國立臺灣大學醫學院分子醫學所 碩士論文
Institute of Molecular Medicine College of Medicine
National Taiwan University Master Thesis
玻尿酸藉由結合CD44活化中性白血球的分子機轉研究 Studies on the Molecular Basis of CD44 and
Hyaluronan in Neutrophils Activation
林佳慧 Chia-Hui Lin
指導教授:余家利 博士 Advisor: Chia-Li Yu, M.D., Ph.D.
中華民國 102 年 7 月 July, 2013
中文摘要
CD44 是一個具有多重功能且位於細胞表面的醣蛋白質,其功能涉及細胞之間的相 互作用、細胞附著、遷移、增殖和血管新生,而且在大多數的哺乳動物細胞中皆 有表現。玻尿酸(hyaluromic acid; HA)是 CD44 一個非常重要的配體,廣泛分佈 在整個結締組織、上皮細胞或神經組織中。CD44 與玻尿酸結合會活化其下游訊息
傳遞路徑而引發許多作用。在之前的研究中已經證明CD44 具有誘導淋巴細胞活
化、再循環和吸引淋巴細胞的功能。一些研究報告更指出CD44 和玻尿酸的結合可
以調控發炎反應、拮抗組織損傷和修復由發炎所造成的細胞傷害、減少發炎因子
的釋放和增加幹細胞遷移。然而CD44 與玻尿酸結合後對於中性白血球的影響目前
仍不清楚。在我們初步的研究中,發現了一些有趣的結果。首先,脂多醣(LPS)
和anti-CD44 會誘導多核型白血球 (PMN)中 CD44 的表現量增加。而玻尿酸的刺
激可以增加多核型白血球的吞噬作用並誘導細胞骨架的重組。CD44 與玻尿酸的結
合能誘導IL-8 的產生。接下來我們更發現,玻尿酸可以誘導 MAP kinase (p38)、
ERK1/2 磷酸化增加。在未來的研究方面,我們將添加不同的促發炎因子,觀察多
核型白血球CD44 的表現量。並使用抑製劑阻斷訊息傳遞路徑研究其對多核型白血
球的吞噬作用和cytokine 產生的影響以及所扮演的角色。此外,我們也將使用玻
尿酸酶分解玻尿酸,以研究CD44-HA 的結合對於多核型白血球的影響。最後,我
們藉由降低CD44 的表現量確認玻尿酸與 CD44 結合對於發炎反應的調解具有重要
性。
關鍵字:CD44、玻尿酸、MAP kinase (p38)、ERK1/2、多核型白血球吞噬作用、
細胞骨架的重組
Abstract
CD44 is a multifunctional cell-surface glycoprotein that is involved in cell-cell
interaction, cell adhesion, migration, proliferation and angiogenesis and is widely
expressed in a large number of mammalian cell types. Hyaluronic acid (HA), one of
important ligand of CD44, is distributed widely throughout connective, epithelial, and
neural tissues. After CD44-HA binding initiates CD44 downstream signaling pathways,
the signaling induces multiple functions. Previous studies have demonstrated that CD44
induces the lymphocyte activation, recirculation, and homing. Some paper have shown
that CD44-HA interaction could regulate inflammation, tissue injury and repair through
regulating inflammatory cell recruitment, release of inflammatory cytokines, and stem
cell migration. However, the effects of CD44-HA interaction on neutrophils remained
unclear. In our preliminary study, several interesting results were found. First, we found
that the LPS and anti-CD44 could increase the expression of CD44 on PMN. HA could
also enhance the phagocytosis activity and cytoskeleton rearrangement of PMN and its
binding to CD44 could induce the interlukin-8 (IL-8) production. Furthermore, we
found that HA could induce phosphorylation of MAP kinase (p38), and ERK1/2
signaling pathways. In the future, we will test the effect of different pro-inflammatory
cytokines on the surface expression of CD44 on PMN. The signaling pathways will be
selectively blocked for the observation of their effects on phagocytosis and cytokines
production of PMN. In addition, the hyaluronidase will be used to evaluate the
CD44-HA interaction on PMN. Lastly, the knockdown of CD44 expression in PMN
will be carried out to define the CD44-independent mechanisms by which HA can
mediate inflammation.
keywords:CD44、hyaluronic acid、MAP kinase (p38)、ERK1/2、PMN phagocytosis、
cytoskeleton rearrangement
目 錄
口試委員會審定書………... i
中文摘要 ... ii
Abstract ... iii
Chapter 1. Introduction ... 4
Chapter 2. Materials and methods ... 7
2.1 Isolation of PMN and MNC from normal human peripheral blood 7 2.2 Cell culture ... 7
2.3 Detection of CD44 expression treated with inflammatory molecule by flow cytometry ... 8
2.4 Cytokine measurements ... 9
2.5 Detection of PMN phagocytosis-enhancing activity of HA by flow
cytometry ... 9
2.6 Preparation of whole cell extraction... 10
2.7 Western blotting ... 10
2.8 Immunofluorescence microscopic observation of cytoskeleton change in activated-PMN ... 11
2.9 Statistical analysis ... 12
Chapter 3. Results ... 13
3.1 The expression of CD44 on MNC and PMN ... 13
3.2 HA induced IL-8 production ... 14
3.3 HA enhanced PMN phagocytosis activity ... 14
3.4 HA induces cytoskeleton rearrangement on PMN ... 15
3.5 HA induced phosphorylation of MAPK and ERK1/2 on PMN and MNC ... 16
3.6 HA abolished induced IL-8 production on differiated-HL60 ... 16
Chapter 4. Discussion ... 18
Chapter 5. Conclusions and Future directions ... 21
Chapter 6. Figures ... 22
Chapter 7. References ... 38
Chapter 1. Introduction
CD44 is a multifunctional cell-surface glycoprotein and widely expressed in a large
number of mammalian cell types. The most abundant standard isoform of human CD44
protein (CD44s) contains 363 amino acids and its molecular weight is approximately 37
kDa. The structure of CD44s consists of three regions, a C-terminal cytoplasmic domain,
to which numerous signaling molecules bind directly or indirectly on activation of the
ligand binding, a transmembrane domain, and an extracellular domain which is the
so-called link modules of hyaluronan (HA) binding proteins (1, 2) (Figure 1A). The
variant isoforms of CD44 (CD44v) inserts an alternatively spliced exons within the
extracellular domain, a membrane proximal domain, which can alter the binding affinity
for HA and confer interaction with alternative ligands. The CD44 isoform go through
the post-translational modification with a molecular weight of about 80 kDa (2). The
CD44s is widely expressed in a large nimber of mammalian cell types, and the
expression of CD44v is detectable in hematopoietic cells (3), particularly in MNCs (4)
and in reactive lymph node cells (3, 5). The principal and important ligand of CD44 is
hyaluronic acid (HA), a ubiquitous component of the extracellular matrix (ECM). But it
can also interact with other ligands, such as osteopontin (6), collagens (7, 8), and
fibronectin (9). HA is a linear, polymeric glycosaminoglycan composed of repeating
disaccharides D-glucuronic acid and N-acetyl-D-glucosamine linked by a glucuronidic β
(1→3) bond (10, 11) (Figure 1B). HA is widely distributed throughout connective,
epithelial, and neural tissues.
