western blot analysis
Tissue was homogenized in cold lysis buffers as described previously.8,23 Fifty-microgram samples were resolved in 10% SDS-PAGE gels and blotted electrophoretically to nitrocellulose membranes. Membranes were incubated with primary antibodies, and immunoreactivity was detected using horseradish-conjugated secondary antibody or mouse immunoglobulin G antibody (1:5000; Chemicon) and visualized with enhanced chemiluminescence. The following primary antibodies were used: anti-CREB (1:1000; Upstate Biotechnology, Charlottesville, VA), anti-pCREB (1:1000; Upstate Biotechnology), and β-actin (1:1000; Chemicon).
Statistics
Statistical significance (p < 0.05) was determined using one-way ANOVA. When data were not normally distributed, analyses were performed using a non-parametric ANOVA by ranks
(Kruskal-Wallis one way test) followed by Dunn's multiple comparison tests. Repeated measures ANOVA was employed where days or quadrants were treated as a within-subject factor and groups as a between-subject factor. Continuous data are expressed as mean ± standard error of the mean (SEM) unless indicated otherwise.
Results
Low- But Not High-Dose Fluoxetine Pretreatment Protects Against Hypoxic Brain Damage at the Behavioral Level
First, we examined whether a daily 5-mg/kg injection of fluoxetine was neuroprotective against HI injury. During the acquisition phase of the MWM, the Ns, 5-Flu-HI, and Ns-HI groups all learned to find the submerged platform (p < 0.001). However, the total distance covered to reach the submerged platform was significantly different between the three groups (p < 0.001), and there was no interaction between the training sessions and treatments. Post hoc multiple comparisons revealed that the Ns-HI-group rats traveled significantly longer distances when finding the platform than did the Ns- (p < 0.001) or the 5-Flu-HI-group (p < 0.05) rats. There was also a significant difference between the 5-Flu-HI and the Ns groups (p < 0.001) (Fig 1A). During the reversal learning phase, the 5-Flu-HI group improved compared with the Ns-HI group. Multiple comparisons revealed that both HI groups traveled significantly longer distances than did the
Ns-group (both p < 0.001). The 5-Flu-HI-group also traveled a significantly shorter distance finding the platform than the Ns-HI group (p < 0.005) did (Fig 1A).
During probe test 1, Ns-group rats spent more time in the target quadrant than did the two HI-group rats (p < 0.001). There was no significant difference between the 5-Flu-HI and Ns-HI groups. However, in probe test 2, the 5-Flu-HI group showed much improvement and spent significantly more time searching in the new target quadrant than did the Ns-HI group (p < 0.05). There was no significant difference between the 5-Flu-HI and the Ns group (Fig 1B).
In the cued version, there was a significant difference (p < 0.001) between the three groups in time spent reaching the visible platform. The 5-Flu-HI group spent significantly (p < 0.001) less time than the Ns-HI group finding the visible platform, and the 5-Flu-HI group spent significantly (p < 0.01) more time than the Ns group. The Ns group spent significantly less time than the Ns-HI (p < 0.001) or the 5-Flu-HI group (p < 0.01) (Fig 1C). In contrast, there was no significant
1D).
We next examined whether a higher dose of fluoxetine pretreatment (15-mg/kg daily) was also neuroprotective. In contrast to the 5-Flu-HI group, the 15-Flu-HI group performed even worse than the Ns-HI group in all phases of training (Fig 2A). In both the acquisition and reversal training, the 15-Flu-HI-group rats had to be categorized as complete non-learners because their travel distance did not decrease over time. Multiple comparisons revealed that the 15-Flu-HI group traveled a significantly longer distance searching for the submerged platform during the acquisition and reversal phases than did the Ns (both p < 0.001) or Ns-HI (both p < 0.005) group. There was no significant difference between the 15-Flu-HI and Ns-HI groups in either probe test (Fig 2B). In the cued version, there was also no significant difference between the 15-Flu-HI and Ns-HI groups in time spent reaching the visible platform (Fig 2C). Again, there was no significant difference between the three groups in swimming speed(Fig 2D). Therefore, we found that a low dose of fluoxetine protected rat pups from HI brain damage at functional levels, but that a high dose exaggerated the damage.
Fluoxetine Pretreatment Did Not Protect Against Hypoxic Brain Damage at the Pathological Level The degree of brain injury, as measured by cerebral hemisphere weight reduction, showed no significant difference between the 5-Flu-HI, 15-Flu-HI, and Ns-HI groups (Fig 3A).
