國立宜蘭大學森林暨自然資源學系碩士論文
Department of Forestry and Natural Resources National Ilan University
Master Thesis
大礁溪實驗林場疏伐木初期木棲性真菌調查 The Primary Wood-inhabiting Fungi on Thinning Tree
Trunks of the Experimental Forest of National Ilan University
指導教授:林亞立博士
Ya-Lih Lin Ph.D.
研究生:曾詩涵
Shih-Han Tseng
中華民國九十九年一月
摘要
本研究以大礁溪實驗林場木棲性真菌為研究對象,於 2008 年 3 月 至2009 年 2 月,每季採樣 2 次,合計共 8 次,時間為期 1 年,對堆置於 標本園、肖楠林及楓香林試驗區內之相思樹、樟樹及白匏子樣木進行木 棲性真菌的取樣、分離、培養及保存,並詳細紀錄試驗區環境及樣木上 子實體出現等相關資料,目的在了解疏伐木上初期木棲性真菌之種類、
出現次序及可能之影響因子。
樣木圓盤菌種分離鑑定結果得,擔子菌 6 種、子囊菌類 1 種、不完全 菌 44 種,調查期間出現在樣木上的大型真菌子實體分別有擔子菌類的木 耳 (Auricularia auricular-judae)、毛木耳 (Auricularia polytricha)、簇生 鬼傘 (Coprinus disseminatus)、裂褶菌 (Schizophyllum commune)、略薄多 孔菌 (Polyporus tenuiculus)與平伏菌類等 6 種;子囊菌類的炭皮菌類 (Biscogniauxia sp.)、炭角菌類 (Xylaria sp.)等 2 種,在本次調查共確認發 現擔子菌12 種、子囊菌 3 種及不完全菌種 44 種。整體而言,不完全菌類 在本研究中出現之菌種數是最高的,其中
Botryosphaeria
、Fusarium
、Pencillium
、Phaeoacremonium
、Trichoderma 這五個屬之菌種為分離次數
較高之菌屬。在研究中也發現下四種菌種Campylocarpon fasciculare
、Pleurostoma ootheca
、Phaeoacremonium inflatipes
、Phaeoacremonium rubrigenum 疑似台灣新紀錄種。
在研究過程中發現不同樹種的菌種出現數量有相當大的差異性,菌種 出現之種類數以樟樹樣木之出現種類數最少,最多為白匏子,但是在三試 驗區比較下則是以標本園最多。於各樹種樣木分邊心材計算時,菌種種類 出現最高還是白匏子樣木,但在不論哪種樹種樣木,在菌種種類出現數邊 材均大於心材。
在環境因子中除了土壤濕度與林內光度外均無明顯之差異性。因此,
推測影響菌種出現可能之原因在於以上兩個因子。除了環境因子的影響 外,調查期間觀察到菌種對於樹種可能具有偏好性,且邊心材的差異也會 影響菌種出現及分離的頻率。有 7 種菌種僅出現在相思樹樣木,4 種在樟 樹樣木上及白匏子樣木上有6 種菌種。同時出現在三種樣木上則有 9 種菌 種。
關鍵詞:大礁溪實驗林場、木棲性真菌、疏伐木
Abstract
A survey on the wood-inhabiting fungi on thinning tree trunks of the Experimental Forest of National Ilan University was conducted from March 2008 to February 2009. Sample discs from piled trunks were collected twice per season and eight times in the period of one year. Trunks of Taiwan Acacia (Acacia confusa), Comphor Tree (Cinnamommum camphora) and Turn-in-the-wind (Mallotus paniculatus) were stacked in three locations, the specimen garden, the , Taiwan Incense-cedar and the Formosan sweet gum stands as specimens for fungal isolation sampling. Environment factors, such as temperature and humidity of air and soil, and the fruiting bodies presented on the trunks were collected and recorded.
Isolation and identification of fungi from the sample discs reveal 6 basidiomycetes and 1 ascomycetes and 44 deuteromycetes in the first year of piling. Six basidiomycetous fruiting bodies of Auricularia auricular-judae,
Auricularia polytrich, Coprinus disseminatus, Schizophyllum commune, Polyporus tenuiculu, and one with resupinate fruiting body alone with two
other ascomyces , Biscogniauxia sp. and Xylaria sp. were collected from the piled trunks.The total fungi species isolated and recognized from this study were 12 basidiomyces, 3 ascomyces, and 44 deuteromyces. Among these fungi, species of Botryosphaeria, Fusarium, Penicillium, Phaeoacremonium, and
Trichoderma genera were most frequently encountered ones. Four suspected
newly recorded species of Taiwan, Campylocarpon fasciculare, Pleurostomaootheca, Phaeoacremonium inflatipes, Phaeoacremonium rubrigenum were
isolated and identified.Differences of species and their isolation frequency among the three tree species and piling location were found, of which the Turn-in-the-wind and the
specimen garden showed highest fungal species number and isolation frequency. When comparing the fungal isolation frequency from between heartwood and softwood of a tree species, the sapwood possesses higher isolation frequency than the heartwood.
Soil humidity and canopy coverage were the only environmental factors indicated obvious differences among the three piling locations which may be taken as the influence factors on the presents of fungal species and their isolation frequencies. Besides the environmental factors, differences occur with varied piled tree species and between heartwood and softwood obviously rendered the varied species appearance and their isolation frequencies.
Host specificity was noticed by the 7 unique species isolated from the specimens of Taiwan Acacia and the 4 and 6 unique species from the specimens of Comphor Tree and Turn-in-the-wind separately. Fungi with generality were also observed by the 9 fungal species occurred on all the tree species piled.