CD44-HA interactions mediate cell adhesion and migration in a variety of physiological
and pathophysiological processes, including tumour metastasis, wound healing and
leukocyte extravasation at sides of inflammation (11, 12, 13, 14). Previous studies have
demonstrated that CD44 induces the lymphocyte activation, recirculation, and homing
(15, 16). According to Paul W. et al. have indicated that the role of CD44 in regulating
HA interactions depends on the cell types, and the effect and mechanism of CD44-HA
interaction on macrophage have reported (11, 17). However, the effects and mechanism
of CD44 binding on neutrophils remain unclear.
CD45, as leukocyte common antigen (LCA), is glycoproteins uniquely expressed on the
surface of all leukocytes and their hemopoietic progenitor cells. CD44 is also a one of
common LCA on neutrophils. CD45 is a family of high molecular weight
transmembrane protein tyrosine phosphatase (PTPase) expressed on all nucleated
haematopoietic cells (18). Many authors have reported the possible roles of CD45 and
its isoforms in T and B cell differentiation (19), natural killer T cells and cytotoxic T
lymphocyte functions (20), cytokine production by MNC (21) and TCR-associated
signalling in T cells (22).
Previous studied have shown that the lower CD44 expression in immunodeficiency
animal model or disease patient. However, the expression of CD44 is higher in
inflammation condition (1, 2). CD44 has a vital role in involving in immune response.
So we want to know the role and mechanism of CD44-HA interaction on PMN. To
figure out the relationship of CD44 and CD45, and the effects and mechanism of CD44
binding to HA on neutrophils. In the present study, we found that anti-CD44 could
increase the expression of CD44 on PMN. CD44-HA interaction could induce IL-8
production through MAP kinase (p38), and ERK1/2 signaling pathways. In addition, it
could enhance phagocytosis and cytoskeleton rearrangement of the PMN.
Chapter 2. Materials and methods
2.1 Isolation of PMN and MNC from normal human peripheral blood
Heparinized venous blood obtained from normal individuals was mixed with
one-quarter volume of 2% dextran solution in 37℃ (molecular wight: 425000-575000)
and incubated at room temperature for 30 minutes. The cell suspension was gently
layered over Ficoll-Hypaque density gradient solution (specific gravity 1.077; GE
Healthcare, Waukesha, Wisconsin, USA) and centrifuged at 500 x g for 30 minutes. The
MNC were aspirated from the interphase whereas the PMN were collected from the
bottom. The residual RBC in PMN was lysed in cold 0.85% ammonium chloride
solution. These cells were then rinsed twice with PBS and re-suspended in RPMI 1640
(Gibco/BRL, Grand Island, New York, USA) supplemented with 10% (v/v) fetal bovine
serum (FBS), 2 mM L-glutamine (hereafter referred to as complete medium). The
viability of PMN and MNC were detected by trypan blue dye and the cell concentration
was adjusted to 2×106/ml in complete medium.
2.2 Cell culture
Human promyelocytic leukemia cell line (HL-60) was maintained in complete medium
at 37℃ in a humidified atmosphere containing 5% CO2. HL-60, MNC and PMN (2×106
cells/ml) were treated with LPS (20ng/ml), h-IgG (1μg/ml), anti-CD44 (1μg/ml),
anti-CD45 (1μg/ml), anti-CD3 (1μg/ml)/ anti-CD28 (1μg/ml), and hyaluronic acid (HA
1mg/ml and 2mg/ml) at 37℃, then cultured and harvested at the indicated time points.
Induction of differentiation was obtained by seeding the cells at a concentration of 5 x
105/ml in the presence of DMSO (Microbiological Associates, Rockville, Md.) at a final
concentration of 1.3% (v/v) for 5 days. After exposure, the cells were resuspended in
DMSO-free medium. Next, differentiated-HL60 were treated with LPS (20ng/ml),
anti-CD44 (1μg/ml), and hyaluronic acid (HA 1mg/ml and 2mg/ml) at 37℃, then
cultured and harvested at the indicated time points.
2.3 Detection of CD44 expression treated with inflammatory molecule by flow
cytometry
MNC and PMN were treated with anti-CD3 (1μg/ml)/ anti-CD28 (1μg/ml) or LPS
(20ng/ml) at 37℃, before being cultured and harvested at the indicated time points. The
cells were washed twice with PBS. The cells were then fixed with 4%
paraformaldehyde for 30 minutes at room temperature and incubated with
anti-CD44-FITC overnight to detect the CD44 expression. The percentage (%) and
mean fiuorescence intensity (MFI, denoted by mean channel number) of CD44
expression were determined by FACSort flow cytometry (Becton Dickinson) at wave
length 488nm excitation.