The onset of abnormal behaviors was seen immediately after fluoxetine injection in 70% of the 15-mg/kg-group rats. The rat pups became agitated and showed hyperactivity, head shaking, clonic twitching of hind-limbs, wide-running, myoclonic jerk, ataxic walking, circling, or falling, which gradually subsided within 30 minutes after injection. None of these adverse behaviors were seen in the Ns- or 5-Flu-group rats. The mortality rate during HI was significantly decreased with an increasing dose of pretreatment fluoxetine (Ns-HI = 13.4%, 5-Flu-HI = 6.1%, 15-Flu-HI = 0%) (chi-square = 4.17, p = 0.04). There were no significant differences in body weight from P1 to P7 or from P14 to P28 between the Ns-HI, 5-Flu-HI, 15-Flu-HI, and Ns groups (Fig 3B). There were also no significant differences in the degree of rectal temperature measured before and after HI between
the three HI groups.
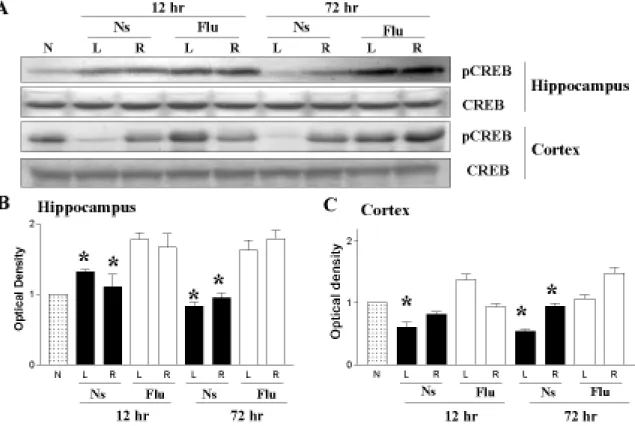
Low-Dose Fluoxetine Pretreatment Increased Posthypoxia CREB Phosphorylation in the Hippocampus and Cortex
Chronic fluoxetine treatment in adult rats increases CREB phosphorylation, which induces cell survival signals and promotes synaptic plasticity.6,9 We therefore investigated, using Western
blotting, the levels of CREB phosphorylation after HI in rat pups with or without fluoxetine pretreatment. The 5-Flu-HI group had significantly higher levels of pCREB expression in the ipsilateral hippocampus 12 hours and 72 hours after HI than did the Ns-HI group at the respective time points (both p < 0.05). The 5-Flu-HI group also had significantly higher pCREB levels in the ipsilateral cortex 72 hours after HI than did the Ns-HI group (p < 0.05) (Fig 4A-C). Compared with the Ns-HI group, the 5-Flu-HI group also had significantly higher pCREB levels in the contralateral hippocampus and cortex 12 hours and 72 hours after HI (both p < 0.05). Total CREB was
unchanged in the hippocampus and cortex between the two HI groups.
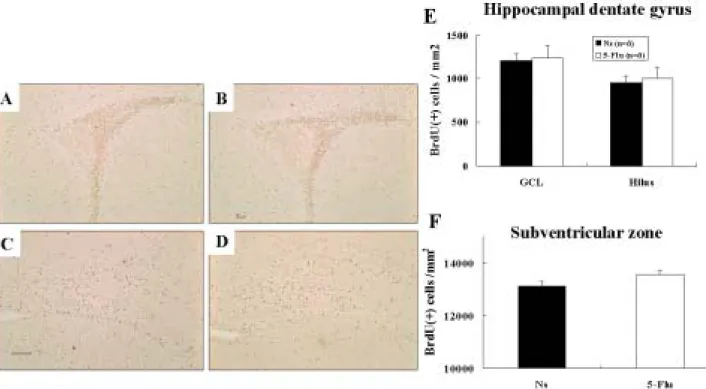
Low-Dose Fluoxetine Pretreatment Increased Posthypoxia Neurogenesis in the Hippocampal Dentate Gyrus
We examined whether fluoxetine pretreatment increased neurogenesis after HI in the immature brain. Increased precursor cell proliferation, survival, or differentiation upregulates neurogenesis. We first assessed progenitor cell proliferation by investigating BrdU labeling 3 hours after the last fluoxetine or normal saline administration on P7. Unbiased stereological cell counts revealed no significant differences in the numbers of BrdU(+) cells in the SVZ between the 5-Flu and Ns groups (Fig 5A, B, F). BrdU(+) cell density in the hippocampal GCL and hilus of dentate gyrus in the 5-Flu group also did not differ from that of the Ns group (Fig 5C, D, E). These data indicated that the 7 days of low-dose fluoxetine treatment did not alter progenitor cell proliferation in the immature rat brain.