Keywords:Experimental Forest of National Ilan University、wood-inhabiting
fungi、
thinning tree trunks謝誌
在研究所就讀的這段期間,感謝林亞立老師在這段時間對學生的提攜 與指導,在此深深的感謝老師,謝謝老師在學生研究的這一路上以自我學 習的方式讓學生成長,除了讓學生學到獨立研究的精神外,並讓學生能在 研究的過程中從錯誤學習經驗,不斷的精進自己。文稿初成,除感謝林亞 立老師耐心的指導外,並感謝曾顯雄博士、張東柱博士以及陳素瓊博士費 心審稿,並賜諸多寶貴意見,使論文能更臻嚴謹與完善,在此由衷的感謝。
在實驗過程中,感謝系上林場技師黃文哲先生、黃鶯小姐、陳小玲小 姐以及系上的各位老師們在研究過程中,提供諸多的幫忙與協助。在就讀 的這段時間,感謝同學銘倉、莉純、彥新、廷諭;學弟妹小布、文傑、小 多、博純、當能、思蜜、婕瑜、姿文、致儀、台大的子葳在研究的過程中,
協助我的實驗、陪伴著我,讓我在這宜蘭的這段時間裡,過的多彩多姿。
另外要感謝我的那群好朋友玫利、宜佳、書婷、秀蓉、婉婷、文斌,謝謝 你們,每當我碰到困境時,總是第一時間跳出來陪伴著我,真的很謝謝你 們。
最後,要感謝我的家人,最強力的後盾,總是默默的關心著我,支持 著我,讓我很安心的在宜蘭從大學讀到研究所畢業,在此謝謝我可愛的家 人,有你們我的真的很幸福。最後~最後~感謝鏡光,一直以來總給我鼓勵 與支持,每當我在鬧情緒時,默默的聽我說,陪著我,謝謝你對我的付出,
謝謝。
一個階段的結束,代表人生又要走到下一個階段,電影總是會散場 的,宜蘭~研究生活在此劃下據點,在此感謝所有曾經幫助過我的人,謝 謝你們,謹將此論文獻給你們。
詩涵 2010.01.27
目 錄
摘要... i
Abstract ...iii
謝誌... v
目 錄 ... vi
圖目錄...viii
表目錄... ix
附表目錄... x
附圖目錄...xii
壹、前言... 1
貳、文獻回顧... 3
一、台灣真菌調查研究... 3
二、真菌鑑別... 5
三、環境與樹種之探討... 6
叁、材料方法... 8
一、大礁溪實驗林場概述... 8
二、試驗區概述... 9
三、試驗樣木的選取... 10
四、樣木堆積與樣本採取... 10
五、樣本圓盤處理... 11
六、環境因子蒐集... 12
七、菌種分離與保存... 13
八、菌種鑑定... 14
十、資料分析與處理... 18
肆、結果與討論... 19
一、菌種名錄... 19
二、實驗區環境因子... 24
三、研究期間菌種依序出現之頻率... 35
四、試驗區與樣木樹種上出現菌種之比較... 42
五、新記錄種之描述... 47
伍、結論... 50
陸、參考文獻... 52
柒、附錄... 56
附錄一、各試驗區不同樹種在不同培養基之分離鑑定結果... 56
附錄二、試驗樣區微環境資料整理... 83
附錄三、菌種圖片... 87
圖目錄
圖1 大礁溪實驗林場分區與試驗區位置圖。... 9
圖2 圓盤取樣示意圖。... 11
圖3 聚合酶連鎖反應所增幅之rDNA之重複序列片段,及所用之廣泛引 子對(ITS-4、ITS-5)之示意圖。 ... 17
圖4 調查期間試驗區月平均溫度變化。... 25
圖5 調查期間試驗區月平均相對濕度變化。... 25
圖6 調查期間試驗區光度變化。... 26
圖7 調查期間試驗區林內鬱閉度變化。... 27
圖8 調查期間試驗區土壤溫度變化。... 28
圖9 調查期間試驗區土壤濕度變化。... 29
圖10 調查期間各試驗區中相思樹樣木含水率變化。... 32
圖11 調查期間各試驗區中樟樹樣木含水率變化。... 33
圖12 調查期間各試驗區中白匏子樣木含水率變化。... 34
圖13 調查全年各菌類群菌種出現頻率。... 35
圖14 調查期間各樣木樹種上擔子菌類出現次數。... 36
圖15 調查期間各樣木樹種上子囊菌類出現次數。... 37
圖16 調查期間各樣木樹種上不完全菌類出現次數。... 37
圖17 調查期間各樣木樹種上細菌類出現次數。... 37
圖18 Campylocarpon fasciculare在MEA培養基培養 30 天之菌落。 .... 47
圖19 Pleurostoma ootheca在MEA培養基培養 30 天之菌落。... 48
圖20 Phaeoacremonium inflatipes在MEA培養基培養 30 天之菌落。... 48 圖21 Phaeoacremonium rubrigenum在MEA培養基培養 30 天之菌落。49
表目錄
表1 擔子菌、子囊菌及細菌菌種名稱與出現次數... 20
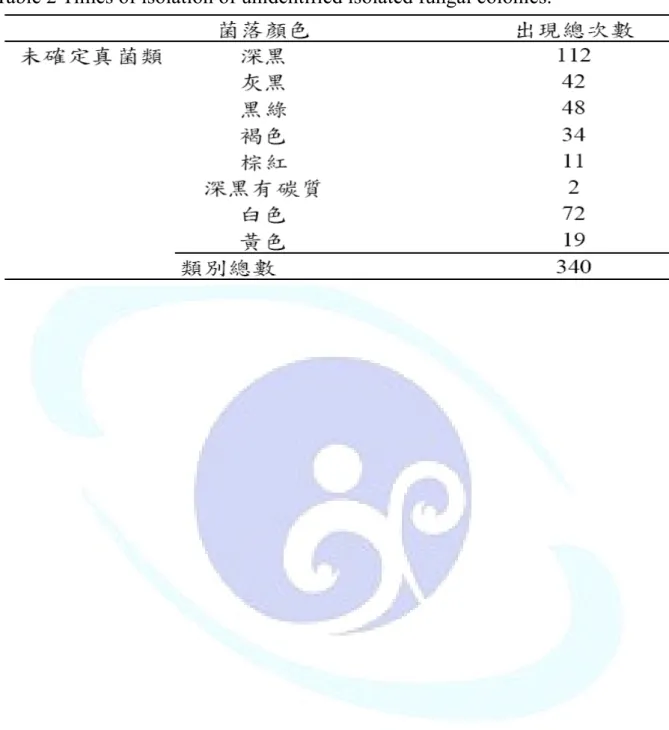
表 各項環境因子之年平均值 表 樟樹樣木上各類群菌種出現之次序 表 不同樹種合計試驗區中菌種出現表 表2 未確定之真菌種類出現次數... 21
表3 不完全菌菌種名稱與出現次數... 22
表4 各試驗區樣木上大型真菌子實體出現之描述... 23
5 ... 30
表6 調查期間各樣木平均pH值 ... 31
表7 相思樹樣木上各類群菌種出現之次序... 39
8 ... 40
表9 白匏子樣木上各類群菌種出現之次序... 41
表10 不同試驗區與樹種邊心材菌種出現表... 43
11 ... 44
表12 不同試驗區菌種出現表... 45
表13 菌種在各試驗區與樣木上出現之種類與次數... 46
附表目錄
附表1-1 標本園內相思樹樣木於MEA中分離出之菌種次數 ... 56 附表1-2 標本園內相思樹樣木於Rose Bengal-MEA中分離出之菌種次
數... 57 附表1-3 標本園內相思樹樣木於Benomyl -MEA中分離出之菌種次數5
標本園內樟樹樣木於MEA中分離出之菌種次數 ... 59 8 附表1-4
1 附表1-7
4 附表1-10
數
附表 肖楠林內相思樹樣木於 中分離出之菌種次數
附表
附表1-14 肖楠林內樟樹樣木於
附表 肖楠林內樟樹樣木於
附表1-16
附表 1-17 肖楠林內白匏子樣木於Rose Bengal-MEA中分離出之菌種次 數... 72 附表 1-18 肖楠林內白匏子樣木於Benomyl -MEA中分離出之菌種次數73 附表1-19 楓香林內相思樹樣木於MEA中分離出之菌種次數 ... 74 附表1-5 標本園內樟樹樣木於Rose Bengal-MEA中分離出之菌種次數60 附表1-6 標本園內樟樹樣木於Benomyl -MEA中分離出之菌種次數 ... 6
標本園內白匏子樣木於MEA中分離出之菌種次數 ... 62 附表1-8 標本園內白匏子樣木於Rose Bengal-MEA中分離出之菌種次
數... 63 附表 1-9 標本園內白匏子樣木於Benomyl -MEA中分離出之菌種次數6
肖楠林內相思樹樣木於MEA中分離出之菌種次數 ... 65 附表1-11 肖楠林內相思樹樣木於Rose Bengal-MEA中分離出之菌種次
... 66
1-12 Benomyl -MEA 67
1-13 肖楠林內樟樹於MEA中分離出之菌種次數... 68 Rose Bengal-MEA中分離出之菌種次數69 1-15 Benomyl -MEA中分離出之菌種次數 ... 70 肖楠林內白匏子樣木於MEA中分離出之菌種次數 ... 71
附表1-20 楓香林內相思樹樣木於Rose Bengal-MEA中分離出之菌種次
數... 7
楓香林內相思樹樣木於Benomyl -MEA中分離出之菌種次數76 5 附表1-21 7 附表1-23 9 附表1-25 附表1-27 3 附表2-3 試 4 附表2-7 各 附表 值 附表1-22 楓香林內樟樹樣木於MEA中分離出之菌種次數 ... 7
楓香林內樟樹要樣木於Rose Bengal-MEA中分離出之菌種次 數... 78
附表1-24 楓香林內樟樹樣木於Benomyl -MEA中分離出之菌種次數 .. 7
楓香林內白匏子樣木於MEA中分離出之菌種次數 ... 80
附表1-26 楓香林內白匏子樣木Rose Bengal-MEA中分離出之菌種次數81 楓香林內白匏子樣木於Benomyl -MEA中分離出之菌種次數82 附表2-1 試驗樣區月平均溫度(℃) ... 83
附表2-2 試驗樣區月平均相對溼度(%)... 8
驗區各次林內鬱閉度測量結果(%) ... 84
附表2-4 試驗區各次光度測量結果(μmol m
-2
s-1
)... 84附表2-5 試驗區各次土壤濕度測量結果(%) ... 84
附表2-6 試驗區土壤溫度測量結果(℃)... 8
試驗區樣木各次邊心材含水率(%) ... 85
2-8 各試驗區樣木各次pH ... 86
附圖目錄
附圖1-1 Phellinus於MEA培養基上所呈現之顯微特徵 ... 87
附圖1-2 Arthrinium sp. 於MEA培養基上所呈現之特徵... 87
附圖1-3 Arthrographis sp.1 於MEA培養基上所呈現之孢子形態 ... 88
附圖1-4 Arthrographis sp.2 於MEA培養基上所呈現之孢子形態... 88
附圖1-5 Aspergillus sp. 於MEA培養基上所呈現之特徵... 88
附圖1-6 Botryosphaeria rhodina於MEA培養基上所呈現之特徵... 89
附圖1-7 Campylocarpon fasciculare於MEA培養基上所呈現之特徵... 89
附圖1-8 Cladsporium sp. 於MEA培養基上所呈現之孢子形態... 89
附圖1-9 Cuvularia sp. 於MEA培養基上所呈現之特徵... 90