2.4 Cytokine measurements
After treatment with HA and pro-inflammatory molecule at indicated time points, the
cells were centrifuged at 800 x g for 10 minutes for the detectection of cytokine
concentrations in cell culture supernatant. The concentrations of cytokines including
IL-8 were quantified using their respective ELISA kits (R&D Systems, Minneapolis,
Minnesota, USA).
2.5 Detection of PMN phagocytosis-enhancing activity of HA by flow cytometry
Fluoresbrit carboxylate microspheres (0.75 μm in diameter, Polyscience Inc.) were
washed with PBS in advance twice and opsonized by incubation with fresh human
serum at 37℃ for 2 hours. Fresh prepared PMN (2×106 cells/ml) were treated with LPS
(20ng/ml), h-IgG (1μg/ml), anti-CD44 (1μg/ml), anti-CD45 (1μg/ml), and HA (1mg/ml
and 2mg/ml) at 37℃ for 1 hours. The mixture was let reacting with opsonized beads
(1×108 beads/ml) at 37℃ for 1 hour in 5% CO2-95% air. After incubation and wash by
PBS twice, the percentage (%) and mean fluorescence intensity (MFI, denoted by mean
channel number) of PMN phagocytosis were determined by FACSort flow cytometry
(Becton Dickinson) at wave length of 488nm excitation.
2.6 Preparation of whole cell extraction
After treatment with HA or control medium at different time points at 37℃, the cells
were centrifuged at 800 x g for 10 minutes followed by wash with cold PBS. After
centrifugation at 800 x g for 10 minutes, the pelleted cells were lysed with cold RIPA
buffer (25 mM Tris-HCl pH 7.6, 150 mM NaCl, 1%NP-40, 1% sodium deoxycholate,
0.1% SDS ) containing protease inhibitor cocktail and phosphatase inhibitor cocktail
(Roche) and kept on ice for 30 minutes. The cell lysates were centrifuged at 10,000 x g
at 4℃ for 20 minutes to remove the debris and the supernatants were applied for
Western blot. The protein concentration of the cell extraction was measured using the
BCA Protein Assay (Pierce).
2.7 Western blotting
Proteins were separated by 10% SDS-PAGE and transfer to polyvinylidene fluoride
(PVDF) membrane (Millipore Inc.) in a Mini Trans-Blot cell (Bio-Rad) for 2 hours at
350 mA. The PVDF membranes were blocked with Tris-buffered saline and Tween 20
(TBST, 50 mM Tris, 150 mM NaCl, 0.05% Tween 20, pH 7.6) containing 1% BSA at
room temperature for 30 minutes, and probed with a specific antibody overnight at 4℃.
After washing the membrane three times with TBST buffer each for 5 minutes, the
complexes were detected by HRP-conjucated-secondary antibody (Jackson ImmunoLab)
and ECL Western Blotting Substrate (Pierce) chemifluorescence detecting system.
2.8 Immunofluorescence microscopic observation of cytoskeleton change in
activated-PMN
After stimulation at 37℃ by HA or LPS for 2 hours, the activated-PMN were wash with
PBS and then fixed for 30 minutes at room temperature with a solution of 4%
paraformaldehyde followed by washing with PBS. Then mixture was centrifuged and
the supernatant was discarded. The cells were resuspended in 200 μL of PBS. 5 to 10
μL of the cell suspension was smeared above a gelatin-coated slide. Placed the slide on
a hot plate (low heat setting), and allowed the liquid to evaporate. The PMN were
permeabilized with 0.1% Triton X-100 for 15 minutes at room temperature, and washed
three times with PBS. The blocking solution (10% FBS in PBS) was added to inhibit the
non-specific binding. PMNs were stained with fluorescent phalloidin (1:200) for
visualization of actin filament at room temperature for 30 to 60 minutes. The slides
were washed three times with PBS and stained with DAPI
(4’,6-diamidino-2-phenylindole, 1:1000) at room temperature for 5 minutes. After three
times washes with PBS, the slides were mounted with glycerol and gently covered the
cover slip avoiding producing the air bubbles. The cells were visualized the
cytoskeleton was observed using a fluorescence microscope (Olympus, Japan).
2.9 Statistical analysis
The statistical analyses were performed using the analysis of variance (ANOVA).
Statistical significance was defined as p<0.05.