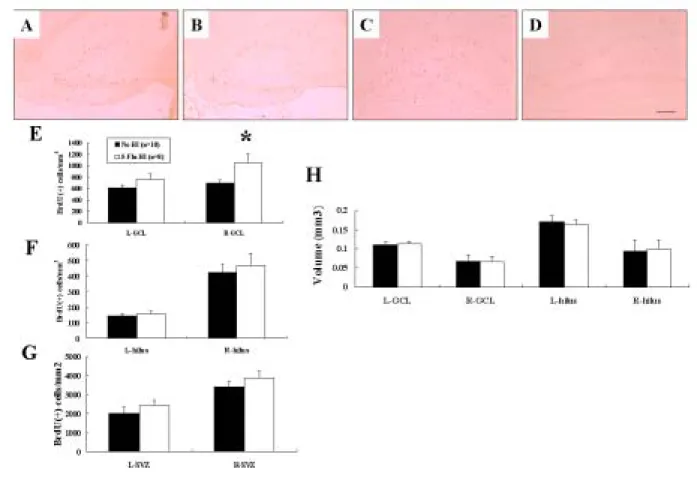
To address the survival of newly born cells after HI, a second group of rats was killed 14 days after BrdU injection and HI. The density of BrdU(+) cells in the ipsilateral hippocampal GCL of
the 5-Flu-HI group was significantly higher (~50%) than in the Ns-HI group (p < 0.01) (Fig 6A-E). The 5-Flu-HI group also had a non-significant trend of increased BrdU(+) cell density in the contralateral hippocampal GCL than the Ns-HI group. There were no differences in the newly born cell density in the hippocampal dentate hilus or the SVZ between the two groups in either cerebral hemisphere (Fig 6F, G). There was also no significant differences in volume in the hippocampal GCL or dentate hilus in either cerebral hemisphere between the two groups (Fig 6H). These results suggested that fluoxetine pretreatment increased neurogenesis by increasing precursor cell survival in the ipsilateral hippocampal GCL after HI.
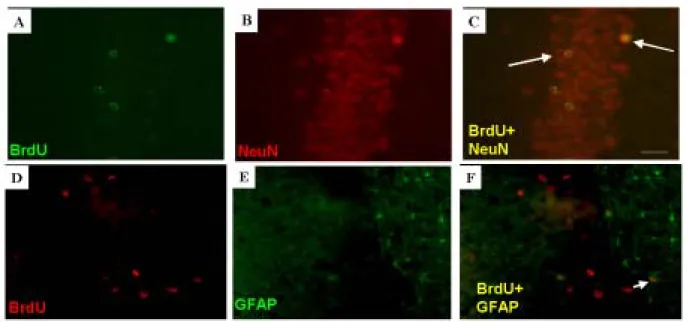
To determine the neuronal identity of BrdU(+) cells in the hippocampus, brain sections were co-labeled with antibodies to BrdU and NeuN (neuronal marker) or BrdU and GFAP (astroglial marker). The BrdU(+) cells that co-expressed NeuN in the hippocampal GCL had the
morphological characteristics of granular cells (a round or oval cell body ~20 um in diameter with long dendrites extending toward the molecular layer) (Fig 7). The morphology of these newly born cells was not different between the 5-Flu-HI and Ns-HI groups. The percentage of newly born cells that differentiated into neurons in the hippocampal GCL was similar in the 5-Flu-HI and Ns-HI groups (5-Flu-HI group left side: 78.9 ± 4 %, right side: 75.3 ± 6 %; Ns-HI group left side: 78.5 ± 4 %, right side: 73.5 ± 4 %), indicating that the differentiation was not influenced by fluoxetine treatment.
Discussion
Our study suggests that fluoxetine, a widely used SSRI antidepressant, reduces, in a dose-related manner, functional deficits in adult rats with neonatal HI brain injury. The
neuroprotective mechanisms of fluoxetine treatment before HI are associated with 1) sustained increased CREB phosphorylation in both the ipsilateral and contralateral hippocampus after HI, 2) increased neurogenesis by raising newly-born cell survival in the ipsilateral hippocampal GCL after HI. These findings provide evidence that functional recovery after HI injury in the developing brain can be induced by agents that increase neurogenesis and neural plasticity through the CREB
activation mechanism, which may suggest a potential strategy for influencing the cognitive outcome of HI injury in neonates.