附圖1-10 Cylindrocarpon sp.於MEA培養基上所呈現之特徵 ... 90
附圖1-11 Eutypella sp.於MEA培養基上所呈現之特徵... 90
附圖1-12 Fusarium oxysporum於MEA培養基上所呈現之特徵 ... 91
附圖1-13 Fusarium solani於MEA培養基上所呈現之特徵 ... 91
附圖1-14 Glicocladiopsis tenuis於MEA培養基上所呈現之特徵... 91
附圖1-15 Penicillium radicum於MEA培養基上所呈現之特徵... 92
附圖1-16 penicillium sp.1 於MEA培養基上所呈現之特徵... 92
附圖1-17 penicillium sp.2 於MEA培養基上所呈現之特徵... 92
附圖1-18 Pestalotiopsis sp. 於MEA培養基上所呈現之孢子形態 ... 93
附圖1-19 Pleurostoma ootheca於MEA培養基上所呈現之產孢結構 ... 93
附圖1-20 Phaeoacremonium inflatipes於MEA培養基上所呈現之特徵 . 93 附圖1-21 Phaeoacremonium rubrigenum 於MEA培養基上所呈現之特 徵... 94
附圖1-22 Phaeoacremonium sp.1 於MEA培養基上所呈現之特徵... 94
附圖1-23 Phaeoacremonium sp.2 於MEA培養基上所呈現之特徵... 94
附圖1-24 phialophora sp. 於MEA培養基上所呈現之特徵... 95 附圖1-25 Rhinocladiella sp. 於MEA培養基上所呈現之特徵... 95 附圖1-26 Scytalidium sp. 於MEA培養基上所呈現之孢子形態... 95 附圖1-27 Trichoderma aureoviride於MEA培養培養基上所呈現之產孢
結構。... 96 附圖1-28 Trichoderma harzianum於MEA培養基上所呈現之產孢結
構。... 96 附圖1-29 放線菌類於MEA培養基上所呈現之顯微特徵 ... 96
真菌為真核多細胞生物 ,行體外消化。早期真菌因有 特徵與植物相似而被歸類於植物界,近年來生物學家普遍接受
於1969 所提出的生物五界說,將真菌獨立為一界(刑與李,2001)。
真菌分布於地球各地
赤道或是兩極均可發現其蹤跡 ,且分布廣
泛。多數的真菌為寄生性或腐生性 解者的角色,特別是在森林生
。
森林生態系中,土壤腐植質以及枯立倒木等可提供含纖維素的物質,
是腐生性真菌所喜好的生存場所。生存於枯立倒木上的真菌通常被稱為木 棲性真菌(wood-inhabiting fungi)。各類群木棲性真菌與細菌等合作分解木 材中各項組成之有機質,是森林生態系養分循環中重要的一環(池等,
2004)。木棲性真菌包含在木材上能夠生長的腐朽菌、軟腐菌、變色菌及 黴菌等,於分類上主要包含有擔子菌類、子囊菌類、盤菌類以及部份不完 全菌類,這類群真菌在森林生態系中與各類生物與非生物因子具有密切相 關,所以在森林生態系的調查與保護上具有重要意義(魏與戴,2004)。
大型真菌為能產生肉眼可見有性繁殖構造(子實體,fruiting boby)的真 菌,多數屬於擔子菌類(Basidiomycota)的銀耳目(Tremellales)、木耳目 (Auriculariales)、傘菌目(Agaricales)、多孔菌目(Polyporales)、雞油菌目 (Cantharellales)、牛肝菌目(Boletales)等 16 個目及子囊菌類(Ascomycota) 的肉座菌目(Halosphaeriales)、炭角菌目(Xylariales)及盤菌目(Pezizales) 等。在森林生態系中除了是含纖維素物質的主要分解者外,亦有與林木產 生共生關係的菌根菌。許多大型真菌具有明顯且具特色的子實體,經常引 起人們的注意,長久以來在世界各地人類飲食、醫藥、宗教、文學等方面
壹、 前言
,不具葉綠素 些
Whittaker
,舉凡高山、湖泊、草原、森林、海洋,甚至於
。生態系中,真菌族群相當龐大
,少數為共生性,在生態系中常扮演分 態系中,更是不可缺少的族群 (刑與李,
2001)
均提供了豐富的素材 (刑與李,
台灣地區目前已知的大型真菌約有 1,100 種(擔子菌約 900 種,子囊 菌約
倍的 方式
真菌在分類中包含變色菌、黴菌等的不完全菌及一些子囊菌類。
木棲
在
的研
2001) 。
200 種),以每採集 100 種大型真菌就有 20 至 40 種是台灣新紀錄種 或新種的比例推估(張等,2005),臺灣地區大型真菌約有 1,500 種。若依 英國真菌學者Hawksworth (1991),以真菌種數為維管束植物種數 4-6
估計台灣地區真菌總數,搭配1999 年台灣真菌名錄(行政院農委會,
1999),以大型真菌佔台灣所有已知真菌總數約 1/6 的比例推估,則臺灣地 區大型真菌種數應較1,500 種高出許多。姑不論哪個推估方式較為正確,
以面積比例而言,台灣地區大型真菌資源可謂十分豐富。
小型
性小型真菌在初期侵入會較快,在侵入的過程中,營造出適合其他菌 類侵入之環境條件,利於其他菌類,如腐朽菌之入侵,最後小型真菌則扮 演著清除者的角色,所以大型和小型真菌在生態系中,它們各司其職,在 森林中扮演不同角色,不論是哪種類型的真菌(土棲型、木棲型),對於 森林中生長與棲息的研究中,都非常值得去探討菌種的差異性與環境的關 係及影響。
大礁溪實驗林場真菌調查曾有伐倒木初期木生性真菌之調查(林,
1996)與大礁溪實驗林場大型真菌之調查(曾與林,2008)對此地區做初步性 究。參考上述研究後,發現林場真菌種類數在木棲性真菌這一部分特 別豐富,致使對此現象感到有相當大的興趣。本研究於試驗樣區堆積不同 樹種疏伐木之段木,藉此嘗試了解菌種在不同樹種及堆置位置上可能出現 的關係為何。期望建立基礎性資料,對於往後在於森林管理上與相關研究 上能提供協助。
、毛黴目、鐮胞菌屬、
尾子菌屬
(2000)於福山樣區調查北部低海拔常綠闊葉林之大型真 菌,記錄 47 種子囊菌和 228 種擔子菌,徐希世(1997) 於惠蓀林場水源 地針對木材腐朽菌調查研究,鑑定出 17 屬 28 種菌種;羅光傑(1998) 於 關刀溪流域對擔子菌類大型真菌調查,紀錄包含24 科 72 屬的 181 種擔子
貳、文獻回顧
一、台灣真菌調查研究
依陳瑞青教授(1991)所述,台灣地區真菌研究始於二十世紀初期,
台灣真菌調查最大貢獻者首推為日據時期的澤田兼吉氏,自1919 年開始 出版台灣菌類調查報告第一篇,歷時三十六年,總共發表了十篇,台灣地 區由日本菌類研究人員所做菌類調查共記錄發現了2,464 種真菌種類,多 數(2,382 種)為動、植物的寄生菌。
台灣光復初期至 1970 年代,真菌調查多以植病學者為主導,此時台 灣之菌類調查多限於植物寄生菌且藉各大學及農林業試驗機構之植病人 員完成。1970 年後,台灣菌類研究人員才開始增加並多元化,1975 年蔡 雲鵬博士主編統計,自1959 年至 1978 年間菌類種數增加大約 493 種,而 台灣菌類種數則累計為2,957 種。1983 年國科會生物處主動召開「台灣菌 類調查」籌備會議,並在 1985 年成立「台灣菌類調查及菌類誌編撰」計 畫,第一年度共執行七個子計畫包括水生不完全菌類
、線菌綱、黏菌綱、子囊菌及擔子菌亞群。第二年又增加兩個子 計畫分別為炭疽病屬及白粉病菌目。經過三年的努力後共得 1,984 種菌 類,分屬於518 屬,其中新紀錄種 525 種,新紀錄屬有 51 屬,世界新種 達65 種(陳,1991)。
整理近年來台灣地區大型真菌調查結果,其中吳聲華等人於1995 至 1996 年調查玉山國家公園濁水區真菌相,發現 147 種大型真菌(吳等,
1996),張東柱等人
菌標本;王也珍(1999)調查南仁山森林生態系,為低地雨林區,調查期間 發現137 種真菌,其中擔子菌類 10 種;周文能(2000) 調查蓮華池樣區,為中部低海拔常綠闊葉林,採得 276 種擔子菌類;陳 中海拔落葉林,紀錄100 多種大型真菌子實體;
王也珍與周文能
僅侷限在中央山脈的北半部,東北 部及
,亦是建 立森林資源資訊與了解木材病害的基礎工作
象 多
127 種,子囊菌類類
昇明(2001)於關刀溪調查
(2004)調查觀霧地區發現 119 種真菌,包括擔子菌 111 種,子囊菌 8 種;蘇毓智(2006)也對藤枝與扇平地區調查,發現 46 種子 囊盤菌;陳朝圳等(2006)於北大武山調查,為針闊葉混合林,也發現 65 種大型真菌;綜觀以上調查研究發現台灣地區之菌種豐富度是相當的大,
但全面性之研究,因為人力上的不足,
東部較少有人研究。
羅光傑(1998)於關刀溪流域對擔子菌類大型真菌調查及徐希世(1997) 在惠蓀林場水源地針對木材腐朽菌調查研究中,都曾經搭配生態習性、環 境因子與區域性環境來比較菌種出現頻率與菌相。本研究要探討的內容及 調查使用的方式都與前兩位大略相同,如調查次數、調查使用器具與方 法、分離純化步驟等皆得自前人研究之啟發,而後再針對本研究略予修正。
一般而言,包括細菌在內,木質殘體上木棲微生物(wood-inhabiting microorganisms)出現的先後順序為:細菌、不完全菌、子囊菌、擔子菌、
不完全菌。此順序的發生及各階段微生物出現的種類,除受外在環境條件 影響,亦與殘體內的微環境有關。伐倒木上木生性真菌出現種類與發生順 序調查,不僅為探討森林生態系殘體分解與養分循環的首要工作