Chapter 3. Results
3.1 The expression of CD44 on MNC and PMN
Previous investigations have shown that during inflammation, CD44 expression is
upregulated on hematopoietic and parenchymal cells (23, 24), and it plays a crucial role
in an variety of inflammatory diseases including murine models of inflammatory bowel
disease, collagen- and proteoglycan-induced arthritis, cutaneous inflammation,
experimental autoimmune encephalomyelitis, and IL-2-induced vascular leak syndrome
(25, 26, 27, 28). To understand whether the inflammatory molecules could enhance the
CD44 expression on MNC and PMN, we used pro-inflammatory molecules anti-CD3/
anti-CD28 on MNC. The results indicated that co-stimulated anti-CD3/ anti-CD28 as
well as LPS provided a strong pro-inflammatory signal on PMN (29, 30). As
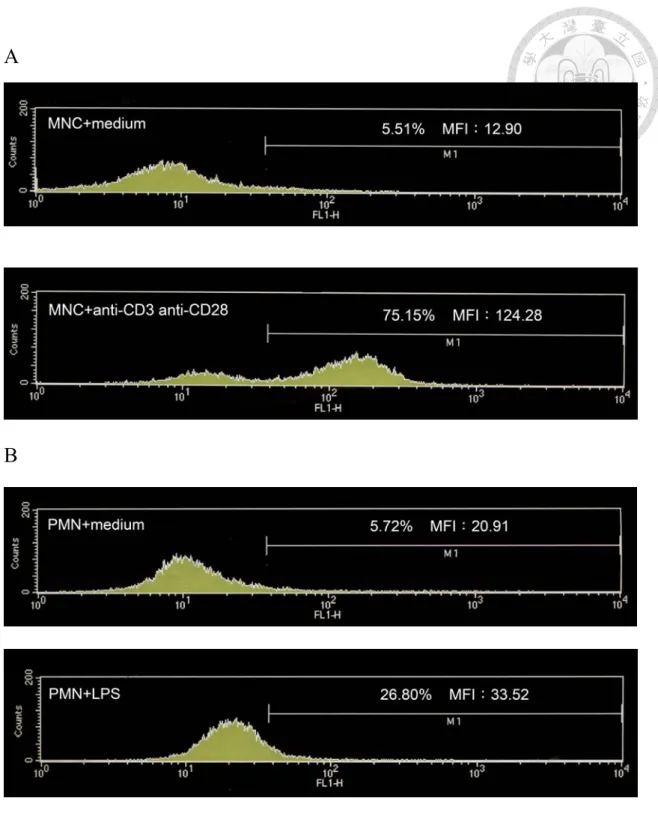
determined and demonstrated by flow cytometry data, anti-CD3/ anti-CD28 had
significantly enhanced the percentage of CD44 expression on MNC compared to the
control (Figure 2A). On the other hand, LPS also slightly increased the CD44
expression on PMN (Figure 2B). Accordingly, we concluded that pro-inflammatory
cytokines could increase the CD44 expression on PMN.
3.2 HA induced IL-8 production on PMN
Next, we determined whether CD44-HA interaction could induce the PMN into the
activation condition and enhance inflammation. We used LPS, anti-CD44, anti-CD45
and HA to stimulate the PMN and then detect the interleukin-8 (IL-8) concentration,
which is a one of the neutrophil-specific CXC subfamily of chemokines. IL-8 is also a
potent neutrophil chemotactic and activating factor and is a primary pro-inflammatory
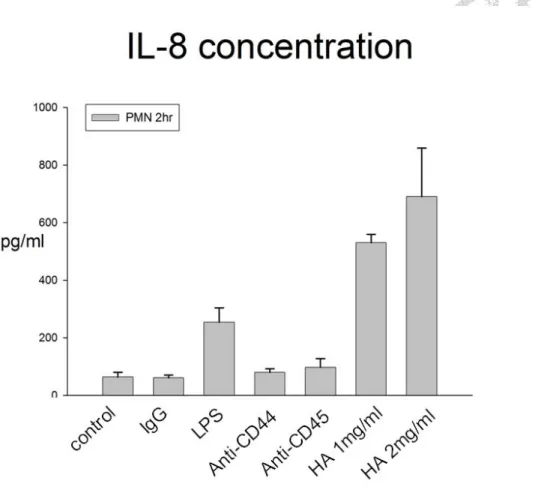
cytokine produced by many cells (31, 32, 33, 34). We could demonstrate that LPS
increases the IL-8 production and HA has significantly increased IL-8 concentration,
compared to the controls. However, anti-CD44 and anti-CD45 might have little
increasing effect on IL-8 production. Therefore, CD44-HA interaction could promote
PMN to go inyo the activated-form.
3.3 HA enhanced PMN phagocytosis activity
Previous studied have shown that the role of CD44 in binding, ingestion (phagocytosis),
and clearance of apoptotic cells (35) as well as microbial pathogens (36). A direct
antibody ligation of CD44 on macrophages enhances the subsequent uptake of apoptotic
cells (37, 38). To further confirm the CD44-HA interaction could enhance the PMN
phagocytosis, we used HA to stimulate PMN and Fluoresbrit carboxylate microspheres
to determine the PMN phagocytosis by flow cytometry. LPS could enhance the PMN
phagocytosis compared to the controls (49.23% to 75.57%) and so was taken as positive
controls. The CD44-HA interaction significantly increased the phagocytosis activity
(49.23% to 93.01%), to an extent of even greater than LPS stimulation. Anti-CD44
(49.23% to 62.87%) and anti-CD45 (49.23% to 63.57%) also stimulated the PMN
phagocytosis (Figure 4), but this increase was smaller than that exerted by LPS and HA.
These results of anti-CD45 were consistent with previous reports (39). It suggested that
CD44-HA interaction could enhance the PMN phagocytosis activity.
3.4 HA induces cytoskeleton rearrangement on PMN
Previous investigations have indicated that the cytoplasmic domain of CD44 has a
linker protein- ankyrin specific binding site (36, 40). Ankyrins proteins are a family of
adaptor proteins that mediate the attachment of integral membrane proteins to the
spectrin-actin based membrane cytoskeleton (41). To understand if CD44-HA
interaction on PMN could initiate the CD44 downstream signaling pathway and induce
cytoskeleton rearrangement, the cells were treated with HA. After treatment with HA,
we used phalloidin to stain actin filaments and observed the aggregation of actin
filaments. We found that not only HA could induce cytoskeleton rearrangement but also
anti-CD44 and anti-CD45 could partially induce the actin aggregation (Figure 5). These
results indicated that CD44 on PMN could mediate the cytoskeleton change and
CD44-HA interaction could enhance cytoskeleton rearrangement to achieve its
biological function.