Sparing neurons from death may not necessarily translate into sparing them from dysfunction, however. Furthermore, differences in cognitive function deficits may occur despite no difference in the degree of brain injury at morphological levels. Therefore, in addition to pathological evaluation, sophisticated behavioral outcome is important for evaluating the efficacy of potential treatment that may improve the consequences of experimental brain insult.28 Performance in the MWM, a device for investigating learning and memory in laboratory rats, appears to depend upon the coordinated action of different brain regions and neurotransmitter systems constituting a
functionally integrated neural network. Lesions in distinct brain regions such as the hippocampus, striatum, basal forebrain, cerebellum, or cerebral cortex impair MWM performance.29
In this study, animals were subjected to a sequence of MWM tasks that measured different aspects of learning, such as spatial orientation, long-term retention, reversal learning, and visual-oriented motor learning. Although fluoxetine pretreatment, at a low or high dose, did not provide neuroprotection at pathological levels, we found that low-dose fluoxetine reduced and, in contrast, high-dose fluoxetine exacerbated functional deficits in adult rats with neonatal HI brain injury. The functional, but not structural, benefit derived from low-dose fluoxetine pretreatment in neonatal HI brain injury may be related to increased neurogenesis in the hippocampal GCL. Recent
data suggests that the behavioral and neuroprotective effects of chronic antidepressants in adult rats are mediated by the stimulation of neurogenesis in the hippocampus.16,30 There is also
evidence that neurogenesis plays a contributory role in functional recovery after cerebral ischemia in adult rats.31 Most experimental studies of perinatal HI brain injury are concerned with how cell death can be prevented or at least minimized. However, many investigators found strong increases in neurogenesis in adult rats following a variety of brain injury paradigms, including ischemia and seizures.32,33 Furthermore, regenerated neurons are actively integrated into the existing brain circuitry and contribute to forming hippocampal-dependent memory or ameliorating neurological deficits.18 These findings suggest that instead of minimizing cell death, functional outcome after injury can be improved by the stimulation of endogenous neurogenesis. Our results suggest that strategies aimed at stimulating neurogenesis can provide new avenues for the treatment of neonatal HI brain injury.
We found that fluoxetine pretreatment was specifically beneficial for the survival, but not proliferation, of precursor cells that differentiated into new neurons in the hippocampal GCL after HI in rat pups. It may be related to significantly sustained increases in CREB phosphorylation, especially in the hippocampus after HI in fluoxetine-pretreated rats. Several signals control the proliferation, differentiation, and survival of endogenous progenitors.34 Of these, CREB activation seems to be both necessary and sufficient for ischemia-induced neurogenesis in the hippocampal dentate gyrus of adult rats. Furthermore, inhibition of CREB activation did not change
proliferation activity, but it did reduce the survival rate of precursor cells in the hippocampal GCL.35 Our results were consistent with other studies in which chronic treatment of adult rats with fluoxetine, which increases the levels of pCREB in the dentate gyrus, increased the survival of immature neurons.15,16 CREB activation and its facilitation of the survival of neuronal precursor cells may also be responsible for the hippocampus-associated learning-dependent changes of dentate gyrus neurogenesis.19,36 All our data taken together suggested that fluoxetine pretreatment increased CREB activation and neurogenesis by facilitating the survival of neuronal precursor cells
after HI injury in rat pups.
The present study also demonstrated that antidepressant regulation of neurogenesis and synaptogenesis in immature brains may be region-specific. Long-term antidepressant treatment specifically increases neurogenesis in the hippocampus but not in the SVZ,16 which suggests that antidepressants do not have a general effect on neurogenesis. CREB regulation of neurogenesis seems to be specific to the hippocampal dentate gyrus because overexpression of CREB did not alter the survival of BrdU-labeled cells in the SVZ.35 We also found a significant increase in CREB phosphorylation in the hippocampus after HI injury. Taken together, these findings suggest that CREB activation plays a contributory role in region-specific neurogenesis.