(林,1996)。在池玉杰等人 (2004)的研究報告中論述木材上的微生物類群以及各類群在生物分解過 程中的演替規律,其中也提出木材分解的不同時期菌種具有明顯之協調與 演替之現 , 半是由細菌、一部份不完全菌、接合菌以及子囊菌先行侵 入,而後木材腐朽菌進入,維持一段分解穩定階段後,最後木材腐質化階 段則由青黴屬(Penicillium)、木黴屬(Trichoderma)、細菌、放線菌以及鐮刀
。Stalpers (1978)亦發表相關研究結果,其鑑 定方
不完
菌屬(Fusarium)等所取代。
二、真菌鑑別
大型真菌初始的鑑定皆是以子實體的特徵為依據,但在木材上無子實 體的出現時只能分離菌絲與菌落,在鑑定上較為困難。Nobles (1948,1965) 針對大型真菌菌絲與菌落特徵做了一系列的鑑定,其內容包含酵素產生與 反應、外觀特徵及顯微特徵等。形態上,外觀特徵包含菌落生長的速率、
顏色、味道、菌絲先端狀況、菌落邊緣等;顯微特徵則包含菌絲的樣式(骨 骼菌絲、纏繞菌絲、生殖菌絲等)、扣子體的有無、形狀、大小樣式等,
而此種鑑定方式也逐漸被採用
式除印證 Nobles 之方法外並加以修正,而在這之後菌落與菌絲之鑑 定方式也趨於成熟階段。
在菌物學概論一書中(姚與李,2002)歸納小型真菌的鑑定,說明在 1910 年 Vuilemin 是最早以分生孢子的個體發育為基礎來進行分生孢子真 菌的分類研究,而後在1933 Mason 也作相關研究。於 1953 年 Hughes 把 全菌的分類建立在產分生孢子細胞和分生孢子發育模式的的基礎 上,是為現今不完全菌形態鑑別的主要的主要依據。
Wang and Zabel (1990)針對電線桿上出現的真菌做研究,不僅僅以 Nobles 及 Stalpers 的鑑定方法探討大型真菌,另外也對小型真菌做了鑑定 方法之描述。在此時後,利用菌落鑑定,不論大型與小型木棲性真菌之鑑 定都趨向成熟。
近年來食品工業發展研究所,也針對台灣之菌種出版一系列之菌種圖 鑑(王與華,1991;彭等,1991;張等,1997;吳等,2000 ),其中包含菌 種之外觀與顯微特徵描述、不同培養基菌落之各時期、溫度表現等特徵 外,更增加了以生化特徵的蛋白質電泳圖譜依據,使得以菌落鑑定加入生 化特徵。現今在菌落鑑定時,則為形態鑑定與生化鑑定相輔相成的階段。
三、環境與樹種之探討
(heterotrophs),在森林中主要能量來源為樹木及地 面之
微生物包含細菌、真菌、放線菌、藻類及原生 生物
。各類真菌皆有其 最適
)於人工林不同疏伐強度作業對真菌組成結構與多樣性 之影
真菌是異營性生物
植被。而真菌出現之多樣性和森林有機質的分布與樹木種類、林相的 演替(sussession)以及氣候等環境條件有密切相關性(陳昇明,2001、2007)。
弓明欽等(1997)在菌根研究及應用一書中,對菌根菌的研究中指出,
真菌菌落需在合宜的環境下才能達到最適生長,土壤的養分、溫度、溼度、
pH 值及水分與林地的光照等,均是影響菌種形成的因子。在汪碧涵(2005) 報告中提及在 1988 胡弘道亦指出影響菌根菌之生長因子為溫度、溼度、
通透性、光照等,其中以溫度對於菌根菌之生長以及新陳代謝影響最為重 要。張怡珮(2009)於惠蓀林場天然闊葉林與杉木林土壤呼吸之季節變化的 碩士論文中也提到,土壤中
等,其中植物是微生物最主要的能量來源,枯枝落葉物的多寡以及組 成亦會影響微生物的出現及生長,在環境因子中最主要的則是土壤溫度。
在陳昇明(2001)的報告中亦提及真菌出現數量與土壤含水率及林內透光 率有顯著的相關性,林內透光率是決定真菌子實體出現最重要的因子,其 次為土壤含水率,多出現在於土壤含水率達45%的林地
的生存環境及條件。一般來說,木生性真菌最適的生活條件為環境溫 度22-32℃、相對溼度 70%以上,木材的酸鹼值介於 4.5-6.5、含水率 20-80%
之間。
汪碧涵(2005
響的研究報告中,提出人工林與天然林相於菌種上之差異度有明顯的 不同,在人工林中亦會因疏伐開闊的程度而有不同結果產生。陳昇明(2001) 期中報告結果也發現,於水源地之天然闊葉林較針葉林區具有較多之真菌 種類的出現。
在樹木生長過程中,常會發展某種抗禦真菌與蟲害之性質,如形成緊
密之組織結構,或由二次代謝中產生具有抑制真菌與蟲害之化學成分,此 1999)。吳季玲(2005)於台灣杉心材抗菌成分之 分析
義的
稱為天然耐腐性(王與李,
、鑑定及其抑制機制的論文中提到,天然耐腐性則泛指木材能抵抗各 種劣化因子如,天候(weathering)、腐朽菌(decay fungi)、木蠹蟲(wood borers)、白蟻(termite)及海蟲(marine borers)等之危害,而天然耐腐又可狹
定義為樹種的心材具有抗真菌腐朽之能力。通常不同樹種的天耐腐性 均有差異,而同樹種間心材與邊材的真菌抗性又以心材大於邊材,主要歸 因於心材中含有較高之抽出物成分。Hawley 等人(1924)之研究也證實心材 耐腐性大於邊材,並証明心材中微量抽出成分對於真菌以及一些微生物具 有抑制生長之效果。
從相關文獻中可見,台灣東北部菌類資源調查資料不如其他地區完 整,以宜蘭地區來看,除了近年來僅有福山植物園(張等,2000 )與宜蘭大 學大礁溪實驗林場(林,1996;曾與林,2008)有菌類資源調查外,其餘均 無相關類似調查研究。而在以上大礁溪實驗林場菌類研究中,發現木棲性 真菌為佔有較大數量之類群,因對此現象具有濃厚興趣,故本研究,以木 棲性真菌作為探討主題,希望藉此能更加了解木棲性真菌在環境中扮演的 角色與其對環境資源的利用方式。
布在全區中;土壤呈酸性,pH 值介 4.6-5.0 之間。
(四)植群與分區
大礁溪實驗林場以蘭花溪為界,分東西兩區。東區主要是以保安 林與次生林為主之竹仔崙山,其地形陡峭、脆弱;西半部則是以次生 林區與經濟林區為主,如圖1。林場以次生闊葉林為主,依林分區分 分為1.經濟林區,此區地形平緩,受風情形小,主要造林樹種有木荷、
烏心石、殼斗科、肖楠、台灣櫸、光臘樹、相思樹、木油桐及楓香等。
2.景觀林區,位於實驗林場東北方,區內為陡峭地與平坦地交錯之地 形,目前該區以相思樹及木油桐等人工造林地居多,且有天然下種之
叁、材料方法
一、大礁溪實驗林場概述
(一)位置
大礁溪實驗林場位於宜蘭縣礁溪鄉二結村山區,經度121°40' 51",緯度 24°47'31",全境面積 173.45 公頃,東、北與私有地為界,
西鄰政府公有地,地形由西北向東南傾斜,海拔在125-670 公尺之間,
蘭花溪貫穿全區,將林場分為東西二部分。
(二)氣候概況
大礁溪實驗林場,因受周圍高山阻擋,雖位於東北季風區,但冬 季吹西風及西南風;夏季則吹東風及東北風。因處於季風區內故年平 均降雨量達2,700 ㎜以上,年降雨日在 200 日以上,年平均相對溼度 約86%,年平均溫度約為 22.5℃。
(三)土壤
大 礁 溪 實 驗 林 場 土 壤 質 地 為 細 質 地 的 坋 質 粘 壤 土 , 新 成 土 (Entisols)與弱育土(Inceptisols)分
於
樹種侵入造林地。3.保安林區,此區為實驗林場主要溪流-蘭花溪二
側之河谷地,地勢陡峭、 蘭花溪為實驗林場主要水源
地,因此做為水源涵養功能為主的保安林區。4.次生林區,此區為實 形險惡,交通開發困難,且此區早期人工造林 然更新為次生林相,為次生林之保護區。
二、試驗區概述
0、260 及 500 公尺之標 楓香林等三處,如圖1 所示。
地質不穩,但因
驗林場東南方,該處地 失敗後已天
(一)試驗區位置
試驗區設於實驗林場內海拔高分別約為20 本園、肖楠林與
●為試驗區中樣木堆積位置。
Compartments of the experimental forest and locations of sample logs piling
圖1 大礁溪實驗林場分區與試驗區位置圖。虛線 為林道路線;
Fig.1
trails, ● indicates piling spots.
( )
spots. indicates
取
園進行疏伐作業,作業後以數量較多之相思樹 (Acacia
Eber uell.)大戟科等三個樹
種,選擇直徑介於15-25 公分,長度約 100 公分的段木各三支,作為試驗 樣木。
四、樣木堆積與樣本採取
從2008 年 3 月至 2009 年 2 月,每季採樣 2 次,合計共 8 次,時間為 期1 年。堆積疏伐木真菌調查中以主要木棲性真菌為主要對象,包含大型 與小型真菌,試圖探討其在不同環境、樹種與微環境條件下,菌種出現之 關係與順序為何。以下分別介紹堆積與採樣之方法。
(一)堆積方法
在標本園、肖楠林、楓香林三個試驗區中,選定離林道5 公尺以 上無雜草覆蓋之空隙,分別放置相思樹、樟樹及白匏子樣木各一段。
(二)取樣方法
在每一試驗區於三個樹種樣木中,以鏈鋸向內離端2 公分處鋸 2 公分厚度之圓盤,將其帶回實驗室進行菌種分離與培養。
(二)微環境概述
三個試驗區中,標本園環境與土壤偏乾燥且含石率高;肖楠林環 境與土壤偏潮濕且含石率低;楓香林則是環境與土壤偏潮濕,但土壤 中含石率高。在三個試驗區中,楓香林與標本園的土壤構成型態為新 成土,肖楠林為弱育土。
三、試驗樣木的選 2008 年 3 月標本
confuse Merr. )豆科、樟樹(Cinnamommum camphora (L.) Nees &
m)樟科及白匏子(Mallotus paniculatus (Lamk.)M
五、樣本圓盤處理
心材分別進行取樣。
(一)菌種分離用
菌種採集與分離時,將圓盤分為三等分,也就是每120°取一樣 木條(如圖 2),並將邊材、
圖2 圓盤取樣示意圖,S 表邊材;H 表心材
Fig. 2 Indication of sample strips collection from the sample disc.
:heartwood.