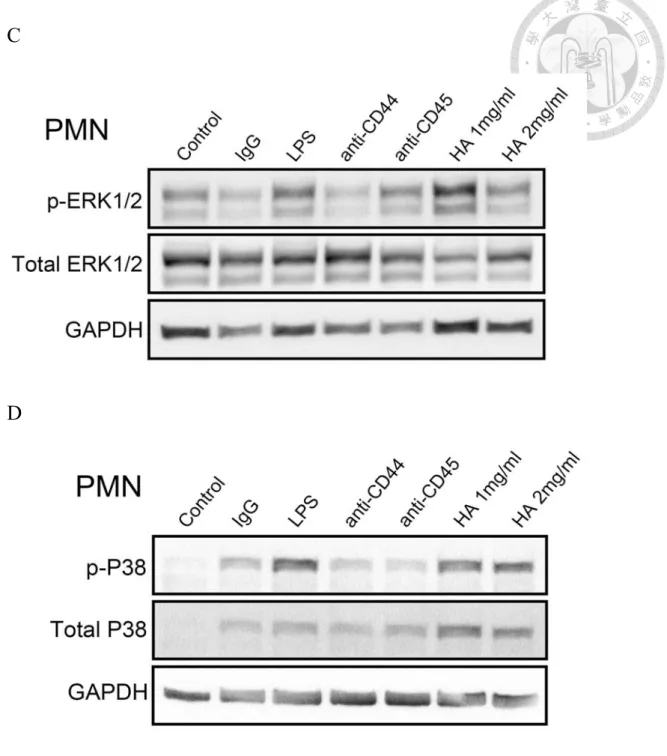
3.5 HA induced phosphorylation of MAPK and ERK1/2 on PMN and MNC
Previous studied have demonstrated that CD44-HA interaction could activate several
signaling pathways (42, 43). To further understanded the molecule mechanism of the
CD44-HA interaction on activated-PMN and activated-MNC, we observed the
phosphorylation of signal transduction. In Fig 6A and 6B, you can see that HA could
increase the phosphorylation of MAPK (P38) and ERK1/2 on MNC. On the other hand,
CD44-HA interaction could also significantly increase the phosphorylation of MAPK
(P38) and ERK1/2 on PMN (Figure 6C and 6D). These results indicated that HA could
induce and activate MAPK (P38) and ERK1/2 signaling pathway.
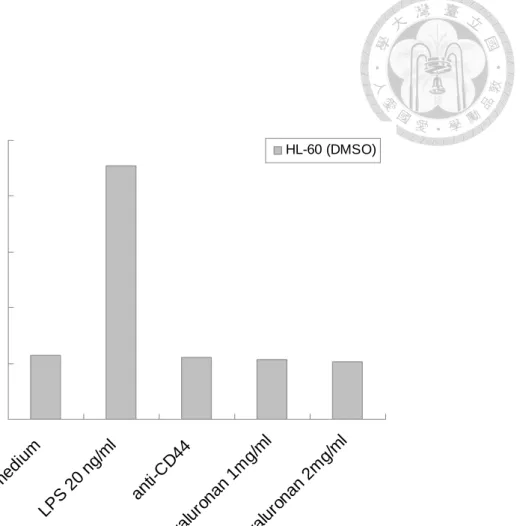
3.6 HA abolished to induce IL-8 production on differiated-HL60 cells
According to Liu J. et al. have shown that down-regulation of CD44 contributes to the
differentiated-HL60 cells (44). To investigate the effect of CD44-HA interaction on the
differentiation of HL-60 cells, we treated with DMSO to induce cells differentiation and
detect IL-8 production after treatment with HA. We found that LPS increased the IL-8
production, however HA and anti-CD44 abolished to induce IL-8 production on
differiated-HL60 cells (Figure 7). Therefore, it indicated that HA is specific binding to
CD44 and induce downstream signaling pathway.
Chapter 4. Discussion
1. The role of hyaluronic acid in tissue and cell
Hyaluronic acid, a ubiquitous component of the extracellular matrix (ECM), is widely
distributed throughout connective, epithelial, and neural tissues. CD44 is expressed in
numerous cell types. Previous papers have shown that CD44 is involved in and
mediated many biological functions. In normal condition, HA is enriched in ECM. How
the cell determines and regulates HA binding to CD44 further prevents HA induced
downstream signaling pathway from activating PMN or MNC is unknown. Otherwise,
many authors have reported that the CD44 molecule plays a central role in the
development of collagen- or proteoglycan (cartilage delivered)-induced arthritis (45, 46,
47, 48, 49). The expression of CD44 and extractable HA is increased in the arthritic
inflamed joint tissues, suggesting that they are associated with the inflammatory process
(50). Hyaluronic acid is primarily used to increase mobility of the joints, clear eye
vision, combat signs of aging such as wrinkles, relief fibromyalgia and help wound heal.
The phenomenon is obviously contrary to our current results showing that HA could
induce tissue inflammation. So, it might have another mechanism underlying the
mediation and regulation of the CD44-HA interaction. There have several possibilities
to affect the CD44-HA interaction: one is hyaluronan synthases (HAS). Previous papers
have indicated that inflammation molecules, TNFα, increases the transcription of HAS
and the expression on cell membrane (51). And another is hyaluronidase, it may be have
some mechanism to regulate it enzyme activity or concentration in cells to increase or
decrease the concentration of HA. There have the papers shown that the HA oligomer
compete for endogenous polymeric HA, thus replacing high affinity, multivalent and
cooperative interactions with low affinity, low valency receptor interactions (52). The
last is the expression of CD44, according to figure 7 that down-regulation of CD44
abolished CD44-HA activation. And these possibilities need further improve and
understand its regulation and detail mechanism.
2. The relationship of CD44 and CD45
CD45, as leukocyte common antigen, is a family of high molecular weight
transmembrane protein tyrosine phosphatase (PTPase) expressed on all nucleated
haematopoietic cells (17). Many authors have reported the possible roles of CD45 and
its isoforms in cell differentiation (18), cytokine production (20) and TCR-associated
signalling (21). Recent studied have shown that CD45, negatively regulatory role for
CD45 in CD44 signaling leading to actin rearrangement and cell spreading in activated
thymocytes and T cells. According to Yu C. L. et al. 2002 and our present data the
CD45 and CD44 had similar function on PMN. The relationship of CD44 and CD45
mediating and regulating PMN remains to be confirmed.