Human fetal exposure to SSRIs may cause transient neurobehavior disturbance in the
newborn period.42 Our data in rat pups shows that the manifestations of serotonergic symptoms is related with the treatment of high-dose fluoxetine: 70% of the 15-mg/kg-fluoxetine-group rats presented with serotonergic symptoms, while none of the 5-mg/kg-fluoxetine- or saline-group rats did. The occurrence of the serotonergic symptoms during high-dose fluoxetine injection may be related with worsened neurologic deficits after HI in the 15-Flu-HI group. Although clinical studies suggest that children with prenatal or perinatal exposure to fluoxetine did not have an adverse long-term outcome in their neurocognition and language development,43 fluoxetine treatment during early brain development altered emotional behavior in adult mice.25 Our data show no long-term adverse cognitive or emotional effects of early-life low-dose fluoxetine treatment, as revealed by the MWM, open field, and elevated plus maze tests in adult rats. This discrepancy in animal studies may be because we used a lower dose (5 mg/kg vs. 10 mg/kg) and a shorter period of treatment (7 days vs. 18 days).
We found that the mortality rate during HI could also be significantly reduced in rat pups pretreated with fluoxetine. Apnea arising from failures of the respiratory center during hypoxia is one of the important causes of mortality during HI. Serotonergic axons originating from the caudal raphe nuclei in the ventral brainstem project to the brain and spinal cord, including the lower
brainstem and the pons, where essential elements for the respiratory network are located. Respiratory long-term facilitation, a serotonin-dependent central neural mechanism, is a long-lasting increase in ventilation or respiratory motor nerve activity that can be induced by hypoxia.44 In addition, drugs directed to the defined 5-HT receptor subtypes, such as 5-HT1AR, 5-HT4R, and 5-HT7R, have successfully been used to treat various respiratory disturbances, including apnea or respiratory depression.45 Therefore, it is likely that more stable breathing induced by serotonin-receptor signaling-activation in rat pups with fluoxetine pretreatment is involved in their reduced mortality rate during HI.
In adult rats, only long-term (> 14 days) treatment with fluoxetine can increase neurogenesis, and CREB expression in the dentate gyrus. Fourteen days is consistent with the time required for the initiation of therapeutic action of fluoxetine in human adults. However, our results indicated that short-term (7 days) fluoxetine pretreatment reduced functional deficits after HI injury in rat pups. Furthermore, there were corresponding increases in neurogenesis, and CREB phosphorylation after HI in the hippocampus. The delayed effects of fluoxetine in adult rats suggest the slow structural and long-term gene changes taking place within the limbic target areas. It appears that the
upregulated cAMP-CREB signaling cascade can accelerate the time course of gene alterations. The shortened interval for the molecular and cellular alterations after fluoxetine treatment in immature brains may be related to the development of cAMP signaling, including the developmental changes of receptor adenylate cyclase and phosphodiesterase activity.46,47 Therefore, we hypothesize that it will be more advantageous to induce neurogenesis and neuroplasticity for cell survival using low-dose fluoxetine in immature brains than in mature brains. To our knowledge, these are the first data to demonstrate that fluoxetine contributes to protecting against HI brain injury in rat pups through activating CREB-mediated cascades. Our findings raise the possibility that short-term perinatal low-dose fluoxetine treatment may have therapeutic benefits against HI injury in neonates.
References
1. Volpe JJ. Neurology of the newborn. 4th ed. Philadelphia: Saunders, 2001.
2. Huang CC, Wang ST, Chang YC, et al. Measurement of the urinary lactate:creatinine ratio for
the early identification of newborn infants at risk for hypoxic-ischemic encephalopathy. N Engl
J Med 1999;34:328-335.
3. Ferriero DM. Neonatal brain injury. N Engl J Med 2004;351:1985-1995.
4. Welberg LA, Seckl JR. Prenatal stress, glucocorticoids and the programming of the brain. J
Neuroendocrinol 2001;13:113-128.
5. Shanks N, Lightman SL. The maternal-neonatal neuro-immune interface: are there long-term
implications for inflammatory or stress-related disease? J Clin Invest 2001;108:1567-1573.
6. Lonze BE, Ginty DD. Function and regulation of CREB family transcription factors in the
nervous system. Neuron 2002;35:605-623.
7. Walton MR, Dragunow I. Is CREB a key to neuronal survival? Trends Neurosci
2000;23:48-53.
8. Lee HT, Chang YC, Wang LY, et al. cAMP response element-binding protein activation in
ligation preconditioning in neonatal brain. Ann Neurol 2004;56:611-623.
9. Thome J, Sakai N, Shin K, et al. cAMP response element-mediated gene transcription is
upregulated by chronic antidepressant treatment. J Neurosci 2000;20:4030-4036.