一項因子。試材含 方法為,將樣本圓盤取完木條後剩餘部份,分邊心材 取試
:
W
2
:試材絕乾重量 S:sapwood, H(二)含水率測定
由於木材是一種吸濕性的材料,它的重量與體積容易隨著水分而 變化,在林地中堆積木材時,菌種的存在與木材中含水率的高低常有 明顯的相關性,所以了解樣木的含水率是很重要的
水率測定採用的
材放置於烘箱,在103±2℃的溫度下將試材烘至恆重(絕乾),操 作程序依中國國家標準CNS 452 木材含水率試驗法進行之。含水率 依下列公式計算
含水率(%) = ((W
1
-W2
)/ W2
)*100 W1
:試材乾燥前重量(三)pH 值測定
的方法。將各次取得之樣本圓盤分邊心材磨成 過濾篩網過濾後,取等同於1 g 絕乾重之氣乾木粉 加入
15 分鐘,取上清液 以pH meter ( Suntex, SP-701 )測定酸鹼值。
六、環境因子蒐集
(一)溫度與相對溼度
氣溫和相對濕度的測定,是以全天候溫度/濕度監測盒( HOBO, H8 Pro Series, Onset., USA )置於三個試驗區內,記錄全年度區內溫濕 度。
(二)光照
光度測定使用光度計量器測量(Light meter, LI-COR, USA),每次 測量時以樣木擺放的中心點,朝向樣木堆積的下坡處為基準,分別測 定中、上、下、左、右五點後,予以平均。
(三)林內樹冠鬱閉度
以 林 內 樹 冠 鬱 閉 度 測 定 儀( Spherical Densiometer Model-C, Forestry Suppliers, Inc, USA )觀測每次取樣時林內鬱閉度值。觀測時,
分別於堆積樣木之左邊與右邊各測定一次,每次測定點之位置均固 定,以確保數值之可靠性。因樹冠鬱閉度儀內數值相加僅能達96%;
為使計算率值達 100%,故在計算時,需將測定後之數值乘以 1.04,
使能符合100% 之計算率。
(四)土壤溫度與溼度
土壤溫度使用Rixen 公司 T-600 digital thermometer;土壤濕度使 用Eijkelkamp 公司的 TDR 土壤水分測定儀器作測量,每次調查時放
參照Hatton (1970) 木粉,用1 mm 的
20 ml 的蒸餾水中,用震盪培養機,以 250 rpm 震盪 1 分鐘後,
靜置15 分鐘後,再震盪 15 分鐘,震盪完後靜置
壤中一分鐘後測定數值,重複兩次並將數值予以平 均。
七、 種
菌種的分離在於為了將單一菌落菌種分離出,方便後續之研究與鑑定
,可對後續菌種在相關研究時 有實
(一)培
芽抽出物培養基(MEA),木塊培養時每 1L 純水中加入 15 g malt 20 g agar。純化菌株時,每 1L 純水中加入 20 g malt extract 及20
l 是一種染劑,能抑制細菌生長,並控制快速生
0 ppm,即 1 L MEA 中分別加入 0.01 g 置於各試驗區土
本研究在測定土壤溫溼度時,選用之樣點為樣木堆積範圍內半徑 30 公分處,為確定每次調查之數值之有效性,每次取樣調查時,均 固定測定區域。
菌 分離與保存
時之用,且鑑定後利用保存技術將菌種留存 際之依據與憑證。
養基
採用Difco 公司的麥芽抽出物(malt extract)及瓊脂粉末(agar)所製 成之麥
extract 及 g agar
(二)抑制藥劑
1.孟加拉玫瑰紅(Rose Bengal) Rose Benga
長的黴菌,須在無光下使用,如照設紫外燈則會失去效用。本實 驗搭配 MEA 使用劑量為 200 ppm,即 1 LMEA 中加入 0.2 g Rose Bengal。
2.Benomyl
此藥劑主要為選擇擔子菌類的真菌使用,本實驗搭配 MEA 使用劑量為 10 ppm 與 10
與 0.1 g benomyl。
EA 使用劑量為 30 ppm,即 1 LMEA 中加入 0.03 g 鏈黴素。
(Penicillin G sodium)
6 g 青黴素-G 鈉。
取
培養基中培養,俟菌落長出後加以分離。
化目的在於分離出單一純系的菌 種,以方
探 通
(五
,將菌種接種於培養管之MEA 斜面,於 25℃下培 養1-2 星
八、菌種
菌種鑑定之目的除確定菌種屬別與分類群外,並藉由鑑定之過程,了 菌種之特性並且加以紀錄,以利後續相關研究使用。以往真菌在鑑定
3.鏈黴素(Streptomycin sulfate)
此藥劑主要為在分離過程中,因常會出現細菌之干擾,致使 單一菌落無法發育良好,故添加在培養基中具抑制細菌之效果。
本實驗搭配 M
4.青黴素-G 鈉
此藥劑與鏈黴素使用之用途相同,亦是添加於培養基中,具 有抑制細菌之效果。本實驗搭配MEA 使用劑量為 60 ppm,即 1 L MEA 中加入 0.0
(三)樣點採
將圓盤攜回實驗室後,依120°角分三等分採取木條樣本,蔭乾 後再將各木條分邊心材取樣,如圖2。取樣時剪取厚度約 2 mm、邊 長約 5 mm 之等腰三角形的樣片 10-15 個備用,樣片用火表面殺菌 後,在放置
(四)分離
將培養出的菌落進行純化,純
便後續鑑定工作的進行。在進行純化時,將想要的菌種用 針或適當的器具,如解剖刀片,挑起一部份接種至另一培養基,
常這個步驟必需重複 2 次以上才能得到純化的菌種。
)保存
菌種之保存
期,待菌落長滿斜面後,將培養管移至 4℃冰箱進行保存。
鑑定
解
時,常利 微鏡下觀察其各種特徵作為鑑定 依據;而現今
以下為本
(一)
實體在鑑定時,會依照其外觀型態先初步分類(如子
實體 再依照其他相關特徵鑑定到
科或屬,
(二)
在培養大型與小型真菌時,於培養基之選定上就已經 有
型真菌將菌種以 MEA 培養,待至產出產孢結構後,而後 於
繪,如各 產孢結構特徵、產孢方式、孢子形狀、大小、顏色、紋路 等
型態特徵參酌相關之期刊、論文與專書等,(Samuels et al., 2002;
2009)加以比對觀察分其大類後,再依照其各屬別之顯 用培養出之單一菌種菌落,在顯
因為分子鑑定之興起,在形態鑑定外可加此方法作為輔助,
研究中使用之鑑定方法。
大型真菌子實體 大型真菌子
的型態與質地、子實層的樣式等),
最後觀察其顯微特徵鑑定到種。(周與張,2005) 菌落型態
1.光學顯微鏡觀察 本研究
所分別,主要是以加入benomyl 藥劑之 MEA 培養,先將大型 真菌的菌落分出,而後在觀察時,除以觀察菌落是否具有扣子體 為主要依據外,另外利用分子生物技術(PCR)之方法將菌種分別 鑑定。
小
顯微鏡下觀察;而針對產孢結構較不一之菌株,則利用馬鈴薯 -紅蘿蔔培養基(potato-carrot agar,PCA)促進菌種產孢。於菌種產 孢後,將菌種先於實體顯微鏡下,挑出一小部份製成玻片,而後 置於光學顯微鏡下,觀察菌種之各項特徵並且紀錄與描
菌種之
。且於觀察期間將其菌落外觀以及菌種顯微特徵攝影記錄存 檔,以便為後續觀察時之參考依據。將其發現之各項顯微與外觀
曾與黃,
微細部構造鑑定至種。
( )DNA
1.DNA ( (2009)之方法略予修改)
於已長滿單一菌落之培養皿表面置入 1 ml 的純水,將表面的 氣生菌絲與孢子刮下
1000- 1300 rpm
5 min 後,將上清液去除後,加入 500-600μl 6 CTAB (Hexadecyl trimethyl ammonium bromide),加入後先放置於 65℃
15 min 65℃下之乾浴槽
中 30 min-1hr 。 而 後 加 入 500-600 μ l 的 P. C. I.
(Phenol/Chloroform/Isoamy ,酚/氯 戊醇,等體積比 25 24 1) 1000- 1300 rpm 5 min
400 P. C. I.
於離心機中用1000 - 1300 rpm 離心 5 min 後。取 200μl 上清液 入新的微量離心管中 而後加入0.6 isopropanol ( )
DNA -20 10 min 1000 - 1300 rpm
5 min 後,取出 0 ash buffer 清洗,在 1000 - 1300 rpm 5 min 400 95
1000 - 1300 rpm 5 min
乾30-60 min,最後加入 50μl 的 TE buffer (Tris EDTA buffer) 回 -20
2.
聚合酶連鎖反應(polymerase chain reat
DNA 3
White 人(1990)、曾顯雄與黃子葳(2009)之方 法,選用增幅 ITS 序列片段之引子。
二 序列比對
的萃取方法 參照曾顯雄與黃子葳
,製成懸浮液後置入微量離心管中。將內含
孢子水的微量離心管,放置於離心機中,以 離心
5℃的 2%
之乾浴槽 後,用研磨棒研磨;再放置於
l alcohol 仿/異
: : 混勻,於離心機中以 離心 後,
取 μl 上清液入新的微量離心管中,加入等量的 混勻,
, 倍的 異丙醇 沉
澱 ,置入 ℃冰箱下 ,再以 離心約
1 上清液,加入 50 μl w
下離心約 後,取出上清液,加入 μl %
酒精清洗,再以 離心 取出上清液,而後風
溶,最後放置於 ℃冰箱下保存備用。
聚合酶連鎖反應
ion, PCR)的目標為增 幅核糖體 RNA 之 片段,如圖 所示,以作為定序用材料。
引子部份,依據 等
ITS 引子
(TCCTC
c
g)溫度 54.5 ℃,30 sec 共循環 33 次 合酶連鎖反應所增幅之rDNA 之重複序列片段,及所用之廣泛引
ITS-5)
dication of amplified rDNA sequence by PCR and the universal er pair applied.