Chapter 5. Conclusions and Future directions
This study showed the effect of CD44-HA interaction on PMN. First, we found that
pro-inflammatory molecule (LPS and anti-CD3/anti-CD28) could increase the
expression of CD44. Next, we showed that HA could activate PMN and increase the
IL-8 production. We also demonstrated CD44-HA interaction not only could enhance
the phagocytosis activity but also induced cytoskeleton reorganization. We observed
that CD44-HA interaction could induce phosphorylation of MAPK and ERK1/2
signaling pathway. Last, we investigated that HA abolished induce the IL-8 production
on the differentiation of HL60 cells induced by DMSO. In the future, we are going to
use signaling transduction inhibitor to block the signaling pathway and to investigate
the effects of phagocytosis as well as cytokines production on PMN. In addition, we
will use the hyaluronidase to investigate the CD44-HA interaction on PMN. Last, we
will knock down the CD44 in PMN expression to define the CD44-independent
mechanisms by which HA can mediate inflammation.
Chapter 6. Figures
A
B
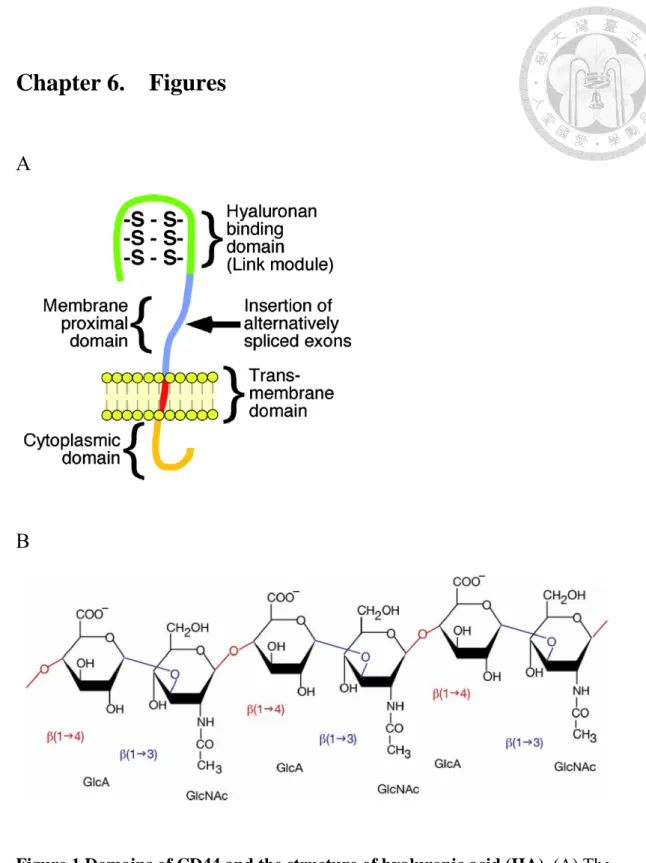
Figure 1 Domains of CD44 and the structure of hyaluronic acid (HA). (A) The
major domains of the standard isoform of human CD44 are shown. They consist of
three regions, a C-terminal cytoplasmic domain which signals molecules directly or
indirectly to binding region, a hydrophobic transmembrane domain, and a extracellular
domain, which is the so-called link modules of hyaluronan binding proteins. However,
the variant isoforms of CD44 inserts a alternatively spliced exons within the
extracellular domain, a membrane proximal domain, which can alter the binding affinity
for HA and confer interaction with alternative ligands. (B) Hyaluronic acid is composed
of repeating polymeric disaccharides D-glucuronic acid (GlcA) and
N-acetyl-D-glucosamine (GlcNAc) linked by a glucuronidic β (1→3) bond.
A
B
Figure 2 Pro-inflammatory molecule enhances the expression of CD44 on MNC
and PMN. (A) MNC added with anti-CD3 and anti-CD28, (B) PMN added with LPS and inflammatory molecule could enhance the expression of CD44.
Figure 3 HA induced IL-8 production. The detailed procedures were described in
“Materials and Mathods” LPS as positive control. HA could induce the IL-8 production
by PMN. However, anti-CD44 or anti-CD45 had little effect on IL-8 concentration.
Figure 4 HA enhanced PMN phagocytosis activity. LPS enhances the phagocytosis
and is used as positive control. HA had greater enhancement of phagocytosis compared
to control and LPS. And anti-CD44 and anti-CD45 induced increase in phagocytosis,
compared to the controls to an extent less them that by LPS.
A
B
C
D
E
F
Figure 5 HA induced cytoskeleton rearrangement on PMN. (A) medium, (B) IgG,
(C) LPS, (D) anti-CD44, (E) anti-CD45, (F) HA. LPS induce cytoskeleton
rearrangement and could enhance the actin aggregation phenomenon. In (F) HA could
clearly induce the aggregation of actin, compared to the control. (D) and (E) also show
the structure of actin aggregation in PMN. HA, anti-CD44 and anti-CD45 are presumed
to induce cytoskeleton rearrangement in PMN.
A
B
C
D
Figure 6 HA induced phosphorylation of MAPK and ERK1/2 on PMN and MNC.
(A) phosphor-ERK1/2, (B) phosphor-P38 on MNC. (C) phosphor-ERK1/2, (D)
phosphor-P38 in PMN. In (A) and (B) HA increase the phosphorylation of MAPK and
ERK1/2 in MNC. Anti-CD3 and anti-CD28 is positive control. In (C) and (D) HA
increase the phosphorylation of MAPK and ERK1/2 in PMN. LPS is positive control.
0 500 1000 1500 2000 2500
medium LPS
20 n g/ml
anti-CD44 hya
luron an 1m
g/m l
hyaluronan 2mg/ml
(pg/ml)
HL-60 (DMSO)
Figure 7 HA abolished to induced IL-8 production on differiated-HL60. The
detailed procedures were described in “Materials and Mathods” LPS as positive control.