10. Nibuya M, Nestler EJ, Duman RS. Chronic antidepressant administration increases the
Neurosci 1996;16:2365-2372.
11. Dowlatshahi D, MacQueen GM, Wang JF, Young LT. Increased temporal cortex CREB
concentrations and antidepressant treatment in major depression. Lancet 1998;352:1754-1755.
12. Nibuya M, Morinobu S, Duman RS. Regulation of BDNF and trkB mRNA in rat brain by
chronic electroconvulsive seizure and antidepressant drug treatments. J Neurosci
1995;15:7539-7547.
13. Saarelainen T, Hendolin P, Lucas G, et al. Activation of the TrkB neurotrophin receptor is
induced by antidepressant drugs and is required for antidepressant-induced behavioral effects. J
Neurosci 2003;23:349-357.
14. Nestler E, Barrot M, Dileone RJ, et al. Neurobiology of depression. Neuron 2002;34:13-25.
15. Nakagawa S, Kim JE, Lee R, et al. Regulation of neurogenesis in adult mouse hippocampus by
cAMP and the cAMP response element-binding protein. J Neurosci 2002;22:3673-3682.
16. Malberg J, Eisch A, Nestler E, Duman RS. Chronic antidepressant treatment increases
neurogenesis in adult rat hippocampus. J Neurosci 2000;20:9104-9110.
17. Fujioka T, Fujioka A, Duman RS. Activation of cAMP signaling facilitates the morphological
maturation of newborn neurons in adult hippocampus. J Neurosci 2004;24:319-328.
18. Nakatomi H, Kuriu T, Okabe S, et al. Regeneration of hippocampal pyramidal neurons after
ischemic brain injury by recruitment of endogenous neural progenitors. Cell 2002;110:429-441.
19. Gould E, Beylin A, Tanapat P, et al., Learning enhances adult neurogenesis in the hippocampal
20. Dam M, Tonin P, De Boni A, et al. Effects of fluoxetine and maprotiline on functional recovery
in poststroke hemiplegic patients undergoing rehabilitation therapy. Stroke 1996;27:1211-1214.
21. Pariente J, Loubinoux I, Carel C, et al. Fluoxetine modulates motor performance and cerebral
activation of patients recovering from stroke. Ann Neurol 2000;50:718-729.
22. Horsfield SA, Rosse RB, Tomasino V, et al. Fluoxetine's effects on cognitive performance in
patients with traumatic brain injury. Int J Psychiatry Med 2002;32:337-344.
23. Chang YC, Huang AM, Kuo YM, et al. Febrile seizures impair memory and cAMP
response-element binding protein activation. Ann Neurol 2003;54:706-718.
24. Balduini W, De Angelis V, Mazzoni E, Cimino M. Simvastatin protects against long-lasting
behavioral and morphological consequences of neonatal hypoxic/ischemic brain injury. Stroke
2001;32:2185-2191.
25. Ansorge MS, Zhou M, Lira A, et al. Early-life blockade of the 5-HT transporter alters emotional
behavior in adult mice. Science 2004;306:879-881.
26. Nowakowski RS, Lewin SB, Miller MW. Bromodeoxyuridine immunohistochemical
determinations of the lengths of cell cycle and the DNA-synthetic phase for an anatomically
defined population. J Neurocytol 1989;18:311-318.
27. Tzeng SF, Wu JP. Responses of microglial and neural progenitors to mechanical brain injury.
Neuroreport 1999;10:2287-2292.
28. Corbett D, Nurse S. The problem of assessing effective neuroprotection in experimental cerebral
29. D’Hooge R, De Deyn PP. Applications of the Morris water maze in the study of learning and
memory. Brain Res Rev 2001;36:60-90.
30. Santarelli L, Saxe M, Gross C, et al. Requirement of hippocampal neurogenesis for the
behavioral effects of antidepressants. Science 2003;301:805-809.
31. Raber J, Fan Y, Matsumori Y, et al. Irradiation attenuates neurogenesis and exacerbates
ischemia-induced deficits. Ann Neurol 2004;55:381-389.
32. Liu J, Solway K, Messing RO, Sharp FR. Increased neurogenesis in the dentate gyrus after
transient global ischemia in gerbils. J Neurosci 1998;18:7768-7778.
33. Madsen TM, Treschow A, Bengzon J, et al. Increased neurogenesis in a model of
electroconvulsive therapy. Biol Psychiatry 2000;47:1043-1049.