圖3 聚
子對(ITS-4、 之示意圖(改繪自曾與黃(2009))。
Fig.3 In prim
組為:ITS4/ITS5
CGCTTATTGATATGC/GGAAGTAAAAGTCGTAACAAGG)。
PCR 反應中每一管各物質含量為:DNA 溶液(約 10ng/μl)
5μl、10X PCR buffer 5μl、10mM dNTP 0.5μl、primer(10μM)1μl、
Taq DNA polymerase(5u/μl)0.25μl、ddH
2
O 37.25μl,總體積為 50μl。而 PCR 反應條件如下:起始變性溫度94℃,5 min
變性(denaturation)溫度 94℃,45 se 煉合(annealin
延展(elongation)溫度 72℃,45 sec 最後延展溫度72℃,7 min 1 個循環 反應完成後,自動設定保持於4℃。
待反應結束後,取部份PCR 產物,以 1%的洋菜膠體(友和貿 易公司,UR-AGA001 , Agarrose PS1200 )進行電泳,確認所增 幅者為目標 DNA 片段,而其餘 PCR 產物則委託基龍米克斯生技 公司代為定序。
3.比對
以 ITS 序列結果至 NCBI 網站內,利用該網站之 BLAST 工 具進行序列比對,確定增幅之序列是否有效;另則可藉由此方式 輔助鑑定及推測出菌種之屬種名。
十、資料分析與處理
,並對於三個試驗區中環境的不同進 培養出之菌種整理並鑑定後,分別將菌種分成
真菌類;而後依照地區以及樹種的不同將其分別整理後,依照發現之次數 除以總採樣點數後
較樹種與
由整年度之菌種出現之順序中 與發展順序。
將調查完後之各項影響因子整理 行初步的比較。將每次調查
5 大類群,分別為擔子菌類、子囊菌類、不完全菌類、細菌類以及未確定
,所算出之百分比加以分析比較。其目的在於探討並比 地區之不同,對於菌種之出現是否菌有相關聯性以及專一性。並
,可試圖看出菌種在堆積時,不同菌種侵入
一、菌種
此次調查研究中,共分離出各類菌種1265 分離株,依照分離出之菌種 類別分為擔子菌類、不完全菌類、未確定真菌類、細菌等五個類群。類群 總分離次數的計算是將所有類群在三個試驗區
培養基以及依樹種邊心材各取
為樣片7776 125 種次,不完全菌類 679 種次,
未確定真菌類340 種次及細菌類 121 種次。分離與鑑定結果歸納於表 1 至 3。
樣本圓盤菌種分離鑑定結果得,擔子菌 菌
屬名者 菌種。在表 的 , 其 中
Tr
下 四 種 菌 種
Phaeoacr
、Phaeoacremonium rubrigenum 疑似台灣新紀錄
種。調查期間,出現在樣木上的大型 菌子實體分別有擔子菌類的木耳 ( Auricularia auricular-judae )、毛木耳 ( Auricularia polytricha )、簇生鬼 傘 ( Coprinus
disseminatus )、裂褶菌 ( Schizophyllum commune )、略薄多
孔菌 ( Polyporus tenuiculus )與平伏菌類等 6 種;子囊菌類的炭皮菌類 ( Biscogniauxia sp.)、炭角菌類 ( Xylaria sp. )等 2 種;子實體發現情形整理 於表4。在表 4 中,可明顯看出三個試驗區,在楓香林堆置的相思樹與樟樹 樣木上沒有發現大型真菌子實體的出現,單就三個試驗區中,以子實體出肆、結果與討論
名錄
、三種樹種、8 次調查、三種 3 點的 2 次重複下發現之次數相加而來,共 點次,其中擔子菌類分離出
6 種、子囊菌類 1 種、不完全 44 種;擔子菌中確定屬名者 3 種,不完全菌中確定種名者 11 種,確定 19 種,不確定名稱者 5 種;其餘 3 種擔子菌及 3 種不完全菌為未知 1- 3 中,可以看到不完全菌類在本研究中出現之菌種數是最高
Botryosphaeria
、Fusarium
、Pencillium
、Phaeoacremonium
、ichoderma 以上這五個屬之菌種為分離次數較高之菌屬。在研究中也發現 Campylocarpon fasciculare
、Pleurostoma ootheca
、emonium inflatipes
真
現之次數來說,楓香林試驗區整體上也較其他兩個試驗區來的低。而就子 實體出現之種類來看,肖楠林試驗區大型真菌子實體出現之種類為三個試 驗區之首。整體而言,在本次調查共確認發現擔子菌12 種、子囊菌 3 種及
不 。
表 Table 1
菌種出現 屬別出現 完全菌種44 種
1 擔子菌、子囊菌菌種名稱與出現次數
The species and times of isolation of basidiomycota, ascomycota from this study.
菌類分類群 屬名 種名
總次數 總次數
擔子菌類
Auricularia* auricula-juade
6 14polytricha
8Coprinus* disseminatus
2 2Schizophyllum* commune
2 2Polyporus* tenuiculus
1 1平伏菌* 2 2
Fomitopsis
sp. 1 1Phellinus
sp.1 22 26sp.2 4
unkonwn sp.1 5 98
sp.2 9
sp.3 84
類別總數 146
子囊菌類
Biscogniauxia*
sp. 3 3Botryosphaeria rhodina
159 159Xylaria*
sp. 1 1類別總數 163
附註:*號表示為調查期間發現之大型真菌子實體。
Table 2 Times of isolation of unidentified isolated fungal colonies.
表2 未確定之真菌種類出現次數
is study.
表 不完全菌菌種名稱與出現次數
Table 3 The species and times of isolation of deuteromycota from th 3
屬名 種名 單一菌種出現次數 單一屬出現總次數
不完全菌類 Arthrinium
sp.
1 1Arthrographis
sp.1
1 3sp.2
2Aspergillus
sp.
2 2Campylocarpon fasciculare 2 2
Cladsporium
sp.
1 1Cuvularia
sp.
1 1Cylindrocarpon
sp.
1 1Eutypella
sp.
1 1Fusarium oxysporum 19 60
solani 37
sp.1
2sp.2
1sp.3
1Glicocladiopsis tenuis 1 1
Penicillium radicum 7 168
sp.1
70sp.2
91Pestalotiopsis
sp.
1 1Pleurostoma ootheca 1 1
Phaeoacremonium inflatipes 6 60
rubrigenum 39
sp.1
1sp.2
14Phialophora
sp.
6 6Rhinocladiella
sp.
37 37Scytalidium
sp.
5 5Trichoderma aureovirid 34 144
harzianum 7
sp.1
9sp.2
2sp.3
2sp.4
13sp.5
1sp.6
3sp.7
73分離出不確定菌種名
Annulohypoxylon
sp.
1 25Hypocreales
sp.
1Nectria pseudotrichi 15
Paecilomyces
sp.
1Polyporus
sp.
4unknown sp.1
1unknown sp.2
1unknown sp.3
1類別總數 520
e
a
(一)標本園
表 4 各試驗區樣木上大型真菌子實體出現之描述
Table 4 Description on the marcofungal fruiting bodies discovered on the sample logs in the three piling spots.