The differentiation of HL60 cells were induced by DMSO contributes to decrease the
expression of CD44. HA could abolish to induce IL-8 production on differiated-HL60
as well as anti-CD44.
Chapter 7. References
1. Toole B. P., Slomiany M. G. Hyaluronan, CD44 and Emmprin: partners in cancer
cell chemoresistance. Drug Resist Updat. 11, 110-121. (2008)
2. Goodison S., Urquidi V., Tarin D. CD44 cell adhesion molecules. Mol Pathol. 52,
189-196. (1999)
3. Stauder R., EistererW., Thaler J. et al. CD44 variant isoforms in non-Hodgkin’s
lymphoma: a new independent prognostic factor. Blood 85, 2885–2899. (1995)
4. Salles G., Zain M., Jiang W. M. et al. Alternatively spliced CD44 transcripts in
diffuse large-cell lymphomas: characterization and comparison with normal
activated B cells and epithelial malignancies. Blood 82, 3539–3547. (1993)
5. Arch R., Wirth K., Hofmann M. et al. Participation in normal immune responses of
a metastasis-inducing splice variant of CD44. Science 257, 682–685. (1992)
6. Zohar R., Suzuki N., Suzuki K., Arora P., Glogauer M., McCulloch C. A., Sodek J.
Intracellular osteopontin is an integral component of the CD44-ERM complex
involved in cell migration. J Cell Physiol. 184, 118-130. (2000)
7. Faassen A. E., Schrager J. A., Klein D. J., Oegema T. R., Couchman J. R.,
McCarthy J. B. A cell surface chondroitin sulfate proteoglycan, immunologically
related to CD44, is involved in type I collagen-mediated melanoma cell motility and
invasion. J Cell Biol. 116, 521-531. (1992)
8. Knutson J. R., Iida J., Fields G. B., McCarthy J. B. CD44/chondroitin sulfate
proteoglycan and alpha 2 beta 1 integrin mediate human melanoma cell migration
ontype IV collagen and invasion of basement membranes. Mol Biol Cell. 7, 383-396.
(1996)
9. Jalkanen S., Jalkanen M. Lymphocyte CD44 binds the COOH-terminal
heparin-binding domain of fibronectin. J Cell Biol. 116. 817-825. (1992)
10. Paul W. N., Jiurong L., Dianhua J. Hyaluronan as an immune regulator in human
diseases. Physiol Rev. 91, 221-264. (2011)
11. Bajorath J. Molecular organization, structural features, and ligand binding
characteristics of CD44, a highly variable cellsurface glycoprotein with multiple
functions. Proteins. 39, 103-111. (2000)
12. Lesley J., Hyman R., Kincade P. W. CD44 and its interaction with extracellular
matrix. Adv Immunol. 54, 271-335. (1993)
13. Martin T. A., Harrison G., Mansel R. E., Jiang W. G. The role of the CD44/ezrin
complex in cancer metastasis. Crit Rev Oncol Hematol. 46, 165-186. (2003)
14. Ponta H., Sherman L., Herrlich P. A. CD44: from adhesion molecules to signaling
regulators. Nat Rev Mol Cell Biol. 4, 33-45. (2003)
15. Protin U., Schweighoffer T., Jochum W., Hilberg F. CD44-deficient mice develop
normally with changes in subpopulations and recirculation of lymphocyte subsets. J
Immunol. 163, 4917-4923. (1999)
16. Khan A. I., Kerfoot S. M., Heit B., Liu L., Andonegui G., Ruffell B., Johnson P.,
Kubes P. Role of CD44 and hyaluronan in neutrophil recruitment. J Immunol. 173,
7594-7601. (2004)
17. Vivers S., Dransfield I., Hart S. P. Role of macrophage CD44 in the disposal of
inflammatory cell corpses. Clin Sci. 103, 441-449. (2002)
18. Thomas M. L. The leukocyte common antigen family. Annu Rev Immunol. 7,
339–369. (1989)
19. Lodbetter J. A., Rose L. M., Spooner C. E., Beatty P. G., Martin P. J., Clark E. A.
Antibodies to leukocyte common antigen p220 influence human T cell proliferation
by modifying IL-2 receptor expression. J Immunol. 135, 1819–1825 (1985)
20. Newman W, Fast L. D., Rose L. M. Blockade of NK cell lysis is a property of
monoclonal antibodies that bind to distinct regions of T-200. J Immunol. 131,
1742–1747. (1993)
21. Bottomly K., Lugman M., Greenbaum L. et al. A monoclonal antibody murine
CD45R distinguished CD45+ T cell population that produce different cytokines. Eur
J Immunol. 19, 617–623. (1989)
22. Spertini F., Wang A. V. T., Chatila T., Geha R. S. Engagement of the common
leukocyte antigen CD45 induces homotypic adhesion of activated human T cells. J
Immunol. 153, 1593–1602. (1994)
23. Budd, R. C. et al. Distinction of virgin and memory T lymphocytes: stable
acquisition of the Pgp-1 glycoprotein concomitant with antigenic stimulation. J.