34. Barnabe-Heider F, Miller FD. Endogenously produced neurotrophins regulate survival and
differentiation of cortical progenitors via distinct signaling pathways. J Neurosci
2003;23:5149-5160.
35. Zhu DY, Lau L, Liu SH, et al. Activation of cAMP-response-element-binding protein (CREB)
after focal cerebral ischemia stimulates neurogenesis in the adult dentate gyrus. Proc Natl Acad
Sci USA 2004;101:9453-9457.
36. Tully T, Bourtchouladze R, Scott R, Tallman J. Targeting the CREB pathway for memory
enhancers. Nature Rev Drug Discovery 2003;2:267-277.
37. D’Sa C, Duman RS. Antidepressants and neuroplasticity. Bipolar Disord 2002;4:183-194.
39. Jovanovic JN, Czernik AJ, Fienberg AA, et al. Synapsins as mediators of BDNF-enhanced
neurotransmitter release. Nat Neurosci 2000;3:323-329.
40. Greengard P, Valtorta F, Czernik AJ, Benfenati F. Synaptic vesicle phosphoproteins and
regulation of synaptic function. Science 1993;259:780-785.
41. Gomez-Pinilla F, So V, Kesslak JP. Spatial learning induces neurotrophin receptor and synapsin
I in the hippocampus. Brain Res 2001;904:13-19.
42. Zeskind PS, Stephens LE. Maternal selective serotonin reuptake inhibitor use during pregnancy
and newborn neurobehavior. Pediatrics 2004;113:368-375.
43. Nulman I, Rovet J, Stewart D, et al. Neurodevelopment of children exposed in utero to
antidepressant drugs. N Engl J Med 1997;336:258-262.
44. Feldman JL, Mitchell GS, Nattie EE. Breathing: rhythmicity, plasticity, chemosensitivity. Annu
Rev Neurosci 2003;26:239-266.
45. Richter DW, Manzke T, Wilken B, Ponimaskin E. Serotonin receptors: guardians of stable
breathing. Trends Mol Med 2003;9:542-548.
46. Slotkin TA, Tate CA, Cousins MM, Seidler FJ. Beta-adrenoceptor signaling in the developing
brain: sensitization or desensitization in response to terbutaline. Dev Brain Res
2001;131:113-125.
47. Lannutti BJ, Schneider LE. Gprk2 controls cAMP levels in Drosophila development. Dev Biol
Figure 1. Low-dose fluoxetine pretreatment reduced functional deficits after hypoxic-ischemia (HI)
as measured by the Morris water maze. (A) During the acquisition and reversal phases, the normal saline (Ns), 5-mg/kg fluoxetine-pretreated (5-Flu-HI), and normal saline-pretreated (Ns-HI) groups all learned to find the submerged platform (p < 0.001). However, the total distance covered to reach the submerged platform was significantly different between the three groups (p < 0.001). Multiple comparisons revealed that both HI groups traveled significantly longer distances than did the
Ns-group (both p < 0.001). The 5-Flu-HI-group also traveled a significantly shorter distance finding the platform than did the Ns-HI group (*p < 0.05 for the acquisition; #p < 0.005 for reversal phase). (B) During probe test 1, the Ns group spent more time in the target quadrant than did the two HI groups (p < 0.001);there was no significant difference between the 5-Flu-HI and Ns-HI groups. However, in probe test 2, the 5-Flu-HI group spent significantly more time searching in the new target quadrant than did the Ns-HI group (*p < 0.05). There was no significant difference between the 5-Flu-HI and the Ns group. (C) In the cued version, the 5-Flu-HI group spent significantly less time reaching the visible platform than the Ns-HI group did (‡ p < 0.001), and the 5-Flu-HI group spent significantly more time than the Ns group (p < 0.01) did. The Ns group spent significantly less time than the other two groups. (D) In contrast, the three groups did not differ in swimming speed measured across the training sections.
Figure 2. High-dose fluoxetine pretreatment exaggerated functional deficits after hypoxic-ischemia
(HI) as measured by the Morris water maze. (A) In both the acquisition and reversal training, the travel distance searching for the submerged platform in the 15-mg/kg fluoxetine-pretreated (15-Flu-HI) group did not decrease over time. Multiple comparisons showed that the 15-Flu-HI group traveled a significantly longer distance than the normal saline (Ns) (both p < 0.001) or normal-saline-pretreated (Ns-HI) group did (both #p < 0.005). (B) There was no significant difference between the 15-Flu-HI and Ns-HI groups in either probe test. The Ns-group rats spent more time in the target quadrant than did the two HI groups (both p < 0.001). (C) In the cued version, there was also no significant difference between the 15-Flu-HI and Ns-HI groups in time spent reaching the visible platform. The Ns group spent significantly less time than did the two HI groups (both p < 0.001). (D) There was also no significant difference between the three groups in swimming speed.