調查次序
相思樹 樟樹 白匏子0
NA NA NA
1
NA NA NA
2
NA NA NA
3
樹皮開裂,黑線紋* NA 黑線紋*
4
黑線紋*,裂褶菌 NA 黑線紋*,毛木耳
5
黑線紋* 黑線紋* 黑線紋*、毛木耳
6
黑線紋* 黑線紋* 黑線紋*,毛木耳,炭角菌
7
黑線紋* 黑線紋* 黑線紋*
8
裂褶菌,黑線紋*(邊材) 炭皮菌,黑線紋* (邊材) 炭皮菌,黑線紋*,毛木耳
(二)肖楠林
調查次序
相思樹 樟樹 白匏子0
NA NA NA
1
NA NA NA
2
NA NA NA
3
黑線紋* NA 黑線紋*
4
木耳 NA 黑線紋*,木耳
5
黑線紋* 黑線紋* 黑線紋*,木耳
6
黑線紋* 黑線紋* 黑線紋*,毛木耳
7
黑線紋*(邊材) 黑線紋* 黑線紋*,木耳,毛木耳,略薄多孔菌,炭皮菌
8
毛木耳,黑線紋*(邊材) 黑線紋*(邊材),平伏菌 黑線紋*,平伏菌,毛木耳,簇生鬼傘
(三)楓香林
調查次序
相思樹 樟樹 白匏子0 NA NA NA
1 NA NA NA
2 NA NA NA
3 NA NA NA
4 NA NA 木耳
5 NA NA 黑線紋*,木耳,簇生鬼傘
6 NA NA 黑線紋*,毛木耳
7 NA NA 黑線紋*
8 NA NA 簇生鬼傘
附註:*號出現表示白腐菌出現之跡象。
顧中陳昇明、汪碧涵等人之研究已簡單說明,試驗區環境的 差異,對於菌種出現之頻率與組成狀況具有明顯之影響,所以依據此項觀 點,本研究過程中對於實驗區中之環境變化做出整理,其整理如下
(一)環境溫度與溼度
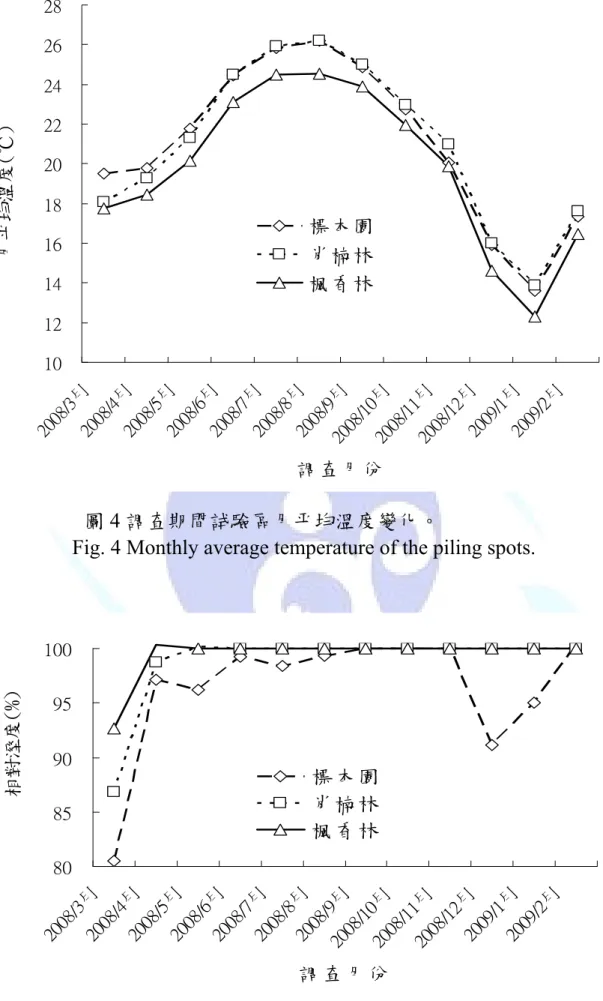
環境溫溼度是利用溫溼度測定儀紀錄調查整年度中,試驗區每月 環境溫度與相對濕度之變化。目的在探討環境的溫溼變化,對於菌種 之種類以及出現數量是否具有影響性。實驗整年度期間環境溫度與相 對溼度整理於圖4 與 5,在下圖 4 中可見楓香林試驗區之溫度明顯低 於標本園與肖楠林試驗區約2℃;而標本園與肖楠林兩個試驗區之溫 度變化則較為相近。在相對濕度的變化上則為標本園試驗區相對濕度 明顯低於肖楠林與楓香林試驗區,如圖5。楓香林試驗區,在整年度 調查中在環境溫度上是三個試驗樣區中最低的,但是在相對濕度中卻 和肖楠林樣區是幾乎整年度均處於相對濕度在100%之情況下;反之 標本園雖然在相對濕度之全年度之變化相較於其他兩試驗區來的低 一些,但在溫度上卻較楓香林試驗區來的高。而綜觀以上結果,實際 探討三個試驗區環境可發現,在標本園中,因為是本校之樹木標本 園,除草與疏伐作業較頻繁,所以環境中之相對濕度會較低;在楓香 林中,則是處於所有試驗區中海拔較高處,所以有較低的環境溫度。
以上結果因子在本研究探討中,則可在後續討論中與菌種出現之數量 與種類相互搭配探討
二、實驗區環境因子 在文獻回
10 12 14 1 26
20 22 24
度(℃)
18 6
20 08 / 20 08 /
20 08 / 20 08 /
20 08 / 20 08 /8
20 08 / 00 8/ 10
008 /11
00 8/ 12 20 09 /
20 09 /
調查月份28
3月 4月 5月 6月 7月 月 9月
2 月
2 月
2
月 1月 2月
月平均溫
標本園 肖楠林 楓香林
Fig. 4 Monthly average temperature of the piling spots.
圖 4 調查期間試驗區月平均溫度變化。
80 85 90 95
相溼%)
100
200 8/ 3月
2008 /4 月
200 8/ 5月
2008 /6 月
20 08 /7 月 2008
/8 月 2008
/9 月 20 08 /10 月
20 08 /11 月 20 08 /12 月
2009 /1 月
20 09 /2 月
調查月份對度(
標本園 肖楠林 楓香林
圖5 調查期間試驗區月平均相對濕度變化。
Fig. 5 Monthly average relative humidity of the piling spots.
(二)光度
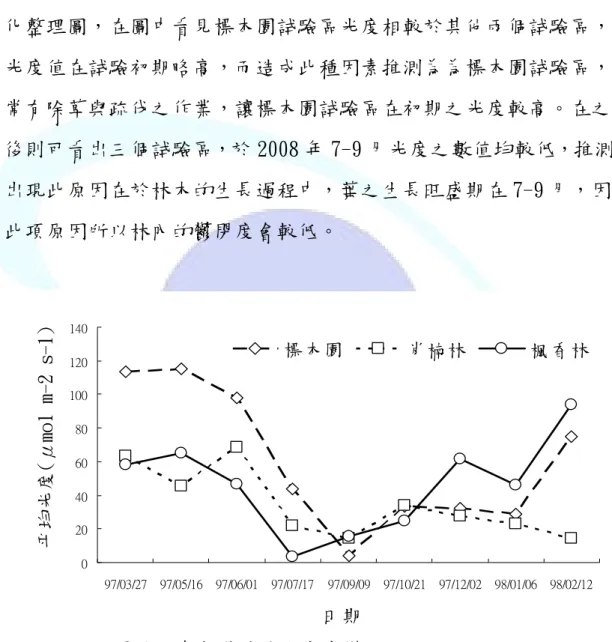
光度的變化,對於大型真菌子實體的出現會有很重要的影響,
因為子實體的出現關係於光的影響,在陳昇明(2001)的研究中就有 提出過類似之論點。本研究是利用光度計量儀在調查期間測量各試 驗區內的光度值。下圖 6 中,為三個試驗區中光度在調查期間的變 化整理圖,在圖中看見標本園試驗區光度相較於其他兩個試驗區,
光度值在試驗初期略高,而造成此種因素推測為為標本園試驗區,
常有除草與疏伐之作業,讓標本園試驗區在初期之光度較高。在之 後則可看出三個試驗區,於 2008 年 7-9 月光度之數值均較低,推測 出現此原因在於林木的生長過程中,葉之生長旺盛期在 7-9 月,因 此項原因所以林內的鬱閉度會較低。
0 20 40 60 80 100 120 140
97/03/27 97/05/16 97/06/01 97/07/17 97/09/09 97/10/21 97/12/02 98/01/06 98/02/12
日期
平均光度(μmol m-2 s-1)
標本園 肖楠林 楓香林
圖 6 調查期間試驗區光度變化。
Fig. 6 Luminosity of piling spots during study period.
冠鬱閉度 林
種 觀 圖 明 在 可 到
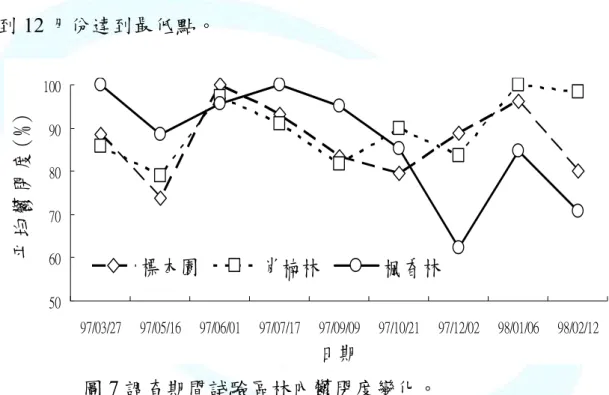
(三)林內樹
內鬱閉度測定的目的在輔助了解林內光度值,鬱閉度變異對菌 子實體的出現亦具有影響性,本研究利用林內鬱閉度計在調查期間 測林內鬱閉度的變化,調查期間的鬱閉度值整理出於下圖7 中。在 中,可見標本園與肖楠林試驗區兩者在調查期間之變化與差異並不 顯,但楓香林試驗區其林內樹冠鬱閉度則有較大之變化。探究原因 於楓香林試驗區中,楓香此樹種為多年生落葉型植物,所以在圖中 見楓香林試驗區,林內樹冠鬱閉度於10 月份開始往下逐漸減低,
12 月份達到最低點。
50 60 70 80 90 100
97/03/27 97/05/16 97/06/01 97/07/17 97/09/09 97/10/21 97/12/02 98/01/06 98/02/12
日期
平均鬱閉度(%)
標本園 肖楠林 楓香林
圖 7 調查期間試驗區林內鬱閉度變化。
Fig. 7 Canopy coverage of piling spots during study period.