Immunol. 138, 3120–3129. (1987)
24. Foster, L. C. et al. Regulation of CD44 gene expression by the pro-inflammatory
cytokine interleukin-1β in vascular smooth muscle cells. J. Biol. Chem. 273,
20341–20346. (1998)
25. Zeidler, A. et al. Therapeutic effects of antibodies against adhesion molecules in
murine collagen type II-induced arthritis. Autoimmunity 21, 245–252. (1995)
26. Camp, R. L. et al. CD44 is necessary for optimal contact allergic responses but is
not required for normal leukocyte extravasation. J. Exp. Med. 178, 497–508. (1993)
27. Brocke, S. et al. Antibodies to CD44 and integrin α4, but not L-selectin, prevent
central nervous system inflammation and experimental encephalomyelitis by
blocking secondary leukocyte recruitment. Proc. Natl. Acad. Sci. U. S. A. 96,
6896–6901. (1999)
28. Rafi-Janajreh, A. Q. et al. Evidence for the involvement of CD44 in endothelial cell
injury and induction of vascular leak syndrome by IL-2. J. Immunol. 163,
1619–1627. (1999)
29. Hara T., Fu S. M., Hansen J. A. Human T cell activation, II: a new activation
pathway used by a major T cell population via a disulfidebonded dimer of a 44
kilodalton polypeptide (9.3 antigen). J Exp Med. 16, 11513- 11524. (1985)
30. June C. H., Bluestone J. A., Nadler L. M., Thompson C. B. The B7 and CD28
receptor families. Immunol Today. 15, 321- 333. (1994)
31. Oppenheim J. J., C. O. C. Zachariae N., Mukaida K., Matsushima. K. Properties of
the novel proinflammatory supergene "intercrine" cytokine family. Ann. Rev.
Immunol. 9, 617-648. (1991)
32. Miller M. D., Krangel M. S. Biology and biochemistry of the chemokines: a family
of chemotactic and inflammatory cytokines. Crit Rev Immunol 12, 17-46. (1992)
33. Matsushima K., Baldwin E. T., Mukaida N. Interleukin-8 and MCAF: novel
leukocyte recruitment and activating cytokines. Chem Immunol. 51, 236–265.
(1992)
34. Hack C. E., Aarden L. A., Thijs L. G. Role of cytokines in sepsis. Adv Immunol 66,
101-195. (1997)
35. Teder P., Vandivier R. W., Jiang D., Liang J., Cohn L., Puré E., Henson P. M.,
Noble P. W. Resolution of lung inflammation by CD44. Science 296, 155-158.
(2002)
36. Moffat F. L. Jr., Han T., Li Z. M., Peck M. D., Falk R. E., Spalding P. B., Jy
W., Ahn Y. S., Chu A. J., Bourguignon L. Y. Involvement of CD44 and the
cytoskeletal linker protein ankyrin in human neutrophil bacterial phagocytosis. J
Cell Physiol. 168, 638-647. (1996)
37. Hart S. P., Dougherty G. J., Haslett C., Dransfield I. CD44 regulates phagocytosis of
apoptotic neutrophil granulocytes, but not apoptotic lymphocytes, by human
macrophages. J Immunol. 159, 919-925. (1997)
38. Vachon E., Martin R., Plumb J., Kwok V., Vandivier R. W., Glogauer M., Kapus
A., Wang X., Chow C. W., Grinstein S., Downey G. P. CD44 is a phagocytic
receptor. Blood 107, 4149-4158. (2006)
39. Yu C. L., Yu H. S., Sun K. H., Hsieh S. C., Tsai C. Y. Anti-CD45 isoform
antibodies enhance phagocytosis and gene expression of IL-8 and TNF-alpha in
human neutrophils by differential suppression on protein tyrosine phosphorylation
and p56lck tyrosine kinase. Clin Exp Immunol. 129, 78-85. (2002)
40. Lokeshwar V. B., Fregien N., Bourguignon L. Y. Ankyrin-binding domain of CD44
(GP85) is required for the expression of hyaluronic acid-mediated adhesionfunction.
J Cell Biol. 126, 1099-1109. (1994)
41. Bennett V., Baines A. J. Spectrin and ankyrin-based pathways: metazoan inventions
for integrating cells into tissues. Physiol Rev. 81, 1353-1392. (2001)
42. Lee J. Y., Spicer A. P. Hyaluronan: a multifunctional, megaDalton, stealth molecule.
Curr Opin Cell Biol. 12, 581-586. (2000)
43. Ghosh P., Guidolin D. Potential mechanism of action of intra-articular hyaluronan
therapy in osteoarthritis: are the effects molecular weight dependent? Semin
Arthritis Rheum. 32, 10-37. (2002)
44. Liu J., Bi G., Wen P., Yang W., Ren X., Tang T., Xie C., Dong W., Jiang G.
Down-regulation of CD44 contributes to the differentiation of HL-60 cells induced
by ATRA or HMBA. Cell Mol Immunol. 4, 59-63. (2007)
45. Nedvetzki, S., Walmsley M., Alpert E. et al. CD44 involvement in experimental
collagen-induced arthritis (CIA). J. Autoimmun. 13, 39–47. (1999)
46. Nedvetzki, S., Gonen E., Assayag N. et al. RHAMM, a receptor for
hyaluronan-mediated motility, compensates for CD44 in inflamed CD44-knockout
mice: a different interpretation of redundancy. Proc. Natl. Acad. Sci. 101,
18081–18086. (2004)
47. Mikecz, K., Brennan F. R., Kim J. H. et al. Anti-CD44 treatment abrogates tissue
oedema and leukocyte infiltration in murine arthritis. Nat. Med. 1, 558–563. (1995)
48. Verdrengh, M., Holmdahl R., Tarkowski A. Administration of antibodies to
hyaluronan receptor (CD44) delays the start and ameliorates the severity of collagen
II arthritis. Scand. J. Immunol. 42, 353–358. (1995)
49. Zeidler A., Brauer R., Thoss K. et al. Therapeutic effects of antibodies against
adhesion molecules in murine collagen type II-induced arthritis. Autoimmunity 21,
245–252. (1995)
50. Naor, D., Nedvetzki S. CD44 in rheumatoid arthritis. Arthritis Res. Ther. 5,
105–115. (2003)
51. Zöller M. CD44: can a cancer-initiating cell profit from an abundantly expressed
molecule? Nat Rev Cancer. 11, 254-267. (2011)
52. Lesley J., Hascall V. C., Tammi M., Hyman R. Hyaluronan binding by cell surface
CD44. J Biol Chem. 275, 26967-26975. (2000)