Figure 3. Fluoxetine pretreatment did not induce neuroprotection against hypoxic-ischemia (HI) as
measured by cerebral hemispheric weight reduction. (A) The degree of brain injury, as measured by hemispheric weight reduction, showed no significant difference between the 5-mg/kg
fluoxetine-pretreated (5-Flu-HI), 15-mg/kg fluoxetine-pretreated (15-Flu-HI), and
normal-saline-pretreated (Ns-HI) groups. (B) There were no significant differences in body weight measured from P1 to P7 or from P14 to P28 between the Ns-HI, 5-Flu-HI, 15-Flu-HI, and normal saline (Ns) groups.
Figure 4. Low-dose fluoxetine pretreatment increased CREB phosphorylation (pCREB) in the
hippocampus and cortex after hypoxic-ischemia (HI). (A-C) The 5-mg/kg fluoxetine-pretreated (5-Flu-HI) group had significantly higher pCREB levels in the ipsilateral hippocampus 12 hours and 72 hours after HI than did the normal-saline-pretreated (Ns-HI) group at the respective time point (*both p < 0.05). The 5-Flu-HI group also had significantly higher pCREB levels in the ipsilateral cortex 72 hours after HI than did the Ns-HI group (*p < 0.05). Compared with the Ns-HI group, the 5-Flu-HI group had significantly higher hippocampal pCREB levels in the contralateral hippocampus and cortex 12 hours and 72 hours after HI (*all p < 0.05). Total CREB was unchanged between the two HI groups. L: left hemisphere, R: right hemisphere.
Figure 5. Daily 5-mg/kg fluoxetine for 7 days from P1 to P7 did not alter progenitor cell
proliferation in the immature rat brain. Unbiased stereological cell counts revealed no significant differences in the numbers of bromodeoxyuridine (BrdU)(+) cells in the subventricular zone (SVZ) between the normal-saline (Ns) (A) and 5-mg/kg fluoxetine (5-Flu) groups (B, F). BrdU(+) cell density in the hippocampal granule cell layer (GCL) and hilus of the dentate gyrus in the 5-Flu group (D) also did not differ from that of the Ns group (C, E). Scale bar = 100 µm.
Figure 6. Fluoxetine pretreatment increased neurogenesis by increasing precursor cell survival in
the ipsilateral hippocampal granule cell layer (GCL) 14 days after hypoxic-ischemia (HI). Quantitative analysis of bromodeoxyuridine (BrdU) immunoreactivity revealed that the 5-mg/kg fluoxetine-pretreated (5-Flu-HI) group had an ~50% higher density of BrdU(+) cells in the
ipsilateral hippocampal GCL (A) than did the normal-saline-pretreated (Ns-HI) group (B) (E) (*p < 0.01). The 5-Flu-HI group also had a non-significant trend of increased BrdU(+)-cell density in the contralateral hippocampal GCL (C) than the Ns-HI group did (D). There were no differences in the density of the newly born cells in the hippocampal dentate hilus (F) or the SVZ (G) between the two groups in either cerebral hemisphere. Nor was there any significant difference in volume (H) in the hippocampal GCL or dentate hilus in either cerebral hemisphere between the two groups. Scale bar = 100 µm. L: left hemisphere, R: right hemisphere.
Figure 7. The neuronal identity of bromodeoxyuridine (BrdU)(+) cells in the hippocampal dentate
gyrus after hypoxic-ischemia (HI). Double immunofluorescent staining showed that the BrdU(+) cells (A, D) were neuron-specific nuclear-protein (NeuN) immunoreactive (B), not glial-fibrillary acidic-protein (GFAP) immunoreactive (E). Merged image C (from A and B) shows the
co-localization of BrdU(+) (green) with NeuN(+) (red) in cells indicated as yellow nuclei (arrows). Merged image F (from D and E) shows that most of the BrdU(+) (red) cells in the granular cell layer were not co-localized with GFAP(+) cell (green). Co-localization of BrdU with GFAP is shown in cells in the hilus with green cytoplasm surrounding red nuclei (small arrow). Scale bar = 25 µm.