(四)土壤溫度與溼度
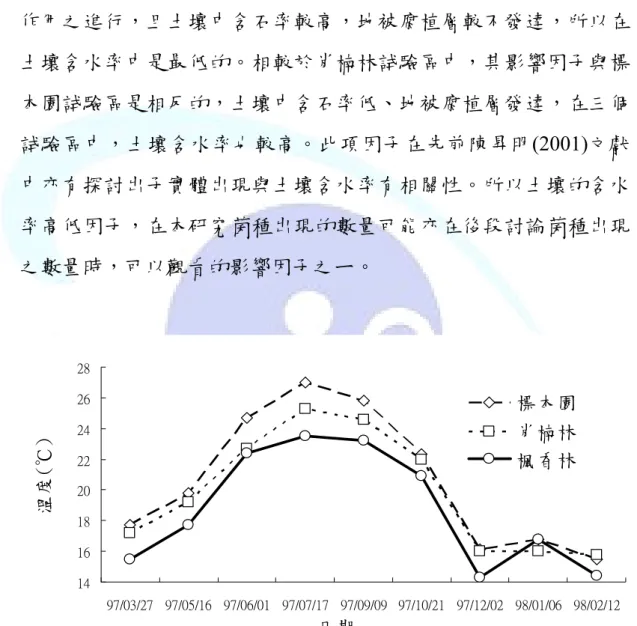
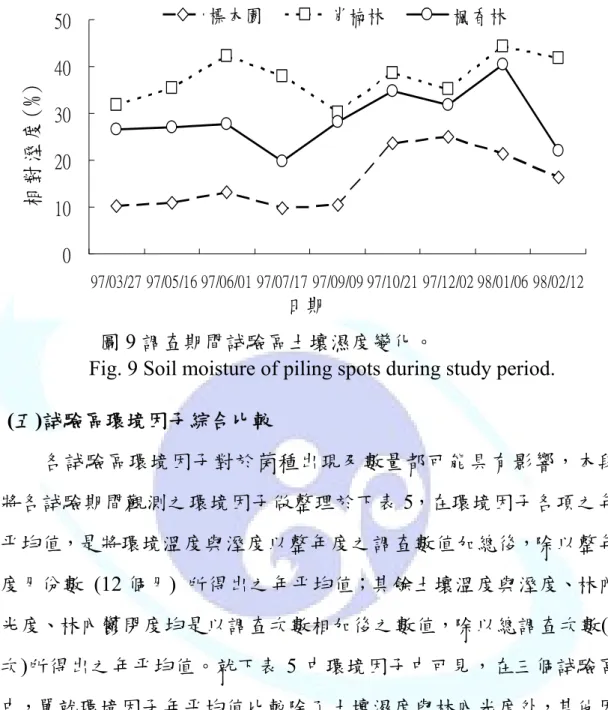
於先前文獻回顧中,提及土壤的溫度與溼度對於菌種出現具有 明顯的影響,本試驗是利用土壤測定儀於每次調查期間,在於每一 試驗區同一位置下進行測量,重複兩次,以兩次測值之平均值則為 當次測值,所以在此探討三個試驗區,其土壤溫度與溼度之差異性 (圖 8 與 9)。在圖 8 中可見土壤溫度的變化和季節環境溫度之變化相 符合的。在試驗區中,標本園試驗區土壤溫度較高;而楓香林試驗
造成三試驗區環境溼度之原因是一致的,與試 驗區
較高。此項因子在先前陳昇明(2001)文獻 中亦有探討出子實體出現與土壤含水率有相關性。所以土壤的含水 率高低因子,在本研究菌種出現的數量可能亦在後段討論菌種出現 之數量時,可以觀看的影響因子之ㄧ。
區則較低。其原因和
本身條件因子,如海拔有關,而環境的溫度與土壤的溫度是具 有連帶關係的。圖 9 中探討土壤濕度變化,其變化原因除與環境溼 度有關係外,另與各試驗區中的地被腐植質及土壤結構有相關性,
在三個試驗區中由圖 9 可見,標本園的環境因為常有除草與疏伐之 作用之進行,且土壤中含石率較高,地被腐植層較不發達,所以在 土壤含水率中是最低的。相較於肖楠林試驗區中,其影響因子與標 本園試驗區是相反的,土壤中含石率低、地被腐植層發達,在三個 試驗區中,土壤含水率也
14
97/03/27 97/05/
16 22 24 26 28
16 97/06/01 97/07/17 97/09/09 97/10/21 97/12/02 98/01/06 98/02/12
標本園 肖楠林
溫度(℃)
18 20
日期
楓香林
圖 8 調查期間試驗區土壤溫度變化。
Fig. 8 Soil temperature of piling spots during study period.
0 10 20 30
97/03/27 97/05/16 97/06/01 97/07/17 97/09/09 97/10/21 97/12/02 98/01/06 98/02/12
日期
相對溼度(%) 40
50 標本園 肖楠林 楓香林
圖 9 調查期間試驗區土壤濕度變化。
Fig. 9 Soil moisture of piling spots during study period.
(五)試驗區環境因子綜合比較
各試驗區環境因子對於菌種出現及數量都可能具有影響,本段 將各試驗期間觀測之環境因子做整理於下表5,在環境因子各項之年 平均值,是將環境溫度與溼度以整年度之調查數值加總後,除以整年 度月份數 (12 個月) 所得出之年平均值;其餘土壤溫度與溼度、林內 光度、林內鬱閉度均是以調查次數相加後之數值,除以總調查次數(8 次)所得出之年平均值。就下表 5 中環境因子中可見,在三個試驗區 中,單就環境因子年平均值比較除了土壤濕度與林內光度外,其他因 子在試驗區中比較上均無明顯之差異性。因此,推測之後影響菌種出 現與數量可能之原因在於以上兩個因子之影響。此兩項因子在本研究 後段菌種出現及數量的變化上亦可將其與之探討。
表5 各項環境因子之年平均值
Table 5 Yearly average value of individual environment factor.
測量因子 標本園 肖楠林 楓香林
土壤溫度(℃) 23.2 22.4 21.1
鬱閉度(%) 98.1 100 98.3
土壤濕度(%) 17.6 42.2 32.3
林內光度
(μmol m
-2s
-1)
82.5 39.1 51.9環境溫度(℃) 21.01 20.98 19.80
環境溼度(%) 96.9 100 100
(六) 調查期間樣木 pH 值之變化
不同的樹種在本身就有不同的差異,不論是組織、硬度或是木 材內部中各種物質的種類與含量多寡都會有所不同,在調查期間樹 種的各項因子亦會因外在因素之影響(如環境溫溼度、昆蟲之侵 害),使得樹種本身內部因子,如 pH 值會有所改變。在樣木上因為 在菌種侵入後,會讓樹種內部的各項因子略為改變,而所有因子中,
在樣木上可很明顯看出的不同的在於樣木中的含水率以及pH 值,測 定pH 值的目的主因,真菌在生長的過程中有喜好的 pH 值,所以在 此可藉由測量 pH 值,試圖找出可能影響菌種出現與數量多寡之原 因。本試驗中在樣木擺置於三個試驗區期間,依照其樹種及區域性 的不同,將所有調查時每次樣木之pH 值,依照邊材與心材的不同分 別測定,主要為探討不同樹種、試驗區下,pH 值的差異對菌種之出 現可能的影響。試驗期間不同試驗區各樹種pH 值整理於表 6。
表 調查期間各樣木平均 pH 值
sample logs during study period.
6
Table 6 The average pH value of
在試驗中發現,不同樹種的pH 值是有差異的,而在上表 6 中,
以樣木平均 pH 值來比較三種樹中之差異性,樟樹的 pH 值在三種試 驗樹種中是最高的,而在這裡可以看到在三個試驗區中,在同種樣木 的 pH 值差異不大,但是樹種間卻是有差異,所以可以推測 pH 值的 高低對於菌種的出現可能具有影響。而在本研究後段研究菌種出現與 數量上可以將本因子放入其中做探討。
(七) 調查期間樣木含水率之變化
在調查期間,樣木含水率的變化反應環境中溫度與溼度的變化,
以及影響菌種可能之出現數量,本研究在調查期間將試驗樣木的含水 率,分邊材與心材參照 CNS 之方法測定樣木含水率。測定結果依照 試驗區以及試驗樣木的不同整理於圖10 至 12。於圖 10 至 12 可發現,
在調查期間白匏子樣木的含水率是三種樹種間最高的,最低的為相思 樹樣木。推測原因在於樹種本身質地密度的不同、試驗區的土壤濕度 與環境溼度之不同,可能造成樣木的吸濕性會有所差異性,所以含水 率上才會有所不同。
0 50 100 150 200
標本園 肖楠林 楓香林
97/05/16 97/06/01 97/07/17 97/09/09 97/10/21 97/12/02 98/01/06 98/02/12
日期
含水率(%)
(A)
0 200
標本園 肖楠林
100 150
含水率(%)
50
97/05/16 97/06/01 97/07/17 97/09/09 97/10/21 97/12/02 98/01/06 98/02/12
日期
楓香林
(B)
圖10 調查期間各試驗區中相思樹樣木含水率變化。(A)邊材;(B)心材。
Fig. 10 Moisture contents of Taiwan Acacia sample logs during study period.
(A)sapwood, (B)heartwood.
0 50 100 150 200
97/05/16 97/06/01 97/07/17 97/09/09 97/10/21 97/12/02 98/01/06 98/02/12
日期
含水率(%)
標本園 肖楠林 楓香林
(A)
0 50 100 150 200
97/05/16 97/06/01 97/07/17 97/09/09 97/10/21 97/12/02 98/01/06 98/02/12
日期
含水率(%)
標本園 肖楠林 楓香林
(B)
圖11 調查期間各試驗區中樟樹樣木含水率變化。(A)邊材;(B)心材。
Fig. 11 Moisture contents of Comphor Tree sample logs during study period.
(A)sapwood, (B)heartwood.
0 50 100 150 200
97/05/16 97/06/01 97/07/17 97/09/09 97/10/21 97/12/02 98/01/06 98/02/12
日期
含水率(%)
標本園 肖楠林 楓香林
(A)
0 50 100 150 200
97/05/16 97/06/01 97/07/17 97/09/09 97/10/21 97/12/02 98/01/06 98/02/12
日期
含水率(%)
標本園 肖楠林 楓香林
(B)
圖12 調查期間各試驗區中白匏子樣木含水率變化。(A)邊材;(B)心材。
Fig. 12 Moisture contents of Turn-in-the-wind sample logs during study period. (A)sapwood, (B)heartwood.